Abstract
Here we sequenced and characterized the complete plastome of Bactris riparia, a species closely related to B. gasipaes and widely distributed in Western Amazonia. We performed a comparative genomic analysis with B. riparia and the other four Bactridinae species retrieved from GenBank. The plastome of B. riparia was 156,715 bp with a quadripartite structure. Gene content included 86 protein-coding genes (CDS), 38 tRNAs, and 8 rRNAs. Bactris riparia has 69 more base pairs than B. gasipaes, with identical numbers in IR, and more in LSC and SSC. The comparative analysis indicated that structure, collinearity, and IR/SSC borders of plastomes within subtribe Bactridinae are, in general, conserved. We predicted 69 SSRs in B. riparia plastome. Among them, ~80% consisted of A/T homopolymers. Among the 52 variable CDS, rbcL showed the highest non-synonymous rate, while the rps15 gene had the highest synonymous rate. Three genes (ccsA, cemA, and rpoC1) presented evidence of positive selection and 22 genes showed evidence of purifying selection. The phylogenetic tree based on plastome sequences set Bactris as more closely related to Astrocaryum than to Acrocomia. These new plastome data of B. riparia will contribute to studies about the diversity, evolutionary history, and conservation of palms.
Keywords
Palms; Amazonian floodplains; plastome; molecular evolution
Palms (family Arecaceae), icons of tropical landscapes, form an economically and ecologically important group of ~2,500 species with remarkable diversity and abundance in the Neotropics (Dransfield et al., 2008Dransfield J, Uhl NW, Lange CBA, Baker WJ, Harley MM and Lewis CE (2008) Genera Palmarum: The Evolution and Classification of Palms. Kew Publishing, London, 732 p.). These species provide valuable ecosystem services, such as provision (food, medicines, and raw materials), support and regulation (contributing to the maintenance of biodiversity), and cultural (leisure and spirituality of various human groups) (Cámara-Leret et al., 2014Cámara-Leret R, Paniagua-Zambrana N, Balslev H, Barfod A, Copete JC and Macía MJ (2014) Ecological community traits and traditional knowledge shape palm ecosystem services in northwestern South America. For Ecol Manage 334:28-42.). Nonetheless, their phylogenetic diversity has not been safeguarded under current and possible future climate conditions by the existing network of Protected Areas (Velazco et al., 2020Velazco, SJE, Svenning, JC, Ribeiro, BR and Laureto, LMO (2020) On opportunities and threats to conserve the phylogenetic diversity of Neotropical palms. Divers Distrib 27:512-523.).
Given such relevance, palms have become a model group for studies on tropical forests (Faurby et al., 2016Faurby S, Eiserhardt WL, Baker WJ and Svenning JC (2016) An all-evidence species-level supertree for the palms (Arecaceae). Mol Phylogenet Evol 100:57-69.). However, as most of these efforts focused on the relationship of major clades of Arecaceae, the phylogenetic relationships at lower taxonomic levels require refinement (Baker and Dransfield, 2016Baker WJ and Dransfield J (2016) Beyond Genera Palmarum: Progress and prospects in palm systematics. Bot J Linn Soc 182:207-233.). For example, Bactridinae (Cocoseae, Arecaceae) consists of five genera (Acrocomia Mart., Aiphanes Willd., Astrocaryum G. Mey., Bactris Jacq. ex Scop., and Desmoncus Mart.), about 150 species, and represents approximately 20 % of the Neotropical palm flora. The subtribe is well-characterized by the presence of epidermal spines and by adherent fibers and deep pores in the endocarp (Dransfield et al., 2008Dransfield J, Uhl NW, Lange CBA, Baker WJ, Harley MM and Lewis CE (2008) Genera Palmarum: The Evolution and Classification of Palms. Kew Publishing, London, 732 p.), but its intergeneric evolutionary relationships require more study (Baker and Dransfield, 2016Baker WJ and Dransfield J (2016) Beyond Genera Palmarum: Progress and prospects in palm systematics. Bot J Linn Soc 182:207-233.).
Previous studies sought to elucidate the phylogenetic relationships within the subtribe using multiple nuclear and plastid markers (Eiserhardt et al., 2011Eiserhardt WL, Pintaud JC, Asmussen-Lange C, Hahn WJ, Bernal R, Balslev H and Borchsenius F (2011) Phylogeny and divergence times of Bactridinae (Arecaceae, Palmae) based on plastid and nuclear DNA sequences. Taxon60:485-498.). However, they did not yield well-defined and supported relationships (Baker and Dransfield, 2016Baker WJ and Dransfield J (2016) Beyond Genera Palmarum: Progress and prospects in palm systematics. Bot J Linn Soc 182:207-233.). Due to structural and evolutionary particularities, the plastome has become a valuable tool for taxonomic, phylogenetic, biogeographic, population genetics, and biotechnological studies (Rogalski et al., 2015Rogalski, M, do Nascimento, Vieira, L, Fraga, HP and Guerra, MP (2015) Plastid genomics in horticultural species: importance and applications for plant population genetics, evolution, and biotechnology. Front Plant Sci6:586.). We sequenced and characterized the plastome of the Amazonian floodplain palm Bactris riparia Mart., a close relative of the economically important Bactris gasipaes Kunth (Couvreur et al., 2007Couvreur TL, Hahn WJ, Granville JJD, Pham JL, Ludena B and Pintaud JC (2007) Phylogenetic relationships of the cultivated Neotropical palm Bactris gasipaes (Arecaceae) with its wild relatives inferred from chloroplast and nuclear DNA polymorphisms. Syst Bot 32:519-530.; Clement et al., 2017Clement CR, Cristo-Araújo M, Coppens d’Eeckenbrugge G, Reis VM, Lehnebach R and Picanço-Rodrigues D (2017) Origin and dispersal of domesticated peach palm. Front Ecol Evol 5:148.), and performed comparative analyzes in the subtribe Bactridinae. For the taxonomic review of the genus Bactris see Henderson (2000Henderson A (2000) Bactris (Palmae). Organization for Flora Neotropica, 181p.).
For this, fresh leaf tissue of Bactris riparia was collected from a single individual (SisGen register number AB0ED60) close to the Solimões River in Manacapuru (AM, Brazil). A sample was deposited in the INPA herbarium, under accession number 290838. The genomic DNA was extracted using DNeasy Plant Mini Kit (Qiagen, Valencia, CA, USA). The DNA was quantified using Qubit fluorometer (Thermo Fisher Scientific) and DNA quality was verified by spectrometry using Nanodrop 2000 (Thermo Fisher Scientific) and by agarose gel electrophoresis (0.8 %, GelRed-stainedTM). Libraries were prepared with genomic DNA (100 ng) using Illumina DNA Prep kit (Illumina, San Diego, CA, USA), and sequenced on an Illumina HiSeq 2500 platform (Illumina Inc.) using a V4 chemistry at NGS Soluções Genômicas (Piracicaba, Brazil). The resulting paired-end reads (2 x 100 bp) were de novo assembled using NOVOPlasty (Dierckxsens et al., 2016Dierckxsens N, Mardulyn P and Smits G (2016) NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res 45:e18.). The annotation of the plastome gene content was performed using Geneious Prime® 2021.1.1 (Biomatters Ltd., San Diego, CA, USA) and manually verified using Bactris gasipaes (MW054718) as reference. The plastome sequence was submitted to GenBank (MZ823390). The comparative analyzes involved Bactris riparia and four species of Bactridinae available in GenBank: Acrocomia aculeata (Jacq.) Lodd. ex Mart. (NC_037084.1), Astrocaryum aculeatum G. Mey (NC_044482.1), Astrocaryum murumuru Mart. (NC_044481.1), and Bactris gasipaes Kunth (MW054718). Possible structural rearrangements were verified using the progressive algorithm of Mauve 2.4.0 (Darling et al., 2004Darling ACE, Mau B, Blattner FR and Perna NT (2004) Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res 14:1394-1403.). The four plastome junctions (IRb/LSC, IRb/SSC, SSC/IRa, IRa/LSC) were analyzed and illustrated by IRScope (Amiryousefi et al., 2018Amiryousefi A, Hyvönen J and Poczai P (2018) IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 34:3030-3031.). Simple sequence repeats (SSRs) were detected using MISA-web (Beier et al., 2017Beier S, Thiel T, Münch T, Scholz U and Mascher M (2017) MISA-web: A web server for microsatellite prediction. Bioinformatics 33:2583-2585.) with thresholds of ten repeat units for mononucleotide SSRs, six repeat units for di- and trinucleotide SSRs, and five repeat units for tetra-, penta-, and hexanucleotide SSRs. The maximum length of sequence between two SSRs to register as compound SSR was set to 100. The 79 protein-coding genes of Bactridinae species (Bactris riparia, Acrocomia aculeata, Astrocaryum aculeatum, Astrocaryum murumuru, and Bactris gasipaes) were extracted using Geneious Prime and individually align with the aid of MAUVE implemented on Geneious Prime. Among them, 27 genes were 100% identity and were not used for selective pressure analysis. The numbers of non-synonymous and synonymous substitutions rate were estimated using DNASP v6.12.03. Then, the Ka/Ks ratio was estimated. For the phylogenetic analysis, we used the plastomes of the five species of Bactridinae and three species as outgroup (Cocos nucifera L. KF285453, Elaeis guineensis Jacq. NC_017602, and Syagrus coronata (Mart.) Becc. NC_029241) to obtain a multiple alignment (one IR excluded) on Mauve 2.0. The locally collinear blocks (LCBs) from the Mauve alignment were extracted and concatenated into a single alignment using Geneious Prime®. Subsequently, the resulting alignment was submitted to Model Selection implemented in W-IQ-TREE (Trifinopoulos et al., 2016Trifinopoulos J, Nguyen L, Haeseler A and Minh BQ (2016) W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res 44:W232-W235.), and the algorithm suggested the best evolutionary model for our data set was HKY+F+I model, according to the Bayesian Information Criterion (BIC). Then, the phylogenetic dendrogram was generated with W-IQ-TREE (Trifinopoulos et al., 2016Trifinopoulos J, Nguyen L, Haeseler A and Minh BQ (2016) W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res 44:W232-W235.), under the Maximum Likelihood (ML) probabilistic model and 1000 bootstrap replications.
A total of 55 million reads were generated by the Illumina Hiseq 2500 platform, of which 378,906 reads were used in the assembly of the Bactris riparia plastome with an average depth of 268×. The assembled plastome was 156,715 bp in length and presented the typical quadripartite structure of angiosperms (LSC - 85,153 bp; IRs - 27,038 bp; SSC - 17,486 bp). The GC content was 37.5 %, consistent with what was expected for Bactridinae plastomes (Table 1). The gene content is conserved among Bactridinae, with a total of 132 genes, including 86 protein-coding genes, 38 tRNAs and 8 rRNAs (Table 1). The ndhD and rpl2 genes had the unusual ACG start codon, which was shown to be corrected by the post-transcriptional process of mRNA editing involving the pentatricopeptide repeat (PPR) proteins (Boussardon et al., 2012Boussardon C, Salone V, Avon A, Berthomé R, Hammani K, Okuda K, Shikanai T, Small I and Lurin C (2012) Two interacting proteins are necessary for the editing of the ndhD-1 site in Arabidopsis plastids. Plant Cell 24:3684-3694.). The cemA gene exhibited the same alternative start codon reported for B. gasipaes, distinct to the other species of Bactridinae sequenced so far (Astrocaryum aculeatum, A. murumuru, and Acrocomia aculeata) (Lopes et al., 2019Lopes, AS, Pacheco, TG, Silva, ON, Cruz, LM, Balsanelli, E, Souza, EM, Pedrosa, FO and Rogalski, M (2019) The plastomes of Astrocaryum aculeatum G. Mey. and A. murumuru Mart. show a flip-flop recombination between two short inverted repeats. Planta250:1229-1246. ; Santos da Silva et al., 2021Santos da Silva, R, Clement, CR, Balsanelli, E, Baura, VA, Souza, EM, Fraga, HPF and do Nascimento Vieira, L (2021) The plastome sequence of Bactris gasipaes and evolutionary analysis in tribe Cocoseae (Arecaceae). PLoS One 16:e0256373. ).
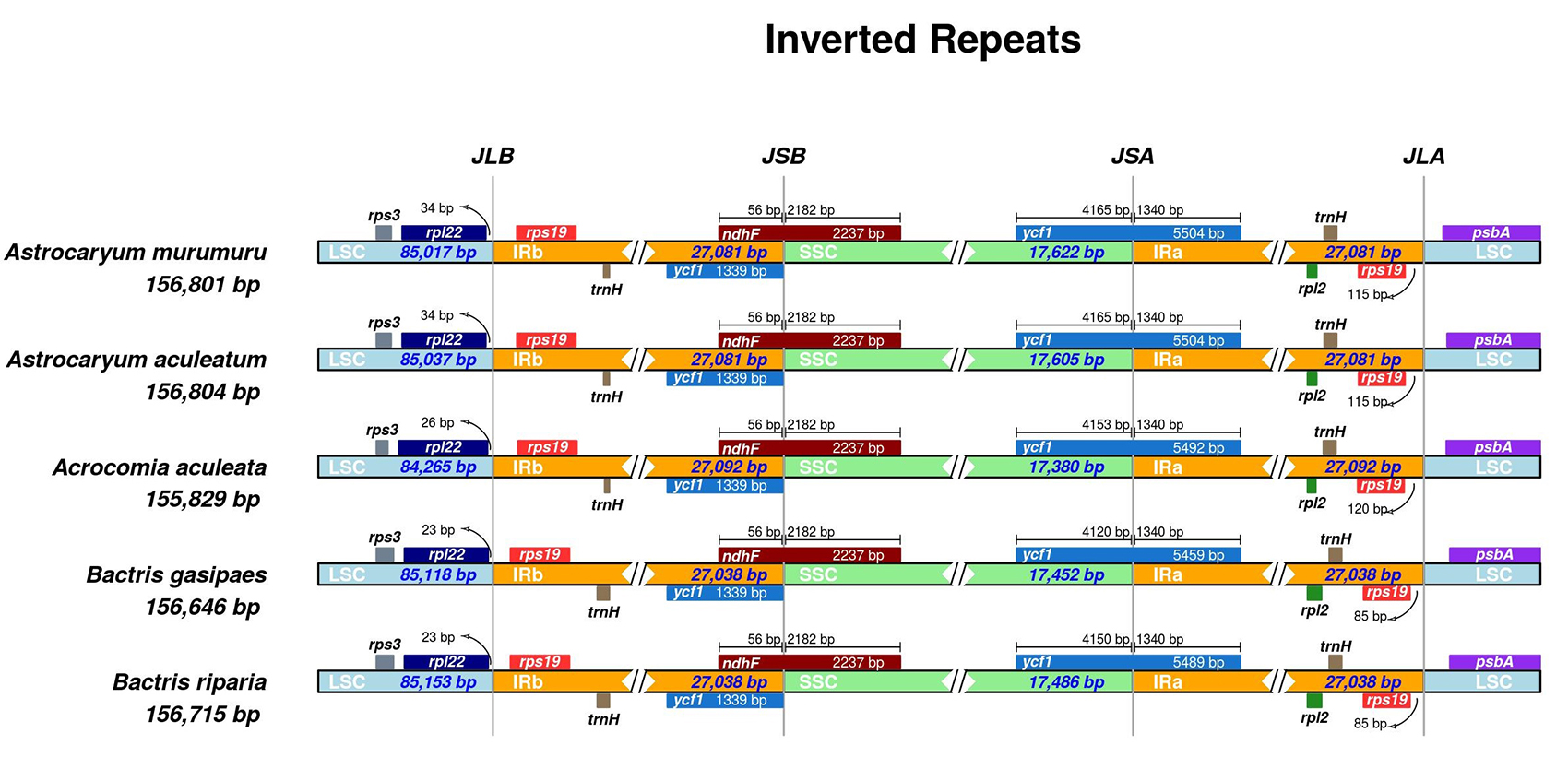
The comparative analysis of the IRs junctions within species of Bactridinae revealed a highly conserved region. The length of the IRs ranged from 27,038 to 27,092 bp. Bactris gasipaes and Bactris riparia have IRs with the same length (27,038 bp) (Table 1). All Bactridinae plastomes exhibited the trnH-rps19 gene cluster and showed the incorporation of part of the ndhF gene into IRb. An expansion of IRa over SSC was observed in the ycf1 gene, which appears as a pseudogene in IRb and with an overlap of 56 bp with ndhF, a characteristic also present in other species of the tribe Cocoseae (de Souza Magnabosco et al., 2020de Souza Magnabosco, JW, de Freitas Fraga, HP, da Silva, RS, Rogalski, M, de Souza, EM, Guerra, MP and Vieira, LDN (2020) Characterization of the complete plastid genome of Butia eriospatha (Arecaceae). Genet Mol Biol 43:e20200023.). There was a marked intergeneric divergence when comparing the length of the rpl22-rps19 (IRb/LSC) and rps19-psbA (IRa/LSC) intergenic spacers (Figure 1). These junctions are widely described in the literature as regions of greater dynamism among angiosperm plastomes (Zhu et al., 2016Zhu A, Guo W, Gupta S, Fan W and Mower JP (2016) Evolutionary dynamics of the plastid inverted repeat: The effects of expansion, contraction, and loss on substitution rates. New Phytol 209:1747-1756. ). The IRs are the most conserved elements of plastomes, and the rate of synonymous substitutions in IRs is on average 3.7 times lower than in SSC due to a dose-dependent effect (Zhu et al., 2016Zhu A, Guo W, Gupta S, Fan W and Mower JP (2016) Evolutionary dynamics of the plastid inverted repeat: The effects of expansion, contraction, and loss on substitution rates. New Phytol 209:1747-1756. ). However, the borders between the IRs and SSC are more dynamic and during evolution they have gone through several lineage-specific contraction and expansion events (Goulding et al., 1996Goulding SE, Wolfe KH, Olmstead RG and Morden CW (1996) Ebb and flow of the chloroplast inverted repeat. Mol Gen Genet 252:195-206).

Plot of Bactridinae plastomes comparing the IR junctions sites. Genes transcribed clockwise and counterclockwise are shown above and below of their corresponding tracks, respectively. JLB - IRb/LSC junction; JSB - IRb/SSC junction; JSA - SSC/IRa junction; and JLA - IRa/LSC junction.
The collinearity analysis indicated the conservation of most of the plastome, as shown by the three LCBs identified by Mauve alignment (Figure 2). Bactris riparia and B. gasipaes are identical in gene content and order, with no rearrangements identified. Divergence was verified in A. murumuru and A. aculeatum, in which there is an inversion of 4.6 kb. This lineage-specific rearrangement, described by Lopes et al. (2019Lopes, AS, Pacheco, TG, Silva, ON, Cruz, LM, Balsanelli, E, Souza, EM, Pedrosa, FO and Rogalski, M (2019) The plastomes of Astrocaryum aculeatum G. Mey. and A. murumuru Mart. show a flip-flop recombination between two short inverted repeats. Planta250:1229-1246. ), is from a flip-flop recombination between 28 bp inverted repeats, which flank the 4.6 kb inversion and is in the trnT-UGU/ndhC and trnL-UAA/trnV-UAC intergenic spacers.

Multiple alignment on Mauve comparing species of Bactridinae. Colors in the alignment represent the Locally Collinear Blocks (LCBs). Annotated genes are shown as boxes below the alignment where annotated CDS features are shown as white boxes, tRNAs are green, rRNAs are red.
The prediction of SSRs revealed a total of 69 SSRs. Among them, homopolymers were the most common (61 occurrences), followed by dipolymers (7 occurrences) and tetrapolymers (1 occurrence). Among the 61 monolymers, 57 were consisted of A/T sequences and all presented 15 repeats or less, which is in accordance to the nature of plastid SSRs of generally <15 mononucleotide repeats. Tri-, penta- and hexapolymers were not present according to the analysis settings used here.
The synonymous (Ks) and non-synonymous (Ka) substitution rates, and the ratio (Ka/Ks) were calculated for 52 genes. Among them, 27 genes had no changes in the synonymous or nonsynonymous rates (Table S1 Table S1 - Non-synonymous (Ka), synonymous (Ks), and Ka/Ks ratio of Bactridinae protein-coding genes. ). The rbcL gene showed the highest non-synonymous rate (Ka = 0.00546), while the rps15 gene had the highest synonymous rate (Ks = 0.02266). Three genes (ccsA, cemA, and rpoC1) presented evidence of positive selection (Ka/Ks ratio > 1.0) and 22 genes showed evidence of purifying selection (Ka/Ks ratio < 1.0; Table S1 Table S1 - Non-synonymous (Ka), synonymous (Ks), and Ka/Ks ratio of Bactridinae protein-coding genes. ).
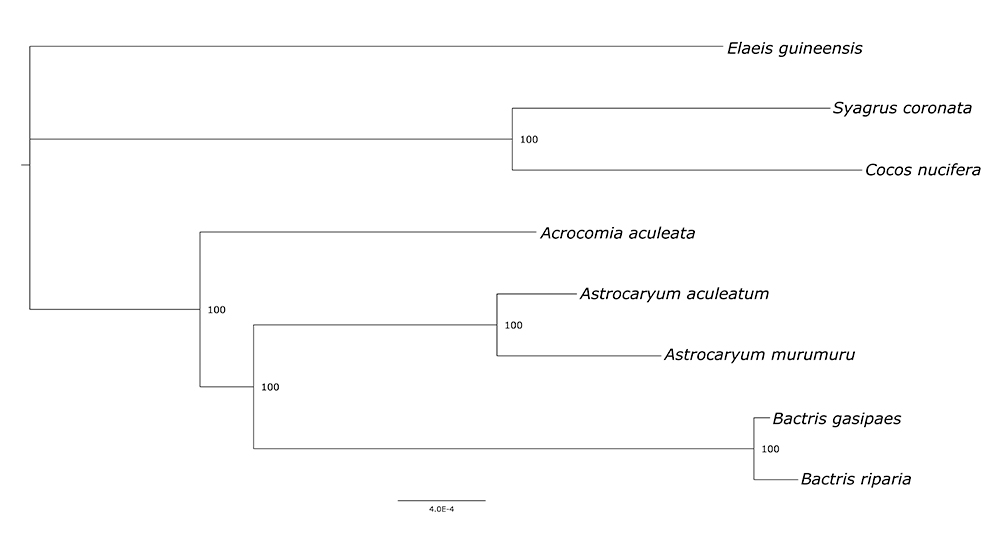
The phylogenetic inference of Bactridinae by ML consisted of an alignment of eight sequences with 133,566 columns, among which were 718 distinct patterns, 787 parsimony-informative sites, 972 singleton sites, and 131,807 constant sites. The resulting phylogeny presented highly supported nodes and intergeneric relationships (Figure 3). The topology obtained here corroborated the well-reported monophyly of the subtribe Bactridinae and the genus Bactris (Eiserhardt et al., 2011Eiserhardt WL, Pintaud JC, Asmussen-Lange C, Hahn WJ, Bernal R, Balslev H and Borchsenius F (2011) Phylogeny and divergence times of Bactridinae (Arecaceae, Palmae) based on plastid and nuclear DNA sequences. Taxon60:485-498.). It also shows Bactris as more closely related to Astrocaryum than to Acrocomia, which was previously recorded by Eiserhardt et al. (2011Eiserhardt WL, Pintaud JC, Asmussen-Lange C, Hahn WJ, Bernal R, Balslev H and Borchsenius F (2011) Phylogeny and divergence times of Bactridinae (Arecaceae, Palmae) based on plastid and nuclear DNA sequences. Taxon60:485-498.), Faurby et al. (2016Faurby S, Eiserhardt WL, Baker WJ and Svenning JC (2016) An all-evidence species-level supertree for the palms (Arecaceae). Mol Phylogenet Evol 100:57-69.) and Santos da Silva et al. (2021Santos da Silva, R, Clement, CR, Balsanelli, E, Baura, VA, Souza, EM, Fraga, HPF and do Nascimento Vieira, L (2021) The plastome sequence of Bactris gasipaes and evolutionary analysis in tribe Cocoseae (Arecaceae). PLoS One 16:e0256373. ). The positioning of Acrocomia in Bactridinae has presented a series of inconsistencies over the years, which could be elucidated by a more complete sampling of Bactridinae plastomes, including Aiphanes and Desmoncus.

Acknowledgements
This research was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) [grant numbers LNV 435200/2018-6, CRC 303477/2018-0] . The authors thank João Bosco Duarte Cintrão for collecting the plant material. This research was also partially supported by Embrapa’s Portfólio de Recursos Genéticos, which helped INPA to maintain the Peach palm Active Germplasm Bank.
References
- Amiryousefi A, Hyvönen J and Poczai P (2018) IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 34:3030-3031.
- Baker WJ and Dransfield J (2016) Beyond Genera Palmarum: Progress and prospects in palm systematics. Bot J Linn Soc 182:207-233.
- Beier S, Thiel T, Münch T, Scholz U and Mascher M (2017) MISA-web: A web server for microsatellite prediction. Bioinformatics 33:2583-2585.
- Boussardon C, Salone V, Avon A, Berthomé R, Hammani K, Okuda K, Shikanai T, Small I and Lurin C (2012) Two interacting proteins are necessary for the editing of the ndhD-1 site in Arabidopsis plastids. Plant Cell 24:3684-3694.
- Cámara-Leret R, Paniagua-Zambrana N, Balslev H, Barfod A, Copete JC and Macía MJ (2014) Ecological community traits and traditional knowledge shape palm ecosystem services in northwestern South America. For Ecol Manage 334:28-42.
- Clement CR, Cristo-Araújo M, Coppens d’Eeckenbrugge G, Reis VM, Lehnebach R and Picanço-Rodrigues D (2017) Origin and dispersal of domesticated peach palm. Front Ecol Evol 5:148.
- Couvreur TL, Hahn WJ, Granville JJD, Pham JL, Ludena B and Pintaud JC (2007) Phylogenetic relationships of the cultivated Neotropical palm Bactris gasipaes (Arecaceae) with its wild relatives inferred from chloroplast and nuclear DNA polymorphisms. Syst Bot 32:519-530.
- Darling ACE, Mau B, Blattner FR and Perna NT (2004) Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res 14:1394-1403.
- de Souza Magnabosco, JW, de Freitas Fraga, HP, da Silva, RS, Rogalski, M, de Souza, EM, Guerra, MP and Vieira, LDN (2020) Characterization of the complete plastid genome of Butia eriospatha (Arecaceae). Genet Mol Biol 43:e20200023.
- Dransfield J, Uhl NW, Lange CBA, Baker WJ, Harley MM and Lewis CE (2008) Genera Palmarum: The Evolution and Classification of Palms. Kew Publishing, London, 732 p.
- Dierckxsens N, Mardulyn P and Smits G (2016) NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res 45:e18.
- Eiserhardt WL, Pintaud JC, Asmussen-Lange C, Hahn WJ, Bernal R, Balslev H and Borchsenius F (2011) Phylogeny and divergence times of Bactridinae (Arecaceae, Palmae) based on plastid and nuclear DNA sequences. Taxon60:485-498.
- Faurby S, Eiserhardt WL, Baker WJ and Svenning JC (2016) An all-evidence species-level supertree for the palms (Arecaceae). Mol Phylogenet Evol 100:57-69.
- Goulding SE, Wolfe KH, Olmstead RG and Morden CW (1996) Ebb and flow of the chloroplast inverted repeat. Mol Gen Genet 252:195-206
- Henderson A (2000) Bactris (Palmae). Organization for Flora Neotropica, 181p.
- Lopes, AS, Pacheco, TG, Silva, ON, Cruz, LM, Balsanelli, E, Souza, EM, Pedrosa, FO and Rogalski, M (2019) The plastomes of Astrocaryum aculeatum G. Mey. and A. murumuru Mart. show a flip-flop recombination between two short inverted repeats. Planta250:1229-1246.
- Trifinopoulos J, Nguyen L, Haeseler A and Minh BQ (2016) W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res 44:W232-W235.
- Rogalski, M, do Nascimento, Vieira, L, Fraga, HP and Guerra, MP (2015) Plastid genomics in horticultural species: importance and applications for plant population genetics, evolution, and biotechnology. Front Plant Sci6:586.
- Santos da Silva, R, Clement, CR, Balsanelli, E, Baura, VA, Souza, EM, Fraga, HPF and do Nascimento Vieira, L (2021) The plastome sequence of Bactris gasipaes and evolutionary analysis in tribe Cocoseae (Arecaceae). PLoS One 16:e0256373.
- Velazco, SJE, Svenning, JC, Ribeiro, BR and Laureto, LMO (2020) On opportunities and threats to conserve the phylogenetic diversity of Neotropical palms. Divers Distrib 27:512-523.
- Zhu A, Guo W, Gupta S, Fan W and Mower JP (2016) Evolutionary dynamics of the plastid inverted repeat: The effects of expansion, contraction, and loss on substitution rates. New Phytol 209:1747-1756.
Supplementary material
The following online material is available for this article:
Edited by
Associate Editor
Publication Dates
-
Publication in this collection
07 Nov 2022 -
Date of issue
2022
History
-
Received
23 Sept 2021 -
Accepted
24 Sept 2022