Abstracts
The infection of the estuarine teleost fishes Mugil gaimardianus Desmarest, 1831 (Mugilidae), Arius phrygiatus Valenciennes, 1839 (Ariidae), Conodon nobilis Linnaeus, 1759 (Haemulidae), Cetengraulis edentulus Cuvier, 1829 (Engraulidae), and Anableps anableps Linnaeus, 1758 (Anablepidae) by praniza larvae of Gnathiidae (Crustacea, Isopoda) was studied in specimens fished off the Atlantic Ocean in Northeast of Pará State, near Bragança, Brazil. The highest infection prevalence value was found in Anableps anableps (42.3%) and the lowest in Conodon nobilis (9.1%). The mean intensity varied from 1 parasitein Conodon nobilis to 19.5 in Arius phrygiatus. A description of the larvae is provided. The morphology of the mouthparts is related to the blood sucking activity, and is compared with the characteristics of other gnathiidae species.
fish; parasites; Isopoda; Gnathiidae; Brazil
Foi estudada a parasitose dos peixes estuarinos Mugil gaimardianus Desmarest, 1831 (Mugilidae), Arius phrygiatus Valenciennes, 1839 (Ariidae), Conodon nobilis Linnaeus, 1759 (Haemulidae), Cetengraulis edentulus Cuvier, 1829 (Engraulidae), e Anableps anableps Linnaeus, 1758 (Anablepidae) por larvas praniza de Gnathiidae (Crustacea, Isopoda) em exemplares pescados no Oceano Atlântico, no Nordeste do Pará, próximo a Bragança, Brasil. O valor mais elevado da prevalência da infecção foi observado em A. anableps (42,3%) e o menor em C. nobilis (9,1%). A intensidade média da parasitose variou entre 1 parasita em C. nobilis até 19,5 em A. phrygiatus. Efetua-se a descrição da larva, verificando-se que a morfologia da armadura bucal está relacionada com a atividade sugadora de sangue, e faz-se a comparação com as características de outras espécies.
peixe; parasita; Isopoda; Gnathiidae; Brasil
BIOLOGICAL SCIENCES
A note on the occurrence of praniza larvae of Gnathiidae (Crustacea, Isopoda) on fishes from Northeast of Pará, Brazil
Daniel G. DinizI; Jamille E.A. VarellaI; Maria Danielle F. GuimarãesI; Arthur F.L. SantosI; Rodrigo Y. FujimotoI; Karla C.F. MonfortI; Marcus A.B. PiresI; Maurício L. MartinsII; Jorge C. EirasIII
IUniversidade Federal do Pará, Campus de Bragança, Alameda Leandro Ribeiro s/n, Bairro Aldeia 68600-000 Bragança, PA, Brasil
IIUniversidade Federal de Santa Catarina, Departamento de Aqüicultura, CCA, Rodovia SC 404 Km 3 88040-900 Florianópolis, SC, Brasil
IIIDepartamento de Zoologia e Antropologia, and CIIMAR, Faculdade de Ciências, Universidade do Porto Rua do Campo Alegre s/n, Edifício FC4, 4169-007 Porto, Portugal
Correspondence to Correspondence to: Daniel Guerreiro Diniz E-mail: danielguerreirodiniz@gmail.com
ABSTRACT
The infection of the estuarine teleost fishes Mugil gaimardianus Desmarest, 1831 (Mugilidae), Arius phrygiatus Valenciennes, 1839 (Ariidae), Conodon nobilis Linnaeus, 1759 (Haemulidae), Cetengraulis edentulus Cuvier, 1829 (Engraulidae), and Anableps anableps Linnaeus, 1758 (Anablepidae) by praniza larvae of Gnathiidae (Crustacea, Isopoda) was studied in specimens fished off the Atlantic Ocean in Northeast of Pará State, near Bragança, Brazil. The highest infection prevalence value was found in Anableps anableps (42.3%) and the lowest in Conodon nobilis (9.1%). The mean intensity varied from 1 parasitein Conodon nobilis to 19.5 in Arius phrygiatus. A description of the larvae is provided. The morphology of the mouthparts is related to the blood sucking activity, and is compared with the characteristics of other gnathiidae species.
Key words: fish, parasites, Isopoda, Gnathiidae, Brazil.
RESUMO
Foi estudada a parasitose dos peixes estuarinos Mugil gaimardianus Desmarest, 1831 (Mugilidae), Arius phrygiatus Valenciennes, 1839 (Ariidae), Conodon nobilis Linnaeus, 1759 (Haemulidae), Cetengraulis edentulus Cuvier, 1829 (Engraulidae), e Anableps anableps Linnaeus, 1758 (Anablepidae) por larvas praniza de Gnathiidae (Crustacea, Isopoda) em exemplares pescados no Oceano Atlântico, no Nordeste do Pará, próximo a Bragança, Brasil. O valor mais elevado da prevalência da infecção foi observado em A. anableps (42,3%) e o menor em C. nobilis (9,1%). A intensidade média da parasitose variou entre 1 parasita em C. nobilis até 19,5 em A. phrygiatus. Efetua-se a descrição da larva, verificando-se que a morfologia da armadura bucal está relacionada com a atividade sugadora de sangue, e faz-se a comparação com as características de outras espécies.
Palavras-chave: peixe, parasita, Isopoda, Gnathiidae, Brasil.
INTRODUCTION
Gnathiid isopods are crustacea which are free-living as adults and have juvenile stages, the praniza larvae, which are temporary ectoparasites feed intermittently on the blood and tissue fluids of fishes, both elasmobranchs and teleosts (Davies 1981). There are twelve genera within the Gnathiidae family, and most of the species belong to the genus Gnathia (Smit and Davies 2004, Hadfield and Smit 2008).
The larval stages attach to the gills and skin andcan cause focal lesions on the place of attachment(Heupel and Bennett 1999, González et al. 2004, Marino et al. 2004), reduce the haematocrit of the host (Jones and Grutter 2005) or even cause the death of the host (Mugridge and Stallybrass 1983).
Apparently these parasites are not host specific and were described from a number of hosts worldwide(Smit and Basson 2002, Smit and Davies 2004, Jones and Grutter 2005, Genc et al. 2005). In some cases they can be very abundant: in Dentex dentex from the Western Mediterranean the prevalence of praniza larvae reached 85%, with a mean intensity of 20 parasites per fish (González et al. 2004), and in Epinephelus aeneus from the North-Eastern Mediterranean 57.1% of the specimens were infected presenting a mean intensity of infection of 43.5 larvae per host (Genc et al. 2005).
In this paper we present some data about the infection of fish from the Pará coastal zone by gnathiidlarvae. Moreover, we describe the morphology of the praniza larvae, and identify the most infected fish species in order to subsequently study their blood parasites since praniza larvae have been shown to transmit haemogregarines to fish (Davies 1982, Davies et al. 1994, Davies and Smit 2001), and the possibility of transmitting other blood parasites can not be ruled out.
MATERIALS AND METHODS
Specimens of the estuarine fish "tainha", Mugil gaimardianus Desmarest, 1831 (Mugilidae - 29 specimens), "peixe-gato", Arius phrygiatus Valenciennes,1839 (Ariidae - 52), "jiquiri", Conodon nobilis Linnaeus, 1759 (Haemulidae - 11), "sardinha", Cetengraulis edentulus Cuvier, 1829 (Engraulidae - 71) and "tralhoto", Anableps anableps Linnaeus, 1758 (Anablepidae - 59) were net-fished in the Atlantic Ocean, in the coastal zone of Pará State, near Bragança, Brazil, from August 2006 to February, 2007, and identified according to Santo et al. 2005. The fish were immediately transported to the laboratory, and were anaesthetized with benzocaine and sacrificed for parasite collection according to Ghiraldelli et al. 2006. Prevalence and mean intensity were calculated according to Bush et al. 1997. Ten praniza larvae, with origin in different hostfish species, (total length: 2.09 ±0.12 mm, 1.8-2.26 mm) were dissected for anatomical studies following the recommendations described elsewhere (Smit et al. 1999, Smit and Basson 2002). No attempts were made to moult the pranizae to adults.
RESULTS
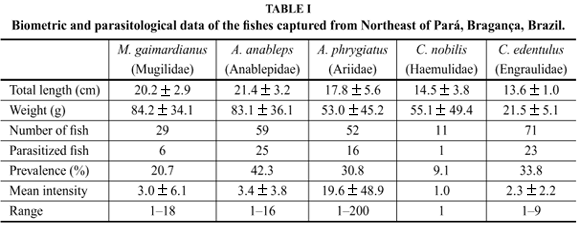
The number of fish, the total length and weight of the specimens, the prevalence and mean intensity of infection are depicted in Table I.
The parasites were found only in the gill chamber with expanded anterior hindgut filled with host blood, attached to the gill filaments with the mouthparts. The gill filaments were not pale and there were apparently not relevant modifications of the host near the place of attachment of the parasites.
The highest prevalence value was found in Anableps anableps (42.3%) and the lowest in Conodonnobilis (9.1%). The mean intensity varied from 1 parasite in C. nobilis to 19.5 in Arius phrygiatus and was lower in the other species (varying between 2.2 and 3.4), the range being very high in A. phrygiatus (1-200 parasites) and much lower in the other species.
LARVAE MORPHOLOGY
The body of the larvae (Fig. 1A) is divided into three parts: the cephalosome (including the antennae and the mouthparts), the peraeon with five pairs of peraeopods, the pleon with five pairs of pleopods, and the telsonwith one pair of uropods.

The posterior margin of the cephalosome is slightly wider than the anterior one, almost as wide as it is long, presenting few setae on the posterior dorsal region. The lateral margins are straight and parallel. The well developed compound eyes are oval-shaped and located on the lateral margins of the cephalosome, and the length of the eye is more or less half of the length of the cephalosome (Fig. 1A). The cephalosome has straight medio-anterior margins with concave lateral excavations to accommodate the first articles of the antennae.
The antennae are straight, the antenna 2 beinglonger than the antenna 1. Antenna 1 has three pedunculate articles, the third of which is the largest (Fig. 1A). The flagellum has four articles, of which article 2 is the largest. Articles 2 and 3 have one aesthetasc seta each, and article 4 ends in one aesthetasc seta and two simple setae. Article 2, 3 and 4 presented few setae. Antenna 2 has four pedunculate articles, the fourth of which is the largest. The flagellum has seven articles, of which article 1 is the largest, article 7 ends in three or four simple setae, and few setae exist on the distal end of each article (Figs. 3A and 3B).

The labrum is prominent and semicircular, with an apical process, and its posterior and anterior margins are concave. The ventral part is gutter-like with a central groove, covering the mandibles both dorsally and laterally (Fig. 1A).
The gnathopods are smaller than the peraeopods, have seven articles, the dactylus is hooked, and they present a few simple setae without scales (Fig. 1B).
The paragnaths are prolonged and end in sharppoints, presenting no teeth (Fig. 1C).
The maxillules are long and thin, exceeding thedistal margin of the labrum. They have six teeth on the distal inner margin and one tooth at the tip (Fig. 1D).
The mandibles are stout, swollen at the base, with the distal margin styliform with seven large triangular teeth on the medial margin, facing backwards, and one small tooth located at the tip of the mandible (Fig. 1E).
The maxilliped is large and cylindrical, with an elongated base. The palp has three articles, the first one being acute, with five teeth placed distally, and a single long simple setae ventrally located. Article 3 has six long simple setae (Fig. 1F).
The pereon is larger than the cephalosome and almost twice as long as it is wide. Pereonite 1 is fused with the cephalon (Fig. 1A), dorsally visible, and has shallow and convex anterior and posterior borders. Pereonite 2 has an anterior constriction separating it medially from pereonite 1. Pereonite 3 is the largest one and when the larvae are full of blood it represents about 50% of the body length. (Fig. 1A). Pereonite 4 is about twice as wide as it is long and has a rounded posterior margin stretching over pereonite 5, with lateral shields at the leg connection. Pereonite 5 consists of an elastic membrane with bulbous shields on its lateral sides at the leg connection. Pereonite 6 is rectangular and its posterior margin is slightly concave. Pereonite 7 is small and dorsally visible, with a rounded posterior margin overlapping the first pleonite (Fig. 1A).
The peraeopods show all six segments with simple setae in varying number (Fig. 4). They are divided atthe base, ischium, merus, carpus, propodus and dactylus. The base is bigger than the others, with one simple seta. The ischium is three quarters of the length of the base, with the same width. The merus is three quarters of the length of the ischium, with an anterior bulbous protrusion and long simple setae. The carpus is the same length as the merus and equipped with a small bulbous protrusion. The size of the propodus is twice as long as the carpus, with one to four simple setae distributed at the back and at the front. The dactylus is the same size as the propodus, terminating in a sharp point with no setae.
The pleotelson is triangular in shape, longer than it is wide, with straight lateral margins, two simple setae on the dorsal surface, the distal tip ending in a pair of simple setae (Fig. 2A).
The exopod and endopod on the pleopods are almost similar in size, both are fringed distally with plumose setae. The endopod has six setae and the exopod nine setae. Short, simple setae are distributed laterally along the margins (Fig. 2B).
The uropods have an endopod extending beyond the tip of the pleotelson, and exopods reaching the tip. They have eight plumose setae on the final appendage, and one plumose setae on the medial region (Fig. 2A).
REMARKS
As shown in Table I there was a considerable variation concerning the prevalence and mean intensity values between the different host species, as well as in therange of the parasites. Considering all the features it can be concluded that A. phrygiatus was the most infected species, in spite of higher values for prevalence obtained for C. edentulus and A. anableps. However, A. phrygiatus had a mean intensity of infection and range values much higher than the other species. This fact is likely to be related to the benthonic behavior of the fish since the adults and larvae of gnathiids are located on the sea-bed, these fish are probably more easily infected.
Comparing our values with literature reports concerning the infection of other species it can be seen that there is a great variation between the prevalence and intensity of infection, in some cases higher (González et al. 2004, Genc et al. 2005) and in others lower than the values obtained for our specimens (Grau et al. 1999, Yuniar et al. 2007). According to Genc et al. (2005), who studied the infection of Epinephelus aneus in the North-eastern Mediterranean, the infestation increased at higher temperatures and varied throughout the year. In our case, in spite of the relatively low number of fish observed in order to draw conclusions about seasonality, there are apparently no differences between the infection from August, 2006 to February, 2007. This may be related to a less variable water temperature around the year at our sampling site.
The pathology induced by these parasites is variable (for review see Smit and Davies 2004). While some authors did not find relevant lesions caused by the parasites others reported severe lesions or even the death of the hosts. Giannetto et al. (2003) reported the mortality of several fish species in an aquarium due to the infestation by Gnathia vorax. Paperna and Overstreet (1981) described lesions caused by G. piscivora in farmed fish in the Red Sea, and also the death of fish from anaemia and stress, and Mugridge and Stallybrass (1983), described mortality of eels attributed to gnathiidae. In other cases focal lesions on the place of attachment of the larvae were described (Heupel and Bennett 1999, González et al. 2004, Marino et al. 2004), as well as the reduction of the haematocrit of the host (Jones and Grutter 2005).
In our specimens apparently there appeared to be no lesions other than the focal ones in the place of attachment, and the condition of the hosts was apparently unaltered.
The morphology of the larvae shows that the mouthparts are well adapted and modified for blood feeding. The backward directed teeth on the mandibles, and the hooked dactylus present at the extremity of the gnathopods, are very likely to contribute to the anchoring of the larvae to the integument of the hosts.
The examination of the present material does notallow us to identify the parasite since we only captured the larvae and the current taxonomy of gnathiidsis based solely on the morphology of free-living adult males (Smit et al. 2003, Smit and Davies 2004). The general characteristics of our specimens are similar to those reported for other species (Davies 1981, Smit et al. 1999, Smit and Basson 2002, M.L. Coetzee, unpublished data). However, the comparison of our specimens with descriptions of other larvae descriptions (Table II) shows some differences, namely the relative small number of teeth on the mandible (7), and the lowest numberof teeth (1) on the tip of the mandible, and the low number of setae in the pleotelson.
It is now well established that gnathiid larvae can transmit blood parasites to the fish, namely Haemogregarina spp., and act as definitive hosts during the life cycle of the parasite (Davies 1982, Davies et al. 1994, Davies and Smit 2001). Therefore, it may be that the effect most detrimental to our host specimens was caused by blood parasites instead of blood sucking by the larvae. We do not know whether our fish hosts are infected by blood parasites and the continuation ofthis research aims to study of blood films to ascertain whether they are infected or not, as well as the examination of the gut content of praniza larvae to try to detect development al stages of blood parasites as has already been demonstrated (Davies 1982, Davies et al. 1994, Davies and Smit 2001).
Manuscript received on February 25, 2008; accepted for publication on June 14, 2008;presented by SERGIO VERJOVSKI-ALMEIDA
- BUSH AO, LAFFERTY KD, LOTZ JM AND SHOSTAK AW. 1997. Parasitology meets ecology on its terms. J Parasitol 83: 575583.
- DAVIES AJ. 1981. A scanning electron microscope study of the praniza larva of Gnathia maxillaries Montagu (Crustacea, Isopoda, Gnathiidae), with special reference to the mouthparts. J Nat Hist 15: 545554.
- DAVIES AJ. 1982. Further studies on Haemogregarina bigemina Laveran & Mesnil, the marine fish Blenius pholis L.,and the isopod Gnathia maxillaries Montagu. J Protozool 29: 5767583.
- DAVIES AJ AND SMIT NJ. 2001. The life cycle of Haemogregarina bigemina (Adeleina: Haemogregarinidae) in South African Hosts. Folia Parasitol 48: 169177.
- DAVIES AJ, EIRAS JC AND AUSTIN TE. 1994. Investigations into the transmission of Haemogregarina bigemina Laveran and Mesnil, 1901 (Apicomplexa: Adeleorina) between fishes in Portugal. J Fish Diseases 17: 283289.
- GENC E, GENC MA, CAN MF, GENC E AND CENGIZLER I.2005. A first documented record of gnathiid infestation on white grouper (Epinephelus aeneus) in Iskenderum Bay (north-eastern Mediterranean), Turkey. J Appl Ichthyol 21: 448450.
- GHIRALDELLI L, MARTINS ML, JERÓNIMO GT, YAMASHITA MM AND ADAMANTE WB. 2006. Ectoparasites communities from Oreochromis niloticus cultivated in the state of Santa Catarina, Brazil. J Fish Aquat Sci 2: 181190.
- GIANNETTO S, MARINO F, PARADISO ML, MACRI D,BOTTARI T AND DE VICO G. 2003. Light and scanning electron microscopy observations on Gnathia vorax (Isopoda: Gnathiidae) larvae. J Submicr Cytol Pathol 35: 161165.
- GONZÁLEZ P, SANCHEZ MI, CHIRIVELLA J, CARBONELL E, RIERE F AND GRAU A. 2004. A preliminary study on gill metazoan parasites of Dentex dentex (Pisces: Sparidae) from the western Mediterranean Sea (Balearic Islands). J Appl Ichthyol 20: 276281.
- GRAU A, RIERA F AND CARBONELL E. 1999. Some protozoan and metazoan parasites of the amberjack from the Balearic Sea (western Mediterranean). Aquacult Int 7: 276281.
- HADFIELD KA AND SMIT NJ. 2008. Description of a new gnathiid, Afrignathia multicavea gen. et sp.n. (Crustacea: Isopoda: Gnathiidae), from South Africa. Afr Zool 43: 8189.
- HEUPEL MR AND BENNETT MB. 1999. The occurrence, distribution and pathology associated with gnathiid isopod larvae infecting the apaulette shark Hemiscylliumocellatum Int J Parasitol 29: 321330.
- JONES CM AND GRUTTER AS. 2005. Parasitic isopods (Gnathia sp.) reduce haematocrit in captive blackeye thicklip (Labridae) on the Great Barrier Reef. J Fish Biol 66: 860864.
- MARINO F, GIANNETTO S, PARADISO ML, BOTTARI T,DE VICO G AND MACRI B. 2004. Tissue damage and haematophagia due to praniza larvae (Isopoda: Gnathiidae) in some aquarium seawater teleosts. Dis Aquat Org 59: 4347.
- MUGRIDGE ER AND STALLYBRAS HG. 1983. A mortality of eels, Anguilla Anguilla L. attributed to Gnathiidae. J Fish Diseases 6: 8182.
- PAPERNA I AND OVERSTREET RM. 1981. Parasites and diseases of mullets (Mugilidae). In: AQUACULTURE OF GREY MULLETS (Oren OH, Ed.), Cambridge Academic Press, Cambridge, UK, p. 411493.
- SANTO RVE, ISAAC VJ, SILVA LMA, DA MARTINELLIJM, HIGUCHI H AND SAINT-PAUL U. 2005. Peixes e camarões do litoral bragantino. MADAM, Belém, PA, Brasil, 268 p.
- SMIT NJ AND BASSON L. 2002. Gnathia pantherina sp.n.(Crustacea: Isopoda: Gnathiidae), a temporary parasite of some elasmobranchs species from southern Africa. Folia Parasitol 49: 137151.
- SMIT NJ AND DAVIES AJ. 2004. The curious lifestyle of the parasitic stages of gnathiid isopods. Adv Parasitol 58: 289391.
- SMIT NJ, VAN AS JG AND BASSON L. 1999. A redescription of the male and praniza of Gnathia Africana Barnard,1914 (Crustacea, Isopoda, Gnathiidae) from southern Africa. Folia Parasitol 46: 229240.
- SMIT NJ, BASSON L AND VAN AS JG. 2003. Life cycle of the temporary fish parasite, Gnathia Africana (Crustacea: Isopoda: Gnathiidae). Folia Parasitol 50: 135142.
- YUNIAR AT, PALM HW AND WALTER T. 2007. Crustacean fish parasites from Segara Anakan Lagoon, Java, Indonesia. Parasitol Res 100: 11931204.
Correspondence to:
Publication Dates
-
Publication in this collection
19 Nov 2008 -
Date of issue
Dec 2008
History
-
Received
25 Feb 2008 -
Accepted
14 June 2008