Resumos
Schizomeris Kützing é uma alga filamentosa sem ramificações e com sistema basal de fixação diferenciado. O gênero contém a espécie-tipo Schizomeris leibleinii Kützing e duas espécies pouco conhecidas, S. irregularis Fritsch & Rich e S. indicum (Ghose) Fritsch & Rich. As espécies são diferenciadas, principalmente, através do tipo de sistema basal de fixação. Contudo, a taxonomia específica do gênero é confusa. Estudos morfométricos e morfológicos foram realizados em populações identificadas como S. leibleinii de acordo com duas abordagens. Na primeira, foram analisadas 11 populações de diferentes localidades e na segunda, duas populações distintas através de coletas mensais durante 11 meses. O diâmetro de filamentos multisseriados, diâmetro e comprimento de células de filamentos multi e unisseriados, presença de constrição, ápice dos filamentos e tipo de sistema basal foram analisados em ambas abordagens. Além disso, estudos cariológicos foram efetuados para complementar as informações taxonômicas. Os resultados revelaram ampla variação morfológica, sendo mais acentuada entre as populações de localidades diferentes que nas mesmas populações ao longo do tempo, mas as características mostraram um gradiente contínuo. Os diferentes tipos de sistemas basais descritos para as espécies do gênero (claviforme, apressório, lobado e rizóides) foram encontrados nas populações estudadas. Esses resultados sugerem que a distinção entre as três espécies é muito frágil e não pode ser apoiada pela diferenciação do sistema basal ou por características métricas. Acredita-se que, por essas razões, o gênero Schizomeris seja composto por uma única espécie. A análise cariológica resultou em n = 12, diferindo de estudos anteriores e algumas possíveis explicações são apresentadas.
cariologia; Chaetophorales; Schizomeris leibleinii; taxonomia; variação morfológica
Schizomeris Kützing is a filamentous alga without ramifications and with differentiated basal attachment system. The genus contains the type species Schizomeris leibleinii Kützing and the two poorly known species S. irregularis Fritsch & Rich and S. indicum (Ghose) Fritsch & Rich. The species are differentiated mostly by the basal attachment system type. However, the specific taxonomy of the genus is confused. Morphometrical and morphological studies were conducted in populations identified as S. leibleinii according to two approaches. In the first one, 11 populations from different localities were analyzed and in the second, two distinct populations were monthly evaluated during 11 months. Diameter of multiseriate filaments, diameter and length of cells from multi and uniseriate filaments, presence of constriction, form of filament summit, and basal system type were evaluated in both approaches. In addition, karyological studies were conducted to complement taxonomic information. Results revealed a wide morphological variation, being more accentuated among populations from different localities than in the same populations along the time, but the characteristics showed a continuous gradient. The different types of the basal system described for the species of the genus (bluntly rounded, holdfast disc, lobed and rhizoid) were found in the populations studied. These findings suggest that distinction among the three species is very fragile and cannot be supported by differentiation of the basal system or metrical characteristics. Based on present results we consider that Schizomeris is composed by a single species. Karyological analyses resulted in n = 12, differing from previous studies and some possible explanations are discussed.
Chaetophorales; karyology; morphological variation; Schizomeris leibleinii; taxonomy
Aspectos taxonômicos de Schizomeris Kützing (Chaetophorales, Chlorophyta)
Taxonomic aspects of Schizomeris Kützing (Chaetophorales, Chlorophyta)
Jeferson Luizi Pereira1 1 . Autor para correspondência: jluizi@ig.com.br ; Luis Henrique Z. Branco
Universidade Estadual Paulista (UNESP), Departamento de Zoologia e Botânica, Rua Cristóvão Colombo, 2265, 15054000 São José do Rio Preto, SP, Brasil
RESUMO
Schizomeris Kützing é uma alga filamentosa sem ramificações e com sistema basal de fixação diferenciado. O gênero contém a espécie-tipo Schizomeris leibleinii Kützing e duas espécies pouco conhecidas, S. irregularis Fritsch & Rich e S. indicum (Ghose) Fritsch & Rich. As espécies são diferenciadas, principalmente, através do tipo de sistema basal de fixação. Contudo, a taxonomia específica do gênero é confusa. Estudos morfométricos e morfológicos foram realizados em populações identificadas como S. leibleinii de acordo com duas abordagens. Na primeira, foram analisadas 11 populações de diferentes localidades e na segunda, duas populações distintas através de coletas mensais durante 11 meses. O diâmetro de filamentos multisseriados, diâmetro e comprimento de células de filamentos multi e unisseriados, presença de constrição, ápice dos filamentos e tipo de sistema basal foram analisados em ambas abordagens. Além disso, estudos cariológicos foram efetuados para complementar as informações taxonômicas. Os resultados revelaram ampla variação morfológica, sendo mais acentuada entre as populações de localidades diferentes que nas mesmas populações ao longo do tempo, mas as características mostraram um gradiente contínuo. Os diferentes tipos de sistemas basais descritos para as espécies do gênero (claviforme, apressório, lobado e rizóides) foram encontrados nas populações estudadas. Esses resultados sugerem que a distinção entre as três espécies é muito frágil e não pode ser apoiada pela diferenciação do sistema basal ou por características métricas. Acredita-se que, por essas razões, o gênero Schizomeris seja composto por uma única espécie. A análise cariológica resultou em n = 12, diferindo de estudos anteriores e algumas possíveis explicações são apresentadas.
Palavras-chave: cariologia, Chaetophorales, Schizomeris leibleinii, taxonomia, variação morfológica
ABSTRACT
Schizomeris Kützing is a filamentous alga without ramifications and with differentiated basal attachment system. The genus contains the type species Schizomeris leibleinii Kützing and the two poorly known species S. irregularis Fritsch & Rich and S. indicum (Ghose) Fritsch & Rich. The species are differentiated mostly by the basal attachment system type. However, the specific taxonomy of the genus is confused. Morphometrical and morphological studies were conducted in populations identified as S. leibleinii according to two approaches. In the first one, 11 populations from different localities were analyzed and in the second, two distinct populations were monthly evaluated during 11 months. Diameter of multiseriate filaments, diameter and length of cells from multi and uniseriate filaments, presence of constriction, form of filament summit, and basal system type were evaluated in both approaches. In addition, karyological studies were conducted to complement taxonomic information. Results revealed a wide morphological variation, being more accentuated among populations from different localities than in the same populations along the time, but the characteristics showed a continuous gradient. The different types of the basal system described for the species of the genus (bluntly rounded, holdfast disc, lobed and rhizoid) were found in the populations studied. These findings suggest that distinction among the three species is very fragile and cannot be supported by differentiation of the basal system or metrical characteristics. Based on present results we consider that Schizomeris is composed by a single species. Karyological analyses resulted in n = 12, differing from previous studies and some possible explanations are discussed.
Key words: Chaetophorales, karyology, morphological variation, Schizomeris leibleinii, taxonomy
Introdução
O gênero de alga verde Schizomeris foi proposto por Kützing (1843a), validado pelo autor com a publicação da descrição de Schizomeris leibleinii Kützing (Kützing 1843b) e incluído na família Hormidieae (Kützing 1845). A validade do gênero foi colocada em dúvida por alguns ficólogos que o consideraram, meramente, um estágio de desenvolvimento de Ulothrix Kützing (Cienkowski 1876, Printz 1927, Fritsch 1935), incluindo-o na família Ulotrichaceae como um gênero duvidoso. Fott (1959) e Bourrelly (1966), no entanto, consideraram-no como membro das Ulvaceae (Ulvales). Tanto Prescott (1962, 1968) como Papenfuss (1955) classificaram esse gênero em uma família própria, Schizomeridaceae, pertencente à ordem Ulotrichales. Atualmente, com base em análises citológicas feitas por Birkbeck et al. (1974) e Mattox et al. (1974), Schizomeris é considerado um gênero bem delimitado e classificado na família Schizomeridaceae e ordem Chaetophorales.
Em 1924, Fritsch & Rich descreveram uma segunda espécie, Schizomeris irregularis Fritsch & Rich, e também concluíram que Uronema indicum Ghose seria melhor designada como Schizomeris indicum (Ghose) Fritsch & Rich, sendo esta a terceira espécie. De todas as espécies citadas, apenas S. leibleinii é mencionada e descrita com freqüência na literatura mundial, enquanto S. irregularis e S. indicum foram citadas, respectivamente, apenas para África do Sul (Fritsch & Rich 1924) e Índia (Ghose 1920).
Segundo a descrição original, Schizomeris leibleinii é uma alga filamentosa, desprovida de ramificações e dotada de sistema basal de fixação do tipo rizoidal coberto por mucilagem. Os filamentos adultos geralmente mostram-se unisseriados próximo à base (região unisseriada) com as células achatadas ou cilíndricas. Os filamentos são multisseriados na região distal (região multisseriada), espessos, com células pequenas dispostas desordenadamente (Kützing 1843b).
Segundo Fritsch & Rich (1924), S. irregularis distingue-se das outras duas espécies pela disposição irregular das células na região multisseriada do filamento, mas Prasad & Srivastava (1963) constataram que esta característica depende do estágio de desenvolvimento do filamento. De acordo com Watson & Tilden (1930), S. indicum diferencia-se das demais espécies pelo ápice acuminado dos filamentos. Contudo, esses mesmos autores coletaram espécimes que apresentavam todas as características de S. leibleinii, mas, em muitos casos, os filamentos manifestavam o ápice acuminado.
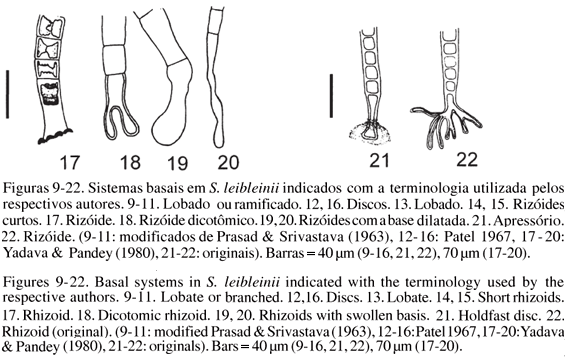
De acordo com Prasad & Srivastava (1963), a característica mais confiável para se distinguir as três espécies seria a diferenciação do sistema basal de fixação dessas algas. Segundo esses autores, S. leibleinii tem a região basal alongada e sub-rizoidal, podendo ser também rizoidal, lobada ou ramificada. Schizomeris indicum apresenta o sistema basal como um apressório mucilaginoso (disco de fixação), enquanto S. irregularis possui sistema de fixação do tipo claviforme.
Várias análises cariológicas foram efetuadas em Schizomeris leibleinii e, no entanto, os dados disponíveis resultam em mais dúvidas ainda com relação ao cariótipo da espécie. Prasad & Srivastava (1963) analisaram amostras dessa alga, encontrando n = 28 cromossomos. Em um estudo citológico dessa mesma espécie, Chowdary (1967) quantificou como n = 15 o número de cromossomos. Patel (1967) analisou amostras que apresentavam 30 cromossomos, sugerindo que esta diferença na contagem em relação aos autores anteriores poderia ser devido à manifestação de poliploidia, raças citológicas ou por alternância de gerações nesta espécie. No entanto, Sarma & Chaudhary (1975) determinaram um n = 14 cromossomos para S. leibleinii, o que diferiu consideravelmente dos cariótipos encontrados até então, sugerindo, assim, que a população estudada seria uma nova raça citológica de S. leibleinii.
Tendo em vista as incertezas em relação a essa espécie e às demais pertencentes ao gênero Schizomeris, o objetivo deste trabalho é avaliar taxonomicamente a espécie S. leibleinii através de dados morfométricos e cariológicos, com a finalidade de delimitar características confiáveis a serem usadas na taxonomia do gênero em questão.
Material e métodos
O estudo da variabilidade morfológica de espécimes de Schizomeris leibleinii Kützing seguiu duas linhas principais: avaliação de populações de diferentes localidades (espacial) e populações de mesma localidade amostradas mensalmente (temporal). A análise da variação morfométrica em populações distintas de S. leibleinii foi realizada a partir de amostras coletadas em 10 localidades diferentes e uma proveniente de material crescendo em laboratório. A maioria das amostras encontrava-se preservada em solução de formaldeído 4% e são originárias de: 1) Herbário do Ibilce/Unesp de São José do Rio Preto/SP - SJRP [abreviação de herbário de acordo com Holmgren et al. (1990)] denominadas Rio Preto (SJRP23461), Canela (SJRP23479), Borá (SJRP23483), Dourados (SJRP27369), São Pedro (SJRP23535); 2) Museu Nacional, Depto. de Botânica/RJ - Fazenda Favorita (R155583); 3) novas coletas feitas nos meses de abril/2001, maio/2001, novembro/2001 e outubro/2002, respectivamente no Ribeirão Claro (Guapiaçu/SP), Rio Verde (Campina Verde/MG), Córrego Piedade (São José do Rio Preto/SP) e Córrego da Maquininha (São José do Rio Preto/SP); e 4) cultura crescendo em Meio Básico de Bold (MBB) por 120 dias (amostra derivada de coleta efetuada no Ribeirão Claro - Guapiaçu/SP).
Para o estudo da variação morfométrica mensal (janeiro/1998, abril/1998-janeiro/1999) foram analisados espécimes coletados em dois trechos do Ribeirão Claro, denominados P3 (20º46'33''S e 49º11'49''W) e P4 (20º46'15'' S e 49º10'39'' W), ambos recebendo descargas de esgotos domésticos e resíduos de dois abatedouros de animais procedentes da cidade de Guapiaçu/SP (Branco & Pereira 2002). Dados referentes às característica físicas e químicas do ambiente coletados mensalmente (Branco & Pereira 2002) também foram utilizados, para avaliar as possíveis fontes de variação dos caracteres morfométricos e/ou morfológicos.
Foram analisados, sob microscopia de luz, 20 filamentos algais completos, ou seja, que apresentavam a região unisseriada e multisseriada, quanto às seguintes características: morfométricas - (i) diâmetro do filamento (região multisseriada), (ii) diâmetro e comprimento celular (região unisseriada e multisseriada) e morfológicas - (iii) tipo de sistema basal de fixação (presença e/ou ausência dos tipos rizóide, apressório, claviforme e lobado). Para as amostras avaliadas mensalmente (P3 e P4) efetuou-se o mesmo procedimento, sendo que, para cada amostra mensal (um total de 11 para cada população), 20 medidas/avaliações foram tomadas de cada característica nas amostras do P3 e P4.
Além dessas características, a forma e/ou posição dos cloroplastos nas células, disposição das células na região multisseriada, forma do ápice e presença de septos transversais ("anéis") foram analisadas nesses mesmos filamentos, para comparação com dados da literatura especializada sobre a morfometria e morfologia de S. leibleinii e de outras espécies do gênero Schizomeris.
O material proveniente do Ribeirão Claro (população denominada Cultura) foi destinado, parcialmente, ao crescimento em meio de cultura unialgal (não axênica) para avaliação taxonômica. O material mantido vivo foi lavado várias vezes com água deionizada esterilizada e, em seguida, três ou quatro filamentos de S. leibleinii foram transferidos para erlenmeyers com meio de cultura MBB (Nichols 1973). Os frascos com os inóculos foram mantidos em câmaras de cultivo sob intensidade luminosa de 100 µmol.m-2.s-1 (± 2,0 µmol.m-2.s -1) com fotoperíodo de 12 h claro/12 h escuro (modificado de Sarma & Chaudhary 1975) e temperatura de 20 ºC (± 1 ºC).
Para a análise cariológica, testes prévios foram realizados para determinar uma maior quantidade de divisões celulares nos filamentos de acordo com Kapraun & Martin (1987). Assim, os filamentos provenientes de espécimes coletados no Ribeirão Claro, no período compreendido entre as 16:30 h e 17:10 h e fixados no próprio local com solução de Carnoy, mostraram-se satisfatórios. Os cromossomos foram analisados a partir de aproximadamente 30 células em divisão mitótica que se encontravam em metafase, já que esta é a melhor etapa para se visualizar e quantificar os cromossomos (Prasad & Srivastava 1963, Patel 1967, Sarma & Chaudhary 1975). A preparação para a contagem de cromossomos foi efetuada segundo a técnica descrita por Wittmann (1965) modificada por meio da diluição da solução de trabalho (corante) a 10% em ácido acético glacial 45%, em virtude da intensa coloração observada no citoplasma das células, o que dificultava a visualização dos cromossomos. Em todas as observações e registros foi utilizado um microscópio fotônico binocular Jenaval equipado com câmara clara e sistema fotográfico. Os cromossomos foram fotografados com filme Plus-X-Pan ASA 100 e ampliados a partir de objetiva de ampliação fotográfica.
Utilizando-se os programas estatísticos Minitab versão 10.1 e KCS (Kovach Computing Services) Multivarite Statistical Package (MVSP versão 3.1), os dados de variação morfométrica foram submetidos a análises descritivas (média, desvio padrão e erro padrão), análise de variância (Anova - uma via), teste t de Student e análise de grupamento, para verificação das diferenças entre as características morfológicas das diferentes populações naturais ou em cultivo e sua relação com a taxonomia da espécie. Os dados das variações morfométricas e morfológicas mensais foram, ainda, submetidos à análise de correlação (r de Pearson) com as características ambientais avaliadas.
Resultados
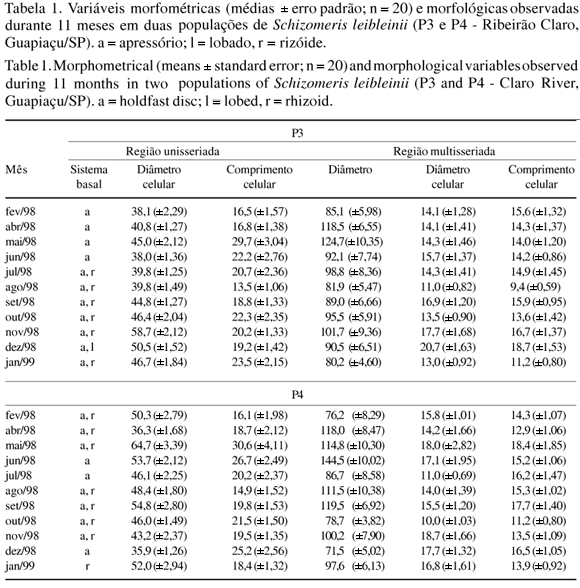
Variação morfométrica intrapopulacional - As duas populações de S. leibleinii Kützing (figuras 1-5) avaliadas temporalmente exibiram ampla variação morfométrica (tabela 1) e ocorreram sob diversas condições ambientais (tabela 2). A Anova das características morfométricas da população P3 mostrou que, na região unisseriada, houve diferenças significativas quanto ao diâmetro celular (F = 6,53; p < 0,001) e ao comprimento celular (F = 4,60; p < 0,001), enquanto que na região multisseriada revelou diferenças significativas na região quanto ao diâmetro do filamento (F = 3,88; p < 0,001), diâmetro celular (F = 4,05; p < 0,001) e comprimento celular (F = 4,37; p < 0,001) nas amostras avaliadas. A análise de grupamento das diferentes amostras (figura 6), baseada na variação das características morfométricas e morfológicas, revelou a formação de cinco grupos. O grupo D incluiu as amostras nov/98, set/98, out/98 e jul/98, apresentando 83,7% de similaridade entre as amostras; o grupo E compreendeu as amostras mai/98, abr/98, jun/98 e fev/98 com 81% de similaridade. As amostras coletadas em dez/98 (grupo A), jan/99 (grupo B) e ago/98 (grupo C) não se agruparam com qualquer das amostras citadas, constituindo grupos distintos. As variações sazonais das características ambientais selecionadas para a população P3 (tabela 2) revelaram correlações significativas principalmente entre o diâmetro das células da região unisseriada e condutividade (r = 0,577; p < 0,05), irradiância (r = 0,685; p < 0,05) e NO3- (r = -0,705; p < 0,05).

A Anova das características morfométricas da população P4 mostrou que na região unisseriada ocorreram diferenças significativas quanto ao diâmetro celular (F = 6,06; p < 0,001) e comprimento celular (F = 4,46; p < 0,001), enquanto na região multisseriada entre as amostras comparadas mostrou que nesta região houve diferenças significativas quanto ao diâmetro do filamento (F = 7,70; p < 0,001), diâmetro celular (F = 3,21; p < 0,001) e comprimento celular (F = 3,19; p < 0,001). A análise de grupamento das diferentes amostras de P4 (figura 7), baseada na variação das características morfométricas e morfológicas, revelou a composição de três grupos. O grupo A, formado pelas amostras coletadas em dez/98, jul/98, e jun/98, apresentou similaridade de 76,2% entre as amostras e o grupo C, compreendendo as amostras jan/99, out/98, nov/98, abr/98, set/98, ago/98 e fev/98, com similaridade de 76,5%. A amostra coletada em maio/1998 (grupo B) não se agrupou com qualquer das amostras anteriormente citadas, constituindo um grupo distinto. A dinâmica sazonal das características ambientais selecionadas para essa população (tabela 2) apresentou correlações significativas principalmente entre o diâmetro dos filamentos na região multisseriada do filamento com irradiância (r = 0,676; p < 0,05), temperatura (r = -0,627; p < 0,05), turbidez (r = -0,773; p < 0,01) e velocidade (r = -0,752; p < 0,01), enquanto na região unisseriada apenas o diâmetro celular correlacionou-se com condutividade (r = 0,601; p < 0,05).
Submetendo-se os dados médios dos caracteres morfométricos, tanto da população P3 quanto da P4, ao teste t de Student com a finalidade de comparar as duas populações observou-se que os valores da região unisseriada: diâmetro celular (t = 1,23) e comprimento celular (t = 0,37); e da região multisseriada: diâmetro do filamento (t = 0,69), diâmetro celular (t = 0,27) e comprimento celular (t = 0,58) não foram significativamente diferentes entre as duas populações.
Variação morfométrica interpopulacional - As 11 populações de S. leibleinii, avaliadas espacialmente, exibiram ampla variação morfométrica (tabela 3). A ocorrência conjunta dos quatro tipos de sistemas basais (apressório, claviforme, lobado e rizóides - figuras 9-22) foi observada apenas na população denominada Cultura. A Anova mostrou que o diâmetro celular da região unisseriada nas populações foi diferente (F = 31,80; p < 0,001). Diferença significativa também foi encontrada quando a mesma análise foi feita para comparar o comprimento celular entre as populações (F = 3,21; p < 0,001). A Anova mostrou, em relação à região multisseriada, que diferenças significativas ocorreram quanto ao diâmetro celular (F = 7,48; p < 0,001), comprimento celular (F = 7,07; p < 0,001) e diâmetro dos filamentos (F = 24,07; p < 0,001) quando as diferentes populações foram comparadas. A análise de grupamento das populações (figura 8) baseada na variação das características morfométricas e morfológicas (tabela 3) revelou a formação de seis grupos. O grupo F incluiu as populações Dourados, Favorita e R. Claro, com 80,9% de similaridade entre si. O grupo E compreendeu as populações do Canela, Borá, S. Pedro e R. Preto, com 77,3% de similaridade entre estas populações. As populações Cultura, Piedade, Maquininha e R. Verde não se agruparam com qualquer das populações citadas no nível de similaridade adotado (75%) constituindo grupos distintos e denominados, respectivamente, A, B, C e D.

As características forma e/ou posição dos cloroplastos nas células, regularidade das células na região multisseriada dos filamentos, forma do ápice e presença de septos transversais ("anéis") nos filamentos foram observadas em todo o material presentemente analisado. Os cloroplastos observados eram laminares e parietais, a disposição das células mostrou-se regular dependendo do diâmetro do filamento (conseqüentemente, dependendo do estágio de desenvolvimento do mesmo), os ápices eram truncados ou arredondados e a presença de "anéis" ocorreu em todos os filamentos.
Cariologia - As células que apresentaram as divisões celulares que permitiram as observações dos cromossomos ocorreram em qualquer região do filamento, sendo que este não apresentou uma área de maior atividade mitótica. As observações cariológicas (figuras 23-30) efetuadas nestas células mostraram um número de cromossomos igual a 12. Verificou-se que os mesmos mostraram-se como pequenos bastões ou pontos; além disso, as células da alga em questão apresentaram um par de cromossomos nitidamente curvos e bem maiores que os demais (figuras 27, 28). Devido ao tamanho reduzido dos cromossomos não foi possível medi-los e, nem mesmo, efetuar observações detalhadas em sua estrutura.

Discussão
Variação morfométrica - As diferenças significativas obtidas através da Anova para diâmetro e comprimento celular da região unisseriada, bem como para diâmetro do filamento, diâmetro e comprimento celular da região multisseriada tanto nas duas populações de Schizomeris leibleinii Kützing estudadas mensalmente (temporalmente), quanto nas 11 populações distintas espacialmente, comprovam a plasticidade dos indivíduos, provavelmente, em resposta às variações ambientais. Alguns indicativos dessa relação foram constatados a partir das correlações significativas entre características morfométricas e ambientais. Dados semelhantes foram encontrados por Branco & Necchi Júnior (1998) para Stigeoclonium helveticum Vischer e Chaetophora elegans (Roth) C. Agardh (Chaetophorales) quando as mesmas tiveram seus microhábitats estudados. Necchi Júnior et al. (1990), durante estudo fenológico em um riacho de 3ª ordem no Estado de São Paulo, também observaram variações nas dimensões de Compsopogon coeruleus (Balbis ex C. Agardh) Montagne (Rhodophyta) associadas às condições ambientais.
Embora essa grande amplitude de variação tenha sido numericamente marcante, os dados da análise de grupamento revelaram grande semelhança entre as amostras dos diferentes meses de coleta das populações, resultando na formação de grupos distintos, mas que apresentaram, entre si, similaridade acima de 60% das características analisadas. Esse dado demonstra que uma população mantém suas características básicas, mesmo apresentando amplos limites de variação. Além disso, a amplitude da variação dos caracteres morfométricos das regiões multi e unisseriadas foi avaliada com o teste t de Student entre as duas populações amostradas mensalmente e mostrou que ambas são morfometricamente semelhantes entre si.
Da mesma maneira que para as populações P3 e P4, a análise de grupamento revelou a formação de grupos distintos que, entretanto, possuem similaridade superior a 52% entre as populações estudadas. É interessante notar que a população denominada Cultura derivou-se da mesma população proveniente do Ribeirão Claro (P3) e, provavelmente, devido às alterações provocadas pelo crescimento em meio artificial, estas duas populações não se associaram intimamente de acordo com a análise efetuada. Alterações fenotípicas são relativamente freqüentes em algas pertencentes à ordem Chaetophorales que se desenvolvem em meios de cultura sob diferentes condições (Johnstone 1978, Francke & Ten Cate 1980, Francke 1982, Simons et al. 1986). Mais uma vez, os resultados obtidos indicam que a variação morfométrica, dentro de alguns limites, é bastante acentuada e depende do ambiente onde o organismo se desenvolve.
Embora a plasticidade fenotípica das Chaetophorales seja reconhecidamente ampla, tais variações ocorrem entre limites relativamente definidos. A interpretação equivocada das expressões morfológicas ou alterações de fenótipos pode levar a afirmações pouco aceitáveis. Campbell & Sarafis (1972), avaliando amostras de Schizomeris leibleinii na natureza e em cultura, afirmaram que a mesma seria uma forma de resistência adotada por Stigeoclonium tenue (C. Agardh) Kützing sob condições desfavoráveis do hábitat. Segundo as referidas autoras, o polimorfismo em Stigeoclonium Kützing é maior do que o conhecido, pois todas as formas e estágios de desenvolvimento que foram obtidos em cultura e encontrados na natureza seriam formas de crescimento desta alga. Assim, Campbell & Sarafis (1972) consideraram Schizomeris Kützing, Pearsoniella Fritsch & Rich, Uronema Lagerheim, Protoderma Kützing emend. Borzi e alguns estágios cocóides como todos sendo ecófenos de Stigeoclonium tenue. No entanto, estudos citológicos realizados por Mattox et al. (1974) em Schizomeris leibleinii mostraram que diferenças quanto aos zoósporos, a estrutura de pirenóides e cloroplastos e ao número de cromossomos seriam suficientes para distinguir o gênero Schizomeris do gênero Stigeoclonium.
A forma do sistema basal de fixação mostrou grande variabilidade entre as populações estudadas e, até mesmo, dentro das populações. Embora o tipo apressório também tenha sido o mais comumente encontrado, outras formas como sistemas rizoidais, lobados e claviformes foram observadas nas populações avaliadas devendo-se salientar que, em uma mesma população, até quatro tipos distintos de sistemas basais foram encontrados. A utilização da forma do sistema basal de fixação como característica taxonômica para distinção de espécies de Schizomeris, como proposto por Prasad & Srivastava (1963), não encontra suporte em dados de literatura e nos resultados obtidos neste trabalho (tabelas 4, 5, figuras 9-22). Kützing (1843b) cita rizóides para Schizomeris leibleinii, no entanto a descrição original deixa margem para a interpretação de apressório ("coberto por mucilagem"). Watson & Tilden (1930) citaram, para amostras de S.leibleinii coletadas em Minnesota (EUA), a ocorrência de sistema basal de fixação do tipo "bulbo" abruptamente arredondado ou alargado (claviforme) que seria típico da espécie S. irregularis Fritsch & Rich. Prasad & Srivastava (1963) observaram, em amostras em cultura de S. leibleinii, apenas os sistemas de fixação do tipo lobado ou ramificado (figuras 9-11). Patel (1967) citou para essa mesma espécie os sistemas de fixação do tipo disco (característico de S. indicum (Ghose) Fritsch & Rich), além dos tipos rizoidal e lobado (figuras 12, 16), destoando de trabalhos anteriores que mencionam apenas os sistemas basais do tipo rizoidal e lobado para S. leibleinii. Rino & Santos (1968) descreveram S. leibleinii portando discos de fixação (apressórios) para material coletado em Portugal. Sarma & Chaudhary (1975) observaram, em amostras de S. leibleinii provenientes da natureza e de material em cultura, a presença de sistemas basais do tipo rizoidal e lobado (claviforme) considerando, assim, esta característica como não confiável taxonomicamente, já que o sistema basal de fixação claviforme seria típico de S. irregularis. Yadava & Pandey (1980) verificaram a presença de rizóides, rizóides dicotômicos e "rizóides com bases dilatadas" (claviformes) em amostras mantidas em cultura e identificadas como S. leibleinii (figuras 17-20). Em nossas observações a presença de sistemas basais do tipo rizóide e apressório, este último considerado típico de S. indicum, foi freqüente, sendo que os tipos claviforme e lobado também foram encontrados, principalmente, no material mantido em cultura. Além disso, os filamentos que foram avaliados do material denominado Cultura, que derivou de subamostras do Ribeirão Claro (P3), apresentaram todos os quatro tipos de sistemas basais de fixação, enquanto este último (P3) deteve apenas três tipos. A análise do presente material sugere que S. leibleinii pode apresentar todos os sistemas basais descritos para as demais espécies do gênero.
Todavia, as observações pertinentes aos sistemas basais efetuadas pelos autores citados referem-se, principalmente, a amostras que cresciam em meios de cultura diversos. Segundo Francke (1982), algumas espécies de Stigeoclonium (Chaetophorales) que cresciam em culturas com altas concentrações de nitrogênio (56 mg.L-1) tiveram a forma dos talos prostrados (sistema basal) afetados (linhagens de Stigeoclonium tenue e S. farctum Berthold), fazendo com que as mesmas manifestassem talos prostrados diferentes dos característicos das espécies. Simons et al. (1986) verificando talos prostrados (sistemas basais) em Stigeoclonium notaram que os mesmos variavam morfologicamente em relação ao material recolhido no campo quando mantidos em cultura. Ainda, de acordo com Simons et al. (1986), a morfologia do sistema prostrado em Stigeoclonium também teria um significado funcional quanto à preferência aos tipos de substratos colonizados por este táxon. Mesmo sabendo que em alguns representantes da ordem Chaetophorales o polimorfismo é freqüente (ex. Stigeoclonium), baseando-se em nossas análises, nas inconsistentes denominações encontradas em literatura e na freqüência de citações de apressório para Schizomeris leibleinii, acreditamos que os sistemas basais de fixação do tipo claviforme e lobado sejam meramente estágios intermediários de desenvolvimento dos tipos apressório e rizóide, limitando a espécie a apenas dois sistemas de fixação ao substrato no indivíduo adulto (figuras 21, 22).
Além dessas características, Watson & Tilden (1930) e Prasad & Srivastava (1963) mencionam que Fritsch & Rich (1924) evidenciaram a diferença entre Schizomeris leibleinii e S. irregularis através da irregularidade na disposição das células na região multisseriada dos filamentos de S. irregularis.A respeito da disposição das células na região multisseriada dos filamentos, tanto no material presentemente analisado quanto no estudado por Yadava & Pandey (1980), ocorreram filamentos com a região multisseriada apresentando disposição regular ou irregular das células. Os estudos de Prasad & Srivastava (1963) indicam que o arranjo das células nessa região depende do estágio de desenvolvimento dos filamentos, sugerindo que Fritsch & Rich (1924) não tenham analisado todos os estágios de desenvolvimento nessa alga.
Outra característica adicional considerada importante é a presença de septos transversais como "anel" em S. leibleinii, enquanto tal estrutura não é conhecida em Schizomeris irregularis e S. indicum (Prasad & Srivastava 1963). A presença de septos transversais como "anéis" foi constatada na maioria das amostras, sendo que, freqüentemente a fragmentação dos filamentos ocorria nesta região.
O conjunto de dados obtido neste trabalho, bem como da literatura especializada (tabela 4), demonstra que as características morfométricas e morfológicas de Schizomeris leibleinii podem ser muito variáveis. No entanto, mesmo que a descrição original de S. leibleinii não faça menção à amplitude morfométrica e mesmo com amplos limites de variação, a espécie pode ser considerada bem definida taxonomicamente.
Dessa maneira, levando-se em consideração os dados obtidos para S. leibleinii, que as informações taxonômicas de Schizomeris irregularis e S. indicum são incompletas (tabela 5) e que as espécies foram citadas e/ou descritas apenas algumas vezes pela literatura, parece-nos que as espécies em questão não estão bem caracterizadas, apresentando sobreposição quanto à morfometria e aos sistemas basais de fixação com S. leibleinii. Assim, sugerimos que o gênero Schizomeris, da família Schizomeridaceae, seja monoespecífico contando apenas com a espécie S. leibleinii e que, baseando-se no princípio da prioridade do Código Internacional de Nomenclatura Botânica, as demais espécies sejam sinônimos taxonômicos.
Cariologia -Os cromossomos observados se mostraram como pequenos bastões, às vezes lembrando pontos, concordando com as observações de Prasad & Srivastava (1963), Chowdary (1967), Patel (1967) e Sarma & Chaudhary (1975). A presença de cromossomos curvos e mais longos que os demais (de formato em V) foi confirmada, assim como Sarma & Chaudhary (1975) encontraram em suas análises. Apesar de não ter sido possível efetuar medidas ou análises mais precisas quanto à forma dos cromossomos, segundo Sarma & Chaudhary (1975) a curvatura dos cromossomos sugere que os mesmos sejam metacêntricos.
A quantificação cromossômica das amostras avaliadas mostrou um número de cromossomos igual a 12, diferindo consideravelmente de todos os autores que efetuaram análises cariológicas em S. leibleinii. Prasad & Srivastava (1963) encontraram um n = 28 cromossomos para os espécimes coletados nas imediações de Lucknow (Utar Pradesh) na Índia. Chowdary (1967) encontrou nas amostras coletadas em Varanasi (Utar Pradesh), também na Índia, um n = 15 cromossomos, sugerindo que, mediante a investigação de Prasad & Srivastava (1963) e seus dados, haveria diversas raças citológicas de Schizomeris leibleinii nas localidades. Patel (1967) verificou, em espécimes provenientes de Vallabh Vidyanagar (Gujarat - Índia), n = 30 cromossomos nas células dessa mesma espécie, achando difícil decidir se a diferença nos números de cromossomos encontrados, quando comparado aos valores citados por Prasad & Srivastava (1963) e Chowdary (1967), seria devido à poliploidia, raças citológicas ou gerações haplóides ou diplóides dessa alga (alternância de gerações).
Contudo, o número de cromossomos (n = 14) determinado por Sarma & Chaudhary (1975) diferiu de todas as contagens anteriormente realizadas para Schizomeris leibleinii. Segundo os autores, parece que a variação no número de cromossomos, particularmente 28 e 30, descritos para S. leibleinii pode ser interpretada como resultado da duplicação das formas contendo 14 e 15 cromossomos. Ainda, de acordo com Sarma & Chaudhary (1975), as formas apresentando 15 e 30 cromossomos, bem como 14 e 28, podem ser interpretadas, também, como as fases haplóide e diplóide no ciclo de vida dessa alga, como suspeitado por Patel (1967). No entanto, informações a respeito do ciclo de vida dessa espécie são extremamente escassas. Kawasaki (1937) sugeriu que S. leibleinii apresenta alternância de gerações envolvendo gametas biflagelados (haplóides), produzidos por filamentos microscópicos (gametófitos), que se uniriam produzindo filamentos diplóides visíveis a olho nu (esporófitos) e ao atingirem a maturidade produziriam macrozoósporos quadriflagelados (haplóides), que reiniciariam o ciclo.
Schizomeris leibleinii pode apresentar diferentes raças citológicas, mas, no entanto, não é possível descartar a hipótese de que a espécie demonstre alternância de gerações quando considerados estudos anteriores. Sendo assim, recomenda-se que novos estudos envolvendo essa espécie sejam realizados, para fins de comparação quanto ao número de cromossomos e o conhecimento do seu histórico de vida.
Agradecimentos - A Maria Helena Carabolante (DZB/Ibilce/Unesp) pelo auxílio nas análises de laboratório, ao Dr. Orlando Necchi Júnior (DZB/Ibilce/Unesp) pela contribuição com os estudos cariológicos e ao Dr. Carlos E. M. Bicudo (IBt/SP, Seção de Ecologia) pelo empréstimo da obra com a descrição original de Schizomeris leibleinii.
(recebido: 19 de agosto de 2004; aceito: 13 de janeiro de 2005)
- BIRKBECK, T.E., STEWART, K.D. & MATTOX, K.R. 1974. The cytology and classification of Schizomeris leibleinii (Chlorophyceae). II. The structure of quadriflagellate zoospores. Phycologia 13:71-79.
- BOURRELLY, P. 1966. Les algues d'eau douce: 1. Les Algues Vertes. N. Boubee & Cie., Paris.
- BRANCO, C.C.Z. & NECCHI JUNIOR, O. 1998. Microhabitat and morphometric variation of two Chaetophoracean (Chaetophorales, Chlorophyta) species in tropical streams of southeastern Brazil. Phycological Research 46:169-174.
- BRANCO, L.H.Z. & PEREIRA, J.L. 2002. Evaluation of seasonal dynamics and bioindication potential of macroalgal communities in polluted tropical stream. Archiv für Hydrobiologie 155:147-161.
- CAMPBELL, E.O. & SARAFIS, V. 1972. Schizomeris - a growth form of Stigeoclonium tenue (Chlorophyta: Chaetophoraceae). Journal of Phycology 8:276-282.
- CHOWDARY, Y.B.K. 1967. A cytological study of Schizomeris leibleinii Kütz. Revue Algologie 8:302-306.
- CIENKOWSKI, L. 1876. Zur morphologie der Ulotricheen. Bulletin Scientific of Academy Imperial of Sciences of St. Petersburg 21:529-557.
- DIAS, I.C.A. 1985. Chlorophyta filamentosas da Lagoa de Juturnaíba, Araruama, Rio de Janeiro. Revista Brasileira de Botânica 8:93-99.
- DIAS, I.C.A. 1989. Chlorophyta filamentosas do município de Cáceres e arredores, Mato Grosso, Brasil: uma contribuição ao seu conhecimento. Acta Botanica Brasilica 3:3-12.
- DIAS, I.C.A. & BICUDO, C.E.M. 2001. Chaetophoraceae, Microsporaceae, Schizomeridaceae, and Ulotrichaceae from Biological Reserve of Poço das Antas, state of Rio de Janeiro, Brazil. Algological Studies 102:73-91.
- FOTT, B. 1959. Algenkunde. Fischer, Jena.
- FRANCKE, J.A. 1982. Morphological plasticity and ecological range in three Stigeoclonium species (Chlorophyceae, Chaetophorales). British Phycological Journal l7:117-133.
- FRANCKE, J.A. & TEN CATE, H.J. 1980. Ecotypic differentiation in response to nutritional factors in the algal genus Stigeoclonium Kütz. (Chlorophyceae). British Phycological Journal 15:343-355.
- FRITSCH, F.E. 1935. Structure and reproduction of algae. Cambridge University Press, London v.1.
- FRITSCH, F.E. & RICH, F. 1924. Contributions to our knowledge of freshwater algae of Africa. 4. Freshwater and subaerial algae from Natal. Transactions of the Royal Society South Africa 11:297-398.
- GHOSE, S.L. 1920. A new species of Uronema from India. Annals of Botany 34:95-98.
- HOLMGREN, P.K., HOLMGREN, N.H. & BARNETT, L.C. 1990. Index Herbariorum, part I, The herbaria of the World, 8th ed. New York Botanical Garden, New York.
- JOHNSTONE, I.M. 1978. Phenotypic plasticity in Draparnaldia (Chlorophyta: Chaetophoraceae). I. Effects of the chemical environment. Journal of Phycology 14:302-308.
- KAPRAUN, D.F. & MARTIN, D.J. 1987. Karyological studies of three species of Codium (Codiales, Chlorophyta) from coastal North Carolina. Phycologia 26:228-234.
- KAWASAKI, Y. 1937. On the life-history of Schizomeris leibleinii Kütz. Botanical Magazine 51:25-30.
- KÜTZING, F.T. 1843a. Ueber die systematische Eintheilung der Algen. Linnaea 17:75-107.
- KÜTZING, F.T. 1843b. Phycologia Generalis, oder Anatomie, Physiologie und Systemkunde der Tange. F.A. Brockhaus, Leipzig.
- KÜTZING, F.T. 1845. Phycologia Germanica. Nordhausen.
- MATTOX, K.R., STEWART, K.D. & FLOYD, G.L. 1974. The cytology and classification of Schizomeris leibleinii (Chlorophyceae). I. The vegetative thallus. Phycologia 13:63-69.
- NECCHI JUNIOR, O., GÓES, R.M. & DIP, M.R. 1990. Phenology of Compsopogon coeruleus (Balbis) Montagne (Compsopogonaceae, Rhodophyta) and evaluation of taxonomic characters of the genus. Japanese Journal of Phycology 38:1-10.
- NECCHI JUNIOR, O., PASCOALOTO, D., BRANCO, C.C.Z. & BRANCO, L.H.Z. 1997. Stream macroalgal flora of the northwest region of São Paulo State, southeastern Brazil. Algological Studies 84:91-112.
- NICHOLS, H.W. 1973. Growth media-freshwater. In Handbook of Phycological Methods: Culture Methods and Growth Measurements (H.W. Nichols, ed.). Cambridge University Press, Cambridge, p.7-24.
- PAPENFUSS, G.F. 1955. Classification of algae in a century of progress in Natural Sciences 1853-1953. In A century of progress in the Natural Sciences 1853-1953 (E.L. Kessel, ed.). California Academy of Sciences in San Francisco, San Francisco, v.8, p.115-224.
- PATEL, R.J. 1967. On morphology and cytology of Schizomeris leibleinii Kuetz. from Gujarat. Phykos 6:87-90.
- PRASAD, B.N. & SRIVASTAVA, P.N. 1963. Observations on the morphology, cytology, and asexual reproduction of Schizomeris leibleinii Phycologia 2:148-156.
- PRESCOTT, G.W. 1962. Algae of the Western Great Lakes Area. Wm C. Brown, Dubuque.
- PRESCOTT, G.W. 1968. The Algae: A Review. Houghton Mifflin, Boston.
- PRINTZ, H. 1927. Chlorophyceae. In Die Näturlichen Pflanzenfamilien (A. Engler & K. Prantl, eds.). Wilhelm Engelmann, Leipzig, v.3, p.463.
- RINO, J.A. & SANTOS, M.F. 1968. Subsídios para o conhecimento das algas de Portugal - III. Anuário da Sociedade Broteriana 34:19-56.
- SARMA, Y.S.R.K. & CHAUDHARY, B.R. 1975. On a new cytological race of Schizomeris leibleinii Kütz. Hydrobiologia 47:181-191.
- SIMONS, J., VAN BEEM, A.P. & DE VRIES, P.J.R. 1986. Morphology of the prostrate thallus of Stigeoclonium (Chlorophyceae, Chaetophorales) and its taxonomic implications. Phycologia 25:210-220.
- TIWARI, M.G. 1994. New record of Schizomeris leibleinii (alga) from polluted habitats of south Chotanagpur, Bihar. Journal of Ecobiology 6:71-72.
- WATSON, J.B. & TILDEN, J.E. 1930. The algal genus Schizomeris and the occurrence of Schizomeris leibleinii Kützing in Minnesota. Transactions of the American Microscopical Society 49:160-167.
- WITTMANN, W. 1965. Aceto-iron-haematoxylin-chloral hydrate for chromosome staining. Stain Technology 40:161-164.
- YADAVA, R.N. & PANDEY, D.C. 1980. Certain cultural observations on Schizomeris leibleinii Kuetz. Phykos 19:204-209.
Datas de Publicação
-
Publicação nesta coleção
13 Jan 2006 -
Data do Fascículo
Jun 2005
Histórico
-
Recebido
19 Ago 2004 -
Aceito
13 Jan 2005