Abstract
In 2000, an outbreak of sylvatic yellow fever possibly occurred in gallery forests of the Grande river in the Paraná basin in the northwestern region of São Paulo state. The aim of this study was to obtain information on the bionomics of Haemagogus and other mosquitoes inside tree holes in that area. Eighteen open tree holes were sampled for immature specimens. Adults were collected twice a month in the forest in Santa Albertina county from July 2000 to June 2001. The seasonal frequency of fourth instars was obtained by the Williams geometric mean (Mw), while the adult frequency was estimated either by hourly arithmetic or the Williams' means. Cole's index was applied to evaluate larval inter-specific associations. Among the ten mosquito species identified, the most abundant was Aedes terrens Walker followed by Sabethes tridentatus Cerqueira and Haemagogus janthinomys Dyar. Larval and adult abundance of these species was higher in summer than in winter. Although larval abundance of Hg. janthinomys peaked in the rainy season, correlation with rainfall was not significant. Six groups of larval associations were distinguished, one of which the most positively stable. The Hg. janthinomys and Ae. terrens association was significant, and Limatus durhamii Theobald was the species with most negative associations.
Sabethes tridentatus; Aedes terrens; Limatus durhamii; sylvatic yellow fever; immature seasonality; adult abundance
PUBLIC HEALTH
Observations on Haemagogus janthinomys Dyar (Diptera: Culicidae) and other mosquito populations within tree holes in a gallery forest in the northwestern region of Sao Paulo state, Brazil
Rosa M TubakiI; Regiane M T de MenezesI; Fabiana T VesgueiroII; Rubens P Cardoso JrIII
ILab de Entomologia Médica, Superintendência de Controle de Endemias, Rua Paula Souza 166/ 5º andar, 1027-000 São Paulo, SP, Brasil; tubaki.rm@gmail.com, rmtironi@gmail.com
IIDepto de Epidemiologia, Fac de Saúde Pública,USP, Av Dr Arnaldo 715, 01246-904 São Paulo, SP, Brasil; fabianatavares@usp.br
IIILab Regional, Av Philadelpho Manoel Gouveia Neto 3101, 15060-040 São José do Rio Preto, SP, Brasil; rcardosojr@yahoo.com.br
ABSTRACT
In 2000, an outbreak of sylvatic yellow fever possibly occurred in gallery forests of the Grande river in the Paraná basin in the northwestern region of São Paulo state. The aim of this study was to obtain information on the bionomics of Haemagogus and other mosquitoes inside tree holes in that area. Eighteen open tree holes were sampled for immature specimens. Adults were collected twice a month in the forest in Santa Albertina county from July 2000 to June 2001. The seasonal frequency of fourth instars was obtained by the Williams geometric mean (Mw), while the adult frequency was estimated either by hourly arithmetic or the Williams' means. Cole's index was applied to evaluate larval inter-specific associations. Among the ten mosquito species identified, the most abundant was Aedes terrens Walker followed by Sabethes tridentatus Cerqueira and Haemagogus janthinomys Dyar. Larval and adult abundance of these species was higher in summer than in winter. Although larval abundance of Hg. janthinomys peaked in the rainy season, correlation with rainfall was not significant. Six groups of larval associations were distinguished, one of which the most positively stable. The Hg. janthinomys and Ae. terrens association was significant, and Limatus durhamii Theobald was the species with most negative associations.
Key words:Sabethes tridentatus, Aedes terrens, Limatus durhamii, sylvatic yellow fever; immature seasonality; adult abundance
The first outbreak of sylvatic yellow fever (SYF) occurred in Brazil in 1908. In the thirties, the disease spread into the States of Goias, Minas Gerais and Mato Grosso. Soper (1936) used the term "jungle yellow fever" to differentiate the sylvatic disease from the rural form transmitted by Aedes aegypti Linnaeus. Further studies (Soper et al 1932, Lane 1936, Antunes & Whitmani 1937, Causey & Kumm 1948, Kumm 1950) described the diurnal sylvatic mosquitoes incriminated in the transmission of the SYF and among these Haemagogus janthinomys Dyar, the primary vector. The epidemy spread throughout the western region of the State of São Paulo, finally reaching the northwestern region of the state of Paraná. Lane (1936) identified Hg. janthinomys in localities with SYF in the western region of São Paulo, but those collections were performed as surveys in the counties. These surveys reported the occurrence of species but they did not add information about the environment or diel mosquito activity.
Studies on adult mosquito activity related to the influence of rainfall were addressed by Kumm (1950) in the locality of Passos, Minas Gerais. Although Dégallier et al (1992) introduced the concept of gallery forests as a phytoclimatic zones, gallery forests were better characterized by Forattini and Gomes (1988) as shelters for Haemagogus mosquitoes and non-human primates involved in maintaining the virus cycle and bridges for propagation along their course, in modified environments for agricultural practices. These sparsely forested areas are nowadays recognized as fragmented areas in the savannah biome.
In 2000, sixty-four years after the first record of Hg. janthinomys, two cases of SYF occurred in the northwestern region of São Paulo, one in the Santa Albertina county and another in the Ouroeste county. There is not much information about the immatures of sylvatic mosquitoes bred inside tree holes that could help to clarify the SYF epidemiology. The aim of this work was to describe the seasonal frequency of immature and adult Hg. janthinomys forms and of other mosquitoes occurring inside tree holes in a gallery forest in Santa Albertina County located in the southeastern region of the savannah, a potential area of SYF transmission.
Material and Methods
The Rancho do Vale (50º 48' W, 19º 58'S) is a patch of gallery forest situated in the Santa Albertina county, Sao Paulo state, on the left margin of the Grande river. Santa Albertina county is located in the northwestern region of Sao Paulo State, Brazil and covers an area of 274 km2, most of which is rural. This county belongs to the eleventh region of the state. The climate is of Aw type, alternately dry and humid in that region, according to Koeppen. The mean values of rainfall and temperature oscillate between 150-1000 mm and 14-31ºC in the dry and wet seasons, respectively (Troppmair 1975). Gallery forests are corridors of evergreen mesophytic vegetation present in the savannah (Tubaki et al 2004).
Larval collections were carried out fortnightly on the Rancho do Vale gallery forest from July 2000 to June 2001. Trees were examined every 5 m, eighteen open tree holes per tree on average. Samples of larvae and pupae were transported to the laboratory and kept alive until the imago state. Adult were then killed and immature ecdyses preserved and mounted for identification. The tree holes with continuous larval production were chosen for analysis and the seasonal abundance of the most frequent larval mosquito species found in the tree holes was estimated by using Williams' mean (Haddow 1960) and standard errors to standardize results.
The adult collections followed larval sampling on two subsequent days. Two men (previously immunized against YF) carried out catches for 15 min every hour. Collections were undertaken on humans in movement, with the aim of improving the efficiency in collecting Hg. janthinomys females through constant human intrusion into the forest. The adult collection period ranged from 9:00h to 16:00h and an hourly average per month was calculated with standard errors to standardize results.
Daily rainfall records were obtained from the Casa da Agricultura in Santa Albertina whereupon monthly records were compared with decennial ones (July 1989 through June 1999) by the non-parametric Man-Whitney (U) test to verify whether they were anomalous at the period of study. Rainfall was accumulated over four weeks prior each larval collection and the relationship between species abundances and accumulated rainfall, was tested for Spearman correlations.
An evaluation of the association between tree hole mosquito species was done to provide information on those species sharing tree holes with Hg. janthinomys in the Rancho do Vale gallery forest. Cole's coefficient of interspecific association (CAB) (1949) takes into consideration the occurrence of two coexistent species in collections. The values change from a perfect negative association (-1), when habitat requirements are distinct or one species has the tendency to exclude the other, to a perfect positive association (+1), when the species have both overlapping habitats and requirements or interact to favor mutual presence. A value of zero represents random or neutral association. The Cole index was tested for significance by using the Fisher test when values were equal or lower than 5.
Results
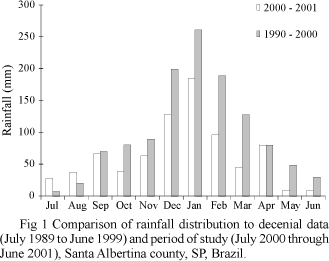
Rainfall data. The records of rainfall over the study period were compared with those of decennial one whereby they showed similar curves (U = 60.0; P > 0.05), thereby suggesting that the distribution of rainfall was typically seasonal during the period. There was a total rainfall of 1, 412, 6 mm throughout with a peak in January (Fig 1).
Larval abundance. A total of 10 mosquito species were identified in the larval survey (Table 1). The most abundant species (50%) was Aedes terrens Walker with 154 specimens (males and females) followed by Sabethes tridentatus Cerqueira (24.4%) and Hg. janthinomys (8.8%) (Table 1).
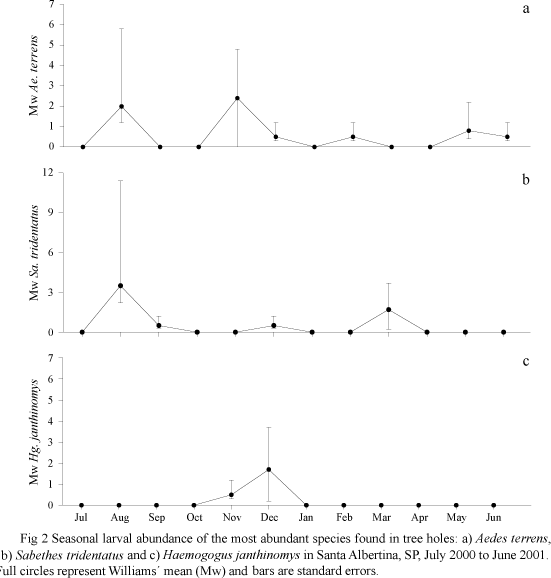
Larval seasonal abundance of Ae. terrens and Sa. tridentatus were highest in the wet season between November and March, with Mw (Williams' means) values equal to 2.4 and 1.7, respectively. Their larval abundance also peaked in the dry season and reached Mw values of 2.4 for Ae. terrens and 3.5 for Sa. tridentatus. The larval abundance of Hg. janthinomys was only slightly correlated with rainfall, but with no statistical support (rs = 0.03, P = 0.8). The larval abundances of Sa. tridentatus (rs = 0.18, P = 0.2) were not correlated with rainfall at all (Fig 2).
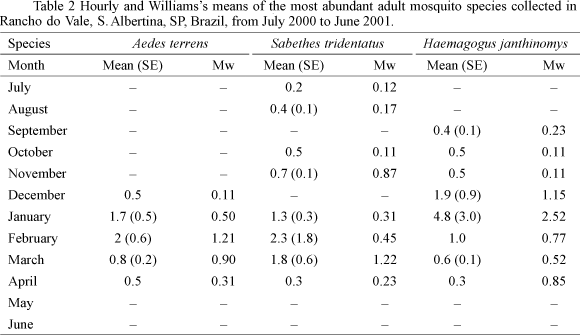
Adult seasonal abundance. Adult abundance of Ae. terrens, Sa. tridentatus and Hg. janthinomys was also highest during the rainy season, with Hg. janthinomys abundance being twice as high the abundance of the other two species. Aedes terrens and Sa. tridentatus adult abundance peaked in February whereas in Hg. janthinomys this occurred in January. Values obtained as arithmetic means were almost twice of those of the Williams' mean. Differences between the arithmetic mean and Williams' mean were explained by the amount of hours considered in the calculation (Table 2).
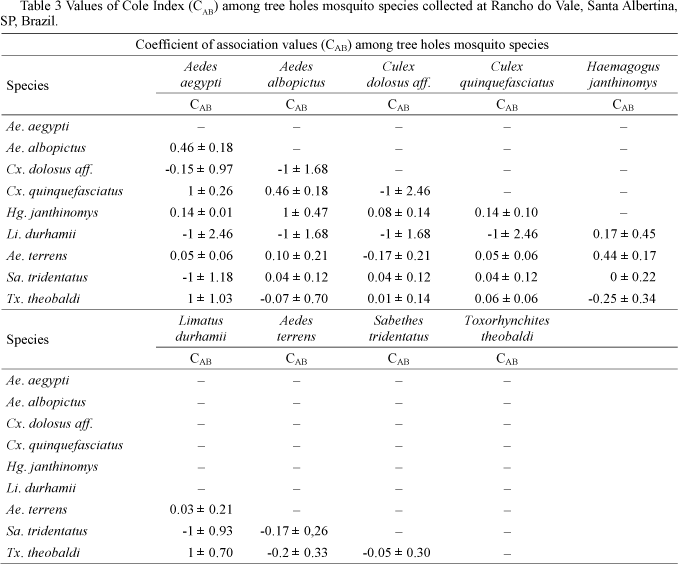
Cole's larval species associations. A comparison of interspecific association coefficient values (CAB) among nine larval species from the tree holes allowed for the discrimination of six association groups. As shown in Table 3, there were four pairs of perfectly negatively associated species (-1 ± 2.46 to -1.18). Limatus durhamii Theobald was the species with the most negative associations (5) from all pairs compared. Completely negative associations occurred between Sa. tridentatus and Ae. aegypti, besides Li. durhamii and Ae. aegypti, Ae. albopictus Skuse, Sa. tridentatus and Cx. quinquefasciatus Say. The second group was slightly negatively associated (-0.25 ± 0.34 to -0.04 ± 0.30). In this group, Tx. theobaldi (Dyar and Knab) was negatively associated with both Hg. janthinomys and Ae. terrens. The negative association between Tx. theobaldi and Ae. terrens was significant (P < 0.05). Twelve pairs of species were included in the third group (0.01 ± 0.14 to 0.16 ± 0.45) whose associations were considered neutral. In this group, Hg. janthinomys was only slightly associated with Cx. dolosus affinis Lynch Arribalzaga and occurred in three of four pairs of associations, which were similar to those associations with Cx. quinquefasciatus and Ae. aegypti. The fourth group was characterized by positive associations (0.44 ± 0.17 to 0.46 ± 0.18) and revealed less variation when compared with the other groups. In this group. Hg. janthinomys was significantly associated with Ae. terrens (P < 0.001). The fifth group was constituted by four pairs with perfect positive associations Ae. aegypti and Cx. quinquefasciatus, Ae. aegypti and Tx. theobaldi, Ae. albopictus and Hg. janthinomys, Tx. theobaldi and Li. durhamii (Table 3).
Fig 3 shows the distribution of the Cole's indexes to mosquito species associated to the two highly synantropic (Ae. aegypti and Ae. albopictus) and to the two sylvatic aedine species (Ae. terrens and Hg janthinomys). Species indexes associated to the most synantropic species are shown in CAB1, and their values are not repeated in CAB2. The variation of the coefficient values was larger in CAB1 than in CAB2.
Discussion
The composition of immature species inside open tree holes is restricted to only a few species and is quite specific, having been previously described in a survey on natural microhabitats undertaken in Tremembe County (Gomes et al 1992), another phytogeographical region in the State of Sao Paulo. Haemagogus janthinomys has not been recorded in Sao Paulo since Lane's reports in 1936. At that time, Lane identified Hg. janthinomys in the Alta Araraquarense, a region that included Santa Albertina County and was formerly named Tanaby. Haemagogus janthinomys and Haemagogus capricornii Lutz are very much alike, they have the same range of distribution in the southeastern region of Brazil and are only distinguishable by the male genitalia. We confirmed the specific status of Hg. janthinomys by rearing the larvae collected in the tree holes until the imago stage and by identifying the specimens by using characters of the male genitalia. Haemagogus janthinomys adult seasonal activity in Santa Albertina was similar to that observed in Trinidad (Chadee 1992), with a small number of females in the dry season and high abundance in the wet season. If Hg. janthinomys number of females collected monthly in Chaguaramas Forest (Chadee 1992) is reduced proportionally to 15 min, the results are similar to those reported in this study. The diel activity of Hg. janthinomys adults is not described in our study although the period of collection varied between 9:00h and 16:00h, coinciding with the period of activity reported for Hg. janthinomys and close related species (Forattini & Gomes 1988, Chadee 1992, Camargo-Neves et al 2005). Our data also suggest that Hg. janthinomys is to be found in drier areas than Hg. capricornii in Sao Paulo, in accordance with the distribution of sylvatic yellow fever risk areas (Dégallier et al 1992).
The correlation of mosquito larval abundance in ephemeral breeding sites with rainfall was tentatively shown in our study area. This relationship is currently pursued in the investigations to determine the risk of YF transmission. Dégallier et al (2006) inferred a time lapse of 11 to 12 days between egg hatching induced by rainfall and adult emergence. They also pointed out the occurrence of YF cases two months after heavy rains corroborating Chadee (1992).
Haemagogus janthinomys biting activity peaked in January when rainfall was more intense and the beginning of symptoms in humans in March, all according to data of the epidemiological vigilance service, seem to be in agreement with Dégallier's hypothesis. This might also suggest that the filling of tree holes by rainfall would be simultaneous with and synchronize the egg hatching and larval development towards immediate imago formation as in other ephemeral breeding sites. Nevertheless, rainfall might cause the flooding of open tree holes, with a subsequent loss or decrease in Hg. janthinomys, Ae. terrens and Sa. tridentatus populations. The difficulty to assign a sampling program adjusted to day-lags and rainfall would explain the low larval abundance and the ineffective attempts to correlate it with rainfall.
The abundance of Sa. tridentatus immatures, increased in the dry and post-wet seasons when compared with the wet season. Nevertheless, the abundance of Sa. tridentatus females in the dry and wet seasons was in agreement with the pattern in Sabethes chloropterus (Von Humboldt) in Trinidad (Chadee 1990). As shown in a prior evaluation (Rocco et al 2003), Sa. tridentatus does not seem to be associated with the transmission of YF.
Aedes terrens is quite adaptive occurring in several habitats (Lopes et al 2004). It occupied natural breeding sites as observed in this study, but may also dwell in artificial containers, mainly in patches of riparian forest in modified environments (Zequi et al 2005).
The Cole index of insterspecific association as related to the two synanthropic species suggests that the associations between Ae. aegypti and Ae. terrens, Ae. aegypti and Hg. janthinomys, Ae. aegypti and Ae. albopictus were casual, whereas the same situation is presented in associations between Ae. albopictus and Tx. theobaldi, and Ae. albopictus and Sa. tridentatus (Fig 3a). On considering the sylvatic species, casual associations should be suggested between Cx. quinquefasciatus and Ae. terrens, Ae. terrens and Li. durhamii and Hg. janthinomys and Sa. tridentatus (Fig 3b). Although the invasive Ae. aegypti was found within the tree holes, it has been pointed out that Ae. aegypti activity made it more vulnerable to predation when compared with other species (Grill & Juliano 1996) from the Aedine tribe. It is likely that some associations with Ae. aegypti are usual and stable compared to others, and might be explained by feeding habits or unknown competitive interactions.
The slight variation in the Cole index obtained for sylvatic species shows that Hg. janthinomys and Cx. quinquefasciatus, and specially Ae. terrens and Hg. janthinomys associations are usual, and possibly the species shared the same requirements in terms of physical environmental conditions and food abundance in tree holes. The associations Ae. albopictus and Ae. terrens on one hand and Ae. albopictus and Hg. janthinomys on the other were positive and stable. It also indicates the restrained behavior of the invasive Ae. albopictus in exploring natural breeding sites as opposed to the synanthropic species (Lounibos et al 2001, Braks et al 2003). Special concern should also be given to the ubiquitous behavior of Ae. albopictus. This observation suggests that positively associated species may not have the same obligatory feeding behavior, but are flexible, depending on the variation in food abundance, be it regulated by predators (Alto et al 2005, Merritt et al 1992) or by the quality of tree hole environment. Flexible habits could possibly explain the nature of mosquito larval associations. Although mosquito species relationships are poorly understood in natural containers, experimental investigation gives evidence of these associations although restricted to invasive species, such as Ae. albopictus and Ae. aegypti (Juliano 2009).
Even if the attempt to correlate rainfall and the larval abundance for the most frequent species was ineffective, we presume that the wet season exerted a generally positive effect on the abundance of Hg. janthinomys and other tree hole mosquitoes, thereby suggesting that greater epidemiological vigilance should be exercised in YF risk during the wet season.
Acknowledgments
We are grateful to Dr Maria A M Sallum for confirming the male specimens of Hg. janthinomys, to the field team of Serviço Regional 6 and Serviço Regional 8, and also to the technicians Maria A dos Santos and Luís M Bonafé for field and laboratory support.
Received 12/III/09.
Accepted 26/I/10.
Edited by Neusa Hamada INPA
- Alto B W, Griswold M W, Lounibos L P (2005) Habitat complexity and sex-dependent predation of mosquito larvae in containers. Oecologia 146: 300-310.
- Antunes P C A, Whitman L (1937) Studies on the capacity of mosquitoes of the genus Haemagogus to transmit yellow fever. Am J Trop Med Hyg 17: 825-831.
- Braks M A, Honorio N A, Lourenco-de-Oliveira R, Juliano S A, Lounibos L P (2003) Convergent habitat segregation of Aedes aegypti and Aedes albopictus (Diptera, Culicidae) in southeastern Brazil and Florida. J Med Entomol 40: 785-794.
- Causey O R, Kumm W H (1948) Dispersion of forest mosquitoes in Brazil. Preliminary studies. Amer J Trop Med Hyg 28: 469-480.
- Camargo-Neves V, Poletto D W, Rodas L A C, Pacchioli M, Cardoso R P, Scandar S A S, Sampaio S M P, Koyanagui P H, Botti M V, Mucci L F, Gomes A C (2005) Investigação entomológica em área de ocorrência de febre amarela silvestre no estado de São Paulo. Cad Saude Públ 21: 1278-1286.
- Chadee D D (1990) Seasonal abundance and diel landing periodicity of Sabethes chloropterus (Diptera: Culicidae) in Trinidad, West Indies. J Med Entomol 27: 1041-1044.
- Chadee D D, Tikasingh E S, Ganesh R (1992) Seasonality, biting cycle and parity of the yellow fever vector mosquito Haemagogus janthinomys in Trinidad. Med Vet Entomol 6: 143-148.
- Cole L C (1949) The measurement of interspecific association. Ecology 30: 411-424.
- Dégallier N, Monteiro H O, Castro F C, Da Silva O, Sá Filho G C, Elguero E (2006) An indirect estimation of the developmental time of Haemagogus janthinomys (Diptera: Culicidae), the main vector of yellow fever in South America. Stud Neotrop Fauna Environ 41: 117-122.
- Dégallier N, Travassos da Rosa A P A, Hervé J P, Rosa J F S T, Vasconcelos P F C, Silva C J M, Barros V L R S, Dias L B, Rosa E S T, Rodrigues S B (1992) A comparative study of yellow fever in Africa and South America. Cienc Cult 44: 143-151.
- Forattini O P, Gomes A C (1988) Biting activity of Aedes scapularis (Rondani) and Haemagogus mosquitoes in Southern Brazil (Diptera: Culicidae). Rev Saude Públ 22: 84-93.
- Gomes A C, Forattini O P, Kakitani I, Marques G R A, Marques C C A, Marucci D, Brito M (1992) Microhabitats de Aedes albopictus (Skuse) na região do Vale do Paraíba, estado de São Paulo, Brasil. Rev Saude Públ 26: 108-118.
- Grill C P, Juliano S A (1996) Predicting species interactions based on behavior: predation and competition in container-dwelling mosquitoes. J Anim Ecol 65: 63-81.
- Haddow A J (1960) Studies on the biting habits and medical importance of East African mosquitoes in the genus Aedes I - Subgenera Aedimorphus, Banksinella and Dunnius Bull Entomol Res 50: 759-779.
- Juliano S A (2009) Species interactions: among larval mosquitoes: context dependence across habitat gradients. Annu Rev Entomol 54: 37-56.
- Kumm H W (1950) Seasonal variations in rainfall: prevalence of Haemagogus and incidence of jungle yellow fever in Brazil and Colombia. Trans R Soc Trop Med Hyg 43: 673-682.
- Lane J (1936) Notas sobre investigações entomológicas em localidades onde houve febre amarela sylvestre em São Paulo. Arq Hyg Saúde Publ 1: 127-133.
- Lopes J, Martins E A C, Oliveira O, Oliveira V, Oliveira Neto B P, Oliveira J E (2004) Dispersion de Aedes aegypti (Linnaeus, 1762) e Aedes albopictus (Skuse, 1894) in the rural zone of North Paraná state. Braz Arch Biol Tecnol 47: 739-746.
- Lounibos L P, O'Meara G F, Escher R L, Nishimura N, Cutwa M, Nelson T, Campos R E, Juliano S (2001) Testing predicted competitive displacement of native Aedes by the invasive Asian tiger mosquito Aedes albopictus in Florida, USA. Biol Invasions 3: 151-166.
- Merritt R W, Dadd R H, Walker E D (1992) Feeding behavior, natural food, and nutritional relationships of larval mosquitoes. Annu Rev Entomol 37: 349-376.
- Reinert J F (1975) Mosquito generic and subgeneric abbreviations (Diptera: Culicidae). Mosq Syst 7: 105-110.
- Reinert J F (1982) Abbreviations form mosquito generic and subgeneric taxa established since 1975 (Diptera: Culicidae). Mosq Syst 14: 124-126.
- Reinert J F (1991) Additional abbreviations of mosquito subgenera: names established since 1982 (Diptera: Culicidae). Mosq Syst 23: 209-210.
- Rocco I M, Katz G, Tubaki R M (2003) Febre amarela silvestre no estado de São Paulo, Brasil: casos humanos autóctones. Rev Inst Adolfo Lutz 62: 201-206.
- Soper F L (1936) Febre amarella silvestre, novo aspecto epidemiológico da doença. Rev Hyg e Saúde Públ 10: 107-144.
- Soper F L, Penna H, Cardoso E, Serafin J, Frobisher M, Pinheiro J (1932) Yellow fever without Aedes aegypti Study of a rural epidemic in the Valle do Canaan, Espírito Santo, Brazil. Am J Hyg 18: 555-587.
- Troppmair H (1975) Regiões ecológicas do estado de São Paulo. Biogeografia 10: 1-24.
- Tubaki R M, Menezes R M T, Cardoso Jr R P, Bergo E S (2004) Studies on entomological monitoring: mosquito species frequency in riverine habitats of the Igarapava dam, southern region, Brazil. Rev Inst Med trop 46: 223-229.
- Zequi J A C, Lopes J, Medri I M (2005) Immature specimens of Culicidae (Diptera) found in installed recipients in forest fragments in the Londrina, Paraná, Brazil. Rev Bras Zool 22: 656-661.
Publication Dates
-
Publication in this collection
14 Sept 2010 -
Date of issue
Aug 2010
History
-
Received
12 Mar 2009 -
Accepted
26 Jan 2010