Abstracts
A unique site at the northern area of Patagonia (Neuquén, Argentina) reveals a terrestrial ecosystem preserved in a detail never reported before in a Late Cretaceous deposit. An extraordinary diversity and abundance of fossils was found concentrated in a 0.5 m horizon in the same quarry, including a new titanosaur sauropod, Futalognkosaurus dukei n.gen., n.sp, which is the most complete giant dinosaur known so far. Several plant leaves, showing a predominance of angiosperms over gymnosperms that likely constituted the diet of F. dukei were found too. Other dinosaurs (sauropods, theropods, ornithopods), crocodylomorphs, pterosaurs, and fishes were also discovered, allowing a partial reconstruction of this Gondwanan continental ecosystem.
Dinosauria; Titanosauria; ecosystem; taphonomy; Upper Cretaceous; Gondwana; Patagonia; Argentina
Um depósito fóssil na região norte da Patagônia (Neuquén, Argentina) revela um ecossistema nunca antes registrado a este nível de detalhes em depósitos do Cretáceo Superior. Uma diversidade e abundância extraordinária de fósseis encontra-se concentrada em uma camada de 0,5 m no mesmo sítio, incluindo um novo saurópodo titanossaurídeo, Futalognkosaurus dukei n. gen, n. sp., que é o mais completo dinossauro gigante encontrado até a presente data. Foram descobertas váriasfolhas de plantas indicando a predominância de angiospermas sobre gimnospermas que possivelmente formavam a base da dieta de F. dukei. Outros dinossauros (saurópodes, terópodes, ornitópodes), crocodilomorfos, pterossauros e peixes foram também encontrados possibilitando a reconstrução parcialdeste ecossistema continental do Gondwana.
Dinosauria; Titanosauria; ecossistema; tafonomia; Cretáceo Superior; Gondwana; Patagônia; Argentina
EARTH SCIENCES
A new Cretaceous terrestrial ecosystem from Gondwana with the description of a new sauropod dinosaur
Jorge O. CalvoI; Juan D. PorfiriI; Bernardo J. González-RigaII; Alexander W.A. KellnerIII, * * Member Academia Brasileira de Ciências
ICentro Paleontológico Lago Barreales, Universidad Nacional del Comahue Proyecto Dino, Ruta Prov. 51, km 65, (8300) Neuquén, Argentina
IILaboratorio de Paleovertebrados, IANIGLA, CRICYT, CONICET, Avda. Ruiz Leal s/n, Parque Gral. San Martín (5500) Mendoza, Argentina/ICB, Universidad Nacional de Cuyo
IIIDepartamento de Geologia e Paleontologia, Museu Nacional, Universidade Federal do Rio de Janeiro Quinta da Boa Vista, São Cristóvão, 20940-040 Rio de Janeiro, RJ, Brasil
Correspondence to Correspondence to: Jorge O. Calvo E-mail: jorgecalvo@digimedia.com.ar
ABSTRACT
A unique site at the northern area of Patagonia (Neuquén, Argentina) reveals a terrestrial ecosystem preserved in a detail never reported before in a Late Cretaceous deposit. An extraordinary diversity and abundance of fossils was found concentrated in a 0.5 m horizon in the same quarry, including a new titanosaur sauropod, Futalognkosaurus dukei n.gen., n.sp, which is the most complete giant dinosaur known so far. Several plant leaves, showing a predominance of angiosperms over gymnosperms that likely constituted the diet of F. dukei were found too. Other dinosaurs (sauropods, theropods, ornithopods), crocodylomorphs, pterosaurs, and fishes were also discovered, allowing a partial reconstruction of this Gondwanan continental ecosystem.
Key words: Dinosauria, Titanosauria, ecosystem, taphonomy, Upper Cretaceous, Gondwana, Patagonia, Argentina.
RESUMO
Um depósito fóssil na região norte da Patagônia (Neuquén, Argentina) revela um ecossistema nunca antes registrado a este nível de detalhes em depósitos do Cretáceo Superior. Uma diversidade e abundância extraordinária de fósseis encontra-se concentrada em uma camada de 0,5 m no mesmo sítio, incluindo um novo saurópodo titanossaurídeo, Futalognkosaurus dukei n. gen, n. sp., que é o mais completo dinossauro gigante encontrado até a presente data. Foram descobertas váriasfolhas de plantas indicando a predominância de angiospermas sobre gimnospermas que possivelmente formavam a base da dieta de F. dukei. Outros dinossauros (saurópodes, terópodes, ornitópodes), crocodilomorfos, pterossauros e peixes foram também encontrados possibilitando a reconstrução parcialdeste ecossistema continental do Gondwana.
Palavras-chave: Dinosauria, Titanosauria, ecossistema, tafonomia, Cretáceo Superior, Gondwana, Patagônia, Argentina.
INTRODUCTION
In March 2000 a very large sauropod cervical vertebra was found in the northern shore of Lake Barreales, some 90 km northwest of Neuquén city, Northwestern Argentina (Fig. 1A). Since then, over 1000 specimens (including 240 fossil plants and 300 teeth) have been collected in an area of about 400 m2 that is called the Futalognko site. The importance of this region led to the establishment of a study center - the Centro Paleontológico Lago Barreales (CePaLB) - that carries out controlled excavations. The sedimentary rocks at this site represent the lower portion of the Upper Cretaceous (Turonian-Coniacian) Portezuelo Formation of the Neuquén Group (Leanza and Hugo 2001) and they are composed mainly of siltstones and sandstones. Fossils are found associated and partially articulated in the same quarry, particularly in a 0.5 m level of siliciclastic fluvial facies (Fig. 1B,C).

Here we describe a new sauropod dinosaur which is the most complete of the giant dinosaurs, and provide information about the Cretaceous terrestrial ecosystem present in this region of former Gondwana.
SYSTEMATIC PALEONTOLOGY
Saurischia Seeley 1887
Sauropodomorpha Huene 1932
Sauropoda Marsh 1878
Titanosauria Bonaparte and Coria 1993
Lognkosauria new taxon
ETYMOLOGY
Lognko from the Mapuche language meaning chief + saurus, from the Greek language meaning lizard.
DEFINITION
Lognkosauria is phylogenetically defined as the most recent common ancestor of Mendozasaurus neguyelap and Futalognkosaurus dukei and all its descendants.
Futalognkosaurus dukei gen. et sp. nov.
ETYMOLOGY
From the Mapuche indigenous language: Futa meaning giant and lognko meaning chief; saurus from the Greek language meaning lizard; and dukei, in honor of the Duke Energy Argentina Company that sponsored theexcavation (2002-2003).
HOLOTYPE
Atlas, axis, five anterior, four middle, and three posterior cervicals; 10 dorsals; ribs; complete sacrum; both ilia; right pubis and ischium; and one anterior caudal. All material is housed at the CePaLB-Universidad del Comahue, Neuquén, Argentina (MUCPv-323; Fig. 2).
HORIZON, AGE AND LOCALITY
Portezuelo Formation, Rio Neuquén Subgroup, Neuquén Group, Late Cretaceous, Turonian-Coniacian (Leanza and Hugo 2001). The material comes from the northern coast of the Lake Barreales, 90 km northwest of Neuquén City, Neuquén Province, Patagonia, Argentina.
DIAGNOSIS
Neurapophysis of the atlas laminar and rectangular, and posteriorly directed; neural spine of the axis high and triangular; posterior border of the neural spine on middle cervical elements concave; ventral depression between parapophyses on middle cervical centra; anterior dorsal vertebrae with horizontal and aliform diapophyses; pre- and postzygapophyses of anterior dorsal vertebrae horizontal; first caudal vertebra with prespinal lamina bifurcated on its base forming two small infraprespinal laminae; supraspinal cavity in first caudal vertebra bordered by the prespinal and lateral laminae; 2 and 3 sacral ribs fused; wide and well developed iliac peduncle on ischia.
DESCRIPTION AND COMPARISON
Futalognkosaurus dukei is the most complete giant sauropod ever found, with an estimated length between 32 to 34 meters (Calvo 2006). The atlas has a laminar,rectangular neurapophysis that expands upward andcurves medially. The axis has a high and robust neural spine with triangular shape. The centrum lacks pleurocoels differing in this respect from Saltasaurus (Powell 2003) and Alamosaurus (Lehman and Coulson 2002). All cervicals have a triangular shaped neural spine that is compressed lateromedially and elongated anteroposteriorly. Like in Malawisaurus dixeyi (Jacobs et al. 1993) and Gondwanatitan faustoi (Kellner and Azevedo 1999), the cervicals lack pleurocoels. Middle cervicals bear high and sail-shaped neural spines as in Malawisaurus and Rapetosaurus (Rogers and Forster 2001). Futalognkosaurus shares with Rapetosaurus neural arches three times higher than the centra in anterior and middle cervicals. In lateral view, the spinoprezygapophyseal border is straight and the spinopostzygapophyseal margin is concave, a feature not generally observed in other members of the Titanosauria. Posterior cervicals have neural arches at least three times higher than the centrum and neural spines compressed proximodistally and expanded laterally as in Mendozasaurus neguyelap (González Riga 2003, 2005). No prespinal lamina as the one reported in Isisaurus (Jain and Bandyopadhyay 1997) is present. A well-developed and deep supradiapophyseal cavity (or fossa) is present in Futalognkosaurus dukei and Mendozasaurus neguyelap (González Riga 2005). Sauroposeidon proteles (Wedel et al. 2000) differs from Futalognkosaurus dukei because it has cervical centra extremely elongated with low neural arches.
The dorsal vertebrae of Futalognkosaurus dukeilack hyposphene-hypantrum, which are present the giant Argentinosaurus huinculensis (Bonaparte and Coria 1993). Diapophyses are laminar, planar and directedlaterally. Anterior dorsals have neural spine formed by a spinodiapophyseal and a spinopostzygapophyseal laminae; no spinoprezygapophyseal lamina is present. Prezygapophyses are placed almost horizontally, differingfrom the inclined condition observed in Mendozasaurus and Argentinosaurus. Futalognkosaurus has the 2nd and 3rd sacral ribs fused unlike any other Titanosauria. The 1st caudal is strongly procoelous with tip of the neural spine expanded. The prespinal lamina is strongly developed and bifurcate on its base forming two small infraprespinal laminae. On anterior view, there are two deep supraspinal cavities on the neural spine. The right pubis (137 cm long) is a robust bone, more than in most other titanosaurids.
PHYLOGENETIC RELATIONSHIPS
The ingroup relationships of the Titanosauria have been extensively discussed in the last years (Salgado et al. 1997, Wilson and Sereno 1998, Upchurch 1998, Wilson and Upchurch 2003, Wilson 2006, among others). Some anatomical features such as the procoelic condition of the caudal vertebra, allows the allocation of Futalognkosaurus dukei in the Titanosauria. In order toassess the phylogenetic position of Futalognkosaurus dukei relative other titanosaurs, we have used a datamatrix (65 characters) proposed by Calvo et al. (in press). In this study we analyze the relation of Futalognkosaurus with 18 taxa using the program PAUP, version 3.0 (Swofford 1989). Camarasaurus grandis Cope 1877 was assigned as outgroup (Fig. 3) and all multi-state characters were considered unordered (see appendicesappendices). The application of the heuristic method with delayed transformation optimization produced one most parsimonious tree (Fig. 3) with a length of 108 steps and high consistency and retention index (C.I. = 0.722; R.I. = 0.780).
The analysis supports the Titanosauria sensu Bonaparte and Coria (1993), defined as the most recent common ancestor of Andesaurus delgadoi and Titanosauridae and all its descendants (Salgado et al. 1997). Futalognkosaurus can be clearly diagnosed as a Titanosauridae (sensu Salgado et al. 1997) or Lithostrotia ( sensu Wilson and Upchurch 2003). The present phylogenetic analysis also shows Malawisaurus as the sister group of Mendozasaurus (sensu González Riga 2003). Mendozasaurus is the sister group of Futalognkosaurus, forming a new clade named Lognkosauria nov., which is supported by five synapomorphies (see character list): (15.1) presence of a laterally expanded posterior cervical neural spines, wider than the centra, (18.1) posterior cervical vertebrae, with a height 1.5 the length of the centra, (19.2) deep and extended supradiapophyseal cavity in posterior cervical vertebrae, (20.2) posterior cervical centra proportions: ratio anteroposterior length/heightof posterior face less than 1,5, and (42.1) anteriormost caudal vertebrae with neural spines transversely elongated. This node has a high bootstrap value (88%) and it is well supported.
DISCUSSION
Futalognkosaurus dukei n.gen, n.sp. has a size comparable to Argentinosaurus huinculensis (Bonaparte andCoria 1993) and Puertasaurus reuili (Novas et al. 2005), up to date, considered the largest of all sauropods but, unfortunately, known by few and incomplete elements. The discovery of Futalognkosaurus and Mendozasaurus indicates the presence of a new lineage of titanosaurs, with strong and huge necks, differing from the remaining members of this group, increasing the diversity of those large dinosaurs that once roamed the Earth.
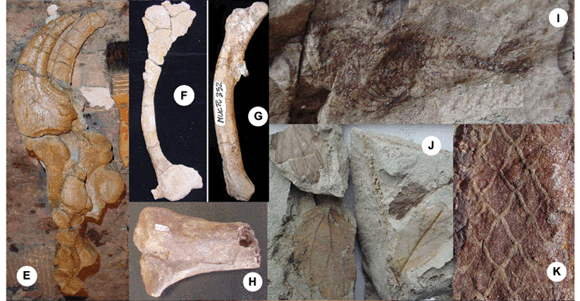
Besides Futalognkosaurus, other sauropod dinosaur remains that represent two additional yet undescribed taxa were also found in the Futalognko site, but none was articulated or nearly as complete. Theropod specimens were also unearthed and indicate the presence of several species, two of which were formally described. One represents Megaraptor namunhuaiquii, previously recovered in the Sierra del Portezuelo, around 80 km South from the Futalognko site, in levels of the Portezuelo Formation (Novas 1998). The material of Megaraptor collected at the Futalognko site consists of a complete and articulated arm (Fig. 4E), showing that the bones originally interpreted as a foot of this taxon (Novas 1998) are, in fact, the hand (Calvo et al. 2004b). The new specimen demonstrates that Megaraptor is not a member of the Coelurosauria, but a basal tetanuran that developed a unique elongated hand with very large sickle-shaped claws, more so than the ones of spinosaurids (Charig and Milner 1997).

The second theropod belongs to the genus Unenlagia that was also first described from the Sierra del Portezuelo (U. comahuensis). This genus has been regarded as the sister group of Aviale ( Archaeopteryx + modern birds) (Novas and Puerta 1997) while others suggest that it was a more basal dromaeosaurid (Norell et al. 2001). The new species recovered from the Futalognko quarry, named Unenlagia paynemili, belongs to a more slender species that has a small anterior process in the pubis (not present in U. comahuensis) and indicates that those enigmatic Unenlagia-type maniraptoran dinosaurs were more common (Calvo et al. 2004a,Fig. 4F). Additional theropod material consists of isolated bones and over 250 teeth that show several morphotypes, suggesting the presence of a rather diverse theropod fauna in this quarry (study in progress). Most are very similar to those reported in dromaeosaurids (Fig. 4B) but some show wrinkled enamel close to the margin of the teeth which are typical of carcharodontosaurids (Sereno et al. 1996, Kellner and Campos 2000) (Fig. 4C).
Remains of ornithischian dinosaurs were also collected in the quarry and consist of isolated bones (dorsal vertebra, femur, and ilium) that are referable to Ornithopoda, which are rare in South America (Fig. 4G). Based on the separated distal condyles of the femur,they are referable to the Iguanodontia.
Other fossil reptiles are crocodylomorph teeth,some referable to Peirosauridae (Fig. 4D), which were found in the Neuquén Group (Digregorio 1972), Morocco (Buffetaut 1994) and Brazil (Price 1955). Recently a lower jaw of a tiny individual was also recovered (Fig. 4L) and belongs to a new species closely related to Comahuesuchus. The new specimen shows that those rare and unusual notosuchid crocodylomorphs have a large posterior tooth that shows coarse serrations similar to theropod dinosaurs, increasing the groups of crocodylomorphs with "theropod-like" teeth (Campos et al. 2001). Among the pterosaur bones recovered is the proximal part of an ulna referable to the Azhdarchoidea sensu Kellner (2003), with an estimated wing span of around 6 m (Kellner et al. 2007, Fig. 4H). Histological sections show the presence of reticular fibro-lamellar bone, with numerous vascular canals, indicating that it was still growing at time of death and therefore can be considered the largest pterosaur recovered from the former Gondwana. Remains of pelomedusoid turtles and undetermined reptilian eggshells were also found.
Unexpectedly, the Futalognko quarry has also yielded delicate fish specimens; the first from the Neuquén Group reported so far (Fig. 4I). There are six specimens representing three different taxa: a small Euteleostei and two clupeomorphs (Gallo et al. 2003). Despite not being complete, all preserved elements are articulated. Ganoid scales of semionotid fishes and freshwater bivalves were also collected.
Lastly, about 240 plant specimens were recovered. Leaves are well preserved, with the majority corresponding to dicotyledonian angiosperms that are shown tohave been abundant in this area (Fig. 4J,K). Some gymnosperm leaves, fructifications and remains of conifers were found too. In this wide diversity of foliar structures there are indications of herbaceous habit for some specimens as well as signs of caducity. Therefore, in this complex association, angiosperms seem to constitute the main element of the ancient flora in this region, and this material consists the first record of those derived plants in the Upper Cretaceous of the Neuquén Basin (Passalia et al., in press).
The fossil content in this region is not limited to the Futalognko quarry. In a range of 1500 m, there are three more quarries at the same level, that have also yielded a variety of specimens, including an articulated partial skeleton of a new theropod (Fig. 4M) and isolated remains of ornithopods, including jaws (Fig. 4N). Very well preserved pleurodiran turtles (including skull) were also found. Two other sites have been discovered in the Plottier Formation that overlies the Portezuelo layers, only 25 m above the Futalognko quarry and also contain large quantities of dinosaur bones, particularly sauropods, bivalves and plant remains, albeit in lower diversity.
Other terrestrial important and rich Cretaceous ecosystems such as the Jehol Biota of China (Chang et al. 2003) and the Santana Formation of Brazil (Maisey 1991, Kellner et al. 1994, Kellner and Campos 1999), have been reconstructed based on fossils collected in large areas and from different layers whose outcrops sometimes are hundreds of meters apart and possible represent a time span of a few million years. The excavated part of the Futalognko site is restricted to a small area (400 m2) and due to a controlled collecting program in which all specimens are plotted on a quarry map, it can be demonstrated that the fossils were found in a bed about 0.5 m thick, representing a time span in the scale of years. We were also able to reconstruct the taphonomic history of this unusual deposit.
Based on geological data, the Portezuelo Formation was deposited in a flat landscape as most of the stratigraphic units of the Neuquén Group. A detailed sedimentological facies analysis (Sánchez et al. 2005) shows that an active and wide meandering river system was installed in this area during the late Cretaceous (Turonian-Coniacian). Moreover, the cyclic deposition of different fluvial facies suggests that the climate was warm and humid, intercalated with short dry seasons.
Three different sedimentological facies deposits that are associated to this meandering system could be recognized: channel, point bar and small lake deposits, the later corresponding to an abandoned meander. Our sedimentological and paleontological studies show that all three facies are present at the Futalognko site (Fig. 1C). The point bar deposits were formed first. During the dry season, fine-grained sediments (that latter onebecome sandstones and show well preserved fossils)were deposited latter. The wet seasons were responsible for flooding events that carried the animal remains downstream the river channel and deposited them in fine conglomerates. Those fossil remains usually are crushed with signs of erosion indicating a moderate transportation.
Regarding the remains of Futalognkosaurus dukei we hypothesize that it died of unknown causes on afloodplain, near the margins of a river. The carcass was possibly partially dismembered by theropod dinosaurs present in this area such as Megaraptor and small dromaeosaurids (and, perhaps also some crocodylomorphs like peirosaurids). A short time after its death, a single flooding event of great intensity washed the carcass of this giant sauropod dinosaur into the river channel. Dueto its giant size the carcass likely acted as a barrier, reducing the competency of the currents generated by subsequent smaller flooding events that are very common inthe wet seasons. This resulted in the deposition of additional organic remains around this huge carcass. At some point, the river changed its course laterally, leaving an oxbow lake behind. Surprisingly some tiny fish skeletons were found together with the dinosaur elements. Since they are complete, the only explanation for their presence along with the large bones is that they were trapped when this lake was formed. The few bivalves preserved in the same horizon were trapped too.
Subsequently wind and rain carried sediments, leaves and branches from the nearby vegetation inside the small lake. Most of the plant material is carbonized indicating that a reducing environment was installed in this lake and that the original connection with the river was broken. These fossil plants are preserved in laminated and rippled siltstone around the dinosaur bones resulting in this very unusual combination of fossils. The events that lead to this accumulation must have happened in a comparatively short time otherwise, the bones of Futalognkosaurus would have been decomposed.
All this sedimentological and paleontological data make us confident to regard the plants and animals found and preserved in the Futalognko site as part of the same terrestrial ecosystem which can be reconstructed as follows. During the Late Cretaceous (Turonian-Coniancian), Patagonia had a warm and humid climate with dry seasons, and a meandering river system crossed a forest formed mainly by angiosperms, with few subordinated gymnosperms. This region was home to small ornithopods, small and large theropods (e.g., Megaraptor, Unenlagia), and giant sauropod dinosaurs such as Futalognkosaurus. Other reptiles present in this ecosystem were peirosaurid and notosuchid crocodylomorphs, pleurodiran turtles and large azhdarchoid pterosaurs. This large diversity of specimens with different potential preservation (e.g., leaves, invertebrates, delicate fish skeleton and large dinosaur bones) associated in one quarry at the same horizon was never reported in Gondwana. This unique site consists a rare occasion where a Late Cretaceous terrestrial ecosystem can be confidently reconstructed.
ACKNOWLEDGMENTS
The research activities at the Futalognko site could not have been done without the help of students and volunteers - too many to be listed here - that have workedthere since 2000. We would also like to thank MarceloN.F. Trotta (Museu Nacional/UFRJ), Valéria Gallo (Universidade do Estado do Rio de Janeiro/UERJ) andRodrigo Santucci (Departamento Nacional de Produção Mineral/Brasília) for several suggestions that improved the manuscript. We specially wish to thank Duke Energy Argentina, Duke Foundation and United Way International for developing and supporting the Proyecto Dino and the research at the new Centro Paleontológico Lago Barreales (CePaLB). This project was also partially funded by the National University of Comahue and Chevron-Texaco for the projects T-021 and I-122 (JOC), the Agencia Nacional de Promoción Científica y Tecnológica project PIDCT 07-08277 (to JOC), Pan American Energy (Proyecto Dino to JOC), Repsol - YPF (CePaLB to JOC), Daniel Andino Trailers (to JOC) and several collaborators of the Proyecto Dino. We also thank Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, proc. no. 486313/2006-9 to AWAK) and Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ, proc. no. E-26/152.885/2006 to AWAK).
REFERENCES
BONAPARTE JF AND CORIA RA. 1993. Un nuevo y gigantesco saurópodo Titanosaurio de la Formación Río Limay (Albiano-Cenomaniano) de la provincia del Neuquén, Argentina. Ameghiniana 30: 271282.
BUFFETAUT E. 1994. A new crocodilian from the Cretaceous of southern Morocco. Comptes Rendus Academy Science 319 (II):15631568.
CALVO J. 2006. Dinossauros e fauna associada de uma nova localidade no Lago Barreales (Formação Portezuelo, Cretáceo Superior), Neuquén, Argentina. Tese (doutorado) UFRJ/Museu Nacional/Programa de Pós-graduação em Ciências Biológicas, Brasil, 139 p.
CALVO JO, PORFIRI J AND KELLNER AWA. 2004a. On a new Maniraptoran Dinosaur (Theropoda) from the Upper Cretaceous of Neuquén, Patagonia, Argentina. Arquivos do Museu Nacional 62: 549566.
CALVO JO, PORFIRI JD, VERALLI C, NOVAS F AND POBLETE F. 2004b. Phylogenetic status of Megaraptor namunhuaiquii Novas based on a new specimen from Neuquén, Patagonia, Argentina. Ameghiniana 41: 565575.
CALVO JO, GONZÁLEZ RIGA BJ AND PORFIRI JD. In press. Muyelensaurus pecheni gen. et sp. nov., a new titanosaur sauropod from the Late Cretaceous of Neuquén, Patagonia, Argentina. Arquivos do Museu Nacional.
CAMPOS DA, SUAREZ JM, RIFF D AND KELLNER AWA. 2001. Short note on a new Baurusuchidae (Crocoyliformes, Metasuchia) from the Upper Cretaceous of Brazil. Boletim do Museu Nacional, Geol 57: 17.
CHANG M, CHEN PJ, WANG YQ AND WANG Y (EDS). 2003. The Jehol Biota. Shanghai Scientific & Technical Publishers, Shanghai, 208 p.
CHARIG AJ AND MILNER AC. 1997. Baryonyx walkeri, a fish-eating dinosaur from de Wealden of Surrey. Bull Nat His Mus, Geology Series 53: 1170.
DIGREGORIO JH. 1972. Neuquén. In: GEOLOGÍA REGIONAL ARGENTINA. LEANZA AF (Ed). Academia Nacional de Ciencias da Argentina (?), p. 439505.
GALLO V, CALVO JO AND KELLNER AWA. 2003. First occurrence of a teleostean fish in the Portezuelo Formation (Upper Cretaceous), Neuquén Group, Patagonia, Argentina. Resumos, III Simpósio Brasileiro de Paleontologia de Vertebrados, 29 p.
GONZÁLEZ RIGA BJ. 2003. A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza Province, Argentina. Ameghiniana 40: 155172.
GONZÁLEZ RIGA BJ. 2005. Nuevos restos fósiles de Mendozasaurus neguyelap (Sauropoda, Titanosauria) del Cretácico Tardío de Mendoza, Argentina. Ameghiniana 42: 535548.
JACOBS LL, WINKLER DA, DOWNS WR AND GOMANI EM. 1993. New material of an early Cretaceous titanosaurid sauropod dinosaur from Malawi. Palaeontology 36: 523534.
JAIN SL AND BANDYOPADHYAY S. 1997. New titanosaurid (Dinosauria: Sauropoda) from the Late Cretaceous of Central India. J Vert Paleont 17: 114136.
KELLNER AWA. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. In: BUFFETAUT E AND MAZIN JM(Eds), Evolution and Palaeobiology of Pterosaurs. Geological Society, Special Publication 217: 105137.
KELLNER AWA AND AZEVEDO SAK. 1999. A new sauropod dinosaur (Titanosauria) from the Late Cretaceous of Brazil. In: PROCEEDINGS OF THE SECOND GONDWANAN DINOSAUR SYMPOSIUM. TOMIDA Y, RICH TH AND VICKERS RICH P (Eds). National Science Museum Monographs 15: 111142.
KELLNER AWA AND CAMPOS DA. 1999. Vertebrate Paleontology in Brazil a review. Episodes 22: 238251.
KELLNER AWA AND CAMPOS DA. 2000. Brief review of dinosaur studies and perspectives in Brazil. An Acad Bras Cienc 72: 509538.
KELLNER AWA, MAISEY JG AND CAMPOS DA. 1994. Fossil down feather from the Lower Cretaceous of Brazil. Paleontology, 37: 489492.
KELLNER AWA, CALVO J, SAYÃO JM AND PORFIRI JD. 2007. Pterosaur bones from the Portezuelo Formation (Cretaceous), Neuquén Group, Patagonia, Argentina. Arquivos do Museu Nacional 64: 369375.
LEANZA HA AND HUGO CA. 2001. Cretaceous red beds from southern Neuquén Basin (Argentina): age, distribution and stratigraphic discontinuities. In: LEANZA HA (Ed), Asociación Paleontológica Argentina. VII International symposium on Mesozoic Terrestrial Ecosystems, Buenos Aires 7: 117122.
LEHMAN TM AND COULSON AB. 2002. A juvenile specimen of the sauropod dinosaur Alamosaurus sanjuanensis from the Upper Cretaceous of Big Bend National Park, Texas. J Paleont 76: 156172.
MAISEY JG (Ed). 1991. Santana fossils: an illustrated atlas. Neptune: T.F.H, 459 p.
NORELL MA, CLARK JM AND MAKOVICKY PJ. 2001. Phylogenetic relationships among coelurosaurian theropods. New Perspectives on the Origin and Early Evolution of Birds: Proceedings of the International Symposium in Honor of John H. Ostrom. Peabody Museum of Natural History, p. 4967.
NOVAS F. 1998. Megaraptor namunhuaiquii, gen. et sp. nov., a large-clawed, late cretaceous theropod from Patagonia. J Vert Paleont 18: 49.
NOVAS FE AND PUERTA P. 1997. New evidence concerning avian origins from the late Cretaceous of Patagonia. Nature 387: 390392.
NOVAS FE, SALGADO L, CALVO J AND AGNOLIN F. 2005. Giant titanosaur (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia. Rev. Del Museo de Ciencias Naturales Bernardino Rivadavia N S 7: 3741.
PASSALIA MG, PRÁMPARO MB, CALVO JO AND HEREDIA S. (in press). Primer registro de hojas de angiospermas en el Grupo Neuquén (Turoniano tardío-Coniaciano temprano), Lago Barreales, Argentina: Reporte preliminar. Ameghiniana.
POWELL JE. 2003. Revision of South American Titanosaurid dinosaurs: palaeobiological, palaeobiogeographical and phylogenetic aspects. Records of the Queen Victoria Museum 111: 1173.
PRICE LI. 1955. Novos crocodileos dos arenitos da série Bauru, Cretáceo do Estado de Minas Gerais. An Acad Bras Cienc 27: 487498.
ROGERS CK AND FOSTER CA. 2001. The last of the dinosaur titans: a new sauropod from Madagascar. Nature 412: 530534.
SALGADO L, CORIA RA AND CALVO JO. 1997. Evolution of titanosaurid sauropods I: phylogenetic analysis based on the postcranial evidence. Ameghiniana 34: 332.
SÁNCHEZ ML, CALVO JO AND HEREDIA S. 2005. Paleoambientes de sedimentación del tramo superior de la Formación Portezuelo, Grupo Neuquén, (Cretácico Superior) Los Barreales, Prov. del Neuquén. Rev. Asoc. Geológica Argentina, 60: 142158.
SERENO PC, DUTHIEL DB, LAROCHENE M, LARSSON HCE, LYON GH, MAGWENW PM, SIDOR CA, VARRICHIO DJ AND WILSON JA. 1996. Predatory dinosaurs from the Sahara and the Late Cretaceous faunal differentiation. Science 272: 986991.
SWOFFORD DL. 1998. PAUP: phylogenetic Analysis using Parsimony. Version 3,0. Sinauer Associates. Inc. Sunderland, Massachusetts.
UPCHURCH P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society. 124: 43103.
WEDEL MJ, CIFELLI RL AND SANDERS RK. 2000. Sauroposeidon proteles, a new sauropod from the Early Cretaceous of Oklahoma. J Vert Paleont 20: 109114.
WILSON JA. 2006. An Overview of titanosaur evolution and phylogeny. In: Colectivo Arqueológico-Paleontológico Salense (Ed), Actas de las III Jornadas sobre Dinosaurios y su Entorno. Burgos, p. 169190.
WILSON JA AND SERENO P. 1998. Early Evolution and Higher-level Phylogeny of Sauropod Dinosaurs. Journal of Vertebrate Paleontology. 18 (Suppl) 2: 168.
WILSON JA AND UPCHURCH P. 2003. A revision of Titanosaurus Lydekker (Dinosauria Sauropoda), the first dinosaur genus with a Gondwana distribution. J Syst Paleontol, London 1: 125160.
Manuscript received on July 12, 2007; accepted for publication on August 1, 2007; contributed by ALEXANDER W.A. KELLNER
APPENDIX 2

- BONAPARTE JF AND CORIA RA. 1993. Un nuevo y gigantesco saurópodo Titanosaurio de la Formación Río Limay (Albiano-Cenomaniano) de la provincia del Neuquén, Argentina. Ameghiniana 30: 271282.
- BUFFETAUT E. 1994. A new crocodilian from the Cretaceous of southern Morocco. Comptes Rendus Academy Science 319 (II):15631568.
- CALVO J. 2006. Dinossauros e fauna associada de uma nova localidade no Lago Barreales (Formação Portezuelo, Cretáceo Superior), Neuquén, Argentina. Tese (doutorado) UFRJ/Museu Nacional/Programa de Pós-graduação em Ciências Biológicas, Brasil, 139 p.
- CALVO JO, PORFIRI J AND KELLNER AWA. 2004a. On a new Maniraptoran Dinosaur (Theropoda) from the Upper Cretaceous of Neuquén, Patagonia, Argentina. Arquivos do Museu Nacional 62: 549566.
- CALVO JO, PORFIRI JD, VERALLI C, NOVAS F AND POBLETE F. 2004b. Phylogenetic status of Megaraptor namunhuaiquii Novas based on a new specimen from Neuquén, Patagonia, Argentina. Ameghiniana 41: 565575.
- CALVO JO, GONZÁLEZ RIGA BJ AND PORFIRI JD. In press. Muyelensaurus pecheni gen. et sp. nov., a new titanosaur sauropod from the Late Cretaceous of Neuquén, Patagonia, Argentina. Arquivos do Museu Nacional.
- CAMPOS DA, SUAREZ JM, RIFF D AND KELLNER AWA. 2001. Short note on a new Baurusuchidae (Crocoyliformes, Metasuchia) from the Upper Cretaceous of Brazil. Boletim do Museu Nacional, Geol 57: 17.
- CHANG M, CHEN PJ, WANG YQ AND WANG Y (EDS). 2003. The Jehol Biota. Shanghai Scientific & Technical Publishers, Shanghai, 208 p.
- CHARIG AJ AND MILNER AC. 1997. Baryonyx walkeri, a fish-eating dinosaur from de Wealden of Surrey. Bull Nat His Mus, Geology Series 53: 1170.
- DIGREGORIO JH. 1972. Neuquén. In: GEOLOGÍA REGIONAL ARGENTINA. LEANZA AF (Ed). Academia Nacional de Ciencias da Argentina (?), p. 439505.
- GALLO V, CALVO JO AND KELLNER AWA. 2003. First occurrence of a teleostean fish in the Portezuelo Formation (Upper Cretaceous), Neuquén Group, Patagonia, Argentina. Resumos, III Simpósio Brasileiro de Paleontologia de Vertebrados, 29 p.
- GONZÁLEZ RIGA BJ. 2003. A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza Province, Argentina. Ameghiniana 40: 155172.
- GONZÁLEZ RIGA BJ. 2005. Nuevos restos fósiles de Mendozasaurus neguyelap (Sauropoda, Titanosauria) del Cretácico Tardío de Mendoza, Argentina. Ameghiniana 42: 535548.
- JACOBS LL, WINKLER DA, DOWNS WR AND GOMANI EM. 1993. New material of an early Cretaceous titanosaurid sauropod dinosaur from Malawi. Palaeontology 36: 523534.
- JAIN SL AND BANDYOPADHYAY S. 1997. New titanosaurid (Dinosauria: Sauropoda) from the Late Cretaceous of Central India. J Vert Paleont 17: 114136.
- KELLNER AWA. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. In: BUFFETAUT E AND MAZIN JM(Eds), Evolution and Palaeobiology of Pterosaurs.
- Geological Society, Special Publication 217: 105137.
- KELLNER AWA AND AZEVEDO SAK. 1999. A new sauropod dinosaur (Titanosauria) from the Late Cretaceous of Brazil. In: PROCEEDINGS OF THE SECOND GONDWANAN DINOSAUR SYMPOSIUM. TOMIDA Y, RICH TH AND VICKERS RICH P (Eds). National Science Museum Monographs 15: 111142.
- KELLNER AWA AND CAMPOS DA. 1999. Vertebrate Paleontology in Brazil a review. Episodes 22: 238251.
- KELLNER AWA AND CAMPOS DA. 2000. Brief review of dinosaur studies and perspectives in Brazil. An Acad Bras Cienc 72: 509538.
- KELLNER AWA, MAISEY JG AND CAMPOS DA. 1994. Fossil down feather from the Lower Cretaceous of Brazil. Paleontology, 37: 489492.
- KELLNER AWA, CALVO J, SAYÃO JM AND PORFIRI JD. 2007. Pterosaur bones from the Portezuelo Formation (Cretaceous), Neuquén Group, Patagonia, Argentina. Arquivos do Museu Nacional 64: 369375.
- LEANZA HA AND HUGO CA. 2001. Cretaceous red beds from southern Neuquén Basin (Argentina): age, distribution and stratigraphic discontinuities. In: LEANZA HA (Ed), Asociación Paleontológica Argentina. VII International symposium on Mesozoic Terrestrial Ecosystems, Buenos Aires 7: 117122.
- LEHMAN TM AND COULSON AB. 2002. A juvenile specimen of the sauropod dinosaur Alamosaurus sanjuanensis from the Upper Cretaceous of Big Bend National Park, Texas. J Paleont 76: 156172.
- MAISEY JG (Ed). 1991. Santana fossils: an illustrated atlas. Neptune: T.F.H, 459 p.
- NORELL MA, CLARK JM AND MAKOVICKY PJ. 2001. Phylogenetic relationships among coelurosaurian theropods. New Perspectives on the Origin and Early Evolution of Birds: Proceedings of the International Symposium in Honor of John H. Ostrom. Peabody Museum of Natural History, p. 4967.
- NOVAS F. 1998. Megaraptor namunhuaiquii, gen. et sp. nov., a large-clawed, late cretaceous theropod from Patagonia. J Vert Paleont 18: 49.
- NOVAS FE AND PUERTA P. 1997. New evidence concerning avian origins from the late Cretaceous of Patagonia. Nature 387: 390392.
- NOVAS FE, SALGADO L, CALVO J AND AGNOLIN F. 2005. Giant titanosaur (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia. Rev. Del Museo de Ciencias Naturales Bernardino Rivadavia N S 7: 3741.
- PASSALIA MG, PRÁMPARO MB, CALVO JO AND HEREDIA S. (in press). Primer registro de hojas de angiospermas en el Grupo Neuquén (Turoniano tardío-Coniaciano temprano), Lago Barreales, Argentina: Reporte preliminar. Ameghiniana.
- POWELL JE. 2003. Revision of South American Titanosaurid dinosaurs: palaeobiological, palaeobiogeographical and phylogenetic aspects. Records of the Queen Victoria Museum 111: 1173.
- PRICE LI. 1955. Novos crocodileos dos arenitos da série Bauru, Cretáceo do Estado de Minas Gerais. An Acad Bras Cienc 27: 487498.
- ROGERS CK AND FOSTER CA. 2001. The last of the dinosaur titans: a new sauropod from Madagascar. Nature 412: 530534.
- SALGADO L, CORIA RA AND CALVO JO. 1997. Evolution of titanosaurid sauropods I: phylogenetic analysis based on the postcranial evidence. Ameghiniana 34: 332.
- SÁNCHEZ ML, CALVO JO AND HEREDIA S. 2005. Paleoambientes de sedimentación del tramo superior de la Formación Portezuelo, Grupo Neuquén, (Cretácico Superior) Los Barreales, Prov. del Neuquén. Rev. Asoc. Geológica Argentina, 60: 142158.
- SERENO PC, DUTHIEL DB, LAROCHENE M, LARSSON HCE, LYON GH, MAGWENW PM, SIDOR CA, VARRICHIO DJ AND WILSON JA. 1996. Predatory dinosaurs from the Sahara and the Late Cretaceous faunal differentiation. Science 272: 986991.
- SWOFFORD DL. 1998. PAUP: phylogenetic Analysis using Parsimony. Version 3,0. Sinauer Associates. Inc. Sunderland, Massachusetts.
- UPCHURCH P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society. 124: 43103.
- WEDEL MJ, CIFELLI RL AND SANDERS RK. 2000. Sauroposeidon proteles, a new sauropod from the Early Cretaceous of Oklahoma. J Vert Paleont 20: 109114.
- WILSON JA. 2006. An Overview of titanosaur evolution and phylogeny. In: Colectivo Arqueológico-Paleontológico Salense (Ed), Actas de las III Jornadas sobre Dinosaurios y su Entorno. Burgos, p. 169190.
- WILSON JA AND SERENO P. 1998. Early Evolution and Higher-level Phylogeny of Sauropod Dinosaurs. Journal of Vertebrate Paleontology. 18 (Suppl) 2: 168.
- WILSON JA AND UPCHURCH P. 2003. A revision of Titanosaurus Lydekker (Dinosauria Sauropoda), the first dinosaur genus with a Gondwana distribution. J Syst Paleontol, London 1: 125160.
appendices

Publication Dates
-
Publication in this collection
27 Aug 2007 -
Date of issue
Sept 2007
History
-
Accepted
01 Aug 2007 -
Received
12 July 2007