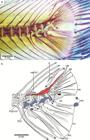
Fig. 1
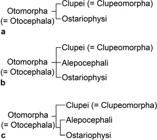
Clupeocephalan relationships proposed from 1973 to 1994. a. after Rosen 1973Rosen DE. Interrelationships of higher euteleostean fishes. In: Interrelationships of Fishes . Greenwood PH, Miles RS, Patterson C, editors. London: Academic Press; 1973. p.397-513., 1974Rosen DE. Phylogeny and zoogeography of salmoniform fishes and relationships of Lepidogalaxias salamandroides. Bull Amer Mus Natur Hist . 1974; 153:265-326.; b. after Rosen, 1982Rosen DE. Teleostean interrelationships, morphological function and evolutionary inference. Amer Zool. 1982; 22:261-73.; c. after Fink, Weitzman, 1982Fink WL, Weitzman SH. Relationships of the stomiiform fishes (Teleostei), with a redescription of Diplophos. Bull Mus Comp Zool. 1982; 150:31-93.; d. after Lauder, Liem, 1983Lauder GV, Liem KF. The evolution and interrelationships of the actinopterygian fishes. Bull Mus Comp Zool . 1983; 150:95-197.; e. after Fink, 1984Fink WL. Basal euteleosts: Relationships. In: Ontogeny and Systematic of Fishes. Moser GH, Richards WJ, Cohen DM, Fahay MP, Kendall AW Jr, Richarson SL, editors. Lawrence, KS: Amer. Soc. Ichthyol. Herpetol. 1984; p.2002-6.; f. after Rosen, 1985Rosen DE. An essay on euteleostean classification. Amer Mus Novitates . 1985; 2827:1-57.; g. after Begle, 1991Begle DP. Relationships of the osmeroid fishes and the use of reductive characters in phylogenetic analysis. Syst Zool. 1991; 40(1):33-53., 1992Begle DP. Monophyly and relationships of the argentinoid fishes. Copeia . 1992;1992(2):350-66.; h. after Lecointre, Nelson, 1996Lecointre G, Nelson GJ. 1996. Clupeomorpha, sister-group of Ostariophysi. In: Interrelationships of Fishes. Stiassny MLJ, Parenti LR, Johnson GD, editors. San Diego: Academic Press ; 1996. p.193-207.. E = Euteleostei. Characters 1 = nuptial tubercles; 2 = stegural; 3 = adipose fin; 4 = absence of tooth plate over basibranchial 4; 5 = pleurostyle; 6 = fusion of hypural 2 with first ural centrum; 7 = fusion of extrascapulars and parietal bones; 8 = fusion of hemal arches and centra anterior to preural centrum 2. (Modified from Lecointre, Nelson, 1996Lecointre G, Nelson GJ. 1996. Clupeomorpha, sister-group of Ostariophysi. In: Interrelationships of Fishes. Stiassny MLJ, Parenti LR, Johnson GD, editors. San Diego: Academic Press ; 1996. p.193-207.: fig. 1).
Fig. 2
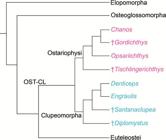
Hypotheses of Otomorpha relationships according to molecular and morphological evidence (a) and only molecular evidence (b and c).
a. after
Lé et al., 1993Lé HLV, Lecointre G, Perasso R. A 28S rRNA-based phylogeny of the gnathostomes: first steps in the analysis of conflict and congruence with morphologically based cladograms. Mol Phyl Evol . 1993; 2(1):31-51.;
Lecointre, 1993Lecointre G. Etude d l’impact de l’echantillonnage des espèces et de la longueur des séquences sur la robustesse des phylogénies moléculaires; implications sur la phylogénie des téléostéens. [PhD Thesis]. Paris: Université Paris VII, 1993.;
Lecointre, Nelson, 1996Lecointre G, Nelson GJ. 1996. Clupeomorpha, sister-group of Ostariophysi. In: Interrelationships of Fishes. Stiassny MLJ, Parenti LR, Johnson GD, editors. San Diego: Academic Press ; 1996. p.193-207.;
Arratia 1996Arratia G, Cione LA. The fossil record of fossil fishes of Southern South America. In: Contributions of Southern South America to Vertebrate Paleontology. Arratia G, editor. Muenchner Geowiss. Abh.. München: Verlag Dr. F. Pfeil ; 1996. p.9-72. (special volume),
1997Arratia G. Basal teleosts and teleostean phylogeny. Palaeo Ichthyologica. 1997; 7:1-168.,
1999Arratia G. The monophyly of Teleostei and stem group teleosts. In: Mesozoic Fishes - Systematics and Fossil Record. Arratia G, Schultze H-P, editors. München: Verlag Dr. F. Pfeil ; 1999. p.265-334.,
2010Arratia G. Clupeocephala re-visited: Analysis of characters and homologies. Revista Biol Mar & Oceanogr. 2010; 45(Suppl.1):635-57.;
Wiley, Johnson, 2010Wiley EO, Johnson GD. A teleost classification based on a monophyletic group. In: Origin and Phylogenetic Relationships on Teleosts. Nelson JS, Schultze H-P, Wilson MVH, editors. München: Verlag. Dr. F. Pfeil; 2010. p.123-182.;
b. after
Lavoué et al., 2005Lavoué S, Miya M, Inoue JG, Saitoh K, Ishiguro NB et al. Molecular systematics of the gonorynchiform fishes (Teleostei) based on whole mitogenome sequences: implications for higher-level relationships within the Otocephala. Mol Phyl Evol. 2005; 37(1):165-77.,
Lavoué et al., 2008Lavoué S, Miya M, Poulsen JY, Møller PR, Nishida M. Monophyly, phylogenetic position and interfamilial relationships of the Alepocephaliformes (Teleostei) based on whole mitogenome sequences. Mol Phyl Evol . 2008; 47(3):1111-21.;
Poulsen et al., 2009Poulsen JY, Møller PR, Lavoué S, Knudsen SW, Nishida M, Miya M. Higher and lower-level relationships of the deep-sea fish order Alepocephaliformes (Teleostei: Otocephala) inferred from whole mitogenome sequences. Biol J Linn Soc. 2009; 98(4):923-36.;
c. after
Near et al., 2012Near TJ, Eytan RI, Dornbur A, Kuhn KL, Moore JA, Davis PM et al. Resolution of ray-finned fish phylogeny and timing of diversification. Proc Nat Acad Sciences USA. 2012; 109(34):13698-13703.;
Betancur-R. et al., 2013Betancur-R R, Broughton RE, Wiley EO, Carpenter K, López JA, Li C et al. The tree of life and a new classification of bony fishes. PLoS Curr. 2013; Available from: doi:10.1371/currents.tol.53ba26640df0ccaee75bb165c8c26288.
https://doi.org/10.1371/currents.tol.53b...
;
Betancur-R. et al., 2017Betancur-R R, Wiley EO, Arratia G, Acero A, Bailly N, Miya M et al. Phylogenetic classification of bony fishes. BMC Evolutionary Biology. 2017; 17(162). Available from: https://doi.org/10.1186/s12862-017-0958-3
https://doi.org/10.1186/s12862-017-0958-...
.
Fig. 3
Hypothesis of phylogenetic relationships of the Ostarioclupeomorpha (= OST-CL; Otocephala) among the crown Teleostei, based on fossil (†) and recent teleosts (abbreviated from Arratia, 1999Arratia G. The monophyly of Teleostei and stem group teleosts. In: Mesozoic Fishes - Systematics and Fossil Record. Arratia G, Schultze H-P, editors. München: Verlag Dr. F. Pfeil ; 1999. p.265-334.:fig. 19).
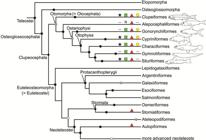
Fig. 4
Diagrammatic representation of different otomorph groups such as: a. clupeiformes; b. alepocephaliforms; c. gonorynchiforms; d. cypriniforms; e. characiforms; f. siluriforms; g. gymnotiforms.
Fig. 5
Fossil otomorphs in lateral view. a. ostariophysan incertae sedis †Tischlingerichthys viohliArratia, 1997Arratia G. Basal teleosts and teleostean phylogeny. Palaeo Ichthyologica. 1997; 7:1-168.; Upper Jurassic, Germany; holotype JM-E Moe 8; photograph courtesy of H. Tischlinger; b. clupeomorph †Santanaclupea silvasantosiMaisey, 1993Maisey J. A new clupeomorph fish from the Santana Formation (Albian) of NE Brazil. Amer Mus Novitates . 1993; 3076:1-15.; Lower Cretaceous, Brazil; paratype DNPM DGM1338-P; photograph courtesy of D. Mayrinck; c. clupeomorph †Ranulfoichthys dorsonudumAlvarado-Ortega, 2014Alvarado-Ortega J. Ancient herring from the Tlayúa Quarry (Cretaceous, Albian) near Tepexi de Rodríguez, Puebla State, central Mexico, closing the gap in the early diversification of Clupeomorpha. Cretaceous Res. 2014; 50:171-80.; Lower Cretaceous, Mexico; paratype IGM 9034; photograph courtesy of J. Alvarado-Ortega; d. gonorynchiform †Gordichthys conquensis Poyato-Ariza, 1994; holotype MCCM-LH-1228; Lower Cretaceous, Spain; photograph by J.A. Gracia; courtesy by F.J. Poyato-Ariza. Scale bars = 1 cm.
Fig. 6
Fossil otomorph in lateral view: †Paleodenticeps sp.; Eocene, Mahenge, Tanzania (MB uncatalogued); photograph courtesy of C. Quezada-Romegialli. Scale bar equals 5 mm.
Fig. 7
Caudal skeleton of Argentina sialis (SIO 66-4) in lateral view. a. photograph of specimen of 144.2 mm standard length; photograph courtesy of C. Quezada-Romegialli; b. interpretative drawing. Note that the posterior hemi-lepidotrichia of principal ray 10 is displaced. The color orange is used to denote a modified uroneural named pseudopleurostyle herein. CC = compound vertebral centrum; E = epural; H1,3,6 = hypurals 1,3,6; hsPU2 = haemal spine of preural centrum 2; naPU1 = neural arch of preural centrum 1; PR1 = 10,11,19 = principal caudal ray 1,10,11,19; PU3 = preural centrum 3; ‘UD’ = tendon-bone urodermal; UN = uroneural.
Fig. 8
Fossil otomorph in lateral view: a.
Carpathichthys polonicusJerzmanska, 1979Jerzmaska A. Oligocene alepocephaloid fishes from the Polish Carpathians. Acta Paleontol Pol. 1979; 24(1):65-76.; Miocene-Oligocene, Polish Carpathians, Europe (ZPALWr a/2004, holotype). b.
Carpathichthys sp. from the same locality and age (ZPALWr N/6116). Photographs courtesy of Małgorzata Bieńkowska-Wasiluk.
Fig. 9
Diagrammatic caudal skeletons in lateral view illustrating the pleurostyle, its relationships (a-c) and early development (d, e). a. clupeoid; b. gonorynchiform; c. cypriniform; d. part of caudal skeleton in lateral view of the cypriniform Catostomus commersoni, 13.4 mm SL (KU:KUI 38655); e. part of caudal skeleton in lateral view of the cypriniform Luxilus zonatus, 8.7 mm SL (KUI uncat.). CC = terminal caudal compound centrum; H3 = hypural 3; naPU1 = neural arch preural centrum 1; naU1, naU2 = neural arch of ural centrum 1, of ural centrum 2 [of the polyural terminology; ural centrum 1 of the diural terminology]; PH = parhypural; Pl = plurostyle; PU1 = PU3 = preural centrum 1, 3; U2+H2 = ural centrum 2 + hypural 2 [= ural centrum 2 of polyural terminology; ural centrum 1 of diural terminology]; U1-3 = ural chordacentrum 1, 2, 3 [of the polyural terminology, each associated with its correspondent hypural].
Fig. 10
Ontogenetic development of the caudal skeleton in alepocephaliforms. a. larva of Talismania bifurcata of 28.18 mm standard length (KU:KUI 41395) illustrating a functional notochord and formation of neural and haemal arches and spines developing from caudad to rostrad direction; b. detail of the caudal skeleton of specimen illustrated in a; note the unusual extension of the haemal cartilage; c. larvae of 68 mm standard length; note the marked differences in the development of chordacentra; small arrows point to the haemal cartilage d.
Searsia koefoedi; specimen of 126 mm standard length; note the development of the first uroneural or stegural-like element and the lack of chordacentra in front of hypurals 1 and 2. Photograph courtesy of C. Quezada-Romegialli. cPU1 = preural chordacentrum 1; cU1, 4 = ural chordacentrum 1, 4 [polyural terminology; ural centrum 1, 2 of diural terminology]; E = epural; h1-6 = hypural 1-6; hd = hypural diastema; hsPU2, hsPU3, hsPU4 = haemal spine of preural centrum 2, 3, 4; mc = median caudal cartilages; naPU1 = neural arch of preural centrum 1; ns PU2, nsPU3 = neural spine of preural centrum 2, 3; no = notochord; op.c = ophistural cartilage; PH = parhypural; UN = uroneurals.
Fig. 11
Caudal skeleton of Alepocephalus tenebrosus (UW 042217 #4). a. photograph of specimen of 363 mm standard length; photograph courtesy of C. Quezada-Romegialli; b. interpretative drawing. Note the articulation between the proximal region of hypural 3 and the haemal cartilage (arrow) uniting hypurals 1 and 2. E = epural; H1, 3, 6 = hypural 1, 3, 6; hd = hypural diastema; hsPU2 = haemal spine of preural centrum 2; mc = median caudal cartilages; naPU1 = neural arch of preural centrum 1; no = notochord; PH = parhypural; UN = uroneurals; the small third one is not shown.
Fig. 12
Abbreviated phylogenetic hypothesis of teleosts (modified from
Betancur-R. et al., 2017Betancur-R R, Wiley EO, Arratia G, Acero A, Bailly N, Miya M et al. Phylogenetic classification of bony fishes. BMC Evolutionary Biology. 2017; 17(162). Available from: https://doi.org/10.1186/s12862-017-0958-3
https://doi.org/10.1186/s12862-017-0958-...
) with distribution of certain character states discussed in the text. Yellow circle = fusion or ankylosis between medial extrascapular and parietal bones; red triangle = vertebrae anterior to preural centrum 2 with haemal arches and centra fused in juvenile-adult individuals; green square = silvery areas associated with gas bladder; black star = hypural 2 fused with ural centrum 2 [of the polyural terminology; ural centrum 1 of the diural terminology]. N = non applicable.
Fig. 13
Posterior cranial region of otomorphs illustrating the relationships of the extrascapular and parietal bones and sensory canals. a. gonorynchiform Chanos chanos of 45 mm standard length; KUI 38796; b.
Chanos chanos of 370 mm standard length; KUI 40935; c. diagrammatic drawing of Chanos chanos; d. diagrammatic drawing of the cypriniform Opsariichthys bidens (CAS 15639); e. diagrammatic drawing of the characiform Xenocharax spilurus (CAS 32512); f. diagrammatic drawing of the alepocephaliforms Leptoderma sp.; g. diagrammatic drawing of the alepocephaliform Searsia koefoedi. c-e, modified from Arratia, Huaquín, 1995Arratia G, Huaquín L. Morphology of the lateral line system and of the skin of diplomystid and certain primitive loricarioid catfishes and systematic and ecological considerations. Bonner zool Monogr . 1995; 36:1-110. and f, g, modified from Greenwood, Rosen, 1971Greenwood PH, Rosen DE. Notes on the structure and relationships of the alepocephaloid fishes. American Mus Novitates. 1971; 2473:1-48.. Exc = ossicle-like extrascapular bones; l. Exc = lateral extrascapula; m.Exc = medial extrascapula; Pa = parietal bone [of traditional terminology]; Ptt = posttemporal; Soc = supraoccipital.
Fig. 14
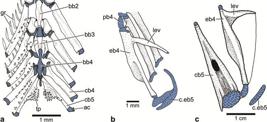
Branchial arches and cartilages. a.
Denticeps clupeoides illustrating a vestigial cartilage associated with ceratogranchial 5 (modified from de Pinna, Di Dario, 2010de Pinna MCC, di Dario F. The branchial arches of the primitive clupeomorpzh fish, Denticeps clupeoides, and their phylogenetic implication (Clupeiformes, Denticipitidae). In: Origin and Phylogenetic Interrelationships of Teleosts . Nelson JS, Schultze H-P, Wilson MVH, editors. München: Verlag Dr. F. Pfeil ; 2010. p.251-268.); b. epibranchials 4 and 5 in left lateral view illustrating the cartilaginous epibranchial 5 in a young specimen of Chanos chanos (modified from Johnson, Patterson, 1997Johnson GD, Patterson C. The gill arches of gonorynchiform fishes. South Afr J Sci. 1997; 93:594-600.); c. ceratobranchial 5 and cartilaginous epibranchial 5 in dorsal view in a large specimen of Chanos chanos of 490 mm SL (KU:KUI 40365). Arrow point to an elastic fiber membrane supporting branchial lamellae. ac = additional cartilage; bb2-4 = basibranchials 2-4; cb4,5 = ceratobranchials 4,5; c.eb5, cartilaginous epibranchial 5eb4 = epibranchial 4gr = gill rakers; lev = levator process; pb4 = pharyngobranchial 4; tp = tooth plate. Note the reduction in size of the cartilaginous epibranchial 5 in Chanos chanos.

 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail
 Thumbnail
Thumbnail













