Resumos
As pastagens formam a base da pecuária brasileira; essas sofrem degradação em larga escala por deficiência de nitrogênio (N). O consórcio com leguminosas, além de fixar N, pode apresentar outros efeitos na fertilidade do solo como acidificação ou retirada de nutrientes de camadas mais profundas para as mais superficiais. Este trabalho objetivou avaliar o estoque de serapilheira e a fertilidade do solo em pastagens degradadas de braquiária (Brachiaria decumbens), após implantar leguminosas arbustivas e arbóreas forrageiras. Para isso, uma amostragem foi realizada em março de 2010 em um experimento no campo, introduzindo pastagem degradada de Brachiaria decumbens, em julho de 2008, com sabiá (Mimosa caesalpiniifolia), leucena (Leucaena leucocephala), mororó (Bauhinia cheilantha) e gliricídia (Gliricidia sepium), além de braquiária adubada e não adubada com N. As amostras de solo e serapilheira foram coletadas aos 0-10, 10-20 e 20-40 cm de profundidade, em três transectos, alternando pontos cobertos por gramíneas e leguminosas, totalizando sete amostras compostas por parcela para determinar pH, P, K, Ca, Mg e Al no solo, enquanto SB, t e m foram calculados. A serapilheira foi separada visualmente em leguminosas, gramíneas e materiais não identificados, em que foram utilizados para quantificação de matéria seca, matéria orgânica, N, P, C, fibra detergente ácido e lignina. A introdução das leguminosas aumentou os teores de N total na serapilheira e reduziu as relações C:N, com destaque para gliricídia e sabiá; entretanto, essa última apresentou elevados teores de lignina. Houve efeito significativo da cobertura por leguminosas, sem diferenças entre essas, para pH e K, na profundidade de 0-10 cm, e para Al e m, aos 10-20 cm de profundidade.
nutrientes; fixação biológica de nitrogênio; silvopastoril; pastagens tropicais
Pastures are the fodder supply of Brazilian livestock, and are exhaustively degraded by nitrogen (N) deficiency. Intercropping with legumes, aside from nitrogen fixation, may have secondary effects on soil fertility, such as acidification or nutrient movement from deeper to the surface soil layers. This study evaluated the soil litter stock and fertility of degraded Brachiaria decumbens pastures after planting leguminous shrubs and forage trees. For this purpose, we sampled (March 2010) degraded Brachiaria decumbens pasture planted in July 2008 in an intercropping experiment with sabiá (Mimosa caesalpiniaefolia), leucaena (Leucaena leucocephala), mororó (Bauhinia cheilantha) and gliricidia (Gliricidia sepium), as well as N-fertilized and unfertilized brachiaria. Soil and litter was sampled (layers 0-10, 10-20 and 20-40 cm) in three transects, at alternating points covered by grasses and by legumes, totaling seven composite samples per plot to determine soil pH, P, K , Ca, Mg, and Al and calculate sum of bases, effective cation exchange capacity and aluminum saturation. Litter was visually separated in legumes, grasses and unidentified material to quantify dry matter, organic matter, N, P, C, acid-detergent fiber, and lignin. The use of legumes increased the levels of total N in litter and reduced the C: N ratios, especially of gliricidia and sabiá, although the lignin levels in the latter were high. There was a significant effect of legume soil cover, with no differences between them, on pH and K (layer 0-10 cm) and on pH, K and Al (layer 10-20 cm).
nutrients; biological nitrogen fixation; forest-pasture; tropical pastures
DIVISÃO 3 - USO E MANEJO DO SOLO
COMISSÃO 3.3 - MANEJO E CONSERVAÇÃO DO SOLO E DA ÁGUA
Estoque de serapilheira e fertilidade do solo em pastagem degradada de Brachiaria decumbens após implantação de leguminosas arbustivas e arbóreas forrageiras(1 (1 ) Parte da Dissertação de Mestrado do primeiro autor. )
Soil litter stock and fertility after planting leguminous shrubs and forage trees on degraded signal grass pasture
André Barbosa SilvaI; Mario Andrade Lira JuniorII; José Carlos Batista Dubeux JuniorIII; Márcia do Vale Barreto FigueiredoIV; Rayssa Pereira VicentinI
IDoutorando(a), Departamento de Agronomia, Universidade Federal Rural de Pernambuco - UFRPE. Rua Dom Manoel de Medeiros, s/n, Dois Irmãos. CEP 52171-900 Recife (PE). E-mail: andreufrpe@gmail.com; rayssavicentin@gmail.com
IIProfessor, Departamento de Agronomia, UFRPE. Bolsista de produtividade do CNPq. E-mail: mario.lira@depa.ufrpe.br
IIIProfessor, Departamento de Zootecnia, UFRPE. Bolsista de produtividade do CNPq. E-mail: dubeux@dz.ufrpe.br
IVPesquisador, Empresa Pernambucana de Pesquisa Agropecuária - IPA. Av. General San Martin, 1371, Bongi. CEP 50761-000 Recife (PE). Bolsista de produtividade do CNPq. E-mail: mbarreto@elogica.com.br
RESUMO
As pastagens formam a base da pecuária brasileira; essas sofrem degradação em larga escala por deficiência de nitrogênio (N). O consórcio com leguminosas, além de fixar N, pode apresentar outros efeitos na fertilidade do solo como acidificação ou retirada de nutrientes de camadas mais profundas para as mais superficiais. Este trabalho objetivou avaliar o estoque de serapilheira e a fertilidade do solo em pastagens degradadas de braquiária (Brachiaria decumbens), após implantar leguminosas arbustivas e arbóreas forrageiras. Para isso, uma amostragem foi realizada em março de 2010 em um experimento no campo, introduzindo pastagem degradada de Brachiaria decumbens, em julho de 2008, com sabiá (Mimosa caesalpiniifolia), leucena (Leucaena leucocephala), mororó (Bauhinia cheilantha) e gliricídia (Gliricidia sepium), além de braquiária adubada e não adubada com N. As amostras de solo e serapilheira foram coletadas aos 0-10, 10-20 e 20-40 cm de profundidade, em três transectos, alternando pontos cobertos por gramíneas e leguminosas, totalizando sete amostras compostas por parcela para determinar pH, P, K, Ca, Mg e Al no solo, enquanto SB, t e m foram calculados. A serapilheira foi separada visualmente em leguminosas, gramíneas e materiais não identificados, em que foram utilizados para quantificação de matéria seca, matéria orgânica, N, P, C, fibra detergente ácido e lignina. A introdução das leguminosas aumentou os teores de N total na serapilheira e reduziu as relações C:N, com destaque para gliricídia e sabiá; entretanto, essa última apresentou elevados teores de lignina. Houve efeito significativo da cobertura por leguminosas, sem diferenças entre essas, para pH e K, na profundidade de 0-10 cm, e para Al e m, aos 10-20 cm de profundidade.
Termos de indexação: nutrientes, fixação biológica de nitrogênio, silvopastoril, pastagens tropicais.
SUMMARY
Pastures are the fodder supply of Brazilian livestock, and are exhaustively degraded by nitrogen (N) deficiency. Intercropping with legumes, aside from nitrogen fixation, may have secondary effects on soil fertility, such as acidification or nutrient movement from deeper to the surface soil layers. This study evaluated the soil litter stock and fertility of degraded Brachiaria decumbens pastures after planting leguminous shrubs and forage trees. For this purpose, we sampled (March 2010) degraded Brachiaria decumbens pasture planted in July 2008 in an intercropping experiment with sabiá (Mimosa caesalpiniaefolia), leucaena (Leucaena leucocephala), mororó (Bauhinia cheilantha) and gliricidia (Gliricidia sepium), as well as N-fertilized and unfertilized brachiaria. Soil and litter was sampled (layers 0-10, 10-20 and 20-40 cm) in three transects, at alternating points covered by grasses and by legumes, totaling seven composite samples per plot to determine soil pH, P, K , Ca, Mg, and Al and calculate sum of bases, effective cation exchange capacity and aluminum saturation. Litter was visually separated in legumes, grasses and unidentified material to quantify dry matter, organic matter, N, P, C, acid-detergent fiber, and lignin. The use of legumes increased the levels of total N in litter and reduced the C: N ratios, especially of gliricidia and sabiá, although the lignin levels in the latter were high. There was a significant effect of legume soil cover, with no differences between them, on pH and K (layer 0-10 cm) and on pH, K and Al (layer 10-20 cm).
Index terms: nutrients, biological nitrogen fixation, forest-pasture, tropical pastures.
INTRODUÇÃO
Os solos sob pastagens no Brasil frequentemente apresentam problemas de fertilidade (Macedo, 2009). Em razão disso, a baixa disponibilidade de nutrientes, principalmente nitrogênio (N) e fósforo (P), e a alta saturação por alumínio (m) são os fatores químicos que mais limitam a produção de forragem (Lopes et al., 2011). Dubeux Jr. et al. (2007) comentam que a diminuição da fertilidade em solo sob pastagens está relacionada principalmente com as perdas de nutrientes, principalmente N e P, sem que ocorram reposições adequadas, acentuando quando o manejo da pastagem não favorece a ciclagem de nutrientes.
A introdução de leguminosas forrageiras fixadoras de N aumenta a disponibilidade desses nutrientes pelo retorno por meio de excreções de animais que as consumem e também via incorporação de material não consumido ao solo (Hakala & Jauhiainen, 2007; Rodrigues et al., 2008). Essas plantas também promovem o acúmulo de quantidades substanciais de carbono (C), que pode favorecer melhorias nas propriedades físicas e químicas do solo, aprimoramento das atividades biológicas e aumento da disponibilidade de nutrientes (Macedo et al., 2008; Cayuela et al., 2009). Diante das dificuldades de manejo do consórcio em pastagens tropicais entre gramíneas e leguminosas herbáceas existe também a possibilidade de uso de espécies arbustivo-arbóreas, em um sistema silvipastoril (Dias et al., 2007; Barcellos et al., 2008), em particular em solos de baixa fertilidade do que naqueles de fertilidade mediana a alta, principalmente na área próxima às árvores (Pandey, 2007; Kimaro et al., 2008).
Nascimento et al. (2003), avaliando o efeito de 12 espécies leguminosas na composição química de um solo degradado, concluíram que houve redução na acidez do solo e elevação dos teores de K e Mg, na profundidade de 0-10 cm. Já Barretos & Fernandes (2001), testando o cultivo de gliricídia e leucena em alamedas de tabuleiros costeiros, perceberam elevação no pH e nos teores de Ca + Mg, sem alteração nos teores de matéria orgânica e na capacidade de troca de cátions. Essas alterações ocorrem provavelmente porque essas plantas possuem sistemas radiculares mais desenvolvidos, que são capazes de absorver nutrientes do solo em horizontes mais profundos, posteriormente depositados como serapilheira (Menezes et al., 2002; Tiessen et al., 2003).
O objetivo deste trabalho foi avaliar o estoque de serapilheira e a fertilidade do solo em pastagens degradadas de Brachiaria decumbens, após implantação de leguminosas arbustivas e arbóreas forrageiras.
MATERIAL E MÉTODOS
Ambiente experimental
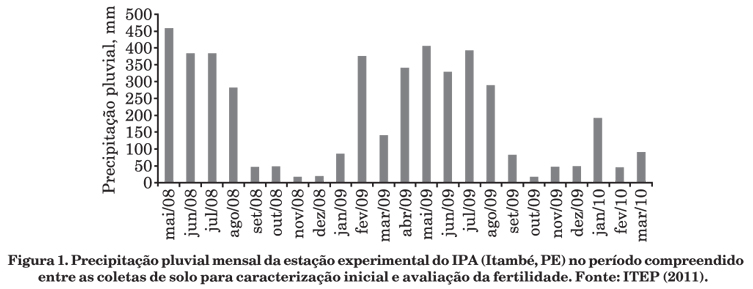
Este trabalho foi conduzido em pastos degradados de braquiária (Brachiaria decumbens), implantados no final da década de 1980, após introdução de leguminosas arbustivas e arbóreas, em julho de 2008, na Estação Experimental do Instituto Agronômico de Pernambuco (IPA), em Itambé, PE (07º 25' S, 35º 06' W, 190 m de altitude). O clima do tipo AS' na classificação de Köppen é quente e úmido, com precipitação pluvial média de 1.200 mm ano-1, temperatura média anual de 24 ºC e umidade relativa média do ar de 80 %, com os meses mais chuvosos ocorrendo geralmente de abril a julho (IPA, 1994; CPRH, 2003). A precipitação pluvial média mensal da implantação do experimento à amostragem (Figura 1) foi de 196,53 mm (ITEP, 2011). Os solos predominantes nessa estação são classificados como Argissolo Vermelho-Amarelo Tb distrófico úmbrico, A proeminente, textura média/argilosa, fase florestal tropical subcaducifólia e relevo suave ondulado (Jacomine et al., 1973; Embrapa, 2006).
Inicialmente, foi realizada a caracterização da fertilidade do solo nas profundidades 0-10, 10-20 e 20-40 cm (Quadro 1), conforme Embrapa (1999), e feita a correção da acidez do solo, aplicando-se 1 Mg ha-1 de calcário e adubação com 50 kg ha-1 cada de P2O5 e K2O.
Tratamentos e delineamento experimental
Os tratamentos foram distribuídos num delineamento em blocos ao acaso com seis tratamentos e quatro repetições, sendo quatro associações gramínea/leguminosa e dois tratamentos mantendo braquiária, um não adubado (braquiária) e o outro com aplicação de 60 kg ha-1 ano-1 de N na forma de ureia (braquiária adubada), sendo 30 kg de N aplicados em maio de 2009 e março de 2010 e outros 30 kg de N, 56 dias após a primeira aplicação.
As leguminosas avaliadas foram sabiá (Mimosa caesalpiniifolia Benth.), leucena (Leucaena leucocephala (Lam.) de Wit), mororó (Bauhinia cheilantha (Bong) Steud) e gliricídia (Gliricidia sepium (Jacq.) Kunthex Walp.). As plantas foram multiplicadas mediante a preparação de mudas em casa de vegetação, visando pleno estabelecimento das espécies. Antes do plantio, as sementes foram escarificadas mecanicamente com lixa, sendo postas para germinar em caixas de germinação de isopor, utilizando vermiculita como substrato, bem como a solução de Hoagland isenta de N para nutrição das plantas. As plântulas foram inoculadas com solução de inoculante específico produzido na UFRPE (Quadro 2 ), com as estirpes recomendadas nacionalmente à época (DAS/MAPA, 2006) e fornecidas pela Embrapa Agrobiologia, exceto o mororó por ser não nodulante.
O transplantio foi realizado quando as mudas apresentaram altura média de 12-15 cm, em fileiras duplas com espaçamento de 10 x 1,0 x 0,5 m, sendo cada parcela média de 660 m2 (33 x 20 m), com três filas duplas de leguminosa por parcela (Figura 2). Cada faixa foi preparada após aplicação de glifosato para controle da braquiária.
Amostragem de serapilheira
A serapilheira foi coletada em março de 2010, no final da estação seca, durante a qual as parcelas experimentais não foram pastejadas de modo similar ao descrito por Machado (2011). Foram utilizados transectos posicionados aos 5, 10 e 15 m do comprimento das faixas de cultivo e aos 2,5; 5,5; 11; 16,5; 22; 27,5; e 30,5 m da largura da parcela (Figura 2). Para a análise de dados, os pontos de coleta foram agrupados em três distâncias das faixas de leguminosas, com o centro das faixas de leguminosas correspondendo à distancia 0 (5,5; 16,5; e 27,5 m), à distância de 2,5 m das faixas de leguminosas (2,5 e 30,5 m) e aos 5,0 m de distância (11 e 22 m).
Em cada ponto, a serapilheira correspondente ao material vegetal senescente depositado na superfície do solo (folhas, frutos e ramos) foi coletada manualmente com o auxílio de uma moldura de 25 x 25 cm e colocada para secar em estufa de circulação forçada a 65 ºC, por aproximadamente 72 h. As amostras secas foram separadas visualmente em leguminosa, gramínea e outros materiais não identificados e pesadas as frações, bem como calculada a quantidade de serapilheira de gramínea e leguminosa depositada. Posteriormente, foram preparadas amostras de aproximadamente 30 g, mantendo a proporção gramínea/leguminosa para cada amostra composta, que foram trituradas em moinhos de facas e armazenadas para determinação da matéria seca (MS) e dos teores de matéria orgânica (MO), conforme Bezerra Neto & Barreto (2004); nitrogênio total (N total), determinado pelo método Kjeldahl; fibra detergente ácido (FDA) e lignina nos tecidos vegetais, de acordo com os métodos descritos em Silva & Queiroz (2006); carbono orgânico (CO), como descrito por Bezerra Neto & Barreto (2004); e fósforo, discriminado a partir de extratos preparados com o resíduo mineral, proveniente da definição da MO, segundo Bezerra Neto & Barreto (2004). Esses resultados também foram utilizados para cálculo dos conteúdos totais de MO e N da serapilheira.
Solo
A amostragem de solo foi conduzida da mesma forma que para a serapilheira, com auxilio de trado holandês, nas profundidades 0-10, 10-20, 20-40 cm, usada para avaliar a fertilidade do solo, segundo os métodos descritos em Embrapa (1999). Foram determinados o pH em água (relação 1:2,5); K, Na e P foram extraídos com Mehlich-1, aferidos por fotometria de chama (Na e K) e por espectrofotometria UV-VIS (P), enquanto Ca, Mg e Al foram extraídos com KCl e dosados por espectrofotometria de absorção atômica (Ca e Mg) e por titulação (Al). Com base nesses teores, foi possível estimar a soma de bases trocáveis (SB), a capacidade de troca de cátions efetiva (t) e a saturação pelo alumínio (m) no complexo de troca.
Análise estatística
Os dados foram inicialmente avaliados quanto à necessidade de transformações e eliminações de outliers utilizando o Guided Data Analysis Procedure do SAS, sendo seguidas as suas recomendações (SAS, 1999). Foi considerado o delineamento em blocos com esquema em faixas. A parcela principal foi formada pelos tratamentos de braquiária e leguminosas e as faixas foram formadas pelas três classes de distância entre leguminosa e gramínea, descritas durante a amostragem. Realizou-se teste de contraste ortogonal entre os tratamentos com braquiária solteira em contrapartida aos com o consórcio com leguminosas e entre os tratamentos de braquiária solteira com e sem adubação nitrogenada, em nível de significância de 5 %. Após essa análise, apenas os tratamentos com leguminosa foram analisados separadamente, utilizando as mesmas transformações e eliminações de outliers da fase anterior, sendo aplicado o teste de Tukey a 5 %, quando apropriado.
RESULTADOS E DISCUSSÃO
Serapilheira
Foram encontradas interações significativas (p<0,05) entre as espécies e as faixas para MO total, teor de N total, FDA, lignina, proporção de leguminosas, relação C:N e conteúdo de N, enquanto para concentração de C, P, conteúdo de P, relações C:P e lig:N só foram observados efeitos isolados para as distâncias avaliadas.
A proporção média de leguminosa na serapilheira foi influenciada tanto pela faixa quanto pela espécie introduzida, com maiores percentuais de leguminosas na distancia 0,0 m e menores nas 2,5 e 5,0 m; contudo, esses últimos não diferiram entre si. O sabiá apresentou maior proporção de leguminosas em todas as faixas, não diferindo de gliricídia e mororó apenas na faixa das leguminosas (Quadro 3). Apesar de a deposição de serapilheira de leguminosas influenciar mais a região sob a copa, há indícios de que para o sabiá essa deposição também contribuiu para alterações nas áreas adjacentes, quando a proporção de leguminosas é avaliada conjuntamente com a MO total da serapilheira.
O elevado acúmulo de serapilheira das gramíneas C4 em relação às leguminosas deve-se à sua maior eficiência na fotossíntese, o que repercute na produção de fitomassa e consequentemente na deposição de material ao solo (Braz et al., 2005; Dias et al., 2007). Os valores encontrados nas faixas de braquiárias superam o 1,58 Mg ha-1 encontrado por Santos et al. (2007), em pastagens em avançado estádio de degradação de B. decumbens com 20 anos de cultivo.
O baixo acúmulo de serapilheira do mororó ocorreu provavelmente por causa da sua menor taxa de crescimento, já observada durante a implantação do experimento (Silva et al., 2009), o que comprometeu sua persistência e produção de biomassa na parte aérea. Foi observado maior consumo da leucena, enquanto na gliricídia e no sabiá quase não houve consumo, o que pode justificar a baixa produção de serapilheira, com apenas 1,38 Mg ha-1 na faixa da leguminosa; menor ainda quando considerada apenas a participação dela na composição da serapilheira. Bertalot et al. (2004) observaram média de 1,39 Mg ha-1 ano-1 em monocultivo e sem animais.
Sabiá e gliricídia apresentaram grande deposição de serapilheira, como descrito por Costa et al. (2004), que observaram deposição anual de 3,3 Mg ha-1 ano-1 na gliricídia e 8,8 Mg ha-1 ano-1 no sabiá, sugerindo maior acúmulo de serapilheira existente para o sabiá, o que se repetiu neste experimento, pois quando foram considerados apenas os materiais dessas leguminosas a serapilheira encontrada foi de 0,44 e 1,56 Mg ha-1 respectivamente para gliricídia e sabiá, com base na proporção de serapilheira da leguminosa na MO total da faixa "0" (Quadro 3), confirmando a superioridade do sabiá.
Além desses autores, Ferreira et al. (2007) e Freire et al. (2010) verificaram, respectivamente, acúmulos de 8,91 e 6,4 Mg ha-1 de serapilheira em bosque de sabiá, bastante superiores aos encontrados neste trabalho; no entanto, nos dois casos, os dados foram obtidos em monocultivo maduro, com base em avaliações mensais, enquanto neste estudo foi realizado apenas no final do período seco.
Os teores de N total variaram significativamente entre as espécies apenas nas faixas de leguminosas, com aumento em relação às faixas de braquiárias. Entre as espécies, o sabiá e a gliricídia (12,3 e 11,4 g kg-1, respectivamente) foram as que apresentaram os maiores teores na serapilheira; mororó (8,1 g kg-1) apresentou o menor; entretanto, nenhum desses diferiram da leucena (9,7 g kg-1), que evidenciou valor intermediário (Quadro 3). Os elevados teores de N total no sabíá, somados à maior massa de serapilheira depositada, permitiram maiores conteúdos de N presente nas faixas cobertas por essa em relação as demais leguminosas, não diferindo apenas da gliricídia. O conteúdo de N no tratamento com sabiá não diferiu aos 0,0 e 2,5 m de distância, mas foi superior aos 5,0 m, o que sugeriu uma contribuição do sabiá fora de sua faixa de plantio (Quadro 3). Além disso, a proporção de leguminosa no sabiá foi maior do que na glirícídia, o que pode ter reduzido a vantagem qualitativa das folhas de gliricídia, em detrimento a do sabiá observada por Costa et al. (2004). Apesar do incremento nos teores de N nos pontos cobertos por leguminosas, esses ainda são inferiores ao valor crítico (1,74 dag kg-1), para que haja mineralização líquida desse nutriente (Palm, 1995).
Os baixos teores de N total no mororó podem ser explicados pelo fato de essa ser uma leguminosa não nodulante (Freitas et al., 2005), mas mesmo assim o teor de N total sob essa leguminosa foi cerca de 35 % maior do que a 2,5 m de distância, o que indica a necessidade de maiores estudos quanto a esse aumento nos teores de N promovidos por essa espécie. Além disso, essa espécie apresentou elevados teores de proteínas, que podem variar de 9,7 a 20,7 dag kg-1; dependendo do estádio vegetativo (Araújo Filho et al., 2002), pode ter participação de até 36 % na dieta do gado na caatinga, principalmente no período seco (Moreira et al., 2006). Os teores de N total encontrados na serapilheira da leucena foram inferiores aos observados por Bertalot et al. (2004), os quais obtiveram de 1,6 a 2,9 dag kg-1, variando conforme a estação do ano, provavelmente em razão da menor proporção da leguminosa na serapilheira.
A relação C:N da serapilheira foi menor nas faixas das leguminosas, com médias de 42,0; 44,6; 49,0; e 58,8 para sabiá, gliricídia, leucena e mororó, respectivamente, contra 80,1 e 83,2 para a faixa de gramínea nos pontos 2,5 e 5,0 m (Quadro 3). As maiores diferenças foram observadas no sabiá e na gliricídia, especialmente no primeiro, com as relações C:N nas faixas de gramínea praticamente o dobro das encontradas sob leguminosa. Como a relação C:N está relacionada com a velocidade de decomposição da serapilheira (Gama-Rodrigues et al., 2007), logo a redução nessa relação favoreceu maior decomposição dos resíduos e contribuiu para o equilíbrio entre a mineralização e a imobilização dos nutrientes. Porém, os valores encontrados ainda são considerados altos, já que esse equilíbrio ocorreria entre 20-30:1 (Sá, 1999).
Apenas o sabiá promoveu elevação nos teores de FDA, com média de 76 dag kg-1 nos pontos cobertos pelo sabiá, muito superior à média das faixas de gramíneas entre 61,1 e 61,5 dag kg-1 (Quadro 3). Para os teores de lignina observados na faixa da leguminosa, a média foi de 38,1 dag kg-1, superior aos cobertos por braquiária, entre 21,9 e 19,1 dag kg-1 (Quadro 3). Elevados teores de FDA indicam maior proporção dos componentes fibrosos resistentes (van Soest, 1994), o que pode dificultar a decomposição desse material e revelar menor qualidade da serapilheira do sabiá, em particular combinado com os valores elevados de lignina encontrados no sabiá, que também podem dificultar a decomposição (Aita & Giacomini, 2003).
Não foram verificadas interações na relação lignina:N, teores de CO, lignina e P (Quadro 4), apenas efeito isolado das faixas, com maiores teores de lignina e P sob as leguminosas. Já quanto ao conteúdo total de P, o menor resultado foi observado na faixa de leguminosa, provavelmente por causa da menor serapilheira.
A ausência de efeito significativo para P em razão das espécies implantadas pode estar relacionada aos baixos teores encontrados no solo, já observados no quadro 1. Salienta-se que a realização da coleta no final do período seco pode ter comprometido a disponibilidade desse nutriente no período (Lima et al., 2011), já que os baixos teores encontrados no solo podem favorecer maior mobilidade desse elemento na planta (Marschner, 1995) e menores teores no material senescente, assim como foi verificado nos estudos de Balieiro et al. (2004) com Acacia mangium Willd.
Solo
Não houve interações significativas entre as espécies de cobertura e os pontos coletados para qualquer característica de fertilidade do solo; somente alguns efeitos isolados da posição da amostra, quando avaliados o pH e K+ nas profundidades 0-10 e 10-20 cm, e Al3+ e m, apenas na profundidade 10-20 cm.
O solo sob leguminosas foi mais ácido (Quadro 5) nos pontos cobertos por leguminosas (pH~5,0) do que nas faixas com braquiária (pH~5,3), nas profundidades 0-10 e 10-20 cm; no entanto, não houve diferença significativa na profundidade 20-40 cm. Essa acidificação pode ter ocorrido em razão de nitrificação e lixiviação do nitrato como observado por Grappegia Júnior et al. (2000) com mucuna cinza (Stizolobium cinereum), possivelmente pela maior absorção de cátions que as gramíneas, liberando mais íons H+ para a solução do solo, combinada com a menor absorção de nitrato por causa da fixação simbiótica (Bohnen et al., 2006). Já o Al3+ apresentou efeito significativo apenas na camada 10-20 cm, observando-se maiores concentrações nos pontos cobertos pelas leguminosas, como poderia ser esperado pela redução do pH, confirmando aumento na saturação por esse elemento (Quadro 5). Além disso, os pontos das gramíneas mais próximos das leguminosas (2,5 m) provavelmente foram influenciados por essas, apresentando valores intermediários da concentração de Al de tal forma que não apresentaram diferenças significativas; entretanto, esse comportamento não foi observado nos valores de pH e m.
As gramíneas costumam ser mais eficientes na remoção de cátions monovalentes do solo do que as leguminosas (Castellane & Fontes, 1983; Silva et al., 2010), o que foi confirmado pelos teores significativamente mais altos de K+, nas duas profundidades, nas faixas cobertas por gramíneas do que na coberta por leguminosa (Quadro 5), provavelmente por causa da maior deposição de material mais rico em K e, consequentemente, maior disponibilidade desse elemento.
Não foram observados efeitos significativos para Ca2+, Mg2+ e P, embora os maiores teores tenham sido observados nas camadas mais superficiais, provavelmente em decorrência da adubação e calagem superficiais, influenciando, portanto, os teores dos nutrientes nessa camada e levando à redução do Al3+, o que permitiu a elevação da CTC (t) e da soma de bases (SB) nas camadas superficiais e a redução da saturação por Al (m) (Quadro 6). Este resultado difere do encontrado por Barretos & Fernandes (2001), os quais, testando o cultivo de gliricídia e leucena solteiras em alamedas de tabuleiros costeiros, verificaram elevação nos teores de Ca e Mg, com a incorporação do material vegetal dessas plantas ao solo. Como esses autores, Nascimento et al. (2003) também verificaram que a introdução de leguminosas em areas degradadas promoveu a elevação dos teores de K e Mg, na profundidade de 0-10 cm, enquanto Santos et al. (2001), assim como neste trabalho, observaram decréscimo nos teores de Ca e Mg em razão da profundidade, independentemente de ser gramínea ou leguminosa. Macharia et al. (2011) observaram que a introdução de leguminosas em pastagens naturais promoveu a redução nos teores de Ca e K, atribuindo essa redução à baixa produção de MS das leguminosas, quando em consórcio, e provável competição por nutrientes entre gramíneas e leguminosas.
Espera-se que a deposição gradual de serapilheira rica em N sob a área de influência de leguminosas arbóreas promova o aumento da fertilidade do solo e dos teores de MO (Oliveira et al., 2000; Andrade et al., 2002), o que não se confirmou neste trabalho. Entretanto, a ausência de resultados neste estudo provavelmente deve-se à sua relação com teores de MO, que em solos tropicais aumenta em longo prazo (Dulormne et al., 2003). Além disso, o solo em questão já apresentava elevados teores de MO em todas as profundidades (entre 4 e 5 %) no início do experimento (Quadro 1), o que provavelmente contribuiu para a baixa diferenciação nos parâmetros de fertilidade. Talvez seja necessário maior tempo para que possam ser constatados mais efeitos significativos sobre a fertilidade. É importante ressaltar também que o P é o principal nutriente limitante da fixação biológica de N e da produção de biomassa nos sistemas naturais (Peoples & Craswell, 1992) e que foram verificados baixos teores desse elemento nas camadas de 10-20 e 20-40 cm, o que pode ter comprometido a absorção pelas leguminosas; logo, a fixação biológica e a produção de biomassa.
CONCLUSÕES
1. A introdução das leguminosas na pastagem melhora a qualidade da serapilheira disponível, especialmente gliricídia e sabiá, sendo a deposição de serapilheira dessas leguminosas importante via de retorno de nitrogênio para o solo.
2. O consórcio com leguminosas promove alterações nos valores de pH, K, Al e m, porém sem distinção entre as espécies e apenas nas profundidades 0-10 cm para os dois primeiros e 10-20 cm para esses e os demais listados.
3. A acidificação do solo nesses sistemas limita-se às camadas mais superficiais e faixas cobertas por leguminosas.
LITERATURA CITADA
Recebido para publicação em 2 de março de 2012 e aprovado em 30 de janeiro de 2013.
- AITA, C. & GIACOMINI, S.J. Decomposição e liberação de nitrogênio de resíduos culturais de plantas de cobertura de solo solteiras e consorciadas. R. Bras. Ci. Solo, 27:601-612, 2003.
- ANDRADE, C.M.S.; VALENTIM, J.F. & CARNEIRO, J.C. Árvores de baginha (Stryphnodendron guianense (Aubl.) Benth.) em ecossistemas de pastagens cultivadas na Amazônia Ocidental. R. Bras. Zootec., 31:574-582, 2002.
- ARAÚJO FILHO, J.A.; CARVALHO, F.C. & SILVA, N.L. Fenologia e valor nutritivo de follajes de algunas espécies forrageiras de la caatinga. Agrofor. Am., 33-34:33-37, 2002.
- BALIEIRO, F.C.; DIAS, L.E.; FRANCO, A.A.; CAMPELLO, E.F.C. & FARIA, S.M. Acúmulo de nutrientes na parte aérea, na serapilheira acumulada sobre o solo e decomposição de filódios de Acacia mangium Willd. Ci. Flor., 14:59-65, 2004.
- BARCELLOS, A.D.O.; RAMOS, A.K.B.; VILELA, L. & MARTHA JUNIOR, G.B. Sustentabilidade da produção animal baseada em pastagens consorciadas e no emprego de leguminosas exclusivas, na forma de banco de proteína, nos trópicos brasileiros. R. Bras. Zootec., 37:51-67, 2008.
- BARRETOS, A. C. & FERNANDES, M. F. Cultivo de Gliricidia sepium e Leucaena leucocephala em alamedas visando a melhora dos solos dos tabuleiros costeiros. Pesq. Agropec. Bras., 36:1287-1293, 2001.
- BERTALOT, M.J.A.; GUERRINI, I.A.; MENDOZA, E.; DUBOC, E.; BARREIROS, R.M. & CORREIA, F.M. Retorno de nutrientes ao solo via deposição de serapilheira de quatro espécies leguminosas arbóreas na região de Botucatu-SP. Sci. For., 65:219-227, 2004.
- BEZERRA NETO, E. & BARRETO, L.P. Métodos de análises químicas em plantas. Recife, Universidade Federal Rural de Pernambuco, 2004. 148p.
- BOHNEN, A.; MEURER, E.J. & BISSANI, C.A. Solos ácidos e solos afetados por sais. In: MEURER, E.J., ed. Fundamentos de química do solo. 3.ed. Porto Alegre, Evangraf, 2006. 285p.
- BRAZ, A.J.B.P.; KLIEMANN, H.J. & SILVEIRA, P.M. Produção de fitomassa de espécies de cobertura em Latossolo Vermelho distrófico. Pesq. Agropec. Trop., 35:55-64, 2005.
- CASTELLANE, P.D. & FONTES, L.A. Alguns aspectos sobre capacidade de troca catiônica radicular. Seiva, 43:16-28, 1983.
- CAYUELA, M.L.; SINICCO, T. & MONDINI, C. Mineralization dynamics and biochemical properties during initial decomposition of plant and animal residues in soil. Appl. Soil Ecol., 41:118-127, 2009.
- COMPANHIA PERNAMBUCANA RECURSOS HÍDRICOS - CPRH. Diagnóstico sócio ambiental do Litoral Norte de Pernambuco. Recife, 2003. 214p.
- COSTA, G.S.; FRANCO, A.A.; DAMASCENO, R.N. & FARIA, S.M. Aporte de nutrientes pela serapilheira em uma área degradada e revegetada com leguminosas arboreas. R. Bras. Ci. Solo, 28:919-927, 2004.
- DIAS, P.F.; SOUTO, S.M.; RESENDE, A.S.; URQUIAGA, S.; ROCHA, G.P.; MOREIRA, J.F. & FRANCO, A.A. Transferência do N fixado por leguminosas arbóreas para o capim Survenola crescido em consórcio. Ci. Rural, 37:352-356, 2007.
- DUBEUX JR., J.C.B.; SOLLENBERGER, L.E.; MATHEWS, B.W.; SCHOLBERG, J.M. & SANTOS, H.Q. Nutrient cycling in warm-climate grasslands. Crop Sci., 47:915-928, 2007.
- DULORMNE, M.; SIERRA, J.; NYGREN, P. & CRUZ, P. Nitrogen-fixation dynamics in a cut-and-carry silvopastoral system in the subhumid conditions of Guadeloupe, French Antilles. Agrofor. Syst., 59:121-129, 2003.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Manual de análises químicas de solos. Brasília, 1999. 370p.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Sistema brasileiro de classificação de solos. 2.ed. Rio de Janeiro, Embrapa Solos, 2006. 306p.
- FERREIRA, R.L.C.; LIRA JUNIOR, M.A.; ROCHA, M.S.; SANTOS, M.V.F.; LIRA, M.A. & BARRETO, L.P. Deposição e acúmulo de matéria seca e nutrientes em serapilheira em um bosque de sabiá (Mimosa caesalpiniaefolia Benth.). R. Árvore, 31:7-12, 2007.
- FREIRE, J.L.; DUBEUX JÚNIOR, J.C.B.; LIRA, M.A.; FERREIRA, R.L.C.; SANTOS, M.V.F. & FREITAS, E.V. Deposição e composição química de serrapilheira em um bosque de sabiá. R. Bras. Zootec., 39:1650-1658, 2010.
- FREITAS, A.D.S.; SAMPAIO, E.V.B.S.; SANTOS, C.E.D.R.; SILVA, A.F.D. & LYRA, M.C.C.P. Nodulação em leguminosas da caatinga. In: CONGRESSO DE ECOLOGIA DO BRASIL, 7., Caxambu, 2005. Anais... Caxambu, SEB, 2005. CD ROM
- GAMA-RODRIGUES, A.C.D.; GAMA-RODRIGUES, E.F.D. & BRITO, E.C.D. Decomposição e liberação de nutrientes de resíduos culturais de plantas de cobertura em Argissolo Vermelho-Amarelo na região noroeste Fluminense (RJ). R. Bras. Ci. Solo, 31:1421-1428, 2007.
- GRAPPEGIA JÚNIOR, G.; AMADO, T.J.C. & PONTELLI, C.B. Acidificação do solo pelo uso prolongado de leguminosas como culturas de cobertura em sistemas de produção de milho. In: REUNIÃO SUL-BRASILEIRA DE CIÊNCIA DO SOLO, 3., Santa Maria, 2000, Anais... Santa Maria, SBCS, 2000. CD ROM
- HAKALA, K. & JAUHIAINEN, L. Yield and nitrogen concentration of above- and below-ground biomasses of red clover cultivars in pure stands and in mixtures with three grass species in northern Europe. Grass Forage Sci., 62:312-321, 2007.
- INSTITUTO AGRONÔMICO DE PERNAMBUCO - IPA. Banco de dados agrometeorológicos. Recife: 1994.100p.
- INSTITUTO DE TECNOLOGIA DE PERNAMBUCO - ITEP. Banco de dados meteorológicos de Pernambuco. Disponível em: <http://www.itep.br/itep_lamepe/site/index.php> Acesso em 10 jan. 2011.
- JACOMINE, P.K.T.; CAVALCANTI, A.C.; BURGOS, N.; PESSOA, S.C.P. & SILVEIRA, C.O. Levantamento exploratório reconhecimento de solos do Estado de Pernambuco. Recife, SUDENE, 1973. v.1. 359p. (Boletim Técnico 26, Série Pedologia, 14)
- KIMARO, A.A.; TIMMER, V.R.; CHAMSHAMA, S.A.O.; MUGASHA, A.G. & KIMARO, D.A. Differential response to tree fallows in rotational woodlot systems in semi-arid Tanzania: Post-fallow maize yield, nutrient uptake, and soil nutrients. Agric. Ecosyst. Environ., 125:73-83, 2008.
- LIMA, S.S.; LEITE, L.F.C.; OLIVEIRA, F.D.C. & DA COSTA, D.B. Atributos químicos e estoques de carbono e nitrogênio em Argissolo Vermelho-Amarelo sob sistemas agroflorestais e agricultura de corte e queima no norte do Piauí. R. Árvore, 35:51-60, 2011.
- LOPES, J.; EVANGELISTA, A.R.; FORTES, C.A.; PINTO, J.C.; FURTINI NETO, A. E. & SOUZA, R.M. Nodulação e produção de raízes do estilosantes mineirão sob efeito de calagem, silicatagem e doses de fósforo. Ci. Agrotec., 35:99-107, 2011.
- MACEDO, M.C.M. Integração lavoura e pecuária: o estado da arte e inovações tecnológicas. R. Bras. Zootec., 38:133-146, 2009.
- MACEDO, M.O.; RESENDE, A.S.; GARCIA, P.C.; BODDEY, R.M.; JANTALIA, C.P.; URQUIAGA, S.; CAMPELLO, E.F.C. & FRANCO, A.A. Changes in soil C and N stocks and nutrient dynamics 13 years after recovery of degraded land using leguminous nitrogen-fixing trees. For. Ecol. Manage., 255:1516-1524, 2008.
- MACHADO, F.A. Avaliação de pastagens de Brachiaria decumbens Stapf. consorciadas com leguminosas na Zona da Mata seca de Pernambuco. Recife, Universidade federal de Pernanbuco, 2011. 132p. (Tese de Doutorado)
- MACHARIA, P.N.; GACHENE, C.K.K.; MUREITHI, J.G.; KINYAMARIO, J.I.; EKAYA, W.N. & THURANIRA, E.G. The effect of introduced forage legumes on improvement of soil fertility in natural pastures of Semi-Arid rangelands of Kajiado District, Kenya. Trop. Subtrop. Agroecosyst., 14:221-227, 2011.
- MARSCHNER, H. Function of mineral nutrients: Macronutrients. In: MARSCHNER, H., ed. Mineral nutrition of higher plants. 2.ed. San Diego, Academic Press, 1995. p.229-312.
- MENEZES, R.S.C.; SALCEDO, I.H. & ELLIOTT, E.T. Microclimate and nutrient dynamics in a silvopastoral system of semiarid northeastern Brazil. Agrofor. Syst., 56:27-38, 2002.
- MOREIRA, J.N.; LIRA, M.A.; SANTOS, M.V.F.; FERREIRA, M.A.; ARAÚJO, G.G.L.; FERREIRA, R.L.C. & SILVA, G.C. Caracterização da vegetação de caatinga e da dieta de novilhos no Sertão de Pernambuco. Pesq. Agropec. Bras., 41:1643-1651, 2006.
- NASCIMENTO, J.T.; SILVA, I.F.; SANTIAGO, R.D. & SILVA NETO, L.F. Efeito de leguminosas nas características químicas e matéria orgânica de um solo degradado. R. Bras. Eng. Agríc. Amb., 7:457-462, 2003.
- OLIVEIRA, M.E.; LEITE, L.L. & CASTRO, L.H.R. Influência de árvores de baru (Dipteryx alata) e pequi (Caryocar brasiliense) no solo sob pastagem de braquiária. In: INTERNATIONAL SYMPOSIUM SOIL FUNCTIONING UNDER PASTURES IN INTERTROPICAL AREAS, 2000, Brasília. Anais... Brasília, Embrapa Cerrados/IRD, 2000. CD ROM.
- PALM, C.A. Contribution of agroforestry trees to nutrient requirements of intercropped plants. Agrofor. Syst., 30:105-124, 1995.
- PANDEY, D.N. Multifunctional agroforestry systems in India. Current Sci., 92:455-463, 2007.
- PEOPLES, M.B. & CRASWELL, E.T. Biological nitrogen fixation: Investments, expectations and actual contributions to agriculture. Plant Soil, 141:13-39, 1992.
- RODRIGUES, A.M.; CECATO, U.; FUKUMOTO, N.M.; GALBEIRO, S.; SANTOS, G.T. & BARBERO, L.M. Concentrações e quantidades de macronutrientes na excreção de animais em pastagem de capim-mombaça fertilizada com fontes de fósforo. R. Bras. Zootec., 37:990-997, 2008.
- SÁ, J.C.M. Manejo da fertilidade do solo no sistema plantio direto. In: SIQUEIRA, J.O.; MOREIRA, F.M.S.; LOPES, A.S.; GUILHERME, L.R.G.; FAQUIN, V.; FURTINI NETO, A.E. & CARVALHO, J.G., eds. Inter-relação fertilidade, biologia do solo e nutrição de plantas. Lavras, UFLA/SBCS, 1999. p.267-321.
- SANTOS, A.C.; SILVA, I.F.; LIMA, J.R.S.; ANDRADE, A.P. & CAVALCANTE, V.R. Gramíneas e leguminosas na recuperação de áreas degradadas: efeito nas características químicas de solo. R. Bras. Ci. Solo, 25:1063-1071, 2001.
- SANTOS, R.; SANTOS, M.; OLIVEIRA, I.P.; MORAIS, R.F.; URQUIAGA, S.C.; BODDEY, R.M. & ALVES, B.J.R. Componentes da parte aérea e raízes de pastagens de Brachiaria spp. em diferentes idades após a reforma, como indicadores de produtividade em ambiente de cerrado. Pesq. Agropec. Trop., 37:119-124, 2007.
- SAS Institute INC. The SAS System for Windows. Cary, 1999.
- SECRETARIA DE DEFESA AGROPECUÁRIA/MINISTÉRIO DA AGRICULTURA, PECUÁRIA E ABASTECIMENTO - SDA/MAPA. Instrução Normativa Nº10, de 21 de março de 2006. Diário Oficial da União - Seção 1, 2006.
- SILVA, G.S.; DUBEUX JUNIOR, J.C.B.; MELO, V.S.; LIRA JUNIOR, M.A.; NASCIMENTO, G.V.C.; MACHADO, F.A. & LIRA, M.A. Estabelecimento de leguminosas arbustivas em consórcio com pastagens de B. decumbens Stapf. In: JORNADA DE ENSINO, PESQUISA E EXTENSÃO, 9., Recife, 2009. Anais... Recife, Universidade Federal Rural de Penambuco, 2009. CD ROM
- SILVA, J.D. & QUEIROZ, A.C. Análise de alimentos: Métodos químicos e biológicos. 3.ed. Viçosa, MG, Universidade Federal de Viçosa, 2006. 235p.
- SILVA, J.O.; ROCHA, F.A. & SILVA, T.O. Determinação da CTC radicular de gramíneas e leguminosas forrageiras. Enciclopédia Biosfera, 6:1-7, 2010.
- TIESSEN, H.; MENEZES, R.S.C.; SALCEDO, I.H. & WICK, B. Organic matter transformations and soil fertility in a treed pasture in semiarid NE Brazil. Plant Soil, 252:195-205, 2003.
- van SOEST, P.J. Nutritional ecology of the rumimant. 2.ed. New York, Cornell University Press, 1994. 476p.
Datas de Publicação
-
Publicação nesta coleção
03 Jun 2013 -
Data do Fascículo
Abr 2013
Histórico
-
Recebido
02 Mar 2012 -
Aceito
30 Jan 2013