Abstract
The invasin gimB (genetic island associated with human newborn meningitis) is usually found in ExPEC (Extraintestinal Pathogenic Escherichia coli) such as UPEC (uropathogenic E. coli), NMEC (neonatal meningitis E. coli) and APEC (avian pathogenic E. coli). In NMEC, gimB is associated with the invasion process of the host cells. Due to the importance of E. coli as a zoonotic agent and the scarce information about the frequency of gimB-carrying strains in different animal species, the aim of this study was to investigate the presence of gimB in isolates from bovine, swine, canine and feline clinical samples. PCR was conducted on 196 isolates and the identity of the amplicons was confirmed by sequencing. Of the samples tested, only E. coli SB278/94 from a bovine specimen was positive (1/47) for gimB, which represents 2.1% of the bovine isolates. The ability of SB278/94 to adhere to and invade eukaryotic cells was confirmed by adherence and gentamicin-protection assays using HeLa cells. This is the first study that investigates for gimB in bovine, canine and feline E. coli isolates and shows E. coli from the intestinal-bovine samples harboring gimB.
gimB ; adherence; invasiveness; zoonotic potential; livestock; companion animals
Introduction
Escherichia coli is a facultative anaerobic bacterium commonly found in the intestinal microbiota of most animal species (Gyles and Fairbrother, 2010Gyles CL, Fairbrother JM (2010) Escherichia coli. In: Gyles CL. Pathogenesis of bacterial infections in animal. 4th ed. Blackwell Publishing, Ames, pp 267–308.). Although typically commensal, various E. coli strains cause intestinal and extraintestinal diseases due to the presence of a range of virulence factors (Kaper et al., 2004Kaper JB, Nataro JP, Mobley HL (2004) Pathogenic E. coli. Nat Rev Microbiol 2:123–140.). The invasin gimB(genetic island associated with newborn meningitis) consists of a sequence of approximately 5,200 bp with six ORFs (Open Reading Frame). It was firstly found by subtractive hybridization in NMEC (neonatal meningitis E. coli) (Bonarcosi et al., 2003Bonacorsi S, Clermont O, Houdouin V et al. (2003) Molecular analysis and experimental virulence of French and North American Escherichia coli neonatal meningitis isolates: identification of a new virulent clone. J Infect Dis 187:1895–1906.). In NMEC, approximately 60% of the strains harbor the gimB sequence, which has been associated with the high levels of bacteremia and ability to the bacteria to invade endothelial cells (Bonacorsi et al., 2003Bonacorsi S, Clermont O, Houdouin V et al. (2003) Molecular analysis and experimental virulence of French and North American Escherichia coli neonatal meningitis isolates: identification of a new virulent clone. J Infect Dis 187:1895–1906.; Ewers et al., 2007Ewers C, Li G, Wilking H et al. (2007) Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related they are? Int J Med Microbiol 297:163–176.). The presence of gimB has also been reported in other ExPEC (extraintestinal pathogenic E. coli) strains, with frequencies of 9% in UPEC (uropathogenic E. coli) and 24% in APEC (avian pathogenic E. coli) (Ewers et al., 2007Ewers C, Li G, Wilking H et al. (2007) Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related they are? Int J Med Microbiol 297:163–176., Barbieri et al., 2013Barbieri NL, de Oliveira AL, Tejkowski TM et al.(2013) Genotypes and pathogenicity of cellulitis isolates reveal traits that modulate APEC virulence. PLoS One 19:e72322.).
Recently, gimB-carrying E. coli strains were isolated from pigs that displayed symptoms of diarrhea as well as asymptomatic pigs. gimB appeared in approximately 3% of both groups. While a 3% frequency is relatively low, this study showed that the gimBvirulence factor may be more frequent and specific in ExPEC strains (Schierack et al., 2011Schierack P, Weinreich J, Ewers C et al. (2011) Hemolytic porcine intestinal Escherichia coli without virulence-associated genes typical of intestinal pathogenic E. coli. Appl Environ Microbiol 77:8451–8455.).
Due to the importance of this bacterium as a zoonotic agent and the scarce studies regarding the frequency of gimB-carrying E. coliin different animal species, the aim of this work was to investigate the presence of gimB in E. coli strains isolated from a variety of animal species.
Material and Methods
E. coli isolates and PCR
In order to detect the presence and origin of gimB in E. coli from different animal species, PCR was performed on DNA isolated from clinical samples of swine, cattle, dogs and cats stored in the LABAC’s collection, UFSM/RS (Table 1). These samples were taken between 1990 and 2012 and the isolates were preserved by lyophilization. The lyophilized samples were plated on 5% sheep-blood agar (Himedia, Mumbai, Índia) and MacConkey agar (Himedia, Mumbai, Índia). Colonies were confirmed as E. coli by Gram staining and biochemical characterization (Quinn et al., 1994Quinn PJ, Carter ME, Markey B et al. (1994) Clinical veterinary microbiology. Wolfe, London.). Subsequently, DNA was extracted from the confirmed colonies (Cheng and Jiang, 2006Cheng Hr, Jiang N (2006) Extremely rapid extraction of DNA from bacteria and Yeasts. Biotechnol Lett 28:55–59.) and was used as a template for PCR assay with primers 6F and 6R (6F: 5′-GCGGGTGCCGATTATATTTC-3′ and 6R: 5′-CTTCGCGCTGCTATTGAA-3′) according to the conditions described by Matter et al. (2011)Matter LB, Barbieri NL, Nordhoff M et al. (2011) Avian Pathogenic Escherichia coli MT78 invades chicken fibroblasts. Vet Microbiol 148:51–59.. The 6F and 6R primers were designed with the gimB sequence available (access number AY170898.1) using the Primer3Plus program. PCR reaction resulted in an amplicon of 724 bp. In order to verify the DNA quality for PCR, species-specific PCR for the detection of E. coli was performed as well using the primer pair ECA75F (5′-GGAAGAAGCTTGCTT CTTTGCTGAC-3′) and ECR619R (5′-AGCCCGGGGAT TTCACATCTGACTTA-3′) (SABAT et al., 2000). The MT 78 strain was used as a positive control in all assays (Matter et al., 2011Matter LB, Barbieri NL, Nordhoff M et al. (2011) Avian Pathogenic Escherichia coli MT78 invades chicken fibroblasts. Vet Microbiol 148:51–59.).
Adhesion and invasion assays
Adhesion and invasion assays were performed according to protocols described by Matter et al., 2011Matter LB, Barbieri NL, Nordhoff M et al. (2011) Avian Pathogenic Escherichia coli MT78 invades chicken fibroblasts. Vet Microbiol 148:51–59.. With the aim of analyzing the adherence (association) profile of the gimB-positive SB278/94 strain, a confluent monolayer of HeLa cells was infected with the bacteria at a multiplicity of infection (MOI) of ~100 cfu per cell in high glucose Dulbecco’s modified Eagles medium (DMEM) (Gibco, Grand Island, NY) plus fetal calf serum (Gibco, Grand Island, NY, USA). After 2 h of incubation at 37 °C under 5% CO2 (Thermo Fisher Scientific, Asheville, NC, USA), the medium was removed, and the cells were washed three times with Phosphate Buffered Saline (PBS) and lysed with 1% (v/v) Triton X-100 (Sigma, Steinheim, Germany) at room temperature. Serial dilutions of the lysate in PBS were plated on Luria Bertani agar (Himedia, Mumbai, Índia) for cfu determination. The experiment was performed at least three separate times with quadruplicates samples of each strain.
For the invasion assay (gentamicin protection assay), HeLa cells were infected with bacteria in the same way as described for the adhesion assay and then washed three times with PBS after 2 h of incubation to allow interaction. HeLa cells and bacteria were again incubated with culture medium plus gentamicin (50 μg.mL−1) for an additional hour. In order to quantify the number of viable, internalized bacteria, the cells were washed twice with PBS and treated with 1% Triton X-100. Serial dilutions were then plated on LB agar. E. coli DH5α and MT78 were used as negative and positive controls for the invasive profile assay, respectively (Matter et al., 2011Matter LB, Barbieri NL, Nordhoff M et al. (2011) Avian Pathogenic Escherichia coli MT78 invades chicken fibroblasts. Vet Microbiol 148:51–59.).
Statistical analysis
Student’s t-test was carried out for multiple comparisons (GraphPad Prism Package 5) of adhesion- and invasion-assay results. P < 0.05 was considered statistically significant.
Results and Discussion
Two strains (SB31/94 and SB278/94) out of 196 total E. coli isolates were PCR-positive for gimB. The amplicons from these strains were sequenced to confirm that the sequences corresponded to gimB(Laboratório de Análise Molecular ACTGene LTDA, Porto Alegre, RS). Only the amplicon from the SB278/94 strain was homologous to gimB (Genbank: AJ810519.1). Thus, 0.5% of the 196 isolates, were gimB-positive, which represents 2.1% of the bovine isolates.
The presence of gimB is relatively well documented in avian and human species (Ewers et al., 2007Ewers C, Li G, Wilking H et al. (2007) Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related they are? Int J Med Microbiol 297:163–176.; Ewers et al., 2009Ewers C, Antão EM, Diehl I et al. (2009) Intestine and environment of the chicken as a reservoirs for extraintestinal pathogenic Escherichia coli strains with zoonotic potential. Appl Environ Microbiol 74:184–192.; Matter et al., 2011Matter LB, Barbieri NL, Nordhoff M et al. (2011) Avian Pathogenic Escherichia coli MT78 invades chicken fibroblasts. Vet Microbiol 148:51–59.; Barbieri et al., 2013Barbieri NL, de Oliveira AL, Tejkowski TM et al.(2013) Genotypes and pathogenicity of cellulitis isolates reveal traits that modulate APEC virulence. PLoS One 19:e72322.); however, few studies have determined the frequency of gimB in other animal species. Schierack et al. (2008Schierack P, Walk N, Ewers C et al. (2008) ExPEC-typical virulence-associated genes correlate with successful colonization by intestinal E. coli in a small piglet group. Environ Microbiol 10:1742–1751.; 2009Schierack P, Kadlec K, Guenther S et al. (2009) Antimicrobial resistances do not affect colonization parameters of intestinal E. coli in a small piglet group. Gut Pathog 1:18.; 2011Schierack P, Weinreich J, Ewers C et al. (2011) Hemolytic porcine intestinal Escherichia coli without virulence-associated genes typical of intestinal pathogenic E. coli. Appl Environ Microbiol 77:8451–8455.; 2013)Schierack P, Rödiger S, Kuhl C et al. (2013) Porcine E. coli: virulence-associated genes, resistance genes and adhesion and probiotic activity tested by a new screening method. PLoS One 26:e59242. found 3.2% (2/62) of the diarrhea isolates and 2.7% (1/37) of E. coli from healthy animals carrying gimB in a study of ExPEC-genes in hemolytic E. coli from swine. None of the non-hemolytic E. colifrom healthy pigs contained gimB (Schierack et al., 2011Schierack P, Weinreich J, Ewers C et al. (2011) Hemolytic porcine intestinal Escherichia coli without virulence-associated genes typical of intestinal pathogenic E. coli. Appl Environ Microbiol 77:8451–8455.). In our study none of the 136 swine isolates carried gimB.
We also did not find gimB in the isolates from canine and feline samples, despite the relatively common presence of this genetic island in extraintestinal-infection isolates (Ewers et al., 2007Ewers C, Li G, Wilking H et al. (2007) Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related they are? Int J Med Microbiol 297:163–176., Barbieri et al., 2013Barbieri NL, de Oliveira AL, Tejkowski TM et al.(2013) Genotypes and pathogenicity of cellulitis isolates reveal traits that modulate APEC virulence. PLoS One 19:e72322.). This result is likely due to the small sample numbers of these groups.
The SB278/94 strain was isolated in 1994 from the intestinal lumen of a calf with diarrhea that died from peritonitis. There are no data about bovine E. coli harboring gimB in the literature. The only data regarding intestinal gimB-containing E. coli are those describing isolates from swine (Schierack et al., 2011Schierack P, Weinreich J, Ewers C et al. (2011) Hemolytic porcine intestinal Escherichia coli without virulence-associated genes typical of intestinal pathogenic E. coli. Appl Environ Microbiol 77:8451–8455.) and human samples (GenBank accession number: CP002167.1). The human intestinal E. coli is an adherent and invasive E. coli (AIEC) pathotype that is associated with Crohn’s disease, a form of inflammatory bowel disease (IBD). AIEC can adhere to and invade enterocytes and can replicate inside macrophages (Krause et al., 2011Krause DO, Little AC, Dowd SE et al. (2011) Complete genome sequence of adherent invasive Escherichia coli UM146 isolated from Ileal Crohn’s disease biopsy tissue. J Bacteriol 193:583.). According to recent studies, AIEC strains share many genetic and phenotypic features with ExPEC strains (Moulin-Schouleur et al., 2006Moulin-Schouler M, Schouler C, Tailliez P et al.(2006) Common virulence factors and genetic relatioships between O18:K1:H7 Escherichia coli isolates of human and avian origin. J Clin Microbiol 44:3484–3492.; Martinez-Medina et al., 2009Martinez-Medina M, Mora A, Blanco M et al. (2009) Similarity and divergence among adherent-invasive Escherichia coli and extraintestinal pathogenic E. colistrains. J Clin Microbiol 47:3968–3970.; Krause et al., 2011Krause DO, Little AC, Dowd SE et al. (2011) Complete genome sequence of adherent invasive Escherichia coli UM146 isolated from Ileal Crohn’s disease biopsy tissue. J Bacteriol 193:583.). The prevalence and importance of this island for this E. coli pathotype is still unknown.
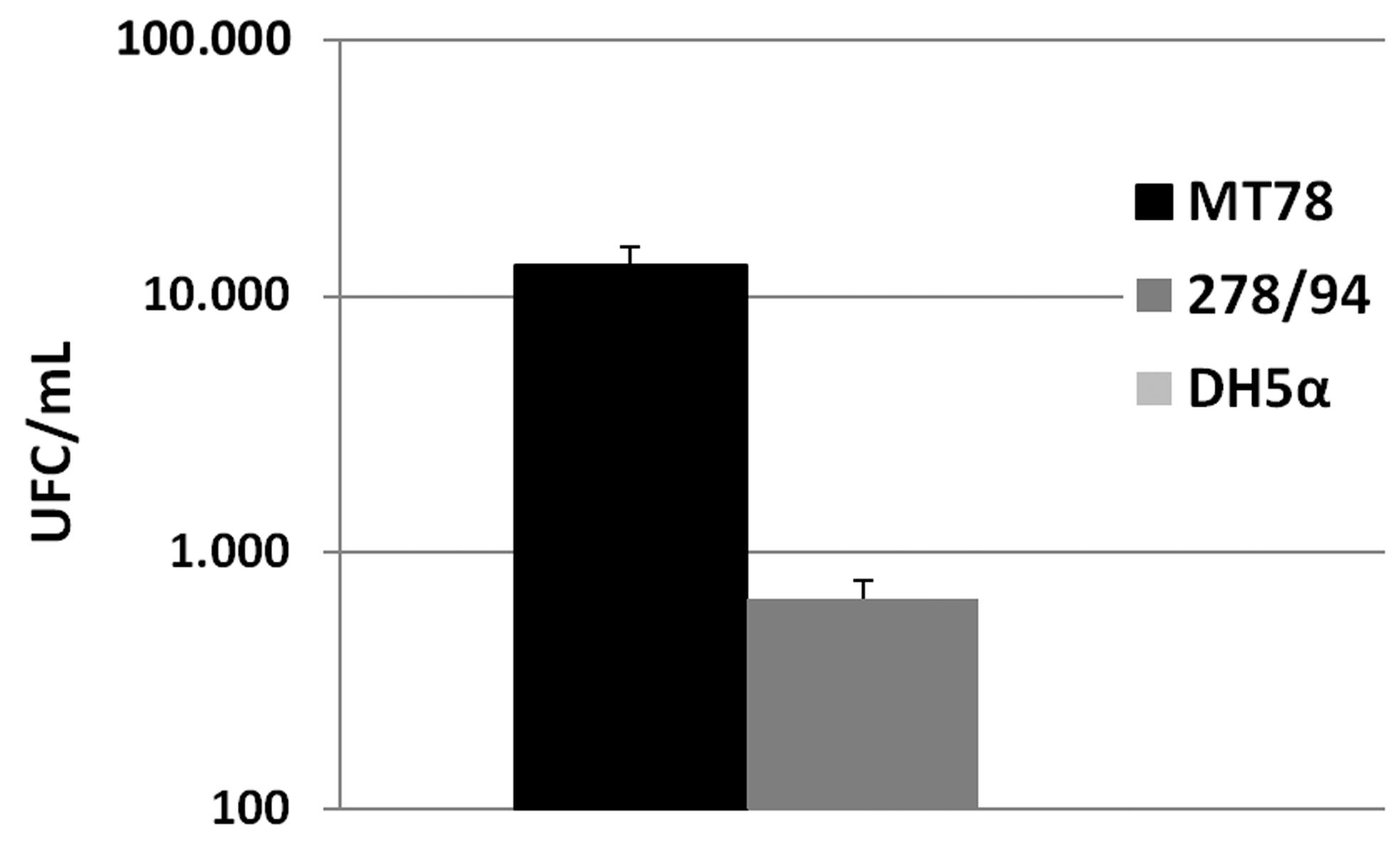
The SB278/94 strain is also capable of adhering to and invading eukaryotic cells such as HeLa cells, at levels comparable to the positive control MT78 strain (Figures 1 and 2). Although our data suggest that this isolate is an AIEC, genetic characterization and in vivo studies with macrophages and enterocytes are still required. In vitro and in vivo studies will also be necessary to investigate the mechanism by which gimB contributes to adherence and invasion in this strain.
Capacity of E. coli strains to adhere to HeLa cells. Data represent the average and standard deviation of at least three assays done in quadruplicates for each strain. MT78 and DH5α strains represent the strain with high and low adherence level. Statistical analysis has showed significant difference among the three strains (p < 0.05)
Capacity of E. coli strains to invade HeLa cells. Results are shown as UFC/mL. Data represent the average and standard deviation of at least three assays done in quadruplicates for each strain. MT78 and DH5α strains represent the positive and negative controls for invasiveness. Statistical analysis showed significant difference between MT78 and 278/94 (p < 0.05). None bacterium was recovered from inside HeLa cells after gentamicin protection assay with DH5α strain
In summary, this study has revealed that E. coli from clinical bovine sources can also harbor gimB. Future studies should be performed to determine the actual clinical impact of this finding and the role of gimB in the pathogenesis of intestinal pathotypes.
Acknowledgments
The authors would like to thank to the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the grant PNPD Institucional 2011 nº 23038007126201144.
References
- Barbieri NL, de Oliveira AL, Tejkowski TM et al.(2013) Genotypes and pathogenicity of cellulitis isolates reveal traits that modulate APEC virulence. PLoS One 19:e72322.
- Bonacorsi S, Clermont O, Houdouin V et al. (2003) Molecular analysis and experimental virulence of French and North American Escherichia coli neonatal meningitis isolates: identification of a new virulent clone. J Infect Dis 187:1895–1906.
- Cheng Hr, Jiang N (2006) Extremely rapid extraction of DNA from bacteria and Yeasts. Biotechnol Lett 28:55–59.
- Ewers C, Li G, Wilking H et al. (2007) Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related they are? Int J Med Microbiol 297:163–176.
- Ewers C, Antão EM, Diehl I et al. (2009) Intestine and environment of the chicken as a reservoirs for extraintestinal pathogenic Escherichia coli strains with zoonotic potential. Appl Environ Microbiol 74:184–192.
- Gyles CL, Fairbrother JM (2010) Escherichia coli In: Gyles CL. Pathogenesis of bacterial infections in animal. 4th ed. Blackwell Publishing, Ames, pp 267–308.
- Kaper JB, Nataro JP, Mobley HL (2004) Pathogenic E. coli Nat Rev Microbiol 2:123–140.
- Krause DO, Little AC, Dowd SE et al. (2011) Complete genome sequence of adherent invasive Escherichia coli UM146 isolated from Ileal Crohn’s disease biopsy tissue. J Bacteriol 193:583.
- Martinez-Medina M, Mora A, Blanco M et al. (2009) Similarity and divergence among adherent-invasive Escherichia coli and extraintestinal pathogenic E. colistrains. J Clin Microbiol 47:3968–3970.
- Matter LB, Barbieri NL, Nordhoff M et al. (2011) Avian Pathogenic Escherichia coli MT78 invades chicken fibroblasts. Vet Microbiol 148:51–59.
- Moulin-Schouler M, Schouler C, Tailliez P et al.(2006) Common virulence factors and genetic relatioships between O18:K1:H7 Escherichia coli isolates of human and avian origin. J Clin Microbiol 44:3484–3492.
- Quinn PJ, Carter ME, Markey B et al. (1994) Clinical veterinary microbiology. Wolfe, London.
- Schierack P, Walk N, Ewers C et al. (2008) ExPEC-typical virulence-associated genes correlate with successful colonization by intestinal E. coli in a small piglet group. Environ Microbiol 10:1742–1751.
- Schierack P, Kadlec K, Guenther S et al. (2009) Antimicrobial resistances do not affect colonization parameters of intestinal E. coli in a small piglet group. Gut Pathog 1:18.
- Schierack P, Weinreich J, Ewers C et al. (2011) Hemolytic porcine intestinal Escherichia coli without virulence-associated genes typical of intestinal pathogenic E. coli Appl Environ Microbiol 77:8451–8455.
- Schierack P, Rödiger S, Kuhl C et al. (2013) Porcine E. coli: virulence-associated genes, resistance genes and adhesion and probiotic activity tested by a new screening method. PLoS One 26:e59242.
Publication Dates
-
Publication in this collection
Jul-Sep 2015
History
-
Received
22 July 2014 -
Accepted
19 Dec 2014