Resumos
Coelhos são susceptíveis à infecção pelo herpes-vírus bovino tipo 5 (BHV-5) e freqüentemente desenvolvem enfermidade neurológica aguda fatal após inoculação intranasal. A cinética da invasão do sistema nervoso central (SNC) de coelhos pelo BHV-5 foi estudada através de pesquisa de vírus em secções do SNC a diferentes intervalos pós-inoculação. Após inoculação intranasal, o vírus foi inicialmente detectado no bulbo olfatório às 48h, seguido do córtex olfatório às 48/72h. Às 72/96h o vírus foi detectado também no gânglio trigêmeo, ponte e córtex cerebral. Dois experimentos foram realizados para avaliar a importância do sistema olfatório na invasão do SNC de coelhos pelo BHV-5. No primeiro experimento, coelhos foram inoculados com duas amostras do BHV-5 no saco conjuntival. Coelhos inoculados por essa via também desenvolveram a enfermidade neurológica, porém com menor freqüência com curso clínico tardio. No segundo experimento, doze coelhos foram submetidos à ablação cirúrgica do bulbo olfatório e posteriormente inoculados com o BHV-5 pela via intranasal. Onze de 12 coelhos controle (91,6%), não submetidos à cirurgia, desenvolveram a doença neurológica, contra quatro de 12 (33,3%) dos animals submetidos à remoção cirúrgica do bulbo olfatório. Esses resultados demonstram que o sistema olfatório constitui-se na principal via de acesso do BHV-5 ao encéfalo de coelhos após inoculação intranasal. No entanto, o desenvolvimento de infecção neurológica em coelhos inoculados pela via conjuntival e em coelhos sem o bulbo olfatório indica que o BHV-5 pode utilizar outras vias para invadir o SNC, provavelmente as fibras sensoriais e autonômicas que compõe o nervo trigêmeo. Os efeitos da imunização com vírus homólogo (BHV-5) e heterólogo (BHV-1) na proteção à infecção neurológica foram investigados. Cinco entre 10 coelhos (50%) imunizados com o BHV-5 apresentaram sinais neurológicos discretos e transitórios e um morreu após o desafio com o BHV-5. Curiosamente, o grau de proteção foi superior nos coelhos imunizados com o BHV-1: apenas dois animais apresentaram sinais clínicos passageiros e recuperaram-se. Portanto, proteção da enfermidade neurológica pelo BHV-5 em coelhos pode ser obtida por imunização com o BHV-5 ou BHV-1, provavelmente devido à extensa reatividade sorológica cruzada entre esses vírus. Estudos adicionais em coelhos podem auxiliar no esclarecimento da patogênese e resposta imunológica a infecção pelo BHV-5.
Herpesvírus Bovino tipo 5; BHV-5; meningoencefalite; coelhos; modelo experimental
Rabbits are susceptible to bovine herpesvirus type 5 (BHV-5) infection and often develop an acute and fatal neurological disease upon intranasal inoculation. The kinetics of viral infection of the central nervous system (CNS) was investigated by testing serial brain sections for infectivity at intervals after virus inoculation. The virus was first detected in the main olfactory bulb at 48h, followed by the olfactory cortex at 48/72h. At 72/96h infectivity was also detected in the trigeminal ganglia, pons and cerebral cortex. Two experiments were conducted to investigate the role of the olfactory system in the invasion of the rabbits' CNS by BHV-5. In the first experiment, rabbits were inoculated with two BHV-5 isolates in the conjunctival sac. Rabbits inoculated by this route developed the neurological disease, yet with a reduced frequency and delayed clinical course. In a second experiment, twelve rabbits were submitted to surgical removal of the olfactory bulb and subsequently inoculated intranasally with BHV-5. Eleven out of 12 (91.6%) of the control rabbits developed the disease, against four out of 12 (33.3%) of the animals lacking the olfactory bulb. These results suggest that the olfactory system is the main pathway utilized by BHV-5 to reach the CNS of rabbits after intranasal inoculation. Nevertheless, the development of neurological infection in rabbits inoculated in the conjunctival sac and in rabbits lacking the olfactory bulb indicate that BHV-5 may utilize an alternative route to invade the CNS, probably the sensory and autonomic fibers of the trigeminal nerve. The effects of immunization with homologous (BHV-5) and heterologous (BHV-1) strains in prevention of neurological disease by BHV-5 were investigated. Five out of 10 rabbits (50%) immunized with BHV-5 showed mild and transient neurological signs and one died upon challenge. Interestingly, the degree of protection against BHV-5 challenge was higher in rabbits immunized with BHV-1: only two rabbits showed transient neurological signs and subsequently recovered. Thus, prevention of neurological disease by BHV-5 in rabbits may be achieved by immunization with either BHV-5 or BHV-1, likely reflecting the extensive serological cross-reactivity between these viruses. Further studies in rabbits may help in understanding the pathogenesis and immune response to BHV-5 infection.
Bovine herpesvirus type 5 (BHV-5); meningo-encephalitis; rabbits; experimental model
Infecção e enfermidade neurológica pelo herpesvírus bovino tipo 5 (BHV-5): coelhos como modelo experimental1 1 Aceito para publicação em 14 de junho de 2000.
Nilzane Beltrão2 2 Mestranda em Medicina Veterinária, Universidade Federal de Santa Maria (UFSM), Santa Maria, RS. , Eduardo F. Flores3 3 Depto Medicina Veterinária Preventiva e Depto Microbiologia e Parasitologia, UFSM. * * Autor para correspondência: DMVP/CCR/UFSM, 97105-900 Santa Maria, RS. Fone/fax:(055)220-8034. E-mail: Flores@ccr.ufsm.br , Rudi Weiblen3 3 Depto Medicina Veterinária Preventiva e Depto Microbiologia e Parasitologia, UFSM. , Adriana M. Silva4 4 União Pioneira de Integração Social, UPIS, Brasília, DF. , Paulo M. Roehe5 5 Centro de Pesquisas Desidério Finamor (CPVDF), Eldorado do Sul, RS e Depto Microbiologia, Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, RS. e Luiz F. Irigoyen6 6 Departamento de Patologia, UFSM.
ABSTRACT .- Beltrão, N., Flores E.F., Weiblen R., Silva A.M., Roehe, P.M. & Irigoyen L.F. 2000. [Acute infection and neurological disease by bovine herpesvirus type-5 (BHV-5): Rabbits as an experimental model.] Infecção aguda e enfermidade neurológica pelo herpesvírus bovino tipo 5 (BHV-5): coelhos como modelo experimental. Pesquisa Veterinária Brasileira 20 (4):144-150. Depto Medicina Veterinária Preventiva, Universidade Federal de Santa Maria, 97105-900 Santa Maria, RS, Brazil.
Rabbits are susceptible to bovine herpesvirus type 5 (BHV-5) infection and often develop an acute and fatal neurological disease upon intranasal inoculation. The kinetics of viral infection of the central nervous system (CNS) was investigated by testing serial brain sections for infectivity at intervals after virus inoculation. The virus was first detected in the main olfactory bulb at 48h, followed by the olfactory cortex at 48/72h. At 72/96h infectivity was also detected in the trigeminal ganglia, pons and cerebral cortex. Two experiments were conducted to investigate the role of the olfactory system in the invasion of the rabbits' CNS by BHV-5. In the first experiment, rabbits were inoculated with two BHV-5 isolates in the conjunctival sac. Rabbits inoculated by this route developed the neurological disease, yet with a reduced frequency and delayed clinical course. In a second experiment, twelve rabbits were submitted to surgical removal of the olfactory bulb and subsequently inoculated intranasally with BHV-5. Eleven out of 12 (91.6%) of the control rabbits developed the disease, against four out of 12 (33.3%) of the animals lacking the olfactory bulb. These results suggest that the olfactory system is the main pathway utilized by BHV-5 to reach the CNS of rabbits after intranasal inoculation. Nevertheless, the development of neurological infection in rabbits inoculated in the conjunctival sac and in rabbits lacking the olfactory bulb indicate that BHV-5 may utilize an alternative route to invade the CNS, probably the sensory and autonomic fibers of the trigeminal nerve. The effects of immunization with homologous (BHV-5) and heterologous (BHV-1) strains in prevention of neurological disease by BHV-5 were investigated. Five out of 10 rabbits (50%) immunized with BHV-5 showed mild and transient neurological signs and one died upon challenge. Interestingly, the degree of protection against BHV-5 challenge was higher in rabbits immunized with BHV-1: only two rabbits showed transient neurological signs and subsequently recovered. Thus, prevention of neurological disease by BHV-5 in rabbits may be achieved by immunization with either BHV-5 or BHV-1, likely reflecting the extensive serological cross-reactivity between these viruses. Further studies in rabbits may help in understanding the pathogenesis and immune response to BHV-5 infection.
INDEX TERMS: Bovine herpesvirus type 5 (BHV-5), meningo-encephalitis, rabbits, experimental model.
RESUMO.- Coelhos são susceptíveis à infecção pelo herpes-vírus bovino tipo 5 (BHV-5) e freqüentemente desenvolvem enfermidade neurológica aguda fatal após inoculação intranasal. A cinética da invasão do sistema nervoso central (SNC) de coelhos pelo BHV-5 foi estudada através de pesquisa de vírus em secções do SNC a diferentes intervalos pós-inoculação. Após inoculação intranasal, o vírus foi inicialmente detectado no bulbo olfatório às 48h, seguido do córtex olfatório às 48/72h. Às 72/96h o vírus foi detectado também no gânglio trigêmeo, ponte e córtex cerebral. Dois experimentos foram realizados para avaliar a importância do sistema olfatório na invasão do SNC de coelhos pelo BHV-5. No primeiro experimento, coelhos foram inoculados com duas amostras do BHV-5 no saco conjuntival. Coelhos inoculados por essa via também desenvolveram a enfermidade neurológica, porém com menor freqüência com curso clínico tardio. No segundo experimento, doze coelhos foram submetidos à ablação cirúrgica do bulbo olfatório e posteriormente inoculados com o BHV-5 pela via intranasal. Onze de 12 coelhos controle (91,6%), não submetidos à cirurgia, desenvolveram a doença neurológica, contra quatro de 12 (33,3%) dos animals submetidos à remoção cirúrgica do bulbo olfatório. Esses resultados demonstram que o sistema olfatório constitui-se na principal via de acesso do BHV-5 ao encéfalo de coelhos após inoculação intranasal. No entanto, o desenvolvimento de infecção neurológica em coelhos inoculados pela via conjuntival e em coelhos sem o bulbo olfatório indica que o BHV-5 pode utilizar outras vias para invadir o SNC, provavelmente as fibras sensoriais e autonômicas que compõe o nervo trigêmeo. Os efeitos da imunização com vírus homólogo (BHV-5) e heterólogo (BHV-1) na proteção à infecção neurológica foram investigados. Cinco entre 10 coelhos (50%) imunizados com o BHV-5 apresentaram sinais neurológicos discretos e transitórios e um morreu após o desafio com o BHV-5. Curiosamente, o grau de proteção foi superior nos coelhos imunizados com o BHV-1: apenas dois animais apresentaram sinais clínicos passageiros e recuperaram-se. Portanto, proteção da enfermidade neurológica pelo BHV-5 em coelhos pode ser obtida por imunização com o BHV-5 ou BHV-1, provavelmente devido à extensa reatividade sorológica cruzada entre esses vírus. Estudos adicionais em coelhos podem auxiliar no esclarecimento da patogênese e resposta imunológica a infecção pelo BHV-5.
TERMOS DE INDEXAÇÃO: Herpesvírus Bovino tipo 5, BHV-5, meningoencefalite, coelhos, modelo experimental.
INTRODUÇÃO
O herpesvírus bovino tipo 5 (BHV-5) é um alfaherpesvírus associado com meningo-encefalite de curso geralmente fatal, que acomete principalmente bovinos jovens (Studdert 1989). Previamente classificado como um subtipo do herpesvírus bovino tipo 1 (BHV-1), o BHV-5 foi recentemente reclassificado com base em algumas características biológicas e moleculares que o distingue dos isolados respiratórios e genitais do BHV-1 (Roizman 1992). Surtos de meningo-encefalite pelo BHV-5 têm sido freqüentemente descritos, principalmente no Brasil e Argentina (Schudel et al. 1986, Weiblen et al. 1989, Salvador et al. 1998). Os sinais clínicos da enfermidade incluem tremores, andar em círculos, incoordenação, nistagmo, bruxismo, ataxia, convulsões e depressão profunda seguida de morte (Hall et al. 1966, Bagust & Clark 1972, Schudel et al. 1986).
Os herpesvírus bovinos tipo 1 e 5 são muito semelhantes entre si em aspectos estruturais, biológicos, antigênicos e moleculares (French 1962, Metzler et al. 1986, Studdert 1989, Bratanich et al. 1991). A principal diferença entre esses vírus parece estar relacionada à sua habilidade de invadir, replicar no sistema nervoso central (SNC) e causar enfermidade neurológica (Bagust & Clark 1972, Studdert 1989, Belknap et al. 1994). Amostras de alfaherpesvírus identificadas como BHV-1 têm sido quase que exclusivamente isoladas de casos de doença respiratória ou genital, enquanto os herpesvírus isolados de casos de doença nervosa têm sido identificados como BHV-5 (Studdert 1989, d'Offay et al. 1995, Roehe et al. 1997, Salvador et al. 1998). Inoculações experimentais de terneiros têm confirmado o potencial neuropatogênico dos isolados de campo identificados como BHV-5 (Belknap et al. 1994, Cascio et al. 1999).
A falta de um modelo experimental têm dificultados o estudo das bases biológicas e moleculares do fenótipo neuropatogênico do BHV-5. Recentemente, a infecção aguda e enfermidade neurológica pelo BHV-5 foram reproduzidas em ovinos (Silva et al. 1998) e coelhos (Chowdhury et al. 1997, Meyer et al. 1997, Silva et al. 1999). Silva et al. (1999) caracterizaram a enfermidade neurológica aguda em coelhos infectados com o BHV-5 e propuseram essa espécie como modelo experimental para estudos de patogenia. O presente artigo relata a utilização do modelo proposto por Silva et al. (1999) para estudar aspectos específicos da neuropatogênese e resposta imunológica ao BHV-5.
MATERIAL E MÉTODOS
Células e vírus
Os procedimentos de multiplicação, detecção e quantificação de vírus e soro-neutralização (SN) foram realizados com células de linhagem de rim de feto bovino (MDBK). As células foram cultivadas em meio essencial mínimo (MEM), contendo penicilina (1,6mg/l), estreptomicina (0,4mg/l) e suplementado com 10% de soro eqüino (Cultilab). A amostra A663 foi isolada por Carrillo et al. (1983) de casos de meningoencefalite em bezerros na Argentina e posteriormente caracterizada (Metzler et al. 1986, Bratanich et al. 1991). A amostra EVI-88 foi isolada de casos de meningoencefalite em bovinos no Brasil e parcialmente caracterizada (Roehe et al. 1997). A amostra 613 foi isolada de um surto de doença neurológica em bovinos na Argentina e gentilmente cedida pelo Dr. Anselmo Odeon (INTA, Balcarce, Argentina). Os vírus foram amplificados em células MDBK, quantificados e estocados a -70ºC.
Desenho experimental, animais e inoculação
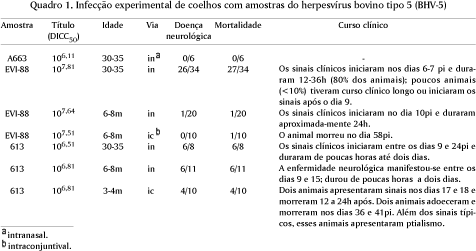
Foram utilizados coelhos da raça Nova Zelândia, independente de sexo, obtidos no Biotério Central e no Colégio Agrícola da Universidade Federal de Santa Maria, Santa Maria, RS. Para a inoculação, foram utilizadas três categorias de animais: coelhos recém-desmamados (30 a 35 dias), com três a quatro meses; e com aproximadamente 6 a 8 meses de idade. Os dados referentes às amostras e títulos virais utilizados e vias de inoculação estão apresentadas no Quadro 1. Os animais foram inoculados com 1ml da suspensão viral pela via intranasal, através de trepanação dos seios paranasais conforme metodologia modificada a partir de técnica descrita por Brown & Field (1990) e descrita com detalhes por Silva et al. (1999). Para a inoculação intranasal, os animais foram previamente anestesiados com Zoletil (Virbac, 100-200ul/animal). Dois grupos de 8 e 10 coelhos, respectivamete, foram inoculados pela via intraconjuntival, através da instilação de 1ml (0,5ml em cada lado) de suspensão viral no saco conjuntival (Quadro 1). Após a inoculação, os animais foram monitorados clinicamente; secções do encéfalo colhidas na necropsia foram submetidas à pesquisa de vírus e exames histológicos.
Cinética e vias de invasão do SNC
Coelhos recém-desmamados (30 a 35 dias de idade) foram inoculados com o BHV-5 (amostra EVI-88, 1ml contendo entre 107,51 e 107,64 DICC50 ) pela via intranasal e sacrificados a intervalos de 24 horas após a inoculação. Suabes nasais e secções de diferentes áreas do SNC foram submetidas à pesquisa de vírus. As secções do encéfalo foram maceradas, homogeneizadas e resuspendidas em MEM (10% peso/volume). Após centrifugação a 2.500 rpm por 10 min, o líquido sobrenadante foi coletado e inoculado em células MDBK. As células foram monitoradas diariamente para o aparecimento de efeito citopático (ECP) e foram consideradas negativas após duas passagens de cinco dias cada sem o aparecimento de ECP.
Dois experimentos foram realizados para investigar a importância da rota olfatória no trânsito do BHV-5 da cavidade nasal até o SNC. No primeiro experimento, coelhos com idade de 3 a 4 meses e 6 a 8 meses foram inoculados com duas amostras do BHV-5, pelas vias intranasal e intraconjuntival, respectivamente (Quadro 1). O curso clínico e os índices de morbidade e mortalidade foram comparados com os índices observados após inoculação intranasal. O encéfalo dos animais que morreram ou foram sacrificados in extremis foram submetidos à pesquisa de vírus. No segundo experimento, doze coelhos foram submetidos à ablação cirúrgica do bulbo olfatório e posteriormente inoculados com o BHV-5 pela via intranasal. Para a cirurgia, os animais foram anestesiados com Fentanil (Citrato de Fentanila, Cristália, São Paulo, SP). A ablação do bulbo olfatório foi realizada através de trepanação seguida de aspiração e curetagem dos seios que abrigam o tecido olfatório. A completa remoção do bulbo olfatório foi confirmada por ocasião da necropsia. Esses animais foram inoculados com o BHV-5 (amostra EVI-88, dose 107,74DICC50/ml) pela via intranasal, juntamente com o grupo controle, cinco dias após a cirurgia. O curso clínico da infecção e os índices de morbidade e mortalidade foram comparados. Secções do SNC dos coelhos que morreram ou foram sacrificados in extremis foram submetidos à pesquisa de vírus.
Efeitos da imunização ativa
Nos experimentos realizados com a finalidade avaliar os efeitos da imunização ativa na infecção e enfermidade neurológica, foram utilizados 28 coelhos com 15 dias de idade. Dez coelhos foram imunizados através da inoculação subcutânea de 1ml de suspensão viral do BHV-5 (EVI-88, 106,81DICC50/ml) e dez foram imunizados com o BHV-1 (amostra Cooper, 1ml contendo 107,64DICC50/ml). Oito coelhos da mesma idade foram inoculados com MEM pela via subcutânea e serviram como controles não-imunizados. Vinte dias após a imunização, os 28 coelhos foram desafiados através de inoculação intranasal do BHV-5 (EVI-88, 1ml com 107,64DICC50/ml) e monitorados clinicamente. Amostras de soro colhidas no dia da imunização e no dia do desafio foram testadas para anticorpos através da técnica de soro-neutralização (SN). Os testes de SN foram realizados em placas de poliestireno de 96 cavidades, utilizando-se diluições 1:2 crescentes de soro (inativado a 56ºC - 30 min) frente a doses constantes de vírus (100 DICC50 /cavidade, amostra EVI-88). A leitura final foi realizada após 96 horas de incubação.
RESULTADOS
Infecção e enfermidade neurológica
Os índices de morbidade e mortalidade, assim como o curso clínico da enfermidade neurológica em coelhos inoculados com o BHV-5 variaram com a amostra viral utilizada, idade dos animais e via de inoculação (Quadro 1). As amostras EVI-88 e 613 foram as que produziram enfermidade com mais freqüência. A amostra EVI-88 produziu índices altos de morbidade em coelhos recém-desmamados, apresentando neuropatogenicidade progressivamente menor quando inoculada em animais com mais idade (Quadro 1). A amostra 613, embora inoculada em títulos virais menores, apresentou alta neuropatogenicidade para coelhos recém-desmamados e também para coelhos adultos. A via conjuntival de inoculação foi menos eficiente na indução da doença nervosa, embora a amostra 613 tenha produzido meningoencefalite em quatro entre dez animais inoculados por essa via. Os sinais clínicos observados foram depressão ou agitação, tremores, bruxismo, reação típica semelhante a opistótono (elevação gradativa dos membros anteriores até estação em posição vertical, com apoio apenas nos membros posteriores), e queda para trás, paralisia nos membros posteriores, depressão profunda e morte.
Cinética e vias de invasão do SNC
A pesquisa de vírus na cavidade nasal e em diferentes secções do SNC de coelhos sacrificados às 24h pós-inoculação detectou replicação viral somente na cavidade nasal. Em coelhos examinados a intervalos maiores após a inoculação, a replicação viral foi inicialmente detectada no bulbo olfatório (48h) e córtex olfatório (48/72h). As 72h o vírus foi detectado também no gânglio trigêmeo e ponte, além do córtex cerebral. No quarto dia após a inoculação, a infecção já encontrava-se distribuida em várias áreas do SNC (Quadro 2). Esses resultados demonstraram que após inoculação intranasal, o vírus atinge inicialmente o bulbo olfatório e sugerem que a invasão do SNC a partir da cavidade nasal ocorre inicialmente pela via olfatória. A detecção de vírus no bulbo olfatório já às 48h pi e a ampla distribuição do vírus no SNC a partir das 72/96h pós-inoculação indicam que a invasão e disseminação do BHV-5 no encéfalo de coelhos é muito rápida e eficiente.
A importância e participação da rota olfatória no acesso do BHV-5 ao encéfalo de coelhos foi investigada através de dois experimentos. No primeiro, coelhos foram inoculados pela via conjuntival com as mesmas amostras do BHV-5 que produziram infecção neurológica após inoculação intranasal. Quatro entre 10 (40%) dos coelhos inoculados com a amostra 613 e 1/10 (10%) dos coelhos inoculados com a amostra EVI-88 apresentaram sinais neurológicos e morreram. Além da morbidade menor, o curso clínico da doença neurológica nesses animais foi mais tardio do que nos animais inoculados pela via intranasal (Quadro 1). No segundo experimento, doze coelhos foram submetidos à ablação cirúrgica do bulbo olfatório e posteriormente inoculados com o BHV-5. Doze coelhos não submetidos à cirurgia foram utilizados como controle. Onze dos 12 animais (91,6%) controles desenvolveram a enfermidade, contra quatro de 12 (33,3%) dos coelhos submetidos à ablação do bulbo olfatório (Quadro 3). Os resultados desses dois experimentos indicam que o sistema olfatório não constitui-se na única via de acesso do BHV-5 ao SNC de coelhos. A interrupção cirúrgica dessa via reduziu drasticamente, mas não preveniu a invasão do SNC pelo BHV-5 a partir da cavidade nasal. Da mesma forma, a inoculação conjuntival, que provavelmente resulta em invasão do SNC por outra via nervosa, também pode resultar em invasão e infecção do SNC, porém com menor eficiência e mais tardiamente.
Proteção por imunização ativa
Para investigar se a imunização ativa com o vírus homólogo (BHV-5) e com um vírus heterólogo (BHV-1) protegeria coelhos da infecção e enfermidade neurológica, coelhos foram imunizados com o BHV-5 (EVI-88) ou com o BHV-1 (Cooper) e 20 dias após foram desafiados com o BHV-5. No dia do desafio, os coelhos imunizados com o BHV-1 apresentaram títulos de anticorpos neutralizantes superiores aos animais imunizados com o BHV-5 (Quadro 4). Os coelhos imunizados com o BHV-1 ficaram protegidos da doença clínica (apenas dois animais apresentaram sinais clínicos neurológicos passageiros) e da mortalidade. Coelhos imunizados com o BHV-5 também ficaram protegidos, porém com menor eficiência. Apenas um animal desenvolveu doença neurológica fatal e outros cinco apresentaram sinais clínicos passageiros e recuperaram-se (Quadro 4). Esses resultados indicam que além da proteção por imunização com o vírus homólogo, proteção contra a infecção e enfermidade neurológica pelo BHV-5 em coelhos pode ser obtida por imunização com o BHV-1.
DISCUSSÃO
Os resultados desse trabalho demonstram que a infecção e enfermidade neurológica pelo BHV-5 em coelhos, caracterizada por Silva et al. (1999), pode ser utilizada para estudar-se vários aspectos da infecção neurológica pelo BHV-5. Ressalvadas as diferenças que certamente existem nas relações entre o BHV-5 com bovinos e coelhos, a infecção neurológica nessas duas espécies apresenta características similares. Variações marcantes de neuropatogenicidade entre as amostras de vírus, assim como diferentes graus de susceptibilidade associados com a idade dos animais puderam ser observados na infecção pelo BHV-5 em coelhos, a exemplo do que tem sido demonstrado em bovinos (Belknap et al. 1994, Cascio et al. 1999). Estudos anteriores em bovinos demonstraram que inoculação de bezerros recém-nascidos (um a três dias de idade) com o BHV-5 produziu infecção cerebral fulminante acompanhada de sinais neurológicos e alterações inflamatórias severas que culminaram com a morte dos animais (Belknap et al. 1994). Em contraste, bezerros com cinco a seis meses de idade inoculados com a mesma amostra de vírus não desenvolveram enfermidade neurológica (Cascio et al. 1999). A enfermidade em coelhos, apesar do curso rápido, dos sinais neurológicos típicos e presença de vírus em diversas àreas do encéfalo, geralmente não é acompanhada de alterações inflamatórias severas, como observado em bovinos (Silva et al. 1999). No entanto, doença neurológica aguda fulminante, sem alterações inflamatórias severas também já foram observadas em bovinos inoculados com o BHV-5 (Beltrão 2000).
A reprodução de enfermidade neurológica pelo BHV-5 em coelhos foi recentemente relatada por vários grupos (Meyer et al. 1997, Chowdhury et al. 1997, Silva et al. 1999). A técnica de inoculação intranasal através de incisão cutânea e trepanação dos seios paranasais (Brown & Field 1990), posteriormente modificada por Silva et al (1999), permite a inoculação de volumes consideráveis de suspensão viral (>1ml por animal), com o mínimo de trauma, resultando em boa distribuição do inóculo na cavidade nasal. A coleta de suabes nasais para o monitoramento da replicação viral, no entanto, segue representando um problema devido à pequena abertura e grande sensibilidade das narinas. Alguns desses estudos relataram a necessidade de tratamento prévio dos animais com dexametasona para a produção da enfermidade (Chowdhury et al. 1997), o que não foi necessário nos experimentos descritos no presente estudo e nos relatos de Silva et al. (1999).
Os estudos de patogenia da infecção pelo BHV-5 sugerem o sistema olfatório como a principal via de acesso do BHV-5 ao encéfalo de coelhos após inoculação intranasal (Chowdhury et al. 1997,1999, Silva et al. 1999). O estudo da cinética da invasão do SNC (Quadro 2) revelou que o vírus atinge inicialmentwe o bulbo olfatório e córtex olfatório já às 48 horas pós-inoculação; e às 72hpi já pode ser detectado no tronco cerebral e córtex cerebral. A ampla disseminação do vírus no SNC após as 72hpi dificultou investigações mais detalhadas sobre outras vias de invasão e de disseminação. Os resultados do experimento com coelhos submetidos à ablação do bulbo olfatório reforçaram a hipótese de que o acesso do BHV-5 ao encéfalo de coelhos após inoculação intranasal ocorre principalmente pelo sistema olfatório. No entanto, esse experimento também demonstrou que o sistema olfatório não é a única via utilizada pelo BHV-5 para invadir o SNC. Recentemente, foi demonstrado que o BHV-5 estabelece infecção latente no gânglio trigêmeo de coelhos após inoculação intranasal e que a reativação da infecção latente é freqüentemente seguida de recrudescência da doença neurológica (Caron et al. 2000). Esses resultados, aliados à ocorrência de meningo-encefalite fatal em coelhos inoculados pela via intra-conjuntival (Quadro 1) indicam que as vias nervosas que compõe o nervo trigêmeo também podem ser utilizadas pelo BHV-5 para invadir o cérebro de coelhos. O curso tardio da enfermidade nesses animais, no entanto, sugere que a provável invasão por essa via é menos eficiente e mais tardia quando comparada com a via olfatória.
Os objetivos dos experimentos de proteção por imunização ativa foram avaliar a adequação desse modelo animal para estudos imunológicos, sem a pretensão de responder conclusivamente as questões pertinentes. Portanto, os resultados desses experimentos devem ser considerados preliminares e interpretados com cautela, enquanto não forem confirmados por estudos adicionais nos hospedeiros naturais. Não obstante, foi demonstrado que o modelo experimental proposto por Silva et al. (1999) parece ser apropriado para estudos de diversos aspectos da resposta imunológica ao BHV-5. Testes preliminares de proteção vacinal, formulações de adjuvantes e vacinas, e regimes de imunização poderiam potencialmente ser realizados em coelhos. Anteriormente, proteção parcial de coelhos através de imunização (imunização das coelhas-mães com o BHV-5) já havia sido demonstrada (Silva et al. 1999).
As diferenças antigênicas entre o BHV-5 e BHV-1 podem ser evidenciadas com o uso de anticorpos monoclonais contra as glicoproteínas do envelope viral (Metzler et al. 1986, Friedli et al. 1987, Roehe et al. 1997). No entanto, testes sorológicos de rotina muitas vezes são incapazes de distinguir entre esses dois vírus. Proteção cruzada entre o BHV-1 e BHV-5, através de imunização ativa e passiva, já foi demonstrada experimentalmente em bovinos (Belknap et al. 1994, Cascio et al. 1999). Essa reatividade sorológica cruzada tem sido responsabilizada, em parte, pela baixa prevalência do BHV-5 e pela ocorrência rara da enfermidade neurológica em países que vacinam regularmente contra o BHV-1 (Studdert 1989, d'Offay et al. 1995). Os resultados obtidos com o experimento de imunização ativa em coelhos, demonstram que a reatividade sorológica cruzada entre o BHV-1 e BHV-5 pode traduzir-se em proteção cruzada. Os maiores índices de proteção observados entre coelhos imunizados com o BHV-1 podem ser atribuídos à maior dose de vírus utilizada para imunização (quase 1log 10 maior), como também a uma possível influência de diferenças antigênicas do BHV-1, levando a uma resposta mais eficaz em termos de proteção.
Em resumo, os resultados obtidos nesse trabalho demonstram a utilidade de coelhos para a investigação de vários aspectos da infecção neurológica pelo BHV-5. Ressalvadas as diferenças existentes entre coelhos e os hospedeiros naturais do BHV-5, as semelhanças observadas na infecção dessas espécies justificam o uso de coelhos como modelo experimental. Certamente, a disponibilidade de um modelo experimental adequado auxiliará no esclarecimento dos mecanismos biológicos e moleculares responsáveis pelo potencial neuropatogênico desse vírus.
Agradecimentos.- Ao Médico Veterinário Antoninho Macuglia (Biotério Central) e à Professora Ione Denardin (Colégio Agrícola da UFSM) pelo fornecimento dos coelhos. Aos colegas Dr Paulo M.Roehe (CPVDF - Centro de Pesquisas Veterinárias Desidério Finamor), Dr Anselmo Odeon (INTA Balcarce, Argentina) e Dra Elba Laura Weber (INTA Castelar, Argentina) pelas amostras de BHV-5. Aos bolsistas e funcionários do Laboratório de Virologia da UFSM pelos cuidados e manutenção dos coelhos. Ao Dr H-Jung Sur (Plum Island Animal Disease Center, Greenport, NY) pela realização dos testes de hibridização in situ.
Trabalho realizado com suporte do MCT-CNPq, Capes e Finep (PRONEX em Virologia Veterinária ¾ 215/96).
- Bagust T. J. & Clark L.1972. Pathogenesis of meningoencephalitis produced in calves by infectious bovine rhinotracheitis herpesvirus. J. Comp. Path. 82:375-383.
- Belknap E.B., Collins J.K., Ayers V.K. & Schultheiss P.C.1994. Experimental infection of neonatal calves with neurovirulent bovine herpesvirus type 5(BHV-5). Vet. Pathol. 31:358-365.
- Beltrăo, N. 2000. Herpesvírus bovino tipo 5 (BHV-5): infecçăo experimental de bovinos e utilizaçăo de coelhos como modelo. Dissertaçăo de Mestrado, Universidade Federal de Santa Maria, Santa Maria, RS. 54p.
- Bratanich A.C., Sardi A.I., Smitstaart E.N. & Schudel A.A.1991. Comparative studies of BHV-1 variants by in vivo and in vitro tests. Zbl.Vet.Med.B. 38:41-48.
- Brown G.A. & Field H.J.1990. Experimental reactivation of bovine herpesvirus (BHV-1) by means of corticosteroids in na intranasal rabbit model. Arch. Virol. 112:81-101.
- Caron L., Flores E.F., Weiblen R., Scherer C.F.C, Irigoyen L.F. & Sue J-H. 1999. Latent infection by bovine herpesvirus type (BHV-5): spontaneous and dexamethasone-induced viral reactivation and recrudescence of nerological disease. In: 19th Annual Meeting, American Society for Virology, Fort Collins, USA.
- Cascio K.E., Belknap E.B., Schulthesis P.C., Ames ŞD. & Collins J.K. 1999. Encephalitis induced by bovine herpesvirus 5 and protection prior vaccination on infection with bovine herpesvirus 1. J. Vet. Diagn. Invest. 11:134-139.
- Carrillo, B.J., Ambrogi, A., Schudel A.A., Vazques M., Dahme, E. & Pospischil, A. 1983. Meningoencephalitis caused by IBR virus in calves in Argentina. Zbl. Vet. Med. B. 30:327-332.
- Chowdhury S.I., Lee B.J., Mosier B.J., Sur J-H., Osorio F.A., Kennedt G. & Weiss M.L. 1997. Neuropathology of bovine herpesvirus type 5 (BHV-5) meningoencephalitis in a rabbit seizure model. J. Comp. Pathol. 117:295-310.
- French E. L. 1962. A specific virus encephalitis in calves: isolation and characterization of the causal agent. Aust.Vet.J. 38:216-221.
- Friedli K. & Metzler A.E.1987. Reactivity of monoclonal antibodies to proteins of a neurotropic bovine herpesvirus 1 (BHV-1) strain and to proteins of representative BHV-1 strains. Arch. Virol. 94:109-122.
- Hall W.T.K., Simmons G.C., French E.L., Snowdon W.A. & Asdell M. 1966. The pathogenesis of meningoencephalitis caused by the infectious bovine rhinotracheitis virus. Aust.Vet.J. 42:229-237.
- Lee, B.J., Weiss, M.L., Mosier, B.J. & Chowdhury S.I. 1999. Spread of bovine herpesvirus type 5 (BHV-5) in the rabbit brain after intranasal inoculation. J.Neurovirol. 5:474-484.
- Metzler A.E., Schudel A.A. & Engels M.1986. Bovine herpesvirus 1: molecular and antigenic characteristics of variant viruses isolated from calves with neurological disease. Arch.Virol. 87:205-217.
- Meyer G., Lemaira M. & Lyaku J.1996. Establishment of a rabbit model for bovine herpesvirus type 5 neurological acute infection. Vet.Microbiol. 51:27-40.
- d'Offay J.M., Ely R.W., Baldwin C.A., Whitenack D.L., Stair E.L. & Collins J.K. 1995. Diagnosis of encephalitic bovine herpesvirus type 5 (BHV-5) infection in cattle: virus isolation and immunohistochemical detection of antigen in formalin-fixed brain tissues. J.Vet.Diagn.Invest. 7:247-251.
- Reed L. & Muench, H. 1938. A simple method of estimating fifty percent endpoints. Am.J.Hyg. 18: 493-494.
- Rock D.L., Lokensgard T., Lewis T & Kutish G.1992. Characterization of dexamethasone-induced reactivation of latent bovine herpesvirus 1. J.Virol. 66:2484-2490.
- Roehe P.M., Silva T.C., Nardi N.B., Oliveira D.G. & Rosa J.C.A 1997. Diferenciaçăo entre o vírus da rinotraqueíte infecciosa bovina (BHV-1) e o herpesvírus da encefalite bovina (BHV-5). Pesq. Vet. Bras. 17:41-44.
- Roizman B. 1992. The family Herpesviridae: an update. Arch.Virol. 123:432-445.
- Schudel A.A., Carrillo B.J., Wyler R. & Metzler A.E. 1986. Infection of calves with antigenic variants of bovine herpesvirus 1 (BHV - 1) and neurological disease. Zbl. Vet. Med. B. 33:303-310.
- Silva A.M., Flores E.F., Weiblen R., Botton S.A., Irigoyen L.F., Roehe P.M., Brum M.C. & Canto M.C.1998. Infecçăo aguda e latente em ovinos inoculados com o herpesvirus bovino tipo 5 (BHV-5). Pesq. Vet. Bras. 18:99-106.
- Silva A.M., Flores E.F., Weiblen R., Canto M.C., Irigoyen L.F., Roehe P.M. & Sousa R.S. 1999. Pathogenesis of meningoencephalitis in rabbits by bovine herpesvirus type-5 (BHV-5). Revta Microbiol., Săo Paulo, 30:22-31.
- Salvador S.C., Lemos R.A.A., Riet-Correa F., Roehe P.M. & Osorio A.L.A.R. 1998. Meningoencephalitis in cattle caused by bovine herpesvírus-5 in Mato Grosso do Sul and Săo Paulo, Brazil. Simpósio Int.BHV-1/5 e BVDV, Santa Maria, RS, 22-24 abril 1998, p.149.
- Studdert, M. J. 1989. Bovine encephalitis herpesvirus. Vet.Rec. 125:584.
- Weiblen R., Barros C.S.L., Canabarro T.F. & Flores I.E. 1989. Bovine meningoencephalitis from IBR virus. Vet. Rec. 124:666-667.
Datas de Publicação
-
Publicação nesta coleção
24 Jan 2001 -
Data do Fascículo
Dez 2000
Histórico
-
Aceito
14 Jun 2000