
RESUMO
As florestas de areias brancas quartzosas constituem um tipo de ecossistema oligotrófico, cujos caracteres fitofisionômicos são afetados pela sazonalidade climática. Entre tais características está a abertura do dossel, cujas variações influenciam na distribuição espacial da luz solar no ambiente. Este estudo avaliou e comparou a estrutura da vegetação e as alterações sazonais do dossel em habitat de areias brancas do Parque Nacional Serra de Itabaiana, Sergipe. Os dados foram coletados em dois ambientes: vegetação aberta de areias brancas strictu sensu (VA) e mata fechada de vegetação ripária (MR). Para estimar a densidade arbórea utilizou-se o método quadrante, no qual foram amostradas árvores com CAP ≥ 5 cm. Para avaliar a dinâmica do dossel foram estabelecidos, em cada área, dois transectos compostos por cinco pontos equidistantes 20 m, totalizando 10 pontos por ambiente. Em cada ponto foram obtidas fotografias hemisféricas do dossel nos meses de janeiro (seca) e junho (chuva). As imagens foram analisadas pelo software Gap Light Analyzer, sendo avaliadas as variáveis abertura do dossel e índice de área foliar (IAF). A vegetação ripária apresentou densidade arbórea de 498 ind ha-1 e a vegetação aberta 296 ind ha-1. Esta última, não evidenciou variações na abertura do dossel (t = 2,0146; p > 0,05) e no IAF (t = -2,0052; p > 0,05) entre os períodos de chuva (52,24% ± 18,13% e 0,68 ± 0,45) e seca (67,39% ± 15,38% e 0,36 ± 0,23). Na mata ripária, a abertura do dossel (t = 7,1739; p < 0,05) e o IAF (t = -5,1609; p < 0,05) variou entre os períodos de chuva (13,70% ± 2,24% e 2,13 ± 0,23) e seca (21,08% ± 2,36% e 1,63 ± 0,20). As diferenças observadas destacam a heterogeneidade ambiental na Serra de Itabaiana e demonstram que as formações florestais de areias bancas respondem de maneira distinta à sazonalidade hídrica deste ecossistema. Por estar localizada em uma área de ecótono entre floresta perene e Floresta Estacional Semidecidual, a mata ripária tende a perder parte de suas folhas durante o período seco. Além disso, o padrão de distribuição das plantas pode contribuir para as diferenças entre os habitat estudados.
Palavras-chave:
abertura do dossel; fotos hemisféricas; índice de área foliar; vegetação aberta; mata ripária
ABSTRACT
The forest formations on quartz white-sand are a kind of oligotrophic ecosystem and its phytophysionomical characteristics are affected by the climate seasonality. The canopy is one of these characteristics, its variations may influence the spatial distribution of the energy in the environment. The objective of this research was to evaluate and compare the tree density and the seasonality change on the canopy openness on habitats of white-sand in Serra de Itabaiana National Park. The data were collected in two environments: gaping vegetation of white-sands strictu sensu (VA) and closed vegetation of riparian forest (MR). To estimate tree density, it was utilized the wandering-quarter, where trees with CAP > 5cm were sampled. To evaluate the canopy dynamics, the transects had 5 points equidistant 20m, ten per environment. Hemispherical pictures were taken in January (dry season) and June (wet season), utilizing a digital camera with a fisheye lens. The images were analyzed on the Gap Light Analyzer software. The results showed that riparian forest have higher tree density (498 ind ha-1) than the opened (296 ind ha-1). The letter do not showed variations in the canopy openness (t = -2,0052; p > 0,05) or in Leaf Area Index (t = -2,0052; p > 0.05) between wet (52,24% ± 18,13% e 0,68 ± 0,45) and dry seasons (67,39% ± 15,38% e 0,36 ± 0,23). In riparian forest, the canopy openness (t = 7,1739; p < 0.05) and LAI values (t = -5,1609; p < 0.05) vary between wet (13,70% ± 2,24% e 2,13 ± 0,23) and dry (21,08% ± 2,36% e 1,63 ± 0,20) seasons showing the seasonality. Differences between the two habitats highlights the environmental heterogeneity in Serra de Itabaiana and show that forest formations on white-sand respond by different ways to the water seasonality of this ecosystem. Because it is located in an ecotone area between evergreen forest and forest Semideciduous, the riparian forest tends to lose some of their leaves during the dry season. Moreover, the distribution pattern of the plants can contribute to the differences between the studied habitats.
Keywords:
opening the canopy; hemispheric photos; leaf area index; gaping vegetation; riparian forest.
INTRODUÇÃO
A comunidade vegetal que cobre as areias brancas quartzosas constitui ecossistema oligotrófico, cuja ciclagem dos nutrientes no solo é fortemente afetada pelo clima (MENDONÇA et al., 2013MENDONÇA, B. A. F. et al. Solos e Geoambientes do Parque Nacional do Viruá e entorno, Roraima: visão integrada da paisagem e serviço ambiental. Ciência Florestal, Santa Maria, v. 23, p. 429-444, 2013.). A vegetação nesse tipo de ambiente é caracterizada por apresentar dossel pouco denso, folhas espessas, escleromórficas e de coloração verde-pálida, resultando em paisagem de exceção quando em meio à matriz florestal (OLIVEIRA et al., 2001OLIVEIRA, A. A. et al. Florestas sobre areia: Campinaranas e Igapós. In: OLIVEIRA, A. A.; DALY, D. C. (Eds.). Florestas do rio Negro. São Paulo: Schwarcz, 2001. p. 179-220.; MENDONÇA et al., 2014). Tais habitats também são encontrados na Amazônia, especialmente na região do alto Rio Negro, sendo popularmente conhecidos como Caatingas amazônicas ou Campinaranas (OLIVEIRA et al., 2001), bem como em domínios de Mata Atlântica, desde o Estado de Sergipe até o Rio Grande do Norte (A.S. Ribeiro, com. pess.). Fora do Brasil, as formações vegetais sobre areis brancas estendem-se sobre territórios da Colômbia, Venezuela, Guianas e até na Ásia Campinaranas (OLIVEIRA et al., 2001). As areias brancas nos Domos de Serras têm sua origem da própria dissolução das rochas metamórficas, enquanto as formações de areias brancas nos tabuleiros têm origem de solos podzólicos, como nas Campinaranas da Amazônia Central (EMBRAPA, 1975; OLIVEIRA et al., 2001).
As características fisionômicas da vegetação nas areias brancas são influenciadas por ciclos sazonais e por alterações nos níveis do lençol freático, de modo que gradientes fitofisionômicos ocorrem associados a diferentes condições hidrológicas do solo (MENDONÇA et al., 2013MENDONÇA, B. A. F. et al. Solos e Geoambientes do Parque Nacional do Viruá e entorno, Roraima: visão integrada da paisagem e serviço ambiental. Ciência Florestal, Santa Maria, v. 23, p. 429-444, 2013.). Além disso, as florestas de areias brancas estão expostas às condições edáficas extremas, que resultam em limitações nutricionais e deficit hídricos periódicos nos solos arenosos (ANDERSON, 1981ANDERSON, A. B. White-Sand Vegetation of Brazilian Amazonia. Biotropica, Zurich, v. 13, n. 3, p. 199-210, 1981.). A natureza oligotrófica desses solos é resultado em grande parte das suas origens, que podem incluir o intemperismo de materiais como arenitos, quartzitos e granitos ou podzolização devido à flutuação do lençol freático que suprime a matéria orgânica resultando em uma areia degradada (ANDERSON, 1981).
Em resposta ao estresse hídrico e nutricional, a vegetação lenhosa das areias brancas apresenta algumas adaptações, como esclerofilia, folhas pequenas e inclinadas (ANDERSON, 1981ANDERSON, A. B. White-Sand Vegetation of Brazilian Amazonia. Biotropica, Zurich, v. 13, n. 3, p. 199-210, 1981.). Do mesmo modo, copas reduzidas e poucas árvores emergentes permitem ao sub-bosque maior exposição à radiação solar (OLIVEIRA et al., 2001OLIVEIRA, A. A. et al. Florestas sobre areia: Campinaranas e Igapós. In: OLIVEIRA, A. A.; DALY, D. C. (Eds.). Florestas do rio Negro. São Paulo: Schwarcz, 2001. p. 179-220.). A luz é fonte essencial e direta de energia para o desenvolvimento das plantas, sendo que diferença na disponibilidade desse recurso influencia a distribuição local das espécies e condiciona grande parte dos processos de crescimento dos vegetais (NICROTA; CHAZDON; IRIARTE, 1999; ZUQUIM; COSTA; PRADO, 2007ZUQUIM, G.; COSTA, F. R. C.; PRADO, J. Fatores que determinam a distribuição de espécies de pteridófitas da Amazônia Central. Revista Brasileira de Biociências , Porto Alegre, v. 5, n. 2, p. 360-362, 2007.). Um dos fatores que influencia na distribuição espacial da energia no ambiente é a dinâmica de abertura e fechamento do dossel, que promove alterações na intensidade e qualidade da luz e suas variáveis associadas (VAN DEN BERG; SANTOS, 2003VAN DEN BERG, E.; SANTOS, F. A. M. Aspectos da variação ambiental em uma floresta de galeria em Itutinga, MG, Brasil. Ciência Florestal , Santa Maria, v. 13, n. 2, p. 83-98, 2003.). Neste caso, a estrutura do dossel pode apresentar variações tanto espacial quanto temporalmente, contribuindo para formação heterogênea do sub-bosque (GARCIA et al., 2007GARCIA, L. C. et al. Heterogeneidade do dossel e quantidade de luz no recrutamento do sub-bosque de uma mata ciliar no Alto São Francisco, Minas Gerais: análise através de fotos hemisféricas. Revista Brasileira de Biociências, Porto Alegre, v. 5, n. 2, p. 99-101, 2007.). Tais variações podem estar relacionadas à complexidade estrutural do ambiente e a fatores como, diferentes estágios sucessionais, deciduidade da copa e abertura de clareiras (BIANCHINI; PIMENTA; SANTOS, 2001BIANCHINI, E.; PIMENTA, J. A.; SANTOS, F. A. M. Spatial and temporal variation in the canopy cover in a tropical semi-deciduous forest. Brazilian Archives Biology Technology, Curitiba, v. 44, n. 3, p. 269- 276, 2001.).
Embora as vegetações abertas e fechadas das areias brancas no Parque Nacional Serra de Itabaiana (PNSI), localizado na zona de transição entre os biomas Mata Atlântica e Caatinga sejam distintas, elas estão sob as mesmas condições edáficas e macroclimáticas. Neste sentido, este estudo teve como objetivo avaliar a estrutura da vegetação arbórea, bem como caracterizar e comparar a abertura do dossel e o índice de área foliar (IAF) em habitat de areias brancas localizados no PNSI, e avaliar alterações sazonais nesses parâmetros dentro e entre os ambientes estudados. Tais avaliações contribuirão para a melhor compreensão das variações sazonais em comunidades vegetais e ecossistemas oligotróficos.
MATERIAIS E MÉTODOS
Área de estudo
O estudo foi realizado no Parque Nacional Serra de Itabaiana - PNSI (10º40’S e 37º25’O), localizado entre os municípios sergipanos de Areia Branca, Itabaiana, Laranjeiras, Itaporanga D’ajuda, Malhador e Campo do Brito (Figura 1). O PNSI abrange área de 7.966 ha, possui elevação máxima de 670 m de altitude e está localizado na zona de transição entre a Mata Atlântica e a Caatinga (CARVALHO; VILAR, 2005CARVALHO, C. M.; VILAR, J. C. Parque Nacional Serra de Itabaiana: levantamento da Biota. São Cristóvão: UFS, 2005. 124 p. (Biologia Geral e Experimental - UFS).). Segundo o sistema de classificação de Köppen, o clima é do tipo As com verão seco (ALVARES et al., 2013ALVARES, C. A. et al. Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift, Stuttgart, v. 22, n. 6, p. 711-728, 2013.). Na região, o período chuvoso compreende os meses de março a agosto e o período da seca entre setembro e fevereiro (DISTRITO FEDERAL, 2012) e a pluviosidade média anual é de 1.100 mm (DANTAS; RIBEIRO, 2010DANTAS, T. V. P. et al. Florística e estrutura da vegetação arbustivo-arbórea das Areias Brancas do Parque Nacional Serra de Itabaiana/Sergipe, Brasil. Revista Brasileira de Botânica, São Paulo, v. 33, p. 575-588, 2010.). As temperaturas mínimas ocorrem entre junho e julho (17ºC a 18ºC) e a média varia com a altitude, sendo 22ºC nos pontos mais baixos e 17ºC nos mais elevados (BEZERRA; FARIAS; CAVALCANTI, 2010BEZERRA, M. F. A.; FARIAS, G. R.; CAVALCANTI, L. H. Mixobiota do Parque Nacional Serra de Itabaiana, SE, Brasil: Trichiales. Acta Botância Brasilica, Belo Horizonte, v. 24, n. 2, p. 510-517, 2010.). Durante o estudo, as temperaturas mínimas e máximas variaram em média de 23,7ºC a 35,2ºC no período seco e de 18,2ºC a 32,5ºC no período chuvoso, enquanto que a pluviosidade acumulada foi de 811 mm no período chuvoso e 159 mm no período seco (DISTRITO FEDERAL, 2012). A Serra de Itabaiana é caracterizada por apresentar mosaico de fitofisionomias, incluindo formações florestais (matas primárias e secundárias), formações abertas naturais (campos graminosos) e áreas antropizadas (capoeiras, áreas agrícolas e solos expostos) (DANTAS; RIBEIRO, 2010). O estudo foi desenvolvido em duas fisionomias: vegetação aberta de areias brancas strictu sensu (VA) e mata fechada de vegetação ciliar (chamada neste estudo de mata ripária, MR).
A vegetação aberta de areias brancas abrange área de 347 ha no PNSI, cuja fisionomia esclerófila com substrato arenoso-quartzoso forma um habitat com árvores agrupadas, associadas a bromélias e cactos, e está localizada na face leste do domo da Serra de Itabaiana (10°45’8,36” S e 037°20’15,67” W). Pode ser caracterizada por apresentar solos sedimentares arenosos de cor branca predominantemente exposta e pobre em nutrientes, cuja principal característica da vegetação é a disposição agregada dos indivíduos (DANTAS; RIBEIRO, 2010DANTAS, T. V. P. et al. Florística e estrutura da vegetação arbustivo-arbórea das Areias Brancas do Parque Nacional Serra de Itabaiana/Sergipe, Brasil. Revista Brasileira de Botânica, São Paulo, v. 33, p. 575-588, 2010.). O estrato arbóreo é pouco denso e baixo podendo chegar a 6 m e as espécies mais comuns são Acosmium bijugum (Vogel) Yakovlev (Fabaceae), Kielmeyera rugosa Choisy (Clusiaseae) e Agarista revoluta (Spreng.) Hook. F. ex Nied. (Ericaceae) (DANTAS et al., 2010). O estrato herbáceo é mais abundante e ocorre de forma dispersa entre os arbustos (DANTAS; RIBEIRO, 2010).
A mata ripária compreende um continuum no riacho Coqueiro, está localizada na encosta sul do domo da Serra de Itabaiana (10°45’59,10” S e 037°20’23,60” W), tem uma extensão de aproximadamente de 150 m, cuja altura do dossel varia de oito a 10 m, na cabeceira e 25 m na jusante, ocorrendo também árvores de maior porte (VICENTE et al., 2005VICENTE, A. et al. Levantamento Botânico. In: CARVALHO, C. M.; VILAR, J. C. (Coord.). Parque Nacional Serra de Itabaiana: levantamento da Biota . São Cristóvão: UFS , 2005. p.15-37. (Biologia Geral e Experimental - UFS).). Ocorrem epífitas, cipós e trepadeiras e a serapilheira é mais densa que outras áreas de vegetação arbórea (VICENTE et al., 2005). As espécies mais abundantes neste ambiente são Sclerolobium densiflorum Benth (Fabaceae), Tapirira guianensis Aubl. (Anacardiaceae), Hymatanthus articulatus (Vahl) Woodson (Apocynaceae), Eschweilera ovata Miers (Lecythidaceae) e Byrsonima sericea DC (Malpighiaceae) (VICENTE et al., 2005).

Parque Nacional Serra de Itabaiana pontos indicam os locais de estudo; areias brancas strictu sensu (VA) e da mata ripária (MR).
FIGURE 1:
Serra de Itabaiana National Park. The points show the study sites; white-sand sensu strictu (VA) and de riparian forest (MR).
Coleta de dados
Para estimar a densidade arbórea utilizou-se método baseado em distâncias, denominado quadrante errante, ou quadrante móvel (CATANA JUNIOR; ANTONY, 1963CATANA JUNIOR, J.; ANTHONY, J. The wandering quarter meted of estimating population density. Ecology, New York, v. 44, n. 2, p. 349-360, 1963.). Esse método é uma variação do ponto quadrante e consiste em amostrar o indivíduo que se encontra mais próximo do vértice (BROWER; ZAR; VON ENDE, 1998BROWER, J. E.; ZAR, J. H.; VON ENDE, C. N. Field and laboratory methods for general ecology, 4th ed. Boston: WCB McGraw-Hill, 1998. 273 p.). Este indivíduo amostrado, por sua vez torna-se o vértice para o ponto seguinte e assim sucessivamente. Foram mensurados os indivíduos mais próximos do vértice com circunferência a altura do peito (CAP = 1,30) ≥ 5 cm, anotando-se também a distância entre a planta e o vértice e a altura total da planta. O diâmetro a altura do peito (DAP) foi calculado pela fórmula DAP = CAP/π. O cálculo da densidade total (DT) teve como base a equação desenvolvida por Pollard (1971POLLARD, J. H. On distance estimators of density in randomly distributed forests. Biometrics, Malden, v. 27, n. 4, p. 991-1002, 1971.):
Em que: u representa o valor da unidade de área usado para expressar densidade (m²); n é o número de indivíduos amostrados e Σdi² soma do quadrado das distâncias entre as plantas amostradas.
Para a estimativa da abertura do dossel e do Índice de Área Foliar (IAF) foram obtidas fotografias hemisféricas nos meses de janeiro (seco) e junho (chuvoso). Ao longo dos transectos determinados inicialmente, estabeleceram-se cinco pontos equidistantes 20 m e georreferenciados, totalizando 10 pontos para cada ambiente.
Em cada um dos pontos dos transectos foi obtida uma fotografia hemisférica do dossel para as estações analisadas. As fotografias foram obtidas com câmera digital, acoplada a uma lente “olho de peixe”. A câmera foi posicionada sempre da mesma maneira, na altura de 1,30 m em relação ao solo e com a parte superior voltada para o norte. As fotografias foram tiradas ou no início da manhã (até as 10h) ou no final da tarde (a partir das 15h) para evitar exposição excessiva ao sol (GARCIA et al., 2007GARCIA, L. C. et al. Heterogeneidade do dossel e quantidade de luz no recrutamento do sub-bosque de uma mata ciliar no Alto São Francisco, Minas Gerais: análise através de fotos hemisféricas. Revista Brasileira de Biociências, Porto Alegre, v. 5, n. 2, p. 99-101, 2007.; NASCIMENTO; FAGG; FAGG, 2007NASCIMENTO, A. R. T.; FAGG, J. M. F.; FAGG, C. W. Canopy openness and lai estimates in two seasonally deciduous forests on limestone outcrops in Central Brazil using hemispherical photographs. Revista Árvore, Viçosa, MG, v. 31, n. 1, p. 167-176, 2007.). Em cada período avaliado (seco e chuvoso) foram obtidas 20 fotografas, sendo 10 na vegetação aberta e 10 na mata ripária, totalizando 40 fotografias. A análise das imagens foi realizada no software Gap Light Analyzer Version 2.0 (FRAZER; CANHAM; LERTZMAN, 1999FRAZER, G. W.; CANHAM, C. D.; LERTZMAN, K. P. Gap Light Analyzer (GLA), Version 2.0: imaging software to extract canopy structure and gap light transmission indices from true-colour fisheye photographs, users manual and program documentation. New York: Simon Fraser University, 1999.). A calibração e correção dos pontos de amostragem foram realizadas no Mangnetic Declination Calculator 1.0, disponível no site Natural Resources Canada(http://geomag.nrcan.gc.ca/calc/mdcal-eng.php).
Análise dos dados
Para verificar a frequência das classes diamétricas nos dois ambientes amostrados, foram eleborados histogramas utilizando a porcentagem de indivíduos em cada classe de DAP. As estimativas de abertura do dossel (%) e IAF foram comparadas entre os períodos seco e chuvoso, para cada ambiente e entre os ambientes, através do teste t. Aplicou-se o coeficiente de correlação de Pearson (r) entre as variáveis IAF x Abertura do dossel ajustado a um modelo de regressão linear (NASCIMENTO; FAGG; FAGG, 2007NASCIMENTO, A. R. T.; FAGG, J. M. F.; FAGG, C. W. Canopy openness and lai estimates in two seasonally deciduous forests on limestone outcrops in Central Brazil using hemispherical photographs. Revista Árvore, Viçosa, MG, v. 31, n. 1, p. 167-176, 2007.).

Frequência de observações do Diâmetro a Altura do Peito (DAP) das árvores na vegetação aberta (A) e na mata ripária (B) no Parque Nacional Serra de Itabaiana, Sergipe, Brasil.
FIGURE 2:
Observation frequencies of Chest High Diameter (DAP) of trees on the opened vegetation (A) and the riparian forest (B) in Serra de Itabaiana National Park, Sergipe state, Brazil.
Os dados foram previamente analisados quanto à normalidade através do teste Shapiro-Wilk (TORMAN; COSTER; RIBOLDI, 2012TORMAN, V. B. L.; COSTER, R.; RIBOLDI, J. Normalidade de variáveis: métodos de verificação e comparação de alguns testes não-paramétricos por simulação. Revista HCPA, Porto Alegre, v. 32, n. 2, p. 227-234, 2012.) e homocedasticidade pelo teste de Levene utilizando o pacote Lawstat do software R. As análises foram realizadas sob o nível de significância de 5%, com o auxílio dos softwares R versão 2.15 (R DEVELOPMENT CORE TEAM, 2006).
RESULTADOS
Nos transectos VA foram amostrados 79 indivíduos, sendo estimada densidade de 296 ind ha-1 e a distribuição de DAP variou de 1,6 a 12,1 cm (Figura 2a). Na MR foram amostrados 242 indivíduos, o que resultou na densidade total estimada de 498 ind ha-1. A distribuição do DAP na mata ripária seguiu o modelo de J-invertido, com maior número de indivíduos nas classes de menor diâmetro (Figura 2b). O DAP nesse ambiente variou de 2 a 51 cm.
Foram observados na VA valores de abertura do dossel de 67,39% em média no período seco, com desvio padrão (DP) ± 15,38, enquanto que no período chuvoso foi registrada abertura média de 52,24%, com DP ± 18,13. Não houve diferença entre os períodos seco e chuvoso (t = 2,0146; p > 0,05). O mesmo foi observado para o IAF cujas médias foram 0,36 (± 0,23) no período seco e 0,68 (± 0,45) no período chuvoso, não havendo diferença sazonal (t = -2,0052; p > 0,05). Na MR, o coeficiente de abertura do dossel variou em média 21,08% (± 2,36) na seca e 13,70% (± 2,24) no período chuvoso, havendo variação sazonal (t = 7, 1739; p < 0,05). Os valores de IAF também diferiram (t = -5.1609; p < 0,05) entre o período seco (1,63 ± 0,20) e chuvoso (2,13 ± 0,23).
A análise das fotografias hemisféricas permitiu observar diferenças significativas no dossel de ambos os ambientes amostrados no PNSI (Figura 3). Sendo que os valores de abertura do dossel na VA diferiram da MR, tanto no período seco (t = -9,4111; p < 0,0001) quanto no período chuvoso (t = -6.6693; p < 0,0001). No período seco, a abertura do dossel na vegetação aberta variou de 48,45% a 94,75% e na mata ripária variou de 17,06% a 23,94%. Enquanto que, no período chuvoso, a abertura do dossel na VA e na MR variou de 28,36% a 83,85% e de 9,33% a 16,83%, respectivamente (Figura 4a).
Os dois ambientes diferiram quanto ao IAF, tanto no período seco (t = 13,3854; p < 0,05) quanto no período chuvoso (t = 9,0899; p < 0,05; Figura 4b), sendo registrados maiores valores na MR, com média de 1,63 (± 0,20) no período seco e 2,13 (± 0,23) no período chuvoso. Adicionalmente, houve correlação negativa entre o IAF e a abertura do dossel, tanto na VA (r = -0,8455; p < 0,005; Figura 5a) como na MR (r = -0,9431; p < 0,05; Figura 5b).

Fotografias hemisféricas do dossel da vegetação aberta nos períodos seco (Abertura do dossel = 57,7%; A) echuvoso (Abertura dodossel= 51,1%; B); mata ripária nosperíodos seco (Abertura do dossel = 23,94% C) e chuvoso (Abertura do dossel = 9,33%; D), fotos obtidas em um único ponto em cada área, Parque Nacional Serra de Itabaiana, Sergipe, Brasil.
FIGURE 3:
Hemispherical picturesofthecanopyofopenedvegetationduringthedry season (Canopy openness = 57,5%, A) and wet season (Canopy openness, = 51,1%; B); riparian forest on dry season (Canopy openness = 23,94% C) and wet (Canopy openness = 9,33%; D), photos taken at a single point in each area, Serra de Itabaiana National Park.

(A) Variação da abertura do dossel na vegetação aberta (VA) e mata ripária (MR) nos períodos seco e chuvoso. (B) Variação do Índice de Área Foliar (IAF) na vegetação aberta (VA) e mata ripária (MR) nos períodos seco e chuvoso, amostrada no Parque Nacional Serra de Itabaiana, Sergipe, Brasil.
FIGURE 4:
(A) Canopy openness variation on opened vegetation (VA) and riparian forest (MR)on dry and wet seasons. (B) Leaf Area Index (IAF) on white-sand (AB) and riparian forest (MR) on dry and wet seasons, sampled in Serra de Itabaiana National Park, Sergipe state, Brazil.
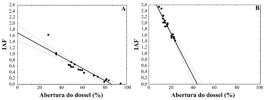
Relação entre Índice de Área Foliar (IAF) e abertura do dossel (%) na vegetação aberta (A) (r2 = 0,8603; r = -0,9275; p < 0,05) e na mata ripária (B) (r2 = 0,9281; r = -0,9634; p < 0,05), amostradas no Parque Nacional Serra de Itabaiana, Sergipe, Brasil.
FIGURE 5:
Relation between leaf Area Index (IAF) and canopy openness (%) on opened vegetation (A) (r² = 0,8603; r = 0,9275; p < 0,05) and on riparian forest (B) (r2 = 0,9281; r = -0,9634; p < 0,05), sampled in Serra de Itabaiana National Park, Sergipe state, Brazil.
DISCUSSÃO
Na vegetação aberta, a baixa densidade de árvores está relacionada às características fisionômicas dessa área, na qual as manchas irregulares de vegetação apresentam poucos indivíduos lenhosos e apresentam distribuição espacial agregada (DANTAS; RIBEIRO, 2010DANTAS, T. V. P. et al. Florística e estrutura da vegetação arbustivo-arbórea das Areias Brancas do Parque Nacional Serra de Itabaiana/Sergipe, Brasil. Revista Brasileira de Botânica, São Paulo, v. 33, p. 575-588, 2010.). Esse tipo de organização, formando associações de indivíduos, é um atributo típico desta comunidade, no qual as associações podem ser constituídas por árvores da mesma espécie, ou por diversas espécies compondo estruturas mais complexas (VILAR et al., 2000; VICENTE et al., 2005VICENTE, A. et al. Levantamento Botânico. In: CARVALHO, C. M.; VILAR, J. C. (Coord.). Parque Nacional Serra de Itabaiana: levantamento da Biota . São Cristóvão: UFS , 2005. p.15-37. (Biologia Geral e Experimental - UFS).; DANTAS et al., 2010). Já a pequena variabilidade nos valores de DAP registrada neste estudo, com concentração de árvores nas classes de menor tamanho e ausência de plantas de maior porte, mostrou que a vegetação aberta encontra-se com predomínio de indivíduos jovens. Dantas et al. (2010DANTAS, T. V. P. et al. Florística e estrutura da vegetação arbustivo-arbórea das Areias Brancas do Parque Nacional Serra de Itabaiana/Sergipe, Brasil. Revista Brasileira de Botânica, São Paulo, v. 33, p. 575-588, 2010.) comprovaram que existe variação na estrutura da vegetação nas areias brancas do PNSI ao analisarem duas áreas deste ambiente e encontrarem diferenças estruturais entre elas, sugerindo que tais distinções ocorrem em função dos estágios sucessionais da vegetação. Além disso, o local também apresenta histórico de degradação resultante de causas antrópicas, principalmente por desmatamentos e queimadas (VICENTE et al., 2005), o que pode justificar a presença das árvores de pequeno porte.
O padrão de distribuição dos indivíduos arbóreos nas matas abertas de areia branca do PNSI é do tipo agregado e com predomínio de indivíduos lenhosos jovens. Este resultado contribui para o entendimento das altas taxas de abertura do dossel encontradas nesta vegetação, com alguns pontos ultrapassando 80% de abertura. Isto é, a distribuição agregada das plantas lenhosas, resultando em lacunas de vegetação, pode condicionar a estrutura do dossel dessa floresta, com áreas mais abertas que outras. De acordo com Bianchini et al. (2001BIANCHINI, E.; PIMENTA, J. A.; SANTOS, F. A. M. Spatial and temporal variation in the canopy cover in a tropical semi-deciduous forest. Brazilian Archives Biology Technology, Curitiba, v. 44, n. 3, p. 269- 276, 2001.), a abertura do dossel pode ser influenciada por diversos aspectos de sua estrutura, como a composição de espécies, o arranjo espacial das plantas, a arquitetura das copas e a área foliar. Este último foi observado no presente estudo, em que o IAF foi altamente correlacionado com a abertura do dossel. Além disso, pesquisas demonstram também a existência de correlação positiva entre o IAF e o diâmetro a altura do peito das árvores (MARSHALL; WARING, 1986MARSHALL, J. D.; WARING, R. H. Comparison of methods of estimating Leaf Area Index in old growth Douglas-fir. Ecology , New York, v. 67, n. 4, p. 975-979, 1986.; CAMPBELL; NORMAN, 1989CAMPBELL, G. S.; NORMAN, J. M. The description and measurement of plant canopy structure. In: RUSSEL, G.; MARSHALL, B.; JARVIS, P. G. Plants canopies: their growth, form and function. Cambridge: Cambridge University Press, 1989. p. 1-19.; WINK et al., 2012WINK, C. et al. Parâmetros da copa e a sua relação com o diâmetro e altura das árvores de eucalipto em diferentes idades. Scientia Forestalis, Piracicaba, v. 40, n. 93, p. 57-67, 2012.), deixando claro que quanto menor o diâmetro das árvores, menor também é a área foliar e, portanto, maior será a abertura do dossel.
Os valores de IAF e de abertura do dossel na vegetação aberta foram semelhantes àqueles descritos para vegetação de Caatinga (NOGUEIRA JUNIOR, 2011NOGUEIRA JÚNIOR, F. C. Estrutura e composição de uma vegetação ripária, relações dendrocronológicas e climáticas na Serra dos Macacos em Tobias Barreto, Sergipe-Brasil. 2011. Dissertação (Mestrado) - Universidade Federal de Sergipe, São Cristóvão, 2011.; OLIVEIRA, 2012OLIVEIRA, D. G. Análise da vegetação em um fragmento de Caatinga no Município de Porto da Folha, Sergipe, Brasil. 2012. 145 f. Dissertação (Mestrado) - Universidade Federal de Sergipe, São Cristóvão, 2012.; PAGOTTO et al., 2015PAGOTTO, M. A. A Vegetação lenhosa da caatinga em assentamento do estado de Sergipe: aspectos fitossociológicos, anatômicos e dendrocronológicos. 2015. Tese (Doutorado) - Universidade Federal de Sergipe, São Cristóvão, 2015.) e outras florestas decíduas (MIRANDA et al., 1997MIRANDA, A. C. et al. Fluxes of carbon, water and energy over Brazilian cerrado: an analysis using eddy covariance and stable isotopes. Plant, Cell and Environment, Malden, v. 20, n. 3, p. 315-328, 1997.; NASCIMENTO; FAGG; FAGG, 2007NASCIMENTO, A. R. T.; FAGG, J. M. F.; FAGG, C. W. Canopy openness and lai estimates in two seasonally deciduous forests on limestone outcrops in Central Brazil using hemispherical photographs. Revista Árvore, Viçosa, MG, v. 31, n. 1, p. 167-176, 2007.), e inferiores àqueles encontrados para áreas semidecíduas (DE PAULA; LEMOS-FILHO, 2001DE PAULA, S. A.; LEMOS FILHO, J. P. Dinâmica do dossel em mata semidecídua no perímetro urbano de Belo Horizonte, MG. Revista Brasileira de Botânica , São Paulo, v. 24, n. 4, p. 545-5, 2001.; LEMOS-FILHO et al., 2010). Mas, de modo oposto às florestas decíduas, os parâmetros analisados não variaram entre os períodos de seca e chuva. Segundo Anderson (1981ANDERSON, A. B. White-Sand Vegetation of Brazilian Amazonia. Biotropica, Zurich, v. 13, n. 3, p. 199-210, 1981.), as plantas deste habitat podem apresentar adaptações que limitam a perda de água e que maximizam a absorção de nutrientes, incluindo a esclerofilia e a associações simbióticas com fungos micorrízicos. No entanto, não é possível afirmar se a ausência de resposta à sazonalidade aqui descrita corresponde de fato a alguma adaptação das plantas ao período seco, ou se a distribuição agregada das árvores, formando mosaicos de vegetação, contribuiu para este resultado. Em outras palavras, uma vez que as plantas lenhosas se encontram distribuídas de modo agregado, alguns pontos amostrados foram situados em áreas abertas, fora dessas agregações, resultando em baixas variações nos valores de IAF e de abertura do dossel entre as estações analisadas. Este fato deixa evidente a necessidade de mais estudos sobre o dossel das florestas de areias brancas strictu sensu.
Em relação à densidade de árvores na mata ripária, esta foi 40% maior que na vegetação aberta, fato este já esperado, dada as diferenças florísticas e estruturais entre os dois ambientes (DANTAS; RIBEIRO, 2010DANTAS, T. V. P. et al. Florística e estrutura da vegetação arbustivo-arbórea das Areias Brancas do Parque Nacional Serra de Itabaiana/Sergipe, Brasil. Revista Brasileira de Botânica, São Paulo, v. 33, p. 575-588, 2010.). Na mata ripária, há maior diversidade de espécies e a estratificação é mais pronunciada, apresentando sub-bosque mais estruturado (VICENTE et al., 2005VICENTE, A. et al. Levantamento Botânico. In: CARVALHO, C. M.; VILAR, J. C. (Coord.). Parque Nacional Serra de Itabaiana: levantamento da Biota . São Cristóvão: UFS , 2005. p.15-37. (Biologia Geral e Experimental - UFS).; DANTAS; RIBEIRO, 2010). A maioria dos indivíduos neste habitat está nas classes de menor DAP, deixando claro o padrão J-invertido dessa formação florestal e corrobora as descrições de Dantas et al. (2010DANTAS, T. V. P. et al. Florística e estrutura da vegetação arbustivo-arbórea das Areias Brancas do Parque Nacional Serra de Itabaiana/Sergipe, Brasil. Revista Brasileira de Botânica, São Paulo, v. 33, p. 575-588, 2010.) para a mesma área de estudo. Este padrão é comum em comunidades florestais que se autorregeneram, pois as árvores de menor dimensão representam a maioria da população, sugerindo que existe um balanço entre mortalidade e o recrutamento dos indivíduos (LOPES et al., 2002LOPES, W. P. et al. Estrutura fitossociológica de um trecho de vegetação arbórea no Parque Estadual do Rio Doce - Minas Gerais, Brasil. Acta Botanica Brasilica, Belo Horizonte, v. 16, n. 443-456, 2002.).
O IAF dessa vegetação foi maior quando comparado com a área aberta, em ambos os períodos estudados, tais valores são semelhantes àqueles encontrados para vegetações semidecíduas (DE PAULA; LEMOS-FILHO, 2001DE PAULA, S. A.; LEMOS FILHO, J. P. Dinâmica do dossel em mata semidecídua no perímetro urbano de Belo Horizonte, MG. Revista Brasileira de Botânica , São Paulo, v. 24, n. 4, p. 545-5, 2001.; LEMOS-FILHO et al., 2010) e corroboram a caracterização de Dantas e Ribeiro (2010DANTAS, T. V. P. et al. Florística e estrutura da vegetação arbustivo-arbórea das Areias Brancas do Parque Nacional Serra de Itabaiana/Sergipe, Brasil. Revista Brasileira de Botânica, São Paulo, v. 33, p. 575-588, 2010.), que classificaram essa vegetação como ecótono entre Floresta Ombrófila Densa e Floresta Estacional Semidecidual, cujas plantas perdem parte de suas folhas durante o período da seca.
Ainda que submetidas às mesmas condições macroclimáticas, na vegetação aberta e na mata ripária, as diferenças nos valores médios de abertura do dossel e IAF, indicam que há variação estrutural do dossel entre as duas fitofisionomias. Essa variação pode ser explicada em função da estrutura vegetal que, por sua vez, está atrelada às condições ambientais locais e aos fatores aleatórios, como limitação da dispersão, ocorrência de patógenos, disponibilidade de nutrientes no solo e o padrão de distribuição espacial das plantas (STROPP et al., 2011STROPP, J. et al. Tree communities of white-sand and terra-firme forests of the upper Rio Negro. Acta Amazonica, Manaus, v. 41, n. 4, p. 521-544, 2011.). Este último parece contribuir significativamente para as diferenças entre os dois habitat estudados. A distribuição agregada das árvores na vegetação aberta permite maior abertura do dossel e maior disponibilidade de luz no sub-bosque, diferentemente da mata ripária que por apresentar distribuição mais uniforme, proporciona um dossel contínuo.
CONCLUSÃO
A vegetação de areia branca no PNSI apresenta diferenças na abertura do dossel de acordo com a estrutura fitofisionômica. Áreas abertas, caracterizadas como vegetação de areias brancas strictu sensu apresentam baixo IAF e dossel mais aberto, em consequência, a distribuição agregada das árvores, que em sua maioria são de pequeno porte. Áreas de vegetação ripária, por sua vez, possuem maior densidade de plantas, com árvores de maior porte, formando dossel contínuo, cujos maiores valores de IAF contribuem para a menor abertura do dossel. As diferenças nos parâmetros analisados em função da sazonalidade, observada somente para a mata ripária, deixa clara a sensibilidade dessa vegetação diante das variações climáticas locais e expõe a necessidade de mais estudos sobre a abertura do dossel da vegetação das areias brancas strictu sensu, uma vez que essas apresentaram respostas distintas à sazonalidade típica deste ambiente. Portanto, as variações estruturais e temporais do dossel das florestas situadas em areias brancas quartzosas estão relacionadas à heterogeneidade e à complexidade estrutural do ambiente.
AGRADECIMENTOS
Agradecemos ao ICMBio/Parque Nacional Serra de Itabaiana pelo suporte e pela infraestrutura concedida durante a realização desta pesquisa
REFERÊNCIAS
- ALVARES, C. A. et al. Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift, Stuttgart, v. 22, n. 6, p. 711-728, 2013.
- ANDERSON, A. B. White-Sand Vegetation of Brazilian Amazonia. Biotropica, Zurich, v. 13, n. 3, p. 199-210, 1981.
- BEZERRA, M. F. A.; FARIAS, G. R.; CAVALCANTI, L. H. Mixobiota do Parque Nacional Serra de Itabaiana, SE, Brasil: Trichiales. Acta Botância Brasilica, Belo Horizonte, v. 24, n. 2, p. 510-517, 2010.
- BIANCHINI, E.; PIMENTA, J. A.; SANTOS, F. A. M. Spatial and temporal variation in the canopy cover in a tropical semi-deciduous forest. Brazilian Archives Biology Technology, Curitiba, v. 44, n. 3, p. 269- 276, 2001.
- BROWER, J. E.; ZAR, J. H.; VON ENDE, C. N. Field and laboratory methods for general ecology, 4th ed. Boston: WCB McGraw-Hill, 1998. 273 p.
- CAMPBELL, G. S.; NORMAN, J. M. The description and measurement of plant canopy structure. In: RUSSEL, G.; MARSHALL, B.; JARVIS, P. G. Plants canopies: their growth, form and function. Cambridge: Cambridge University Press, 1989. p. 1-19.
- CARVALHO, C. M.; VILAR, J. C. Parque Nacional Serra de Itabaiana: levantamento da Biota. São Cristóvão: UFS, 2005. 124 p. (Biologia Geral e Experimental - UFS).
- CATANA JUNIOR, J.; ANTHONY, J. The wandering quarter meted of estimating population density. Ecology, New York, v. 44, n. 2, p. 349-360, 1963.
- DANTAS, T. V. P.; RIBEIRO, A. S. Caracterização da vegetação do Parque Nacional Serra de Itabaiana, Sergipe Brasil. Biotemas, Florianópolis, v. 23. n. 4, p. 9-18, 2010.
- DANTAS, T. V. P. et al. Florística e estrutura da vegetação arbustivo-arbórea das Areias Brancas do Parque Nacional Serra de Itabaiana/Sergipe, Brasil. Revista Brasileira de Botânica, São Paulo, v. 33, p. 575-588, 2010.
- DE PAULA, S. A.; LEMOS FILHO, J. P. Dinâmica do dossel em mata semidecídua no perímetro urbano de Belo Horizonte, MG. Revista Brasileira de Botânica , São Paulo, v. 24, n. 4, p. 545-5, 2001.
- EMBRAPA. Levantamento exploratório: reconhecimento de solos do estado de Sergipe. Recife: EMBRAPA/SUDENE, 1975. 142 p. (Boletim técnico).
- FRAZER, G. W.; CANHAM, C. D.; LERTZMAN, K. P. Gap Light Analyzer (GLA), Version 2.0: imaging software to extract canopy structure and gap light transmission indices from true-colour fisheye photographs, users manual and program documentation. New York: Simon Fraser University, 1999.
- GARCIA, L. C. et al. Heterogeneidade do dossel e quantidade de luz no recrutamento do sub-bosque de uma mata ciliar no Alto São Francisco, Minas Gerais: análise através de fotos hemisféricas. Revista Brasileira de Biociências, Porto Alegre, v. 5, n. 2, p. 99-101, 2007.
- LEMOS FILHO, J. P. et al. Spatial and temporal variability of canopy cover and understory light in a Cerrado of Southern Brazil. Brazilian Journal Biology, São Carlos, v. 70, n. 1, p. 19-24, 2010.
- LOPES, W. P. et al. Estrutura fitossociológica de um trecho de vegetação arbórea no Parque Estadual do Rio Doce - Minas Gerais, Brasil. Acta Botanica Brasilica, Belo Horizonte, v. 16, n. 443-456, 2002.
- MARSHALL, J. D.; WARING, R. H. Comparison of methods of estimating Leaf Area Index in old growth Douglas-fir. Ecology , New York, v. 67, n. 4, p. 975-979, 1986.
- MENDONÇA, B. A. F. et al. Solos e Geoambientes do Parque Nacional do Viruá e entorno, Roraima: visão integrada da paisagem e serviço ambiental. Ciência Florestal, Santa Maria, v. 23, p. 429-444, 2013.
- MENDONÇA, B. A. F. et al. Use of geophysical methods for the study of sandy soils under Campinarana at the National Park of Viruá, Roraima state, Brazilian Amazonia. Journal of Soils and Sediments, Landsberg, v. 14, p. 1-13, 2014.
- MIRANDA, A. C. et al. Fluxes of carbon, water and energy over Brazilian cerrado: an analysis using eddy covariance and stable isotopes. Plant, Cell and Environment, Malden, v. 20, n. 3, p. 315-328, 1997.
- NASCIMENTO, A. R. T.; FAGG, J. M. F.; FAGG, C. W. Canopy openness and lai estimates in two seasonally deciduous forests on limestone outcrops in Central Brazil using hemispherical photographs. Revista Árvore, Viçosa, MG, v. 31, n. 1, p. 167-176, 2007.
- NICOTRA, A. B.; CHAZDON, R. L.; IRIARTE, S. V. B. Spatial heterogeneity of light and woody seedling regeneration in tropical wet forests. Ecology , New Iork, v. 80, n. 6, p. 1908-1926, 1999.
- NOGUEIRA JÚNIOR, F. C. Estrutura e composição de uma vegetação ripária, relações dendrocronológicas e climáticas na Serra dos Macacos em Tobias Barreto, Sergipe-Brasil. 2011. Dissertação (Mestrado) - Universidade Federal de Sergipe, São Cristóvão, 2011.
- OLIVEIRA, A. A. et al. Florestas sobre areia: Campinaranas e Igapós. In: OLIVEIRA, A. A.; DALY, D. C. (Eds.). Florestas do rio Negro. São Paulo: Schwarcz, 2001. p. 179-220.
- OLIVEIRA, D. G. Análise da vegetação em um fragmento de Caatinga no Município de Porto da Folha, Sergipe, Brasil. 2012. 145 f. Dissertação (Mestrado) - Universidade Federal de Sergipe, São Cristóvão, 2012.
- PAGOTTO, M. A. A Vegetação lenhosa da caatinga em assentamento do estado de Sergipe: aspectos fitossociológicos, anatômicos e dendrocronológicos. 2015. Tese (Doutorado) - Universidade Federal de Sergipe, São Cristóvão, 2015.
- POLLARD, J. H. On distance estimators of density in randomly distributed forests. Biometrics, Malden, v. 27, n. 4, p. 991-1002, 1971.
- R DEVELOPMENT CORE TEAM. A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing, 2006.
- DISTRITO FEDERAL (Brasil). Secretaria de Meio Ambiente e Recursos Hídricos. Base de dados do Centro de Meteorologia. Disponível em planilha eletrônica. 2012.
- STROPP, J. et al. Tree communities of white-sand and terra-firme forests of the upper Rio Negro. Acta Amazonica, Manaus, v. 41, n. 4, p. 521-544, 2011.
- TORMAN, V. B. L.; COSTER, R.; RIBOLDI, J. Normalidade de variáveis: métodos de verificação e comparação de alguns testes não-paramétricos por simulação. Revista HCPA, Porto Alegre, v. 32, n. 2, p. 227-234, 2012.
- VAN DEN BERG, E.; SANTOS, F. A. M. Aspectos da variação ambiental em uma floresta de galeria em Itutinga, MG, Brasil. Ciência Florestal , Santa Maria, v. 13, n. 2, p. 83-98, 2003.
- VICENTE, A. et al. Levantamento Botânico. In: CARVALHO, C. M.; VILAR, J. C. (Coord.). Parque Nacional Serra de Itabaiana: levantamento da Biota . São Cristóvão: UFS , 2005. p.15-37. (Biologia Geral e Experimental - UFS).
- WINK, C. et al. Parâmetros da copa e a sua relação com o diâmetro e altura das árvores de eucalipto em diferentes idades. Scientia Forestalis, Piracicaba, v. 40, n. 93, p. 57-67, 2012.
- ZUQUIM, G.; COSTA, F. R. C.; PRADO, J. Fatores que determinam a distribuição de espécies de pteridófitas da Amazônia Central. Revista Brasileira de Biociências , Porto Alegre, v. 5, n. 2, p. 360-362, 2007.
Datas de Publicação
-
Publicação nesta coleção
Apr-Jun 2017
Histórico
-
Recebido
13 Mar 2015 -
Aceito
09 Set 2015





