Resumos
O tegumento das sementes de soja é uma estrutura relacionada à capacidade de germinação, vigor e longevidade das sementes. Entre genótipos de soja existe variabilidade genética quanto aos caracteres do tegumento da semente, a qual pode ser utilizada em programas de melhoramento genético. Diante do exposto, realizou-se esse estudo com os objetivos de: Analisar a dissimilaridade genética de população segregante de soja para caracteres morfológicos de sementes, por meio de marcadores morfológicos e marcadores moleculares (AFLP, SSR e RAPD); Estimar a correlação entre as matrizes de dissimilaridade obtidas por meio da utilização das diferentes classes de marcadores. A população utilizada no estudo originou-se do cruzamento entre dois genótipos contrastantes para os caracteres do tegumento, a cultivar CD 202 (genitor feminino), com tegumento amarelo, e o genótipo TP (genitor masculino), com tegumento preto. Na geração F3, foram avaliados os caracteres morfológicos de cores do tegumento e do hilo, presença de derramamento de hilo, cor da pubescência e cor da flor. Os marcadores moleculares foram obtidos através das técnicas de AFLP, SSR e RAPD. De acordo com os resultados, os marcadores AFLP são os mais eficientes em acessar a variabilidade genética da população estudada, proporcionando a formação de maior número de grupos em relação aos demais marcadores. Apenas a matriz de dissimilaridade e o dendrograma obtido através de marcadores moleculares AFLP avaliados isoladamente apresentam correlação positiva com a matriz de dissimilaridade e o dendograma obtido por avaliação dos dados morfológicos.
Glycine max; tegumento da semente; marcadores moleculares
The soybean seed coat is a structure associated with seed germination, vigor and longevity. Soybean genotypes show genetic variability regarding the seed coat traits, which can be used in soybean breeding programs. The objectives of this study were: to analyze the genetic similarity of a segregant soybean population for morphological seed traits using morphological and molecular markers (AFLP, RAPD and SSR) and to establish the correlation between the dissimilarity matrices obtained using different marker classes. The population was obtained by crossing between two contrasting soybean genotypes: the cultivar CD 202 (female parent) with a yellow coat, and the genotype TP (male parent) with a black coat. The morphological traits evaluated were coat and hilum colors, hilum spread, pubescence and flower colors. Molecular markers were obtained by AFLP, RAPD and SSR techniques. Based on estimated similarity matrices, dendrograms were developed and the correlations between all matrix pairs and dendrograms estimated. The results showed that AFLP markers are more efficient for accessing the genetic variability of this population. Only the dissimilarity matrix and the dendrogram obtained by AFLP markers showed a positive correlation with the dissimilarity matrix and dendrogram obtained by morphological data.
Glycine max; seed coat; molecular markers
Dissimilaridade genética em população segregante de soja com variabilidade para caracteres morfológicos de semente
Soybean genetic dissimilarity in a segregant population with variability in seed morphological traits
Marisa DellagostinI; Fernando Augusto HenningII; Liliane Marcia MertzII; Mauricio Marini KoppIII; Maraisa CrestaniIV; Ivan SchusterI; Paulo Dejalma ZimmerII
ICooperativa Central de Pesquisa Agrícola, BR 467, Km 98, CEP 83813-450 Cascavel, PR. E-mail: marisa@coodetec.com.br; ivan@coodetec.com.br
IIPrograma de Pós-Graduação em Ciência e Tecnologia de Sementes - Depto de Fitotecnia, FAEM, UFPel. CP 354, CEP: 96010-900, Pelotas, RS. E-mail: fernandohenning@yahoo.com.br; lilianemertz@yahoo.com.br; djzimmer@ufpel.edu.br
IIIEmbrapa Gado de Leite, Rua Eugênio do Nascimento, no 610, Dom Bosco, CEP 36038-330 Juiz de Fora, MG. E-mail: kopp@cnpgl.embrapa.br
IVPrograma de Pós-Graduação em Agronomia - Área de concentração Fitomelhoramento, Depto de Fitotecnia, FAEM, UFPel. CP 354, CEP: 96010-900, Pelotas, RS. E-mail: maraisacrestani@yahoo.com.br
RESUMO
O tegumento das sementes de soja é uma estrutura relacionada à capacidade de germinação, vigor e longevidade das sementes. Entre genótipos de soja existe variabilidade genética quanto aos caracteres do tegumento da semente, a qual pode ser utilizada em programas de melhoramento genético. Diante do exposto, realizou-se esse estudo com os objetivos de: Analisar a dissimilaridade genética de população segregante de soja para caracteres morfológicos de sementes, por meio de marcadores morfológicos e marcadores moleculares (AFLP, SSR e RAPD); Estimar a correlação entre as matrizes de dissimilaridade obtidas por meio da utilização das diferentes classes de marcadores. A população utilizada no estudo originou-se do cruzamento entre dois genótipos contrastantes para os caracteres do tegumento, a cultivar CD 202 (genitor feminino), com tegumento amarelo, e o genótipo TP (genitor masculino), com tegumento preto. Na geração F3, foram avaliados os caracteres morfológicos de cores do tegumento e do hilo, presença de derramamento de hilo, cor da pubescência e cor da flor. Os marcadores moleculares foram obtidos através das técnicas de AFLP, SSR e RAPD. De acordo com os resultados, os marcadores AFLP são os mais eficientes em acessar a variabilidade genética da população estudada, proporcionando a formação de maior número de grupos em relação aos demais marcadores. Apenas a matriz de dissimilaridade e o dendrograma obtido através de marcadores moleculares AFLP avaliados isoladamente apresentam correlação positiva com a matriz de dissimilaridade e o dendograma obtido por avaliação dos dados morfológicos.
Termos para indexação:Glycine max, tegumento da semente, marcadores moleculares.
ABSTRACT
The soybean seed coat is a structure associated with seed germination, vigor and longevity. Soybean genotypes show genetic variability regarding the seed coat traits, which can be used in soybean breeding programs. The objectives of this study were: to analyze the genetic similarity of a segregant soybean population for morphological seed traits using morphological and molecular markers (AFLP, RAPD and SSR) and to establish the correlation between the dissimilarity matrices obtained using different marker classes. The population was obtained by crossing between two contrasting soybean genotypes: the cultivar CD 202 (female parent) with a yellow coat, and the genotype TP (male parent) with a black coat. The morphological traits evaluated were coat and hilum colors, hilum spread, pubescence and flower colors. Molecular markers were obtained by AFLP, RAPD and SSR techniques. Based on estimated similarity matrices, dendrograms were developed and the correlations between all matrix pairs and dendrograms estimated. The results showed that AFLP markers are more efficient for accessing the genetic variability of this population. Only the dissimilarity matrix and the dendrogram obtained by AFLP markers showed a positive correlation with the dissimilarity matrix and dendrogram obtained by morphological data.
Index terms:Glycine max, seed coat, molecular markers.
INTRODUÇÃO
O sucesso de um programa de melhoramento depende fundamentalmente da existência de variabilidade genética, a qual permite que os processos seletivos resultem em ganhos genéticos significativos (Bernardo, 2002). Entretanto, para que isso seja possível é necessária à utilização de técnicas direcionadas ao desenvolvimento de materiais genéticos superiores para os caracteres de interesse.
Na cultura da soja (Glycine max), a hibridização artificial tem sido o método mais utilizado e mais comum para ampliação de variabilidade genética intraespecífica (Baenziger et al., 2006). Esse processo é comumente usado por melhoristas visando à recombinação gênica entre diferentes genótipos para a obtenção de plantas que reúnam melhores características. O cruzamento entre indivíduos específicos proporciona a decomposição de caracteres genéticos complexos em seus componentes mendelianos, a localização das regiões genômicas envolvidas em seu controle, a quantificação do efeito destas regiões sobre o caráter em estudo e a canalização de toda esta informação para uso em programas de melhoramentos (Ferreira e Grattapaglia, 1998).
O tegumento das sementes de soja é um dos condicionantes da capacidade de germinação, do vigor e da longevidade das sementes. Grande parte das características do tegumento está associada a problemas específicos apresentados pelas sementes. A susceptibilidade a danos mecânicos está associada ao seu teor de lignina, enquanto que longevidade e potencial de deterioração no campo têm sido relacionados ao grau de permeabilidade do tegumento (Souza e Marcos Filho, 2001). Entre os genótipos de soja existe variabilidade genética quanto à qualidade de sementes, a qual pode ser utilizada em programas de melhoramento genético. Um exemplo é a variabilidade existente entre genótipos de soja com variação na coloração do tegumento. Tegumentos de sementes de soja de coloração preta apresentam menor velocidade de embebição, maior resistência à deterioração no campo, maior espessura, propriedades antifúngicas e maior conteúdo de lignina em relação a tegumentos de coloração clara (Peske e Pereira, 1983; Chachalis e Smith 2000; Mertz et al., 2009).
Em um programa de melhoramento genético é de fundamental importância o conhecimento do germoplasma disponível em termos de desempenho, capacidade de combinação e a dissimiliaridade genética (Vieira et al., 2005). A distância genética pode ser estimada a partir da utilização de caracteres fenotípicos, dados moleculares, coeficiente de parentesco e a combinação destes, afim de compreender a variabilidade existente na população (Kim e Ward, 1997; Bered et al., 2002).
Diante do exposto, os objetivos nesse estudo foram: Analisar a dissimilaridade genética de população segregante de soja para caracteres morfológicos de sementes, por meio de marcadores morfológicos e marcadores moleculares (AFLP, SSR e RAPD); Estimar a correlação entre as matrizes de dissimilaridade obtidas por meio da utilização das diferentes classes de marcadores.
MATERIAL E MÉTODOS
Nesse estudo foi utilizada uma população segregante de 94 indivíduos, e seus dois genitores, oriunda do cruzamento entre a cultivar CD 202 (genitor feminino) com tegumento amarelo, hilo marrom, pubescência cinza e flor branca, a qual apresenta tegumento com menor espessura, maior permeabilidade e baixos teores de lignina, e uma linhagem TP (genitor masculino) com tegumento preto, hilo preto, pubescência marrom e flor branca, a qual se caracteriza por apresentar tegumento com maior espessura, menor permeabilidade e maior quantidade de lignina. A linhagem TP originou-se a partir de sucessivas autofecundações de sementes atípicas, com tegumento de coloração preta, encontradas em lavoura de soja de cultivar de cultivar comercial com tegumento amarelo.
Avaliações dos parâmetros morfológicos
As plantas utilizadas nesse estudo apresentavam-se na geração F2, onde foram avaliados os caracteres de cor da pubescência e cor da flor. Nas sementes F3, oriundas das plantas F2, foram avaliados os caracteres de cor do tegumento, cor do hilo e a ocorrência de derramamento de hilo.
Obtenção dos Marcadores moleculares
Extração do DNA: O DNA genômico foi extraído a partir de plantas na geração F2, através do método Doyle e Doyle (1990). Para todas as classes de marcadores moleculares utilizadas nesse estudo, efetuou-se uma seleção prévia dos primers por meio de avaliação do polimorfismo entre os genitores masculino (TP) e feminino (CD 202).
Marcadores AFLP (Amplified Fragment Length Polymorphism): Foram utilizadas combinações de primers disponíveis no Kit AFLP Starter Primer (Invitrogen®). Foram obtidos 17 marcadores polimórficos entre os genitores utilizando as combinações (E-AGG/M-CAC); (E-ACC/M-CAG) (E-AAC/ M-CAG).
Marcadores RAPD (Random Amplification of Polymorphic DNA): foram utilizados os primers AA11; J11; A10; C04; C07a; C07b; F04; W02a; W02b.
Marcadores SSR Single Sequence Repeat (Microssatélites): foram utilizados os primers Satt 114; Satt 186; Satt 184; Satt 181; Satt 253; Satt 358; Satt 371; Satt 394; Satt 551; Satt 558; Satt 586; Satt 449; Satt 559; Satt 406; Satt 070; Satt 481; Satt 268; Satt 579; Satt 238; Satt 303; Satt 233; Satt 480; Satt 311; Satt 267; Satt 339; Satt 363; Satt 324; Satt 454; Satt 424; Satt 094; Satt 263; Satt 353; Satt 277; Satt 038; Satt 384; Satt 420; Satt 335 e Satt 066.
Análise da dissimilaridade genética
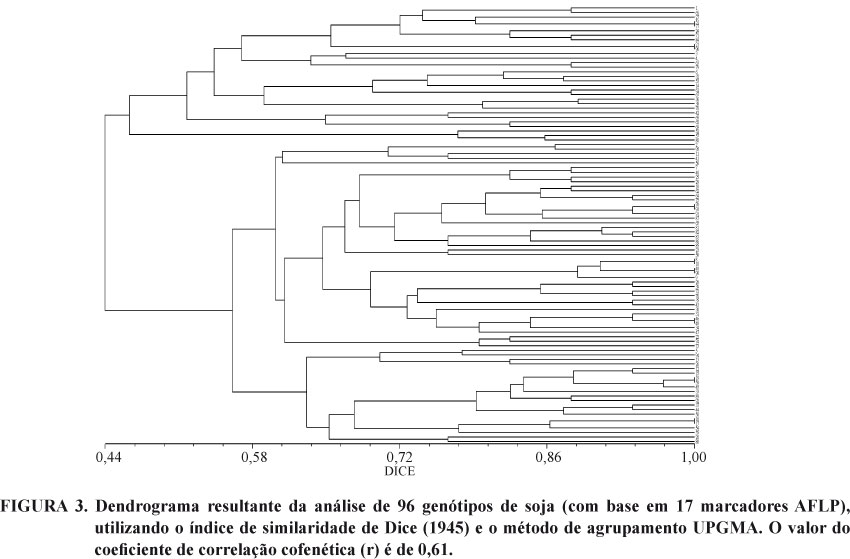
Com base nas avaliações morfológicas (caracteres qualitativos) foi estimada uma matriz de dissimilaridade através do índice "Simple Match" (Sokal e Sneath, 1963) entre todos os pares de genótipos. A partir dos resultados obtidos pelos marcadores moleculares foram obtidas matrizes de similaridade genética (Sg), utilizando o coeficiente de Dice (Dice, 1945). As matrizes de similaridade estimadas foram transformadas em matrizes de dissimilaridade a partir do complemento da similaridade estimada (Dg = 1 - Sg). Baseados nas matrizes de dissimilaridade gerados foram construídos dendrogramas, utilizando o método de agrupamento das médias das distâncias "Unweighted Pair Group Method with Arithmetic Mean" (UPGMA). Para a verificação do ajuste entre a matriz de dissimilaridade e o dendrograma, foi calculado o coeficiente de correlação cofenética (r), conforme (Sokal e Rohlf 1962).
A seguir foram estimadas as correlações entre todos os pares de matrizes obtidas (AFLP, SSR, RAPD, conjunta de marcadores moleculares e morfológica) e entre todos os dendrogramas obtidos de cada um destes conjuntos de dados utilizando o teste de comparação de matrizes de Mantel, com 1000 permutações (Mantel, 1967) e significância testada pelo teste t. Todas as análises foram realizadas com auxílio do programa computacional NTSYS pc 2.1 (Rohlf, 2000).
RESULTADOS E DISCUSSÃO
O cruzamento entre a cultivar CD 202 e o genótipo TP, resultou em uma população F2 com variação nos padrões de coloração da pubescência e com sementes F3 com diferenças para coloração do tegumento e do hilo (Tabela 1). No que se refere à coloração do tegumento, foram encontradas sementes de cor preta, amarela e camurça (Figura 1). Em soja, três locos independentes (I, R e T) controlam a pigmentação do tegumento das sementes. O loco I controla a distribuição de pigmentos, antocianinas e proantocianinas e corresponde a uma região multigênica contendo genes da família chalcone synthase (CHS). O alelo dominante I inibe a pigmentação fazendo com que o tegumento apresente coloração amarela enquanto que o alelo recessivo i promove a pigmentação das sementes. O alelo recessivo i permite a coloração do tegumento da semente por completo enquanto que dois outros alelos, ii e ik restringem a pigmentação à região do hilo (ii) e pigmentação do hilo e parte do tegumento (ik) (Todd e Vodkin, 1996). A interação dos alelos dos locos R e T que também controlam a cor de pubescência são responsáveis pelas colorações preta, marrom e camurça do tegumento e do hilo. Dessa forma tegumentos de sementes de soja com coloração podem ser preto (i, R, T), preto imperfeito (i, R, t), marrom (i, r, T) ou (camurça) (i, r, t) (Zabala e Vodkin, 2003).
Com relação à coloração do hilo, as sementes apresentaram hilo marrom ou preto, sendo que entre as sementes com hilo preto, algumas apresentaram o caráter de hilo esparramado (Tabela 1). Trabalho conduzido por Carpentieri-Pípolo et al. (2007), demonstra que em sementes de soja, o caráter hilo normal é dominante sobre a presença de hilo esparramado, e não há influência de efeito materno sobre a expressão genética desse caráter. Além disso, os autores afirmam que a presença do hilo esparramado é condicionada por um gene recessivo, e sua expressão é dependente do loco T, que controla a cor da pubescência, pois somente ocorre em genótipos com pubescência marrom (T_). Portanto, o alelo T é pleiotrópico, em relação ao alelo que condiciona hilo esparramado.
No que se refere à coloração da pubescência foram encontradas as cores marrom e cinza (Tabela 1). Dois locos são responsáveis pela coloração da pubescência em plantas de soja, sendo eles o loco T e o Td, podendo apresentar as seguintes combinações alelicas TT TdTd, marrom; TT tdtd, marrom claro e tt TdTd ou tt tdtd, cinza (Iwashina et al., 2006).
Com relação à cor da flor, todos os genótipos avaliados apresentaram flores com coloração branca (Tabela 1). A cor roxa e a cor branca são as mais comuns entre a maioria das cultivares de soja, entretanto, alguns genótipos apresentam variações. O gene W tem efeito pleiotrópico para cor da flor e cor do hipocótilo (Takahashi e Fukuyama, 1919), sendo que, o alelo dominante origina flores com coloração roxa (W1W1), enquanto que o recessivo flores com coloração branca (w1w1).
Em se tratando de marcadores moleculares, o número de marcadores polimórficos foi de 17 para os marcadores AFLP, 38 para os marcadores SSR e nove para os marcadores RAPD, totalizado 64 marcadores moleculares. Os três pares de primers utilizados na técnica de AFLP resultaram em 17 marcadores polimórficos. Esse número foi bem inferior ao obtido em estudos com outras espécies como a aveia por exemplo, onde Vieira et al. (2005), obtiveram até 53 marcadores polimórficos em uma única combinação de primer. Em trabalho com trigo, o número de marcadores polimórficos obtidos em cada combinação de primer variou entre 36 a 52 (Vieira et al., 2007). Esse baixo polimorfismo encontrado na soja pode ser atribuído a estreita base genética da espécie, quando comparada as poáceas, por exemplo, além disso, deve-se considerar que o trigo é uma espécie poliploide.
O dendograma obtido a partir da matriz de dissimilaridade gerada a partir dos caracteres morfológicos possibilitou a formação de seis grupos distintos, sendo 18 indivíduos atribuídos ao grupo 1, 28 ao grupo 2, 29 ao grupo 3, 11 ao grupo 4, quatro ao grupo 5 e quatro ao grupo 6 (Figura 2, Tabela 1). Os indivíduos que mais divergiram foram os integrantes do grupo 1, formado por sementes com tegumento preto, hilo preto e pubescência marrom, e os do grupo 6, formado por sementes com tegumento camurça, hilo marrom e pubescência cinza.
Com relação aos dendogramas obtidos a partir dos marcadores AFLP, RAPD, SSR e o dendograma obtido através da associação entre os diferentes marcadores, o número de grupos formados foi bem mais elevado em relação ao dendograma obtido pelos marcadores morfológicos (Figuras 3, 4, 5 e 6 respectivamente). Além disso, as distâncias estimadas a partir da utilização de dados moleculares, com exceção dos marcadores AFLP, se correlacionam negativamente com os resultados obtidos a partir dos marcadores morfológicos (Tabela 2). Essa correlação negativa pode ser atribuída ao pequeno número de marcadores utilizados, considerando que muitos desses marcadores podem estar associados a regiões intergênicas. Uma alternativa para tentar contornar esse tipo de problema, seria a seleção prévia dos marcadores moleculares através de análise in silico. Em trabalho de prospecção de marcadores SSR através de analise in silico, Henning et al. (2009) sugerem que a estratégia da mineração de marcadores moleculares a partir da integração in silico de sequências de marcadores moleculares ainda anônimos, bancos de dados genômicos e bancos contendo seqüências com descrições funcionais dos genes demonstra ser promissora, pois, possibilita prever as funções para estes genes e verificar a associação destes com rotas bioquímicas e metabólicas responsáveis pelas características que se deseja analisar.
Analisando o número de grupos formados nos dendogramas obtidos em cada uma das classes de marcadores moleculares, os marcadores AFLP foram os que resultaram na formação de maior número de grupos, seguida pelos marcadores RAPD e por último os marcadores SSR (Figuras 3, 4 e 5). Levando em consideração ainda que o número de marcadores AFLP foi inferior aos SSR, podemos afirmar que os marcadores AFLP pois foram os mais eficientes em acessar a variabilidade genética da população.
No que se refere ao coeficiente de correlação cofenética (r) dos dendrogramas obtidos, o melhor ajuste entre a representação gráfica das distâncias e a sua matriz original foi observado para o dendograma obtido a partir dos dados morfológicos (r=0,94), seguido dos marcadores SSR (r=0,74), dos marcadores RAPD (r=0,72), e por último dos marcadores AFLP (r=0,61). O coeficiente de correlação obtido no dendograma gerado a partir da associação entre todos os marcadores moleculares resultou em valor de correlação igual a 0,72). Um bom ajuste entre a representação gráfica das distâncias e a sua matriz original, possibilita a realização de inferências por meio da avaliação visual (Rohlf, 2000). Existem casos em que a estrutura de agrupamento dos genótipos avaliados não atende a pressuposição hierárquica, por consequência, outra forma de representação deverá ser adotada, mesmo que sejam os mais recomendados para aplicação em dados biológicos (Everitt, 1993). Em termos de melhoramento genético, estudos indicam que o método hierárquico da distância média (UPGMA) é superior ao vizinho mais próximo e mais distante quando comparados com informações conhecidas da genealogia (Romesburg, 1984). Nesse caso, pode-se afirmar que de maneira geral, os dendogramas obtidos nesse estudo apresentaram valores de coeficiente de correlação cofenética adequados, pois para todas as classes de marcadores utilizadas o valor foi próximo ou superior a 70.
CONCLUSÕES
Os marcadores AFLP são os mais eficientes em acessar a variabilidade genética da população estudada, proporcionando a formação de maior número de grupos em relação aos demais marcadores.
Apenas a matriz de dissimilaridade e o dendograma obtidos através de marcadores moleculares AFLP avaliados isoladamente apresentam correlação positiva com a matriz de dissimilaridade e o dendrograma obtido por avaliação dos dados morfológicos.
AGRADECIMENTOS
Os autores agradecem à COODETEC, CAPES, CNPq e FAPERGS pelo auxilio financeiro e concessão das bolsas de estudo.
Submetido em 12/09/2010.
Aceito para publicação em 21/04/11.
- BAENZIGER, P.S.; RUSSELL, W.K.; GRAEF, G.L.; CAMPBELL, B.T. Improving lives: 50 years of crop breeding, genetics, and cytology (C-1). Crop Science, v.46, p.2230-2244, 2006.
- BERNARDO, R. Breeding for quantitative traits in plants Woodbury: Stemma Press, 2002, 360p.
- BERED, F.; BARBOSA-NETO, J.F.; CARVALHO, F.I.F. Genetic variability in common wheat germplasm based on coefficients of parentage. Genetics and Molecular Biology, v.25, n.2, p.211-215, 2002.
- CARPENTIERI-PÍPOLO, V.; SOUZA, A.; ALMEIDA, L.A.; KIIHL, R.A.S.; PÍPOLO, A.E. Herança de hilo esparramado em sementes de soja. Pesquisa Agropecuária Brasileira, v.42, n.1, p.127-129, 2007.
- CHACHALIS, D.; SMITH, M.L. Imbibition behavior of soybean (Glycine max (L.) Merril) accessions with different testa characteristics. Seed Science andTechnology, v.28, n.2, p.321-331, 2000.
- DICE, L. R. Measures of the amount of ecologic association between species. Ecology, v.26, n.3, p.297-302, 1945.
- DOYLE, J.J.; DOYLE, J.L. Isolation of plant DNA from fresh tissue. Focus, v.12, p.13-15, 1990.
- EVERITT, B.S. Cluster analysis. Cambridge, University Press, 1993. 170p.
- FERREIRA, M.E.; GRATTAPAGLIA, D. Introdução ao uso de marcadores moleculares em análise genética. Brasília: EMBRAPA, CENARGEM, 1998, p.220.
- HENINNG, F.A.; MAIA, L.C.; MERTZ, L.M.; ZIMMER, P.D.; OLIVEIRA, A.C. Predição in silico de marcadores microssatélites relacionados ao tegumento de sementes de soja. Revista Brasileira de Sementes, v.31, n.4, p.49-56, 2009.
- IWASHINA, T.; BENITEZ, E.R.; TAKAHASHI, R. Analysis of flavonoids in pubescence of soybean near-isogenic lines for pubescence color loci. Journal of Heredity, v.97, p.438-443, 2006.
- KIM, H.S.; WARD, R.W. Genetic diversity in Eastern U.S. soft winter wheat (Triticum aestivum L. em Thell.) based on RFLPs and coefficient of parentage. Theoretical and Applied Genetics, v.94, n.3-4, p.472-479, 1997.
- MANTEL, N. The detection of disease clustering and a generalized regression approach. Cancer Research, v.27, n.2, p.209-220, 1967.
- MERTZ, L.M.; HENNING, F.A.; CRUZ, H.L.; MENEGHELLO, G.E.; FERRARI, C.S.; ZIMMER, P.D. Diferenças estruturais entre tegumentos de sementes de soja com permeabilidade contrastante. Revista Brasileira de Sementes, v.31, n.1, p.23-29, 2009.
- PESKE, S.T.; PEREIRA, L.A.G. Tegumento da semente de soja. Tecnologia de Sementes, v.6, p.23-34, 1983.
- ROHLF, F.J. NTSYS-pc: numerical taxonomy and multivariate analysis system, version 2.1. Exeter Software, New York, 2000.
- ROMESBURG, H.C. Cluster analysis for researchers. Califórnia, Lifetime Learning, 1984. 334p.
- SOKAL, R.R.; ROHLF, F.J. The comparison of dendrograms by objective methods. Taxonomy, v.11, n.1, p.30-40, 1962. 359p.
- SOKAL, R.R. SNEATH, P.H.A. Principles of Numeric Taxonomy W.H. Freeman, San Francisco, 1963. 359p.
- SOUZA, F.H.D.; MARCOS-FILHO, J. The seed coat as a modulator of seed-environment relationships in Fabaceae. Revista Brasileira de Botânica, v.24, n.4, p.365-375, 2001.
- TAKAHASHI, Y.; FUKUYAMA, J. Morphological and genetic studies on soybean (in Japanese). Hokkaido Agr Exp Stn Rep v.10, p.1-100, 1919.
- TODD, J.J.; VODKIN, L.O. Duplications that suppress and deletions that restore expression from a chalcone synthase multigene family. Plant Cell, v.8, n.4, p.687-699, 1996.
- VIEIRA, E.A.; CARVALHO, F.I.F; OLIVEIRA, A.C.; BENIN, G.; ZIMMER, P.D.; SILVA, J.A.G.; MARTINS, A.F.; BERTAN, E.; SILVA, G.O.; SCHMIDT, D.A.M. Comparação entre medidas de distância genealógica, morfológica e molecular em aveia em experimentos com e sem a aplicação de fungicida. Bragantia, v.64, n.1, p.51-60, 2005.
- VIEIRA, E.A.; CARVALHO, F.I.F.; OLIVEIRA, A.C.; MARTINS, L.F.; BENIN, G.; SILVA, J.A.G; KOPP, M.M.; HARTWIG, I.; CARVALHO, M.F.; VALÉRIO, I.P. Associação da distância genética em trigo estimada a partir de caracteres morfológicos, caracteres fenológicos e dos componentes do rendimento de grãos. Revista Brasileira de Agrociência, v.13, n.2, p.161-168, 2007.
- ZABALA, G.Z.; VODKIN, L.O. Cloning of the pleiotropic T locus in soybean and two recessive alleles that differentially affect structure and expression of the encoded flavonoid 3' hydroxylase. Genetics, v.163, n.1, p.295-309, 2003.
Datas de Publicação
-
Publicação nesta coleção
06 Jan 2012 -
Data do Fascículo
2011
Histórico
-
Aceito
21 Abr 2011 -
Recebido
12 Set 2010