Abstracts
Pore-linked filaments were visualized in spreads of anuran spermatocyte nuclei using transmission electron microscope. We used Odontophrynus diplo and tetraploid species having the tetraploid frogs reduced metabolic activities. The filaments with 20-40 nm width are connected to a ring component of the nuclear pore complex with 90-120 nm and extend up to 1µm (or more) into the nucleus. The filaments are curved and connect single or neighboring pores. The intranuclear filaments are associated with chromatin fibers and related to RNP particles of 20-25 nm and spheroidal structures of 0.5µm, with variations. The aggregates of several neighboring pores with the filaments are more commonly observed in 4n nuclei. We concluded that the intranuclear filaments may correspond to the fibrillar network described in Xenopus oocyte nucleus being probably related to RNA transport. The molecular basis of this RNA remains elusive. Nevertheless, the morphological aspects of the spheroidal structures indicate they could correspond to nucleolar chromatin or to nucleolus-derived structures. We also speculate whether the complex aggregates of neighboring pores with intranuclear filaments may correspond to pore clustering previously described in these tetraploid animals using freeze-etching experiments.
pore-linked filaments; intranuclear filaments; clustered-like NPCs in polyploid anurans; RNA transport
Filamentos ligados a poros foram visualizados em núcleos de espermatócitos de anuros através da técnica de espalhamento para microscopia eletrônica de transmissão. Os animais usados pertencem ao gênero Odontophrynus com espécies cripticas diplo e tetraplóides naturais, tendo os tetraplóides atividade metabólica reduzida. Os filamentos com 20-40 nm de largura são ligados a um anel componente do complexo poro nuclear de 90-120 nm e estendem-se até 1 µm (ou mais) para dentro do núcleo. Os filamentos são curvos e ligam poros simples ou poros vizinhos. Os filamentos intranucleares são associados a fibras de cromatina e relacionados a partículas de RNP de 20-25 nm e a estruturas esféricas de 0.5µm, com variações. Os agregados de poros vizinhos com os filamentos longos são mais freqüentemente observados em núcleos 4n. Concluímos que os filamentos intranucleares podem corresponder aos emaranhados fibrilares descritos em núcleos de oócitos de Xenopus e possivelmente relacion ados ao transporte de RNA. A base molecular desse RNA não é conhecida. Contudo, os aspectos morfológicos das estruturas esféricas parecem indicar que elas podem corresponder à cromatina nucleolar ou a estruturas derivadas do nucléolo. Também, especulamos se os agregados complexos de poros vizinhos com os filamentos intranucleares podem corresponder aos aglomerados de poros previamente descritos nesses animais tetraplóides através da técnica''freeze-etching''.
filamentos ligados ao poro; filamentos intranucleares; aglomerados de NPCs em anuros poliplóides; transporte de RNA
BIOMEDICAL AND MEDICAL SCIENCES
Pore-linked filaments in anura spermatocyte nuclei
Maria Luiza Beçak; Kazumi Fukuda-Pizzocaro
Laboratório de Genética, Instituto Butantan, Av. Vital Brasil, 1500, 05503-900 São Paulo, SP, Brasil
ABSTRACT
Pore-linked filaments were visualized in spreads of anuran spermatocyte nuclei using transmission electron microscope. We used Odontophrynus diplo and tetraploid species having the tetraploid frogs reduced metabolic activities. The filaments with 20-40 nm width are connected to a ring component of the nuclear pore complex with 90-120 nm and extend up to 1µm (or more) into the nucleus. The filaments are curved and connect single or neighboring pores. The intranuclear filaments are associated with chromatin fibers and related to RNP particles of 20-25 nm and spheroidal structures of 0.5µm, with variations. The aggregates of several neighboring pores with the filaments are more commonly observed in 4n nuclei. We concluded that the intranuclear filaments may correspond to the fibrillar network described in Xenopus oocyte nucleus being probably related to RNA transport. The molecular basis of this RNA remains elusive. Nevertheless, the morphological aspects of the spheroidal structures indicate they could correspond to nucleolar chromatin or to nucleolus-derived structures. We also speculate whether the complex aggregates of neighboring pores with intranuclear filaments may correspond to pore clustering previously described in these tetraploid animals using freeze-etching experiments.
Key words: pore-linked filaments, intranuclear filaments, clustered-like NPCs in polyploid anurans, RNA transport.
RESUMO
Filamentos ligados a poros foram visualizados em núcleos de espermatócitos de anuros através da técnica de espalhamento para microscopia eletrônica de transmissão. Os animais usados pertencem ao gênero Odontophrynus com espécies cripticas diplo e tetraplóides naturais, tendo os tetraplóides atividade metabólica reduzida. Os filamentos com 20-40 nm de largura são ligados a um anel componente do complexo poro nuclear de 90-120 nm e estendem-se até 1 µm (ou mais) para dentro do núcleo. Os filamentos são curvos e ligam poros simples ou poros vizinhos. Os filamentos intranucleares são associados a fibras de cromatina e relacionados a partículas de RNP de 20-25 nm e a estruturas esféricas de 0.5µm, com variações. Os agregados de poros vizinhos com os filamentos longos são mais freqüentemente observados em núcleos 4n. Concluímos que os filamentos intranucleares podem corresponder aos emaranhados fibrilares descritos em núcleos de oócitos de Xenopus e possivelmente relacion ados ao transporte de RNA. A base molecular desse RNA não é conhecida. Contudo, os aspectos morfológicos das estruturas esféricas parecem indicar que elas podem corresponder à cromatina nucleolar ou a estruturas derivadas do nucléolo. Também, especulamos se os agregados complexos de poros vizinhos com os filamentos intranucleares podem corresponder aos aglomerados de poros previamente descritos nesses animais tetraplóides através da técnica''freeze-etching''.
Palavras-chave: filamentos ligados ao poro, filamentos intranucleares, aglomerados de NPCs em anuros poliplóides, transporte de RNA.
INTRODUCTION
The importance of the study of nuclear pore complexes (NPCs) and of intranuclear filaments in nucleocytoplasm transport has been predicted to have many potential applications for gene expression control and antiviral therapies. Also, evidences were presented postulating that the fusion of certain nucleoporins to certain proteins may contribute to tumorigenesis (Nigg 1997). NPCs are structures embedded in the nuclear envelope (NE) that mediate the transport of macromolecules and particles between nucleus and cytoplasm during interphase. The architecture of the NPC was extensively studied in amphibian oocyte nuclei through different electron microscopy methodologies (Franke and Scheer 1970, Unwin and Milligan 1982, Scheer et al. 1988, Akey 1989, Ris 1989, 1991, Jarnik and Aebi 1991, Goldberg and Allen 1992, Panté et al. 1994, Panté and Aebi 1996, Allen et al. 2000). Vertebrate NPCs have an outer diameter of about 125 nm. Basically it has a cylindrical shape located between inner and outer membranes of the NE. The framework of the NPC contains the spoke ring complex which displays a channel called plug or transporter. The spoke ring complex is flanked by cytoplasmic and nuclear rings. The cytoplasmic ring with 125 nm in diameter is decorated with eight 30 to 50 nm long kinked filaments. The nuclear ring with 90 nm is capped with a basket-like assembly built from eight thin filaments with 8-10 nm diameter and 50 to 100 nm long. These filaments join at their distal ends in a smaller terminal ring of 30 to 50 nm diameter. First transmission electron microscope (TEM) studies in amphibian oocyte nuclei reported filaments attached to the nucleoplasmic face of the NPC (Afzelius 1955) that may be baskets, as commented (Goldberg et al. 1997). TEM experiments for NE spreads of Xenopus laevis oocytes showed that the baskets are insensitive to RNase treatment, its structural integrity depending on the presence of divalent cations as well as on the use of 1% tannic acid (Jarnik and Aebi 1991).
Long filaments have been described at the nucleoplasmic face of the NPC. These filaments with 1mm long and about 3-6 nm thick are arranged in a cylinder and seem to anastomose and to branch into subfibrils extending toward different NPCs (Franke and Scheer 1970). Further, fibers attached to the filaments of the baskets and forming a regular network called nuclear envelope lattice (NEL) were observed in Triturus and Xenopus oocyte nuclei using high-resolution electron microscopy (HREM) (Goldberg and Allen 1992). TEM studies in Xenopus oocyte nuclei using spreads and conventional sections also indicated that bundles of these filaments extend inward the nucleus to amplified nucleoli. This finding led to the assumption that the basket would be part of a higher-order structure associated with NPCs and chromatin fibers (Arlucea et al. 1998). According to this idea groups of eight pore-linked filaments form cylinders for diffusion of cargos leading to each NPC. The function of these long intranuclear filaments is still a matter of speculation, although evidences have been reported favoring its role for nucleocytoplasmic transport. As known some models for nucleocytoplasmic transport were previously proposed. Based in data using in situ hybridization optimized for fluorescent detection of nuclear RNA it was suggested that transcripts are transported vectorially from genes to NPCs. This traffic would be visualized through ''tracks'' in the nuclear architecture (Lawrence et al. 1989). An alternative model based in conjugated experiments using in situ hybridization, immunolocalization and confocal microscopy suggested that gene products move from genes to nuclear surface by diffusion through an extrachromosomal channel network (ECN) (Zackar et al. 1993). Recently, field emission electron microscopy (feSEM) studies in Xenopus oocyte nucleus showed a network of pore-linked filaments extending inward the nucleus. These filaments with 40 nm diameter (ranging from 12-120 nm) appear embedded in spherical nuclear bodies demonstrated to be nucleoli. The finding that the filaments contain actin and protein 4.1 led to the suggestion that the network could provide ''diffusion channels'' through chromatin (Kiseleva et al. 2004).
The composition of the NPCs and of the intranuclear filaments are being studied by several authors. In vertebrate nuclei the NPCs contain several nucleoporins some related to the transport. Nup 153 was localized to the nuclear side of the NPC (Sukegawa and Blobel 1993) and to the distal ring of the basket (Panté et al. 1994). This nucleoporin also occurs in the long intranuclear filaments and can be observed connecting NPCs to the nucleolus (Cordes et al. 1993). The long filaments also contain p270/Tpr/Bx34 proteins (Cordes et al. 1997, Zimowska et al. 1997). The function of Nup 153 was investigated in experiments using removal of specific nucleoporins in extracts derived from Xenopus eggs. The results of these genetic assays demonstrated that the removal of Nup 153 leads to structural disturbs of NPC - linked filaments and alters the interaction of the NPC with the nuclear lamina. These alterations may cause mobilization of NPCs and probably NPCs clustering affecting the transport (Walther et al. 2001). Moreover the localization of actin and 4.1 protein in the pore-linked filaments indicated its relation to the extrachromosomal diffusion channels (Kiseleva et al. 2004).
First whole mount experiments followed by critical point drying showed that chromatin fibers are associated with the annuli of the NE (DuPraw 1965). The same procedure allowed the visualization of fibers attached to the nucleoplasmic side of the NE (Aréchaga and Bahr 1985). Though well-preserved baskets were observed in NE spreads with the use of tannic acid fixative and uranyl acetate staining long filaments were not evidenced. This is probably due to their extraction during nucleus removal (Arlucea et al. 1998).
This paper contains the description of pore-linked filaments found in spermatocyte nuclei of the diplo and tetraploid Odontophrynus americanus species using unfixed spread assays for TEM. The intranuclear filaments connect single or neighboring pores and are associated with RNP particles and spheroidal structures of 0.5µm. These round elements may correspond to nucleolar chromatin or to nucleolus-derived structures. It is also reported the presence of complex aggregates of neighboring pores associated with intranuclear filaments in tetraploid nuclei. The morphology of the fibrillar network found in spermatocyte nucleus was compared with feSEM images of pore-linked filaments described in Xenopus oocyte nucleus. Moreover, it is discussed whether the complex aggregates of neighboring pores and intranuclear filaments might be a fenotype of low metabolic activity in the 4n, as known in pore clustering of other cells.
MATERIALS AND METHODS
Spermatocytes and isolated nuclei spreads were obtained from the Brazilian Odontophrynus americanus, two natural cryptic diploid and tetraploid species (2n=22, 4n=44; State of São Paulo). To get isolated cells the testis were surgically removed and small pieces of seminiferous tubules were placed in 0.6% or 0.7% NaCl solution for 1 to 5 min and squashed on a slide. A drop of this suspension was spread on bidistilled water (adjusted to pH 7.0 with sodium borate buffer, 1 to 5 min) in a plastic tray with Teflon bars and collected on 1.5% Parlodium - covered grids. For nuclei isolation the following procedures were used: Small pieces of seminiferous tubules were homogenized with a 0.25M sacarose + 2mMCa++ (or Mg++) solution and filtered through eight layers of tissue cloth. Aliquots of 15 ml of this cellular suspension were put into centrifuge tubules adding equal volume of a 0.35M sacarose solution underlay and centrifuged at 300g for 5-10 min. After repeating this step small pieces of the pellet were left shortly in 0.1M KCl, transferred for 1min to 5:1 of 0.1M KCl + 10-3M Ca (NO3)2 and spread as described above. The grids were stained with aqueous phosphotungstic acid (PTA, 1%, 5 min) at room temperature, or aqueous uranyl acetate (1%, 10 min) then washed in bidistilled water and air-dried. Other grids were positively stained in ethanolic uranyl acetate (1:3 aqueous uranyl acetate plus 95% ethanol, 1 min) and then washed in 95% ethanol and air-dried. Following these procedures, some specimens were shadowed with Paladium (10º angle). For specimen support we used carbon/colloidon-coated cooper grids. Before use the grids were glow-discharged to render then hydrophilic. The following enzymes were used in some specimens before staining:
1. DNase (Worthington), electrophoretically purified deoxyribonuclease (DPFF) RNase free, 50µg/ml acetate buffer with 1mM MgCl2, pH 7.0, for 30-60 sec at 37ºC;
2. RNase A, protease free, type XII - A, Sigma Chem. Company, 100µg/ml in distilled water, heated to 80ºC for 10min. The grids were treated at 37ºC for 30-60 sec; Trypsin - BDH Chemicals Ltd. Pools, England, 0.01% in phosphate buffer, pH 6.8, at 37ºC, 30-60 sec. Electron micrographs were obtained with a 109 Zeiss electron microscope at 50kV and Siemens Elmiskop I, 60kV. All animals were anesthetized before death.
RESULTS
Long intranuclear filaments were observed in unfixed spreads of isolated nuclei or of whole spermatocytes from both 4n and 2n species. The filaments with 20-40 nm in width are attached to a NPC ring of about 90-120 nm and extended up to 1µm (or more) into the nucleus. They appear associated with chromatin fibers (Figure 1). Smaller NPCs diameters of 60 nm were also found.
The long filaments have a tortuous contour and a variable width due to the presence of segments of 6 nm intercalated with expanded regions of 18-20 nm or more up to 40 nm (Figure 2A). These filaments are insensitive to DNase and RNase treatments (Figure 2B). At nucleus periphery the NPCs were visualized with six or more filaments (Figure 2C). In this nuclear region RNP particles appear aligned along thinner filaments that converge to NPCs. These thin filaments seem to be either stretched intranuclear filaments or residues of collapsed NL (Figure 2D).

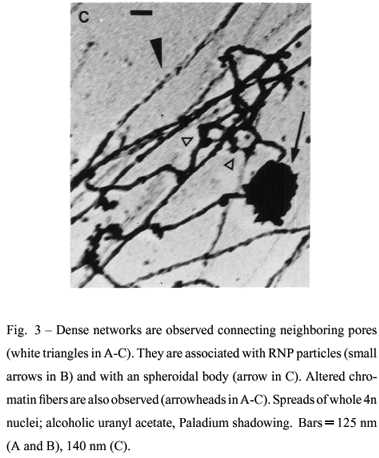
Very electron dense aggregates of intranuclear filaments were also observed in nucleus periphery connecting neighboring NPCs. They appear associated with stretched and altered chromatin fibers (Figures 3, A-C). These aggregates may appear associated with RNP particles of 20 nm-25 nm (Figure 3B) and with spheroidal structures of 0.5µm-1.2µm (Figure 3C). Smaller spheroidal structures with 100 nm-400 nm were also observed. This reduced size can be attributed either to differences of compactness, unfolding or to distinct structures. The dense aspect of the filaments is probably due to the collapse of either eventual subfibrillar components or RNP particles caused by spreading forces. The high compactness of the filaments hinders a clear visualization of individual pores. The aggregates of pores connected by filaments are more commonly found in the 4n species than in the diploids.
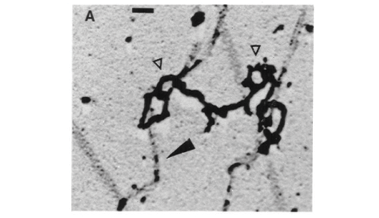
We also observed that the intranuclear filaments form a fibrous network associated with chromatin fibers. The filaments are curved due to serial circumvolutions and connect NPCs. Several filaments may converge to a same NPC (Figure 4) and exhibit RNP particles (Figure 5). The fibrous network can be observed as patches parallel to the chromatin fibers (Figures 4 and 5) or disrupted into filaments at nucleus periphery (Figure 2D).
DISCUSSION
We report here the visualization of pore-linked filaments up to 1µm long (or more) in spermatocyte nuclei belonging to diplo and tetraploid frogs (Beçak et al. 1967) using whole mount preparations for TEM. The tetraploid animals have gene mechanisms reducing metabolic activities revealed either by their low amount of total RNA (Beçak and Goissis 1971) even though 4n present double the number of 18S and 28S ribosomal genes as compared to diploids (Schmidtke et al. 1976, Ruiz et al. 1981) or by the low expression of several enzymes and hemoglobin in the tetraploid (Beçak and Pueyo 1970, Cianciarullo et al. 2000). Probably a presumed higher level of methylation of ribosomal genes would be accounted for the reduction of gene activity at the transcription level (Ruiz and Brison 1989). Based on results by different authors it was proposed that gene dose combined with transcription control show epigenetic effects on gene regulation (reviewed by Beçak and Setsuo-Kobashi 2004). Moreover, the number of NPCs in these anurans was estimated in red blood cells by freeze - etching experiments. These assays were addressed to assess the presumed functional role of NPCs in tetraploids with reduced metabolic activities. The data showed that the number of NPCs is not directly proportional to the DNA content but to the nuclear surface which in the 4n is only 1.6 times larger than in the 2n. This fact suggested that the number of NPCs could be related to transcription activity or to the transport of products to the cytoplasm (Maul et al. 1980).
The intranuclear filaments here described in both 2n and 4n species assemble in a network that connect NPCs and are associated with chromatin fibers, spheroidal structures and RNP particles. The nature of both RNP particles and spheroidal structures are at present a matter of speculation. Our morphological data suggest that the spheroidal elements found may correspond to nucleolar genes or to nucleolus-derived structures. Some subnuclear organelles termed paranucleolar or coiled bodies or nucleolus-derived structures have also comparable morphology. Among these the snurposomes b described in loops of lampbrush chromosomes from amphibian oocytes range in size from less than 1µm to about 4µm. They have splicing sn RNAs and several proteins associated with pre-mRNAs (Gall 1991). Further studies are therefore needed to clarify the identidy of the spheroidal bodies here described.
In summary, our data indicate that the intranuclear filaments may correspond to the fibrillar network previously described in anuran oocyte nuclei with other procedures (Franke and Scheer 1970, Goldberg and Allen 1992, Arlucea et al. 1998, Kiseleva et al. 2004). To our knowledge this is the first time such intranuclear filaments have been reported in spermatocytes. We also suggest that these filaments may be related to the transport of RNA precursors, probably rRNA precursors. Our data also agrees with the idea that this network may serve as ''tracks'' for nucleocytoplasm transport (Arlucea et al. 1998, Kiseleva et al. 2004). The presence of thinner and wider filaments fits well with the idea that groups of eight filaments form cylinders leading to each NPC (Arlucea et al. 1998, Kiseleva et al. 2004).
In addition, the overall aspect of the pore-linked filaments seems to be similar in both 2n and 4n species studied. A possible difference among these species is the common presence of aggregates of pores connected by intranuclear filaments in the tetraploid animals. Previous data showed that the frequency of clusters with four pores increases in these polyploid anurans (Maul et al. 1980). We do not know whether the complex aggregates of neighboring NPCs associated with the intranuclear filaments obtained in the spreads might correspond to the pore clustering described in these 4n frogs using freeze-etching experiments (Maul et al. 1980). This is an interesting question considering that the number of NPCs in the 4n frogs with reduced metabolism seems to be related to transcription activity or to the export (Maul et al. 1980). Moreover, pore clustering is either an early feature of apoptotic cells or of Nup 153 depleted cells with altered import mechanisms (Reipert et al. 1999, Walther et al. 2001). Based in these findings it is reasonable to speculate whether clustered-like pores may indicate disturbances of nucleocytoplasm transport in the tetraploid frogs. The idea that pore clustering phenotype may denote defects of transport machinery infers that the biosynthetic activities in the nucleus and cytoplasm are probably dependent of genes controlling the transport machinery (Paschal 2002).
This paper suggests the utilization of the diplo-tetraploid species of O. americanus to check the eventual relation of pore-linked filaments with gene regulation, considering the reduced metabolism of the 4n.
ACKNOWLEDGMENTS
Special thanks are due to Rita de Cassia Stocco dos Santos for the use of laboratory facilities. Many thanks to Carolina da Paz Sabino and Helir Serralvo for help in the manuscript and in photography. This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP).
Manuscript received on July 27, 2005; accepted for publication on February 17, 2006 presented by WILLY BEçAK
Correspondence to: Dra. Maria Luiza Beçak
E-mail: mlbecak@yahoo.com.br
- AFZELIUS BA. 1955. The ultrastructure of the nuclear membrane of the sea urchin oocyte as studied with the electron microscope. Exp Cell Res 8: 147-158.
- AKEY CW. 1989. Interaction and structure of the nuclear pore complex revealed by cryo-electron microscopy. J Cell Biol 109: 955-970.
- ALLEN TD, CRONSHAW JM, BAGLEY S, KISELEVA E AND GOLDBERG MW. 2000. The nuclear pore complex: mediator of translocation between nucleus and cytoplasm. J Cell Sci 113: 1651-1659.
- ARÉCHAGA J AND BAHR GF. 1985. Nuclear-rings: Portarit of a new organelle. In: SMUCKLER EA AND CLAWSON GA (Eds), Nuclear Envelope Structure and RNA maturation. Alan R Liss, New York, USA, p. 23-50.
- ARLUCEA J, ANDRADE R, ALONSO R AND ARÉCHAGA J. 1998. The nuclear basket of the nuclear pore complex is part of a higher-order filamentous network that is related to chromatin. J Struct Biol 124: 51-58.
- BEÇAK ML AND SETSUO-KOBASHI L. 2004. Evolution by polyploidy and gene regulation in Anura. Genet Mol Res 3: 195-212.
- BEÇAK ML, BEÇAK W AND RABELLO MN. 1967. Further studies on polyploid amphibians (Ceratophrydidae). I. Mitotic and meiotic aspects. Cromossoma 22: 192-201.
- BEÇAK W AND GOISSIS G. 1971. DNA and RNA content in diploid and tetraploid amphibians. Experientia 27:345-346.
- BEÇAK W AND PUEYO MT. 1970. Gene regulation in the polyploid amphibian Odontophrynus americanus Exp Cell Res 63: 448-451.
- CIANCIARULLO AM, NAOUM PC, BERTHO AL, KOBASHI LS, BEÇAK W AND SOARES MJ. 2000. Aspects of gene regulation in the diploid and tetraploid Odontophrynus americanus (Amphibia, Anura, Leptodactylidae). Gen Mol Biol 23: 357-364.
- CORDES VC, REIDENBACH S, KÖHLER A, STUURMAN N, DRIEL R VAN AND FRANKE WW. 1993. Intranuclear filaments containing a nuclear pore complex protein. J Cell Biol 123: 1333-1344.
- CORDES VC, REIDENBACH S, RACKWITZ H-R AND FRANKE WW. 1997. Identification of protein p270/Tpr as a constitutive component of the nuclear pore complex - attached intranuclear filaments. J Cell Biol 136: 515-529.
- DUPRAW EJ. 1965. The organization of nuclei and chromosome in honeybee embryonic cells. Proc Natl Acad Sci, USA 53: 161-168.
- FRANKE WW AND SCHEER U. 1970. The ultrastructure of the nuclear envelope of amphibian oocytes: a reinvestigation. I The mature oocyte. J Ultra-struct Res 30: 288-316.
- GALL JG. 1991. Spliceosomes and Snurposomes. Science 252: 1499-1500.
- GOLDBERG MW AND ALLEN TD. 1992. High resolution scanning electron microscopy of the nuclear envelope: demonstration of a new, regular, fibrous lattice attached to the baskets of the nucleoplasmic face of the nuclear pores. J Cell Biol 119: 1429-1440.
- GOLDBERG MW, SOLOVEI I AND ALLEN TD. 1997. Nuclear pore complex structure in birds. J Struct Biol 119: 284-294.
- JARNIK M AND AEBI U. 1991. Toward a more complete 3-D structure of the nuclear pore complexe. J Struct Biol 107: 291-308.
- KISELEVA E, DRUMMOND SP, GOLDBERG MW, RUTHERFORD SA, ALLEN T AND WILSON KL. 2004. Actin- and protein-4.1 - containing filaments link nuclear pore complexes to subnuclear organelles in Xenopus oocyte nuclei. J Cell Sci 117: 1-10.
- LAWRENCE JB, SINGER RH AND MARSELLE LM. 1989. Highly localized tracks of specific transcripts within interphase nuclei visualized by in situ hybridization. Cell 57: 493-502.
- MAUL GG, DEAVEN LL, FREED JJ, CAMPBELL GLE AND BEÇAK W. 1980. Investigation of the determinants of the nuclear pore number. Cytogenet Cell Genet 26: 175-190.
- NIGG EA. 1997. Nucleocytoplasmic transport: signals, mechanisms and regulation. Nature 386: 779-787.
- PANTÉ N AND AEBI U. 1996. Toward the molecular dissection of protein import into nuclei. Curr Opin Cell Biol 8: 397-406.
- PANTÉ N, BASTOS R, MCMORROW I, BURKE B AND AEBI U. 1994. Interaction and three-dimensional localization of a group of nuclear pore complex proteins. J Cell Biol 123: 603-617.
- PASCHAL BM. 2002. Translocation through the nuclear pore complex. Trends in Biochem Sci 27: 593-596.
- REIPERT S, BENNION G, HICKMAN JA AND ALLEN TD. 1999. Nucleolar segregation during apoptosis of haemopoietic stem cell line FDCP-Mix. Cell Death Differ 6: 334-341.
- RIS H. 1989. Three-dimensional structure of the nuclear pore complex. J Cell Biol 109: 134a.
- RIS H. 1991. The three-dimensional structure of the nuclear pore complex as seen by high voltage electron microscopy and high resolution low voltage scanning electron microscopy. Electron microscopy of nuclear pore complex. Scanning 19: 368-375.
- RUIZ IRG AND BRISON O. 1989. Methylation of ribosomal cistrons in diploid and tetraploid Odontophrynus americanus (Amphibia, Anura) Chromosoma 98: 86-92.
- RUIZ IRG, SOMA M AND BEÇAK W. 1981. Nucleolar organizer regions and constitutive heterochromatin in polyploid species of the genus Odontophrynus (Amphibia, Anura). Cytogenet Cell Genet 29: 84-98.
- SCHEER U, DABAUVALLE M-C, MERKERT H AND BENEVENTE R. 1988. In: NUCLEO-CYTOPLASMIC TRANSPORT, PETERS R (Ed), London, Academic Press, p. 5-25.
- SCHMIDTKE J, BEÇAK W AND ENGEL W. 1976. The reduction of gene activity in the tetraploid Odontophrynus americanus is not due to loss of ribosomal DNA. Experientia 32: 27-28.
- SUKEGAWA J AND BLOBEL G. 1993. A nuclear pore complex protein that contains zinc finger motifs, binds DNA, and faces the nucleoplasm. Cell 72: 29-38.
- UNWIN PNT AND MILLIGAN. 1982. A large particle associated with the perimeter of the nuclear pore complex. J Cell Biol 93: 63-75.
- WALTHER TC, FORNEROD M, PICKERSGILL H, GOLDBERG M, ALLEN TD AND MATTAJ W. 2001. The nucleoporin Nup 153 is required for nuclear pore basket formation, nuclear pore complex anchoring and import of a subset of nuclear proteins. EMBO J 20: 5703-5714.
- ZACHAR Z, KRAMER J, MIMS IP AND BINGHAM PM. 1993. Evidence for channeled diffusion of pre-m RNAs during nuclear RNA transport in Metazoans. J Cell Biol 121: 729-742.
- ZIMOWSKA G, ARIS JP AND PADDY MR. 1997. A Drosophila Tpr protein homolog is localizated both in the extrachromosomal channel network and to nuclear pore complexes. J Cell Sci 110: 927-944.
Publication Dates
-
Publication in this collection
23 Mar 2007 -
Date of issue
Mar 2007
History
-
Accepted
17 Feb 2006 -
Received
27 July 2005