Abstracts
Heparan sulfate proteoglycans are ubiquitously found at the cell surface and extracellular matrix in all the animal species. This review will focus on the structural characteristics of the heparan sulfate proteoglycans related to protein interactions leading to cell signaling. The heparan sulfate chains due to their vast structural diversity are able to bind and interact with a wide variety of proteins, such as growth factors, chemokines, morphogens, extracellular matrix components, enzymes, among others. There is a specificity directing the interactions of heparan sulfates and target proteins, regarding both the fine structure of the polysaccharide chain as well precise protein motifs. Heparan sulfates play a role in cellular signaling either as receptor or co-receptor for different ligands, and the activation of downstream pathways is related to phosphorylation of different cytosolic proteins either directly or involving cytoskeleton interactions leading to gene regulation. The role of the heparan sulfate proteoglycans in cellular signaling and endocytic uptake pathways is also discussed.
glycosaminoglycans and protein interactions; growth factors; focal adhesion; extracellular matrix; cell cycle; cell proliferation
Proteoglicanos de heparam sulfato são encontrados tanto superfície celular quanto na matriz extracelular em todas as espécies animais. Esta revisão tem enfoque nas características estruturais dos proteoglicanos de heparam sulfato e nas interações destes proteoglicanos com proteínas que levam à sinalização celular. As cadeias de heparam sulfato, devido a sua variedade estrutural, são capazes de se ligar e interagir com ampla gama de proteínas, como fatores de crescimento, quimiocinas, morfógenos, componentes da matriz extracelular, enzimas, entreoutros. Existe uma especificidade estrutural que direciona as interações dos heparam sulfatos e proteínas alvo. Esta especificidade está relacionada com a estrutura da cadeia do polissacarídeo e os motivos conservados da cadeia polipeptídica das proteínas envolvidas nesta interação. Os heparam sulfatos possuem papel na sinalização celular como receptores ou coreceptores para diferentes ligantes. Esta ligação dispara vias de sinalização celular levam à fosforilação de diversas proteínas citosólicas ou com ou sem interações diretas com o citoesqueleto, culminando na regulação gênica. O papel dos proteoglicanos de heparam sulfato na sinalização celular e vias de captação endocítica também são discutidas nesta revisão.
glicosaminoglicanos e interações com proteínas; fatores de crescimento; adesão focal; matriz extracelular; ciclo celular; proliferação celular
BIOMEDICAL AND MEDICAL SCIENCES
Heparan sulfate proteoglycans: structure, protein interactions and cell signaling
Juliana L. DreyfussI; Caio V. RegatieriI,II; Thais R. JarrougeI; Renan P. CavalheiroI; Lucia O. SampaioI; Helena B. NaderI
IDisciplina de Biologia Molecular, Departamento de Bioquímica, Universidade Federal de São Paulo Rua Três de Maio, 100, 04044-020 São Paulo, SP, Brasil
IIDepartamento de Oftalmologia, Universidade Federal de São Paulo, Rua Botucatu, 822, 04023-062 São Paulo, SP, Brasil
Correspondence to Correspondence to: Dr. Helena B. Nader E-mail: hbnader.bioq@epm.br
ABSTRACT
Heparan sulfate proteoglycans are ubiquitously found at the cell surface and extracellular matrix in all the animal species. This review will focus on the structural characteristics of the heparan sulfate proteoglycans related to protein interactions leading to cell signaling. The heparan sulfate chains due to their vast structural diversity are able to bind and interact with a wide variety of proteins, such as growth factors, chemokines, morphogens, extracellular matrix components, enzymes, among others. There is a specificity directing the interactions of heparan sulfates and target proteins, regarding both the fine structure of the polysaccharide chain as well precise protein motifs. Heparan sulfates play a role in cellular signaling either as receptor or co-receptor for different ligands, and the activation of downstream pathways is related to phosphorylation of different cytosolic proteins either directly or involving cytoskeleton interactions leading to gene regulation. The role of the heparan sulfate proteoglycans in cellular signaling and endocytic uptake pathways is also discussed.
Key words: glycosaminoglycans and protein interactions, growth factors, focal adhesion, extracellular matrix, cell cycle, cell proliferation.
RESUMO
Proteoglicanos de heparam sulfato são encontrados tanto superfície celular quanto na matriz extracelular em todas as espécies animais. Esta revisão tem enfoque nas características estruturais dos proteoglicanos de heparam sulfato e nas interações destes proteoglicanos com proteínas que levam à sinalização celular. As cadeias de heparam sulfato, devido a sua variedade estrutural, são capazes de se ligar e interagir com ampla gama de proteínas, como fatores de crescimento, quimiocinas, morfógenos, componentes da matriz extracelular, enzimas, entreoutros. Existe uma especificidade estrutural que direciona as interações dos heparam sulfatos e proteínas alvo. Esta especificidade está relacionada com a estrutura da cadeia do polissacarídeo e os motivos conservados da cadeia polipeptídica das proteínas envolvidas nesta interação. Os heparam sulfatos possuem papel na sinalização celular como receptores ou coreceptores para diferentes ligantes. Esta ligação dispara vias de sinalização celular levam à fosforilação de diversas proteínas citosólicas ou com ou sem interações diretas com o citoesqueleto, culminando na regulação gênica. O papel dos proteoglicanos de heparam sulfato na sinalização celular e vias de captação endocítica também são discutidas nesta revisão.
Palavras-chave: glicosaminoglicanos e interações com proteínas, fatores de crescimento, adesão focal, matriz extracelular, ciclo celular, proliferação celular.
STRUCTURAL FEATURES OF HEPARAN SULFATE
The most distinguishing features between heparin and heparan sulfate (HS) are their cellular localization, their occurrence in the animal kingdom, and thus their biological functions. Heparin and heparan sulfate are attached to different core proteins and found in different cellular compartments. Heparin is found exclusively inside storage vesicles of mast cells of some animal species (Nader et al. 1999a, 1980, Straus et al. 1982) whereas heparan sulfates are ubiquitous to the cell surface of both vertebrate and invertebrate species (Cassaro and Dietrich 1977, Dietrich et al. 1980, 1977, Nader et al. 1984).
Heparin and heparan sulfates are polydisperse linear polymers that share structural similarities. They are composed of alternate units of α-D-glucosamine (GlcN) and uronic acid, either β-D-glucuronic acid (GlcA) or α-L-iduronic acid (IdoA), joined together by (1  4) glycosidic linkages. In heparan sulfate the GlcN can be either N-sulfated or N-acetylated, whereas in heparin the N-acetyl groups correspond to less then 5%. Furthermore, heparin shows higher degree of sulfation (2.3-2.8 sulfates/disaccharide) when compared to heparan sulfates (0.6-1.5 sulfates/disaccharide).
4) glycosidic linkages. In heparan sulfate the GlcN can be either N-sulfated or N-acetylated, whereas in heparin the N-acetyl groups correspond to less then 5%. Furthermore, heparin shows higher degree of sulfation (2.3-2.8 sulfates/disaccharide) when compared to heparan sulfates (0.6-1.5 sulfates/disaccharide).
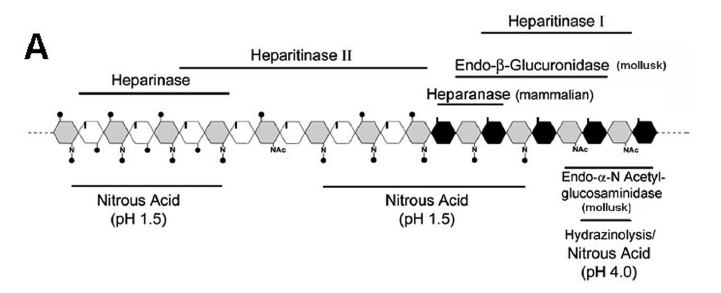
The sequences of the different types of disaccharides in heparan sulfate and heparin were established using chemical, enzymatic and NMR analyses of the intact polymers and their fragments. Figure 1A summarizes the sites of action of the chemical and enzymatic protocols.
Chemical procedures, such as nitrous acid degradation of the polymer at different pHs, can furnish important data on the structure of these compounds. At low pH and room temperature, the N-sulfated GlcNs in the heparan sulfate are susceptible to degradation yielding fragments with ranges of molecular weights that depend on the distributions of the N-sulfated GlcN residues in the chain and bearing an anydromannose at the reducing terminal end of the fragment. Thus, the obtained fragments will contain clusters of N-acetylated GlcN, since N-acetylated amino sugars are not affected (Conrad 2001). On the other hand, hydrazinolysis coupled with nitrous acid treatment at pH 4.0 affects the N-acetylated portion of the polymer.
Bacterial glycosaminoglycan lyases and the animal endo-hydrolases described so far have also been used to ascertain the disaccharide sequences in heparan sulfate chains. Mollusk endo-β-glucuronidase and α-D-Nacetylglucosaminidase degrade heparan sulfates chains yielding oligosaccharides enriched in O-sulfates and IdoA residues. Furthermore, heparan sulfate can be degraded by a class of mammalian endo-hydrolases known as heparanases, which are endo-β-glucuronidases that cleave β-D-glucuronyl (1 4) D-GlcN N-sulfated located after a disaccharide composed of α-L-iduronyl (1 4) D-GlcN N-acetylated. Figure 1A summarizes the sites of action of the chemical and enzymatic procedures.
Characteristic 1H and 13C chemical shifts have been identified for the individual residues, and the relative abundance of these moieties can be quantitatively determined by integrating the proton signals. By a combination of two-dimensional NMR techniques such as correlated spectroscopy (COSY), nuclear overhauser effect (NOESY) and total correlation spectroscopy (TOCSY) for 1H, and heteronuclear single-quantum coherence (HSQC) for 13C, some of the sequences can be determined (Chavante et al. 2000, Chuang et al. 2001, Dietrich et al. 1999, Ferreira et al. 1993, Guerrini et al. 2001, 2002, Nader et al. 1999b, 1990).
The combined used of these approaches made it possible to establish the sequence of characteristic domains in the structure of heparan sulfates from different origins (Fig. 1B). Thus heparan sulfates from both vertebrate and invertebrate tissues contain common structural features such as N-acetylated and N-sulfated GlcN, domains consisting only of GlcA-containing disaccharides with no 6-O-sulfate substitutions (susceptible to heparitinase I, endo-β-glucuronidases, and nitrous acid pH 4.0) and a more sulfated region consisting of IdoAcontaining disaccharides (susceptible to heparitinase II and nitrous acid pH 1.5). At the non-reducing end all polymers contain GlcN N-sulfate or GlcN N,6-disulfate followed by a disaccharide composed of IdoA 2-O-sulfated linked to GlcN 2,6-disulfated (susceptible to heparinase). A peculiar tetrasaccharide is positioned between the two regions and was identified in all heparan sulfates. The N-acetylated GlcA domain is close to the protein core and contains the reducing terminal of the chain (Dietrich et al. 1983, 1998, Ferreira et al. 1993, Nader et al. 1987, 1999b, Tersariol et al. 1994).
Heparan sulfates are absent in protista, plantae and fungi, and their appearance in the animal kingdom coincides with the emergence of eumetazoa, which are animals that display true tissues, being ubiquitously found in all tissues and species analyzed (Cassaro and Dietrich 1977, Dietrich et al. 1980, 1977, Gomes and Dietrich 1982, Nader et al. 1984, Toledo and Dietrich 1977). Non-sulfated version of heparan sulfate, named heparosan, is found in the capsules of some pathogenic bacteria, thus acting as molecular camouflages protecting the microbe and enhancing infection (DeAngelis 2002).
Heparan sulfates are attached to different core proteins and found at the cellular surface and extracellular matrices, such as basal membrane. The chains at the cell surface can be attached to transmembrane proteins as in syndecans or through a glycosylphosphatidylinositolanchored core protein, as in glypicans (Bernfield et al. 1999, Fransson 2003, Fransson et al. 2004, Tantravahi et al. 1986, Tkachenko et al. 2005). Table I shows different proteins that can bear heparan sulfate chains and their cellular localization.
HEPARAN SULFATES AND PROTEIN INTERACTIONS
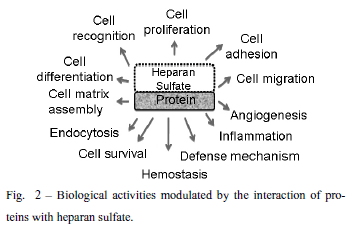
Several works in the literature clearly show that there is a specificity directing the interactions of heparan sulfates and target proteins, regarding both the fine structure of the polysaccharide chain and precise protein motifs. Thus, they can interact with a diverse range of proteins leading to biological activities (Fig. 2). The heparan sulfate chains due to their vast structural diversity are able to bind and interact with a wide variety of proteins, such as growth factors, chemokines, morphogens, extracellular matrix components, and enzymes, among others. Table II lists some of the heparan sulfate binding proteins that modulate different biological processes through this interaction.
These proteins contain relatively large numbers of the basic amino acids (lysine, arginine and in some cases histidine). These basic residues can be found in linear arrangements or in spatial folded clusters. Cardin and Weintraub proposed two consensus motifs, XBBXBX or XBBBXXBX, where B represents basic amino acids and X, hydropathic (neutral or hydrophobic) residue (Cardin andWeintraub 1989). Nevertheless, binding can also involve basic amino acids that are distant in linear sequence of the protein and that are brought together in the protein folded state (Capila and Linhardt 2002, Hileman et al. 1998, Krilleke et al. 2007, Mulloy and Linhardt 2001, Vives et al. 2004).
Considering the heparan sulfates, the specificity seems to be related to the distribution and conformation of β-D-GlcA and α-L-IdoA residues, relative amounts of N-acetyl or N-sulfate groups in the GlcN moiety, as well as the relative amounts and the position of O-sulfation of the uronic acid and GlcN units. Specific sequences of disaccharides can favor the interaction of the molecule with certain proteins and not to others. Up to now, besides specific sugar sequences bearing IdoA and enriched in sulfate groups (S-domain), it has been postulated that the conformational flexibility of the α-L-IdoA residue plays a pivotal role in protein interactions. IdoA residues can assume both 1C4 chair and the 2S0 skew boat conformation, thus allowing appropriate electrostatic interactions with basic amino acids on the protein (Casu et al. 1986, Ferro et al. 1990, Gallagher 2006, Habuchi et al. 2004, Mulloy 2005, Mulloy and Forster 2000, Noti and Seeberger 2005, Ragazzi et al. 1993). It has been shown that a heparin-derived tetrasaccharide that interacts with annexin V shows IdoA on the 2S0 conformation, while the non-interacting tetrasaccharide the 1C4 conformation (Capila et al. 2001, 1999, Ishitsuka et al. 1998). More recently, it has been suggested the N-acetylated region (NA-domain), which is rich in β-D-GlcA residues, also displays structural plasticity and hence could mediate protein interactions (Mobli et al. 2008).
Using heparin derived oligosaccharides and chemically modified molecules, the role of N-sulfated and Nacetylated domains, as well as the position of the O-sulfates, and the conformation of the uronic acid residue were investigated regarding the binding specificity to different proteins.
It is well established that fibroblast growth factors and their receptors are dependent on binding to heparan sulfate, and this interaction is an absolute requirement for full signaling. However most of the studies regarding the minimum structural features of the polysaccharide needed for the binding were concluded using chemico-enzymatically heparin derivatives as heparan sulfate analogs. Even though these heparin derivatives can be produced in high amounts, and thus can be used to elucidate some of the binding characteristics, they do not substitute the high diversity and thus the specificity that is found in the heparan sulfate polymers (Belford et al. 1992, Harmer 2006, Ishihara et al. 1993, Mohammadi et al. 2005a, Presta et al. 2005, Yates et al. 2004).
However, heparan sulfates show large sequences of GlcA linked to N-acetylated GlcN which are not present in heparins. Also, heparan sulfates show lower degree of sulfation, even in the IdoA residue, requisites that are described as important for the protein binding. So, this raises questions on how the binding could be affected by these domains in the heparan sulfate chains, which are the postulated polysaccharide for most of these biological interactions.
The structural requirements involved in the binding vary for each protein. Some important sequences for specific heparin/heparan sulfate-protein interactions, which are dependent mostly on the presence of IdoA, as well as the sulfation of the IdoA and the N-sulfation of the GlcN moiety, have been established (Jastrebova et al. 2006, Patel et al. 2008, Sampaio et al. 2006, Sasisekharan et al. 2002, Yates et al. 2004, Zhang et al. 2007). Furthermore, the protein interaction depends on the size of the chain, and the minimum fragment varies from a tetrasaccharide described for annexin V up to a dodecasaccharide for the FGF-2 receptor.
The sulfation pattern is another important requisite for the binding of heparin/heparan sulfate to proteins. For example, the growth factor PDGF-A is dependent mostly on the amounts of 2-O-sulfate in the IdoA residues (Feyzi et al. 1997). On the other hand, for the chemokine CCL-2, both 2-O-sulfate in the IdoA and Nsulfation of the GlcN are required (Crown et al. 2006). The interaction of heparan sulfate with FGF-4 receptor depends more on the number of 6-O-sulfate groups than on their precise location (Loo et al. 2001), and for FGF-receptor 2 the minimum structure for binding is an octasaccharide containing 2-O- and 6-O-sulfates and for signaling a dodecasaccharide (Walker et al. 1994). In a recent paper, using embryonic fibroblasts derived from knock-out mice for heparan sulfate 6-O-sulfotransferases 1- and 2, it was shown an important role of 6-Osulfation patterns in FGF signaling (Sugaya et al. 2008). The binding of neuregulin-1 to erbB receptor depends mainly on the N-sulfate groups of heparan sulfate, followed by 2-O- and 6-O-sulfate groups (Pankonin et al. 2005). Interaction of endostatin to endothelial heparan sulfate shows differential requirements for specific sulfate groups where 6-O-sulfates play a dominant role in selectivity (Blackhall et al. 2003). Recently, it has been shown that 6-O-sulfation of heparan sulfate differentially regulates various fibroblast growth factor-dependent signalings in culture (Sugaya et al. 2008).
Specific structural features of heparan sulfate involved in protein interactions were illustrated by experiments using FGF-1 and heparan sulfates from various sources that exhibit different disaccharides assembling. The FGF-1 mitogenic activity varies among the different heparan sulfates. The oligosaccharide derived from a heparan sulfate containing only the GlcA domain with no 6-O-sulfation (NA/NS-domain) displays no activity, whereas the counterpart enriched in IdoA and 6-O-sulfation (S-domain) shows around 10 times the activity of the intact polymer (Fig. 1B). Furthermore, heparan sulfate purified from 3T3 fibroblasts has an effect about 100 times higher. These results indicate that endogenous heparan sulfate is the best elicitor for the FGF-1 mitogenic activity, and that the S-domain represents the FGF-1 binding site, indicating a highly specific interaction (Gambarini et al. 1993).
Thus, it appears that the specificity of the interactions heparan sulfate-protein depends on the overall organization of the glycosaminoglycan chain rather than on the fine structure of the individual sequences to achieve its functional role (Gambarini et al. 1993, Kreuger et al. 2006, Sampaio et al. 2006, Suarez et al. 2007).
HEPARAN SULFATE PROTEOGLYCANS AND CELL SIGNALING PATHWAYS
Heparan sulfate chains are located facing the extracellular compartment, and thus their biological roles can be related to assemble the extracellular matrices (Baeg et al. 2001, Iozzo 2005, Peretti et al. 2008), to modulate the activity of enzymes and/or their inhibitors (Almeida et al. 2001, Hausser et al. 2004, Nascimento et al. 2005, Raman et al. 2005, Sasaki et al. 1999, Whitelock et al. 1996, Yu and Woessner 2000, Yu et al. 2000), to provide an extracellular gradient of growth factors and chemokines (Ashikari-Hada et al. 2005, Grunert et al. 2008, Hacker et al. 2005, Kirkpatrick and Selleck 2007, Ng et al. 2006, Nugent and Iozzo 2000, Ruhrberg et al. 2002), and to prevent degradation of growth factors (Saksela et al. 1988), among others. Nevertheless, heparan sulfate proteoglycans can trigger cell response through signal transduction pathways, as well as by translocation to intracellular compartments, due to interactions of the polysaccharide chains and/or the core protein with specific ligands.
Heparan sulfates play a role in cellular signaling either as receptor or co-receptor for different ligands (Fig. 3). The activation of downstream pathways is related to phosphorylation of different cytosolic proteins either directly (Fig. 3A) or involving cytoskeleton interactions (Fig. 3B) leading to gene regulation.
In early 1990's Yayon and co-workers, using CHO cells defected in glycosaminoglycan biosynthesis, suggested heparan sulfates as low affinity receptors required for binding of FGF-2 to the high affinity site (Yayon et al. 1991). Such an obligatory interaction of low and high affinity FGF receptors suggested a novel mechanism for the regulation of growth factor-receptor interactions. Indeed, this effect of heparan sulfate is responsible for FGF receptor dimerization and activation, leading to cellular responses (Rapraeger et al. 1991, Spivak- Kroizman et al. 1994).
As described above, HSPGs can function as lowaffinity receptors required for the activation of growth factor high-affinity receptor, which has tyrosine kinase activity. A substantial body of literature supports the concept of the ternary complex involving HSPG, growth factor and its high-affinity receptor (Fig. 3A-2).
Heparan sulfates as co-receptors for fibroblast growth factors with tyrosine kinase activity have been extensively studied and corroborated in other systems (Czubayko et al. 1997, Duchesne et al. 2006, Mohammadi et al. 2005a, b, Mongiat et al. 2000, Padera et al. 1999, Pellegrini et al. 2000, Powers et al. 2000, Rapraeger et al. 1994, Wiedlocha and Sorensen 2004, Wu et al. 1991). This general model has been also extended to many other growth factors, such as vascular endothelial growth factor (Ashikari-Hada et al. 2005, Gitay-Goren et al. 1992, Iozzo and San Antonio 2001, Stringer 2006), hepatocyte growth factor (Kemp et al. 2006, Rubin et al. 2001, Schwall et al. 1996), plateletderived growth factor (Abramsson et al. 2007, Rolny et al. 2002), placenta growth factor (Athanassiades and Lala 1998), and heparin binding-epidermal growth factor (Aviezer and Yayon 1994).
Heparan sulfate proteoglycans can also interfere with serine/threonine kinase receptors, such as transforming growth factor-β and bone morphogenetic protein (Chen et al. 2006, Cohen 2003, Grunert et al. 2008, Rider 2006, Sasaki et al. 2008), tyrosine phosphatase receptors (Aricescu et al. 2002, Fox and Zinn 2005, Johnson et al. 2006), 7-helix transmembrane receptors coupled to G-protein (Lau et al. 2004, Lortat-Jacob et al. 2002, Parish 2006) and other multiple-helices transmembrane receptors (Hacker et al. 2005, Sasaki et al. 2008).
Different protein cores of the heparan sulfate proteoglycans have been described regarding growth factor and chemokine signaling transduction. Among them, the syndecan family is the most extensively studied (Alexopoulou et al. 2007, Bartlett et al. 2007, Beauvais and Rapraeger 2004, Fears and Woods 2006, Lopes et al. 2006a, Porcionatto et al. 1999, Su et al. 2007, Tkachenko et al. 2005). Nevertheless, other cell surface proteoglycans, such as glypicans (Cano-Gauci et al. 1999, Capurro et al. 2008, Filmus et al. 2008, Gumienny et al. 2007, Kayed et al. 2006, Song et al. 2005, Traister et al. 2008) and betaglycans (Harrison et al. 2005, Lewis et al. 2000), have been also implicated with cellular transduction mechanisms.
Another possibility is that the proteoglycan itself could act as a transducer for cell signaling elicited by a growth factor. Using as a working model, with L6 myoblast cells lacking endogenous functional high-affinity FGF receptors, it was proposed the direct involvement of syndecan with the internalization of FGF-2 and cellular response. This cell signaling is distinct from the better known transmembrane tyrosine kinase receptors (Quarto and Amalric 1994).
Mechanisms leading to gene regulation can also involve interaction of the extracellular cell matrix (ECM) components with cytoskeleton via transmembrane surface receptors, such as integrins and/or heparan sulfate proteoglycans (Fig. 3B). Integrins consist of heterodimers of single helix transmembrane proteins that, like syndecans, do not display enzymatic activity and so their actions as transducers depend on the activation of a number of cytoplasmic kinases. The best evidence for a specific role of integrins in cell adhesion and cell migration comes from studies of focal adhesion formation. Fibronectin, vitronectin, collagen, laminin, among others, including matrix proteoglycans, such as perlecan, collagen XVIII and agrin, are potential ECM ligands of integrins. On the other hand, the intracellular domain interacts with many cytoplasmic proteins including talin, vinculin, paxillin and α-actinin. These set of molecules can activate kinases like FAK (focal adhesion kinase) and Src, which in turn leads to a cascade of protein phosphorylation that regulates genes expression involved in cell spreading, recognition, adhesion, growth control, apoptosis, etc. (Bernfield et al. 1999, Lopes et al. 2006a). The integrins thus link across the plasma membrane two networks: the extracellular and the intracellular actin filamentous system.
The connection of ECM and cytoskeleton can also be mediated by syndecans either directly or as co-receptor of integrins. The cytoplasmic region of syndecans contains two domains that are conserved in each of the four syndecans, and flank a central variable region that is distinct for each family member. The invariant region close to the transmembrane domain contains serine and tyrosine which are potential substrates for phosphorylation, as well as specific motifs that can bind to kinases as Src and Fyn. The variable region is distinct for each of the 4 family members. The function of this variable domain is largely unknown except for syndecan- 4, where it has been shown to bind PIP2 (4,5-bisphosphate phosphatidylinositol) and activate PKC-a, leading to oligomerization of the proteoglycan in focal adhesions. Several studies have shown that the oligomeric status of the cytoplasmic domain is related to the activation of the downstream signaling pathway. The other conserved domain, at the C-terminal of the protein, interacts with specific proteins containing PDZ domains, such as CASK and syntenin which are thought to link membrane components to the underlying actincontaining cytoskeleton. Interestingly, the variable and first conserved domains of syndecan-4 can also bind other proteins related to the cytoskeleton, such as syndesmos and α-actinin (Oh and Couchman 2004, Woods and Couchman 2001).
A direct role of syndecan-4 in focal adhesion regulation was observed using fibroblasts derived from syndecan- 4 or fibronectin null mice. It was shown that the proteoglycan was capable to regulate FAK phosphorylation in a Rho dependent mechanism with no activation of PKC (Wilcox-Adelman et al. 2002). In a recent paper, a novel RGD-independent cell adhesion mechanism is proposed, in which syndecan-4 activates PKC-α and its subsequent interaction with the β1-integrin chain and, thus, initiating the FAK signaling cascade and actin-stress fiber organization (Telci et al. 2008). Recent results identify syndecan-4 as a novel receptor for the N-terminus of TSP-1 (thrombospondin) interfering with cell adhesion through activation of FAK (Nunes et al. 2008).
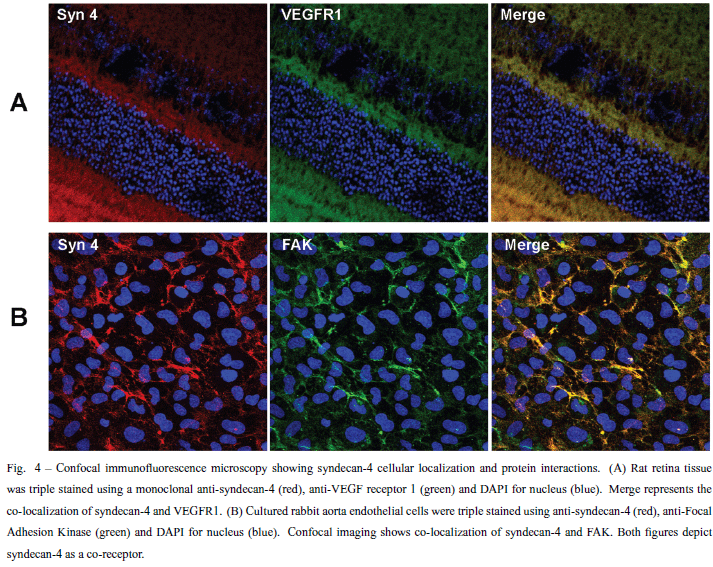
Figure 4 illustrates an experiment using confocal immunofluorescence microscopy, showing the co-localization of syndecan-4 and VEGF-receptor as well as that of syndecan-4 and FAK.
Synergistic control of cell adhesion involving integrins and syndecans were recently reviewed (Alexopoulou et al. 2007, Morgan et al. 2007).
It should also be referred that α5β1 integrin is a part time proteoglycan, and the GAG chains play an essential role in the control of motility of cells on fibronectin and, thus, in the cascade of signaling events (Franco et al. 2001, Veiga et al. 1997).
Furthermore, depending on the biological process investigated as cell migration, adhesion, growth, differentiation and apoptosis, it has been found that the extracellular matrix heparan sulfate proteoglycans, such as perlecan (Baker et al. 2008, Farach-Carson et al. 2008, Farach-Carson and Carson 2007, Giros et al. 2007, Jiang and Couchman 2003, Knox and Whitelock 2006, Lindner et al. 2007, Smirnov et al. 2005), agrin (Fox and Zinn 2005, Glass et al. 1996, Jury et al. 2007, Ngo et al. 2007, Tourovskaia et al. 2008,Williams et al. 2008), collagen XVIII (Fjeldstad and Kolset 2005) and testican (Schnepp et al. 2005) can also modulate the activity of growth factors, cytokines, morphogens and enzymes.
The different cell ligands and receptors trigger downstream pathways that share cytosolic components, leading ultimately to the activation of a complex biomolecular network. This large network of molecular interactions and signaling pathways involve phosphorylation of key substrates including enzymes, microtubules, histones, and transcription factors that play pivotal roles in determining the cellular response.
The signaling systems evoked by the interaction of heparan sulfate proteoglycans with extracellular ligands and/or receptors include pathways such as Ras/Raf/MAPK (Leicht et al. 2007), PIP3/Akt (Carnero et al. 2008), PLC/PKC (Escriba et al. 2007), cAMP/PKA (Murray 2008, Wojtal et al. 2008), among others. They are of great interest and play a key role in normal cell behavior and in diseases such as cancer, arthritis and rheumatism.
PMA (phorbol 12-myristate 13-acetate) is recognized as a strong and specific activator of PKC mimicking diacylglycerol. PMA specifically stimulates the synthesis of syndecan-4 in endothelial cells in a mechanism mediated by PKC activation. The most remarkable aspect of these results, however, was the correlation between the up-regulation of heparan sulfate proteoglycans expression and the blockade of G1-S phase transition triggered by PMA (Moreira et al. 2004, Porcionatto et al. 1998, 1994).
Recently, it has been shown that over-expression of the EJ-ras oncogene in endothelial cells modifies the cell cycle, up-regulates the expression of syndecan-4, and down-regulates several enzymes involved in heparan sulfate biosynthesis, leading to a decrease in the N- and O-sulfation of the chains (Lopes et al. 2006b). These results are in accordance to the structural characteristics of heparan sulfate from neoplastic tissues (Jeronimo et al. 1994, Oba-Shinjo et al. 2006).
The understanding of how cells control proliferation and differentiation, survival and death, migration and adhesion, requires the analyses of the crosstalk of the various pathways involved in these processes.
Although growth factor receptors are generally thought to carry out their role in signal transduction at the cell surface, many of these transmembrane proteins translocate to the nucleus after ligand stimulation.
In the 80's it was reported a nuclear pool of free heparan sulfate chains using radioactive sulfate labeling of a hepatoma cell line (Fedarko and Conrad 1986, Ishihara et al. 1986). Independently, the presence of FGF-2 in the nucleus was also documented using endothelial (Bouche et al. 1987) as well as CHO cells (Caizergues-Ferrer et al. 1984) in G0-G1 transition. On the other hand, the connection in the internalization of both heparan sulfate and FGF-2 was proposed using L6 myoblasts (Quarto and Amalric 1994).
Interestingly, the up-regulation in the expression of heparan sulfate proteoglycan induced by growth factors and PMA in endothelial cells occurs during the G0-G1 transition and has also been described associated with PKC pathway (Porcionatto et al. 1998, 1994). These results have been confirmed using corneal stromal fibroblasts (Hsia et al. 2003).
Lipid rafts seem to play an important role for FGF-2 and heparan sulfate proteoglycan internalization. FGF-2 induces syndecan-4 clustering of the proteoglycan, leading to the internalization by macropinocytosis of both molecules. It requires lipid rafts integrity, occurs in a nonclathrin-, non-dynamin-dependent manner and involves Rac1, which is activated by syndecan-4 clustering (Tkachenko et al. 2004).
The importance of glypican endocytosis as a positive or negative modulator is pointed out in the regulation of Hedgehog (Hh) signaling and in Wingless gradient formation (Beckett et al. 2008, Gagliardi et al. 2008).
Heparan sulfate proteoglycans have also been described in the internalization of ligands other than growth factors (Poon and Gariepy 2007). Syndecans and perlecan have been shown to mediate the clathrin-independent endocytosis of lipoproteins (Fuki et al. 2000, 1997). Also, a physiological role for glypican-1 in the cellular homoeostasis of polyamines was demonstrated in vesicle caveolae-mediated endocytosis (Belting 2003, Cheng et al. 2002).
Endocytic pathway for many cationic ligands mediated by cell surface proteoglycans involving raft-dependent macropinocytosis have been studied and proposed as a delivery of therapeutic genes and drugs to intracellular compartments (Fan et al. 2007, Nascimento et al. 2007).
ACKNOWLEDGMENTS
This work was supported by grants from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). J.L. Dreyfuss is sponsored by Jairo Ramos postdoctoral fellowship awarded by FAP/UNIFESP (Fundação de Apoio à Pesquisa/ Universidade Federal de São Paulo).
Manuscript received on August 26, 2008; accepted for publication on October 8, 2008; contributed by HELENA B. NADER* * Member Academia Brasileira de Ciências
In commemoration of the 75th anniversary of Escola Paulista de Medicina / Universidade Federal de São Paulo. In honor of Prof. Carl P. Dietrich
- ABRAMSSON A ET AL. 2007. Defective N-sulfation of heparan sulfate proteoglycans limits PDGF-BB binding and pericyte recruitment in vascular development. Genes Dev 21: 316-331.
- ALEXOPOULOU AN, MULTHAUPT HA AND COUCHMAN JR. 2007. Syndecans in wound healing, inflammation and vascular biology. Int J Biochem Cell Biol 39: 505-528.
- ALLIEL PM, PERIN JP, JOLLES P AND BONNET FJ. 1993. Testican, a multidomain testicular proteoglycan resembling modulators of cell social behaviour. Eur J Biochem 214: 347-350.
- ALMEIDA PC, NANTES IL, CHAGAS JR, RIZZI CC, FALJONI- ALARIO A, CARMONA E, JULIANO L, NADER HB AND TERSARIOL IL. 2001. Cathepsin B activity regulation. Heparin-like glycosaminogylcans protect human cathepsin B from alkaline pH-induced inactivation. J Biol Chem 276: 944-951.
- ARICESCU AR, MCKINNELL IW, HALFTER W AND STOKER AW. 2002. Heparan sulfate proteoglycans are ligands for receptor protein tyrosine phosphatase sigma. Mol Cell Biol 22: 1881-1892.
- ASHIKARI-HADA S, HABUCHI H, KARIYA Y AND KIMATA K. 2005. Heparin regulates vascular endothelial growth factor165-dependent mitogenic activity, tube formation, and its receptor phosphorylation of human endothelial cells. Comparison of the effects of heparin and modified heparins. J Biol Chem 280: 31508-31515.
- ATHANASSIADES A AND LALA PK. 1998. Role of placenta growth factor (PIGF) in human extravillous trophoblast proliferation, migration and invasiveness. Placenta 19: 465-473.
- AVIEZER D AND YAYON A. 1994. Heparin-dependent binding and autophosphorylation of epidermal growth factor (EGF) receptor by heparin-binding EGF-like growth factor but not by EGF. Proc Natl Acad Sci USA 91: 12173-12177.
- BAEG GH, LIN X, KHARE N, BAUMGARTNER S AND PERRIMON N. 2001. Heparan sulfate proteoglycans are critical for the organization of the extracellular distribution of Wingless. Development 128: 87-94.
- BAKER AB, ETTENSON DS, JONAS M, NUGENT MA, IOZZO RV AND EDELMAN ER. 2008. Endothelial cells provide feedback control for vascular remodeling through a mechanosensitive autocrine TGF-beta signaling pathway. Circ Res 103: 289-297.
- BARTLETT AH, HAYASHIDA K AND PARK PW. 2007. Molecular and cellular mechanisms of syndecans in tissue injury and inflammation. Mol Cells 24: 153-166.
- BEAUVAIS DM AND RAPRAEGER AC. 2004. Syndecans in tumor cell adhesion and signaling. Reprod Biol Endocrinol 2: 3.
- BECKETT K, FRANCH-MARRO X AND VINCENT JP. 2008. Glypican-mediated endocytosis of Hedgehog has opposite effects in flies and mice. Trends Cell Biol 18: 360-363.
- BELFORD DA, HENDRY IA AND PARISH CR. 1992. Ability of different chemically modified heparins to potentiate the biological activity of heparin-binding growth factor 1: lack of correlation with growth factor binding. Biochemistry 31: 6498-6503.
- BELLIN R, CAPILA I, LINCECUM J, PARK PW, REIZES O AND BERNFIELD MR. 2002. Unlocking the secrets of syndecans: transgenic organisms as a potential key. Glycoconj J 19: 295-304.
- BELTING M. 2003. Heparan sulfate proteoglycan as a plasma membrane carrier. Trends Biochem Sci 28: 145-151.
- BEN-ZAKEN O, TZABAN S, TAL Y, HORONCHIK L, ESKO JD, VLODAVSKY I AND TARABOULOS A. 2003. Cellular heparan sulfate participates in the metabolism of prions. J Biol Chem 278: 40041-40049.
- BERNFIELD M, GOTTE M, PARK PW, REIZES O, FITZGERALD ML, LINCECUM J AND ZAKO M. 1999. Functions of cell surface heparan sulfate proteoglycans. Annu Rev Biochem 68: 729-777.
- BEZAKOVA G AND RUEGG MA. 2003. New insights into the roles of agrin. Nat Rev Mol Cell Biol 4: 295-308.
- BLACKHALL FH, MERRY CL, LYON M, JAYSON GC, FOLKMAN J, JAVAHERIAN K AND GALLAGHER JT. 2003. Binding of endostatin to endothelial heparan sulphate shows a differential requirement for specific sulphates. Biochem J 375: 131-139.
- BOUCHE G, GAS N, PRATS H, BALDIN V, TAUBER JP, TEISSIE J AND AMALRIC F. 1987. Basic fibroblast growth factor enters the nucleolus and stimulates the transcription of ribosomal genes in ABAE cells undergoing G0-G1 transition. Proc Natl Acad Sci USA 84: 6770-6774.
- CAIZERGUES-FERRER M, DOUSSEAU F, GAS N, BOUCHE G, STEVENS B AND AMALRIC F. 1984. Induction of new proteins in the nuclear matrix of CHO cells by a heat shock: detection of a specific set in the nucleolar matrix. Biochem Biophys Res Commun 118: 444-450.
- CAMPBELL EJ AND OWEN CA. 2007. The sulfate groups of chondroitin sulfate- and heparan sulfate-containing proteoglycans in neutrophil plasma membranes are novel binding sites for human leukocyte elastase and cathepsin G. J Biol Chem 282: 14645-14654.
- CANO-GAUCI DF ET AL. 1999. Glypican-3-deficient mice exhibit developmental overgrowth and some of the abnormalities typical of Simpson-Golabi-Behmel syndrome. J Cell Biol 146: 255-264.
- CAPILA I AND LINHARDT RJ. 2002. Heparin-protein interactions. Angew Chem Int Ed Engl 41: 391-412.
- CAPILA I, VANDERNOOT VA, MEALY TR, SEATON BA AND LINHARDT RJ. 1999. Interaction of heparin with annexin V. FEBS Lett 446: 327-330.
- CAPILA I, HERNAIZ MJ, MO YD, MEALY TR, CAMPOS B, DEDMAN JR, LINHARDT RJ AND SEATON BA. 2001. Annexin V-heparin oligosaccharide complex suggests heparan sulfate-mediated assembly on cell surfaces. Structure 9: 57-64.
- CAPURRO MI, XU P, SHI W, LI F, JIA A AND FILMUS J. 2008. Glypican-3 inhibits Hedgehog signaling during development by competing with patched for Hedgehog binding. Dev Cell 14: 700-711.
- CARDIN AD AND WEINTRAUB HJ. 1989. Molecular modeling of protein-glycosaminoglycan interactions. Arteriosclerosis 9: 21-32.
- CARNERO A, BLANCO-APARICIO C, RENNER O, LINK W AND LEAL JF. 2008. The PTEN/PI3K/AKT signalling pathway in cancer, therapeutic implications. Curr Cancer Drug Targets 8: 187-198.
- CASSARO CM AND DIETRICH CP. 1977. Distribution of sulfated mucopolysaccharides in invertebrates. J Biol Chem 252: 2254-2261.
- CASU B, CHOAY J, FERRO DR, GATTI G, JACQUINET JC, PETITOU M, PROVASOLI A, RAGAZZI M, SINAY P AND TORRI G. 1986. Controversial glycosaminoglycan conformations. Nature 322: 215-216.
- CHAVANTE SF, SANTOS EA, OLIVEIRA FW, GUERRINI M, TORRI G, CASU B, DIETRICH CP AND NADER HB. 2000. A novel heparan sulphate with high degree of Nsulphation and high heparin cofactor-II activity from the brine shrimp Artemia franciscana Int J Biol Macromol 27: 49-57.
- CHEN CL, HUANG SS AND HUANG JS. 2006. Cellular heparan sulfate negatively modulates transforming growth factor-beta1 (TGF-beta1) responsiveness in epithelial cells. J Biol Chem 281: 11506-11514.
- CHENG F, MANI K, VAN DEN BORN J, DING K, BELTING M AND FRANSSON LA. 2002. Nitric oxide-dependent processing of heparan sulfate in recycling S-nitrosylated glypican-1 takes place in caveolin-1-containing endosomes. J Biol Chem 277: 44431-44439.
- CHUANG WL, CHRIST MD AND RABENSTEIN DL. 2001. Determination of the primary structures of heparin- and heparan sulfate-derived oligosaccharides using band-selective homonuclear-decoupled two-dimensional 1H NMR experiments. Anal Chem 73: 2310-2316.
- COHEN MM JR. 2003. The hedgehog signaling network. Am J Med Genet A 123A: 5-28.
- COLE GJ, LOEWY A AND GLASER L. 1986. Neuronal cell-cell adhesion depends on interactions of N-CAM with heparin-like molecules. Nature 320: 445-447.
- CONRAD HE. 2001. Degradation of heparan sulfate by nitrous acid. Methods Mol Biol 171: 347-351.
- COOMBE DR, WATT SM AND PARISH CR. 1994. Mac-1 (CD11b/CD18) and CD45 mediate the adhesion of hematopoietic progenitor cells to stromal cell elements via recognition of stromal heparan sulfate. Blood 84: 739-752.
- CROWN SE, YU Y, SWEENEY MD, LEARY JA AND HANDEL TM. 2006. Heterodimerization of CCR2 Chemokines and Regulation by Glycosaminoglycan Binding. J Biol Chem 281: 25438-25446
- CZUBAYKO F, LIAUDET-COOPMAN ED, AIGNER A, TUVESON AT, BERCHEM GJ AND WELLSTEIN A. 1997. A secreted FGF-binding protein can serve as the angiogenic switch in human cancer. Nat Med 3: 1137-1140.
- DEANGELIS PL. 2002. Evolution of glycosaminoglycans and their glycosyltransferases: Implications for the extracellular matrices of animals and the capsules of pathogenic bacteria. Anat Rec 268: 317-326.
- DERKSEN PW, KEEHNEN RM, EVERS LM, VAN OERS MH, SPAARGAREN M AND PALS ST. 2002. Cell surface proteoglycan syndecan-1 mediates hepatocyte growth factor binding and promotes Met signaling in multiple myeloma. Blood 99: 1405-1410.
- DIETRICH CP, SAMPAIO LO, TOLEDO OM AND CASSARO CM. 1977. Cell recognition and adhesiveness: a possible biological role for the sulfated mucopolysaccharides. Biochem Biophys Res Commun 75: 329-336.
- DIETRICH CP, SAMPAIO LO, MONTES DE OCA H AND NADER HB. 1980. Role of sulfated mucopolysaccharides in cell recognition and neoplastic transformation. AnAcad Bras Cienc 52: 179-186.
- DIETRICH CP, NADER HB AND STRAUS AH. 1983. Structural differences of heparan sulfates according to the tissue and species of origin. Biochem Biophys Res Commun 111: 865-871.
- DIETRICH CP, TERSARIOL IL, TOMA L, MORAES CT, PORCIONATTO MA, OLIVEIRA FW AND NADER HB. 1998. Structure of heparan sulfate: identification of variable and constant oligosaccharide domains in eight heparan sulfates of different origins. Cell Mol Biol (Noisy-le-grand) 44: 417-429.
- DIETRICH CP, PAIVA JF, CASTRO RA, CHAVANTE SF, JESKE W, FAREED J, GORIN PA, MENDES A AND NADER HB. 1999. Structural features and anticoagulant activities of a novel natural low molecular weight heparin from the shrimp Penaeus brasiliensis Biochim Biophys Acta 1428: 273-283.
- DIETZ F, FRANKEN S, YOSHIDA K, NAKAMURA H, KAPPLER J AND GIESELMANN V. 2002. The family of hepatoma-derived growth factor proteins: characterization of a new member HRP-4 and classification of its subfamilies. Biochem J 366: 491-500.
- DUCHESNE L, TISSOT B, RUDD TR, DELL A AND FERNIG DG. 2006. N-glycosylation of fibroblast growth factor receptor 1 regulates ligand and heparan sulfate co-receptor binding. J Biol Chem 281: 27178-27189.
- ESCRIBA PV, WEDEGAERTNER PB, GONI FM AND VOGLER O. 2007. Lipid-protein interactions in GPCR-associated signaling. Biochim Biophys Acta 1768: 836- 852.
- FAN TC, CHANG HT, CHEN IW, WANG HY AND CHANG MD. 2007. A heparan sulfate-facilitated and raft-dependent macropinocytosis of eosinophil cationic protein. Traffic 8: 1778-1795.
- FARACH-CARSON MC AND CARSON DD. 2007. Perlecan - a multifunctional extracellular proteoglycan scaffold. Glycobiology 17: 897-905.
- FARACH-CARSON MC, BROWN AJ, LYNAM M, SAFRAN JB AND CARSON DD. 2008. A novel peptide sequence in perlecan domain IV supports cell adhesion, spreading and FAK activation. Matrix Biol 27: 150-160.
- FEARS CY AND WOODS A. 2006. The role of syndecans in disease and wound healing. Matrix Biol 25: 443-456.
- FEDARKO NS AND CONRAD HE. 1986. A unique heparan sulfate in the nuclei of hepatocytes: structural changes with the growth state of the cells. J Cell Biol 102: 587-599.
- FERREIRA TM, MEDEIROS MG, DIETRICH CP AND NADER HB. 1993. Structure of heparan sulfate from the fresh water mollusc Anomantidae sp: sequencing of its disaccharide units. Int J Biochem 25: 1219-1225.
- FERRO DR, PROVASOLI A, RAGAZZI M, CASU B, TORRI G, BOSSENNEC V, PERLY B, SINAY P, PETITOU M AND CHOAY J. 1990. Conformer populations of L-iduronic acid residues in glycosaminoglycan sequences. Carbohydr Res 195: 157-167.
- FEYZI E, LUSTIG F, FAGER G, SPILLMANN D, LINDAHL U AND SALMIVIRTA M. 1997. Characterization of heparin and heparan sulfate domains binding to the long splice variant of platelet-derived growth factor A chain. J Biol Chem 272: 5518-5524.
- FILMUS J. 2002. The contribution of in vivo manipulation of gene expression to the understanding of the function of glypicans. Glycoconj J 19: 319-323.
- FILMUS J, CAPURRO M AND RAST J. 2008. Glypicans. Genome Biol 9: 224.
- FJELDSTAD K AND KOLSET SO. 2005. Decreasing the metastatic potential in cancers-targeting the heparan sulfate proteoglycans. Curr Drug Targets 6: 665-682.
- FOX AN AND ZINN K. 2005. The heparan sulfate proteoglycan syndecan is an in vivo ligand for the Drosophila LAR receptor tyrosine phosphatase. Curr Biol 15: 1701-1711.
- FRANCO CR, ROCHA HA, TRINDADE ES, SANTOS IA, LEITE EL, VEIGA SS, NADER HB AND DIETRICH CP. 2001. Heparan sulfate and control of cell division: adhesion and proliferation of mutant CHO-745 cells lacking xylosyl transferase. Braz J Med Biol Res 34: 971-975.
- FRANSSON LA. 2003. Glypicans. Int J Biochem Cell Biol 35: 125-129.
- FRANSSON LA, BELTING M, CHENG F, JONSSON M, MANI K AND SANDGREN S. 2004. Novel aspects of glypican glycobiology. Cell Mol Life Sci 61: 1016-1024.
- FUKI IV, KUHN KM, LOMAZOV IR, ROTHMAN VL, TUSZYNSKI GP, IOZZO RV, SWENSON TL, FISHER EA AND WILLIAMS KJ. 1997. The syndecan family of proteoglycans. Novel receptors mediating internalization of atherogenic lipoproteins in vitro J Clin Invest 100: 1611-1622.
- FUKI IV, IOZZO RV AND WILLIAMS KJ. 2000. Perlecan heparan sulfate proteoglycan: a novel receptor that mediates a distinct pathway for ligand catabolism. J Biol Chem 275: 25742-25750.
- FURUKAWA K AND BHAVANANDAN VP. 1983. Influences of anionic polysaccharides on DNA synthesis in isolated nuclei and by DNA polymerase alpha: correlation of observed effects with properties of the polysaccharides. Biochim Biophys Acta 740: 466-475.
- GAGLIARDI M, PIDDINI E AND VINCENT JP. 2008. Endocytosis: a positive or a negative influence on Wnt signalling? Traffic 9: 1-9.
- GALLAGHER JT. 2006. Multiprotein signalling complexes: regional assembly on heparan sulphate. Biochem Soc Trans 34: 438-441.
- GAMBARINI AG, MIYAMOTO CA, LIMA GA, NADER HB AND DIETRICH CP. 1993. Mitogenic activity of acidic fibroblast growth factor is enhanced by highly sulfated oligosaccharides derived from heparin and heparan sulfate. Mol Cell Biochem 124: 121-129.
- GIROS A, MORANTE J, GIL-SANZ C, FAIREN A AND COSTELL M. 2007. Perlecan controls neurogenesis in the developing telencephalon. BMC Dev Biol 7: 29.
- GITAY-GOREN H, SOKER S, VLODAVSKY I AND NEUFELD G. 1992. The binding of vascular endothelial growth factor to its receptors is dependent on cell surface-associated heparin-like molecules. J Biol Chem 267: 6093-6098.
- GLASS DJ, DECHIARA TM, STITT TN, DISTEFANO PS, VALENZUELA DM AND YANCOPOULOS GD. 1996. The receptor tyrosine kinase MuSK is required for neuromuscular junction formation and is a functional receptor for agrin. Cold Spring Harb Symp Quant Biol 61: 435-444.
- GOMES PB AND DIETRICH CP. 1982. Distribution of heparin and other sulfated glycosaminoglycans in vertebrates. Comp Biochem Physiol B 73: 857-863.
- GRUNERT M, NURCOMBE V AND COOL SM. 2008. Stem cell fate decisions: the role of heparan sulfate in the control of autocrine and paracrine signals. Curr Stem Cell Res Ther 3: 1-8.
- GUERRINI M, BISIO A AND TORRI G. 2001. Combined quantitative (1)H and (13)C nuclear magnetic resonance spectroscopy for characterization of heparin preparations. Semin Thromb Hemost 27: 473-482.
- GUERRINI M, RAMAN R, VENKATARAMAN G, TORRI G, SASISEKHARAN R AND CASU B. 2002. A novel computational approach to integrate NMR spectroscopy and capillary electrophoresis for structure assignment of heparin and heparan sulfate oligosaccharides. Glycobiology 12: 713-719.
- GUMIENNY TL, MACNEIL LT, WANG H, DE BONO M, WRANA JL AND PADGETT RW. 2007. Glypican LON-2 is a conserved negative regulator of BMP-like signaling in Caenorhabditis elegans. Curr Biol 17: 159-164.
- HABUCHI H, HABUCHI O AND KIMATA K. 2004. Sulfation pattern in glycosaminoglycan: does it have a code? Glycoconj J 21: 47-52.
- HACKER U, NYBAKKEN K AND PERRIMON N. 2005. Heparan sulphate proteoglycans: the sweet side of development. Nat Rev Mol Cell Biol 6: 530-541.
- HANDEL TM, JOHNSON Z, CROWN SE, LAU EK AND PROUDFOOT AE. 2005. Regulation of protein function by glycosaminoglycans - as exemplified by chemokines. Annu Rev Biochem 74: 385-410.
- HARMER NJ. 2006. Insights into the role of heparan sulphate in fibroblast growth factor signalling. Biochem Soc Trans 34: 442-445.
- HARRISON CA, GRAY PC, VALE WW AND ROBERTSON DM. 2005. Antagonists of activin signaling: mechanisms and potential biological applications. Trends Endocrinol Metab 16: 73-78.
- HAUSSER HJ, DECKING R AND BRENNER RE. 2004. Testican- 1, an inhibitor of pro-MMP-2 activation, is expressed in cartilage. Osteoarthritis Cartilage 12: 870-877.
- HENKE CA, ROONGTA U, MICKELSON DJ, KNUTSON JR AND MCCARTHY JB. 1996. CD44-related chondroitin sulfate proteoglycan, a cell surface receptor implicated with tumor cell invasion, mediates endothelial cell migration on fibrinogen and invasion into a fibrin matrix. J Clin Invest 97: 2541-2552.
- HILEMAN RE, FROMM JR, WEILER JM AND LINHARDT RJ. 1998. Glycosaminoglycan-protein interactions: definition of consensus sites in glycosaminoglycan binding proteins. Bioessays 20: 156-167.
- HSIA E, RICHARDSON TP AND NUGENT MA. 2003. Nuclear localization of basic fibroblast growth factor is mediated by heparan sulfate proteoglycans through protein kinase C signaling. J Cell Biochem 88: 1214-1225.
- HUBER R, MAZZARELLA R, CHEN CN, CHEN E, IRELAND M, LINDSAY S, PILIA G AND CRISPONI L. 1998. Glypican 3 and glypican 4 are juxtaposed in Xq26.1. Gene 225: 9-16.
- IOZZO RV. 2005. Basement membrane proteoglycans: from cellar to ceiling. Nat Rev Mol Cell Biol 6: 646-656.
- IOZZO RV AND SAN ANTONIO JD. 2001. Heparan sulfate proteoglycans: heavy hitters in the angiogenesis arena. J Clin Invest 108: 349-355.
- ISHIHARA M, FEDARKO NS AND CONRAD HE. 1986. Transport of heparan sulfate into the nuclei of hepatocytes. J Biol Chem 261: 13575-13580.
- ISHIHARA M, GUO Y, WEI Z, YANG Z, SWIEDLER SJ, ORELLANA A AND HIRSCHBERG CB. 1993. Regulation of biosynthesis of the basic fibroblast growth factor binding domains of heparan sulfate by heparan sulfate-N-deacetylase/N-sulfotransferase expression. J Biol Chem 268: 20091-20095.
- ISHITSUKA R, KOJIMA K, UTSUMI H, OGAWA H AND MATSUMOTO I. 1998. Glycosaminoglycan binding properties of annexin IV, V, and VI. J Biol Chem 273: 9935-9941.
- JASTREBOVA N, VANWILDEMEERSCH M, RAPRAEGER AC, GIMENEZ-GALLEGO G, LINDAHL U AND SPILLMANN D. 2006. Heparan sulfate-related oligosaccharides in ternary complex formation with fibroblast growth factors 1 and 2 and their receptors. J Biol Chem 281: 26884-26892.
- JERONIMO SM, SALES AO, FERNANDES MZ, MELO FP, SAMPAIO LO, DIETRICH CP AND NADER HB. 1994. Glycosaminoglycan structure and content differ according to the origins of human tumors. Braz J Med Biol Res 27: 2253-2258.
- JIANG X AND COUCHMAN JR. 2003. Perlecan and tumor angiogenesis. J Histochem Cytochem 51: 1393-1410.
- JOHNSON KG ET AL. 2006. The HSPGs Syndecan and Dally-like bind the receptor phosphatase LAR and exert distinct effects on synaptic development. Neuron 49: 517-531.
- JOHNSON Z, PROUDFOOT AE AND HANDEL TM. 2005. Interaction of chemokines and glycosaminoglycans: a new twist in the regulation of chemokine function with opportunities for therapeutic intervention. Cytokine Growth Factor Rev 16: 625-636.
- JURY EC, ELDRIDGE J, ISENBERG DA AND KABOURIDIS PS. 2007. Agrin signalling contributes to cell activation and is overexpressed inTlymphocytes from lupus patients. J Immunol 179: 7975-7983.
- KAYED H, KLEEFF J, KELEG S, JIANG X, PENZEL R, GIESE T, ZENTGRAF H, BUCHLER MW, KORC M AND FRIESS H. 2006. Correlation of glypican-1 expression with TGF-beta, BMP, and activin receptors in pancreatic ductal adenocarcinoma. Int J Oncol 29: 1139-1148.
- KEMP LE, MULLOY B AND GHERARDI E. 2006. Signalling by HGF/SF and Met: the role of heparan sulphate coreceptors. Biochem Soc Trans 34: 414-417.
- KIRKPATRICK CA AND SELLECK SB. 2007. Heparan sulfate proteoglycans at a glance. J Cell Sci 120: 1829-1832.
- KNOX SM AND WHITELOCK JM. 2006. Perlecan: how does one molecule do so many things? Cell Mol Life Sci 63: 2435-2445.
- KOOPMANN W, EDIRIWICKREMA C AND KRANGEL MS. 1999. Structure and function of the glycosaminoglycan binding site of chemokine macrophage - inflammatory protein-1 beta. J Immunol 163: 2120-2127.
- KREUGER J, SPILLMANN D, LI JP AND LINDAHL U. 2006. Interactions between heparan sulfate and proteins: the concept of specificity. J Cell Biol 174: 323-327.
- KRILLEKE D, DEERKENEZ A, SCHUBERT W, GIRI I, ROBINSON GS, NG YS AND SHIMA DT. 2007. Molecular mapping and functional characterization of the VEGF- 164 heparin-binding domain. J Biol Chem 282: 28045-28056.
- LAU EK, ALLEN S, HSU AR AND HANDEL TM. 2004. Chemokine-receptor interactions: GPCRs, glycosaminoglycans and viral chemokine binding proteins. Adv Protein Chem 68: 351-391.
- LEICHT DT, BALAN V, KAPLUN A, SINGH-GUPTA V, KAPLUN L, DOBSON M AND TZIVION G. 2007. Raf kinases: function, regulation and role in human cancer. Biochim Biophys Acta 1773: 1196-1212.
- LEWIS KA, GRAY PC, BLOUNT AL, MACCONELL LA, WIATER E, BILEZIKJIAN LM AND VALE W. 2000. Betaglycan binds inhibin and can mediate functional antagonism of activin signalling. Nature 404: 411-414.
- LINDNER JR, HILLMAN PR, BARRETT AL, JACKSON MC, PERRY TL, PARK Y AND DATTA S. 2007. The Drosophila Perlecan gene trol regulates multiple signaling pathways in different developmental contexts. BMC Dev Biol 7: 121.
- LOO BM, KREUGER J, JALKANEN M, LINDAHL U AND SALMIVIRTA M. 2001. Binding of heparin/heparan sulfate to fibroblast growth factor receptor 4. J Biol Chem 276: 16868-16876.
- LOPES CC, DIETRICH CP AND NADER HB. 2006a. Specific structural features of syndecans and heparan sulfate chains are needed for cell signaling. Braz J Med Biol Res 39: 157-167.
- LOPES CC, TOMA L, PINHAL MA, PORCIONATTO MA, SOGAYAR MC, DIETRICH CP AND NADER HB. 2006b. EJ-ras oncogene transfection of endothelial cells upregulates the expression of syndecan-4 and downregulates heparan sulfate sulfotransferases and epimerase. Biochimie 88: 1493-1504.
- LORTAT-JACOB H, GROSDIDIER A AND IMBERTY A. 2002. Structural diversity of heparan sulfate binding domains in chemokines. Proc Natl Acad Sci USA 99: 1229-1234.
- MA YQ AND GENG JG. 2000. Heparan sulfate-like proteoglycans mediate adhesion of human malignant melanoma A375 cells to P-selectin under flow. J Immunol 165: 558-565.
- MIYAZONO K. 1997. TGF-beta receptors and signal transduction. Int J Hematol 65: 97-104.
- MOBLI M, NILSSON M AND ALMOND A. 2008. The structural plasticity of heparan sulfate NA-domains and hence their role in mediating multivalent interactions is confirmed by high-accuracy (15)N-NMR relaxation studies. Glycoconj J 25: 401-414.
- MOHAMMADI M, OLSEN SK AND GOETZ R. 2005a. A protein canyon in the FGF-FGF receptor dimer selects from an a la carte menu of heparan sulfate motifs. Curr Opin Struct Biol 15: 506-516.
- MOHAMMADI M, OLSEN SK AND IBRAHIMI OA. 2005b. Structural basis for fibroblast growth factor receptor activation. Cytokine Growth Factor Rev 16: 107-137.
- MONGIAT M, TAYLOR K, OTTO J, AHO S, UITTO J, WHITELOCK JM AND IOZZO RV. 2000. The protein core of the proteoglycan perlecan binds specifically to fibroblast growth factor-7. J Biol Chem 275: 7095-7100.
- MOREIRA CR, LOPES CC, CUCCOVIA IM, PORCIONATTO MA, DIETRICH CP AND NADER HB. 2004. Heparan sulfate and control of endothelial cell proliferation: increased synthesis during the S phase of the cell cycle and inhibition of thymidine incorporation induced by ortho-nitrophenyl- beta-D-xylose. Biochim Biophys Acta 1673: 178-185.
- MORGAN MR, HUMPHRIES MJ AND BASS MD. 2007. Synergistic control of cell adhesion by integrins and syndecans. Nat Rev Mol Cell Biol 8: 957-969.
- MULLOY B. 2005. The specificity of interactions between proteins and sulfated polysaccharides. An Acad Bras Cienc 77: 651-664.
- MULLOY B AND FORSTER MJ. 2000. Conformation and dynamics of heparin and heparan sulfate. Glycobiology 10: 1147-1156.
- MULLOY B AND LINHARDT RJ. 2001. Order out of complexity- protein structures that interact with heparin. Curr Opin Struct Biol 11: 623-628.
- MURRAY AJ. 2008. Pharmacological PKA inhibition: all may not be what it seems. Sci Signal 1: re4.
- NADER HB, TAKAHASHI HK, STRAUS AH AND DIETRICH CP. 1980. Selective distribution of the heparin in mammals: conspicuous presence of heparin in lymphoid tissues. Biochim Biophys Acta 627: 40-48.
- NADER HB, FERREIRA TM, PAIVA JF, MEDEIROS MG, JERONIMO SM, PAIVA VM AND DIETRICH CP. 1984. Isolation and structural studies of heparan sulfates and chondroitin sulfates from three species of molluscs. J Biol Chem 259: 1431-1435.
- NADER HB, DIETRICH CP, BUONASSISI V AND COLBURN P. 1987. Heparin sequences in the heparan sulfate chains of an endothelial cell proteoglycan. Proc Natl Acad Sci USA 84: 3565-3569.
- NADER HB, PORCIONATTO MA, TERSARIOL IL, PINHAL MA, OLIVEIRA FW, MORAES CT AND DIETRICH CP. 1990. Purification and substrate specificity of heparitinase I and heparitinase II from Flavobacterium heparinum Analyses of the heparin and heparan sulfate degradation products by 13C NMR spectroscopy. J Biol Chem 265: 16807-16813.
- NADER HB ET AL. 1999a. Heparan sulfates and heparins: similar compounds performing the same functions in vertebrates and invertebrates? Braz J Med Biol Res 32: 529-538.
- NADER HB, KOBAYASHI EY, CHAVANTE SF, TERSARIOL IL, CASTRO RA, SHINJO SK, NAGGI A, TORRI G, CASU B AND DIETRICH CP. 1999b. New insights on the specificity of heparin and heparan sulfate lyases from Flavobacterium heparinum revealed by the use of synthetic derivatives of K5 polysaccharide from E. coli and 2-O-desulfated heparin. Glycoconj J 16: 265-270.
- NAKADA M, MIYAMORI H, YAMASHITA J AND SATO H. 2003. Testican 2 abrogates inhibition of membrane-type matrix metalloproteinases by other testican family proteins. Cancer Res 63: 3364-3369.
- NASCIMENTO FD, RIZZI CC, NANTES IL, STEFE I, TURK B, CARMONA AK, NADER HB, JULIANO L AND TERSARIOL IL. 2005. Cathepsin X binds to cell surface heparan sulfate proteoglycans. Arch Biochem Biophys 436: 323-332.
- NASCIMENTO FD, HAYASHI MA, KERKIS A, OLIVEIRA V, OLIVEIRA EB, RADIS-BAPTISTA G, NADER HB, YAMANE T, TERSARIOL IL AND KERKIS I. 2007. Crotamine mediates gene delivery into cells through the binding to heparan sulfate proteoglycans. J Biol Chem 282: 21349-21360.
- NG YS, KRILLEKE D AND SHIMA DT. 2006. VEGF function in vascular pathogenesis. Exp Cell Res 312: 527-537.
- NGO ST, NOAKES PG AND PHILLIPS WD. 2007. Neural agrin: a synaptic stabiliser. Int J Biochem Cell Biol 39: 863-867.
- NOTI C AND SEEBERGER PH. 2005. Chemical approaches to define the structure-activity relationship of heparin-like glycosaminoglycans. Chem Biol 12: 731-756.
- NOZIK-GRAYCK E, SULIMAN HB AND PIANTADOSI CA. 2005. Extracellular superoxide dismutase. Int J Biochem Cell Biol 37: 2466-2471.
- NUGENT MA AND IOZZO RV. 2000. Fibroblast growth factor- 2. Int J Biochem Cell Biol 32: 115-120.
- NUNES SS, OUTEIRO-BERNSTEIN MA, JULIANO L, VARDIERO F, NADER HB, WOODS A, LEGRAND C AND MORANDI V. 2008. Syndecan-4 contributes to endothelial tubulogenesis through interactions with two motifs inside the pro-angiogenic N-terminal domain of thrombospondin-1. J Cell Physiol 214: 828-837.
- OBA-SHINJO SM ET AL. 2006. Melanocyte transformation associated with substrate adhesion impediment. Neoplasia 8: 231-241.
- OH ES AND COUCHMAN JR. 2004. Syndecans-2 and -4; close cousins, but not identical twins. Mol Cells 17: 181- 187.
- PADERA R, VENKATARAMAN G, BERRY D, GODAVARTI R AND SASISEKHARAN R. 1999. FGF-2/fibroblast growth factor receptor/heparin-like glycosaminoglycan interactions: a compensation model for FGF-2 signaling. Faseb J 13: 1677-1687.
- PANKONIN MS, GALLAGHER JT AND LOEB JA. 2005. Specific structural features of heparan sulfate proteoglycans potentiate neuregulin-1 signaling. J Biol Chem 280: 383-388.
- PARISH CR. 2006. The role of heparan sulphate in inflammation. Nat Rev Immunol 6: 633-643.
- PATEL VN, LIKAR KM, ZISMAN-ROZEN S, COWHERD SN, LASSITER KS, SHER I, YATES EA, TURNBULL JE, RON D AND HOFFMAN MP. 2008. Specific heparan sulfate structures modulate FGF10-mediated submandibular gland epithelial morphogenesis and differentiation. J Biol Chem 283: 9308-9317.
- PATEY SJ, EDWARDS EA, YATES EA AND TURNBULL JE. 2008. Engineered heparins: novel beta-secretase inhibitors as potential Alzheimer's disease therapeutics. Neurodegener Dis 5: 197-199.
- PELLEGRINI L, BURKE DF, VON DELFT F, MULLOY B AND BLUNDELL TL. 2000. Crystal structure of fibroblast growth factor receptor ectodomain bound to ligand and heparin. Nature 407: 1029-1034.
- PERETTI T, WAISBERG J, MADER AM, DE MATOS LL, DA COSTA RB, CONCEIÇÃO GM, LOPES AC, NADER HB AND PINHAL MA. 2008. Heparanase-2, syndecan-1, and extracellular matrix remodeling in colorectal carcinoma. Eur J Gastroenterol Hepatol 20: 756-765.
- POON GM AND GARIEPY J. 2007. Cell-surface proteoglycans as molecular portals for cationic peptide and polymer entry into cells. Biochem Soc Trans 35: 788-793.
- PORCIONATTO MA, PINTO CR, DIETRICH CP AND NADER HB. 1994. Heparan sulfate proteoglycan and control of cell proliferation: enhanced synthesis induced by phorbol ester (PMA) during G(1)-phase. Braz J Med Biol Res 27: 2185-2190.
- PORCIONATTO MA, MOREIRA CR, LOTFI CF, ARMELIN HA, DIETRICH CP AND NADER HB. 1998. Stimulation of heparan sulfate proteoglycan synthesis and secretion during G1 phase induced by growth factors and PMA. J Cell Biochem 70: 563-572.
- PORCIONATTO MA, NADER HB AND DIETRICH CP. 1999. Heparan sulfate and cell division. Braz J Med Biol Res 32: 539-544.
- POWELL AK, YATES EA, FERNIG DG AND TURNBULL JE. 2004. Interactions of heparin/heparan sulfate with proteins: appraisal of structural factors and experimental approaches. Glycobiology 14: 17R-30R.
- POWERS CJ, MCLESKEY SW AND WELLSTEIN A. 2000. Fibroblast growth factors, their receptors and signaling. Endocr Relat Cancer 7: 165-197.
- PRESTA M, DELL'ERA P, MITOLA S, MORONI E, RONCA R AND RUSNATI M. 2005. Fibroblast growth factor/fibroblast growth factor receptor system in angiogenesis. Cytokine Growth Factor Rev 16: 159-178.
- QUARTO N AND AMALRIC F. 1994. Heparan sulfate proteoglycans as transducers of FGF-2 signalling. J Cell Sci 107 (Pt 11): 3201-3212.
- RAGAZZI M, FERRO DR, PROVASOLI A, PUMILIA P, CASSINARI A, TORRI G, GUERRINI M, CASU B, NADER HB AND DIETRICH CP. 1993. Conformation of the Unsaturated Uronic Acid Residues of Glycosaminoglycan Disaccharides. J Carbohy Chem 12: 513-535.
- RAMAN R, SASISEKHARAN V AND SASISEKHARAN R. 2005. Structural insights into biological roles of proteinglycosaminoglycan interactions. Chem Biol 12: 267-277.
- RAPRAEGER AC, KRUFKA A AND OLWIN BB. 1991. Requirement of heparan sulfate for bFGF-mediated fibroblast growth and myoblast differentiation. Science 252: 1705-1708.
- RAPRAEGER AC, GUIMOND S, KRUFKA A AND OLWIN BB. 1994. Regulation by heparan sulfate in fibroblast growth factor signaling. Methods Enzymol 245: 219-240.
- RIDER CC. 2006. Heparin/heparan sulphate binding in the TGF-beta cytokine superfamily. Biochem Soc Trans 34: 458-460.
- ROHDE LH, JANATPORE MJ, MCMASTER MT, FISHER S, ZHOU Y, LIM KH, FRENCH M, HOKE D, JULIAN J AND CARSON DD. 1998. Complementary expression of HIP, a cell-surface heparan sulfate binding protein, and perlecan at the human fetal-maternal interface. Biol Reprod 58: 1075-1083.
- ROLNY C, SPILLMANN D, LINDAHL U AND CLAESSONWELSH L. 2002. Heparin amplifies platelet-derived growth factor (PDGF)- BB-induced PDGF alpha-receptor but not PDGF beta-receptor tyrosine phosphorylation in heparan sulfate-deficient cells. Effects on signal transduction and biological responses. J Biol Chem 277: 19315-19321.
- RUBIN JS, DAY RM, BRECKENRIDGE D, ATABEY N, TAYLOR WG, STAHL SJ, WINGFIELD PT, KAUFMAN JD, SCHWALL R AND BOTTARO DP. 2001. Dissociation of heparan sulfate and receptor binding domains of hepatocyte growth factor reveals that heparan sulfate-c-met interaction facilitates signaling. J Biol Chem 276: 32977-32983.
- RUHRBERG C, GERHARDT H, GOLDING M, WATSON R, IOANNIDOU S, FUJISAWA H, BETSHOLTZ C AND SHIMA DT. 2002. Spatially restricted patterning cues provided by heparin-binding VEGF-A control blood vessel branching morphogenesis. Genes Dev 16: 2684-2698.
- SAITO Y ET AL. 2007. A peptide derived from tenascin-C induces beta1 integrin activation through syndecan-4. J Biol Chem 282: 34929-34937.
- SAKSELA O, MOSCATELLI D, SOMMER A AND RIFKIN DB. 1988. Endothelial cell-derived heparan sulfate binds basic fibroblast growth factor and protects it from proteolytic degradation. J Cell Biol 107: 743-751.
- SAMPAIO L, TERSARIOL I, LOPES C, BOUÇAS R, NASCIMENTO F, ROCHA H AND NADER H. 2006. Heparins and heparans sulfates. Structure, distribution and protein interactions. In: VERLI H (Ed), INSIGHTS INTO CARBOHYDRATE STRUCTURE AND BIOLOGICAL FUNCTION. Transworld Research Network, Kerala, p. 1-24.
- SANDERSON RD AND YANG Y. 2008. Syndecan-1: a dynamic regulator of the myeloma microenvironment. Clin Exp Metastasis 25: 149-159.
- SASAKI S ET AL. 1999. Ligand-induced recruitment of a histone deacetylase in the negative-feedback regulation of the thyrotropin beta gene. Embo J 18: 5389-5398.
- SASAKI N ET AL. 2008. Heparan sulfate regulates selfrenewal and pluripotency of embryonic stem cells. J Biol Chem 283: 3594-3606.
- SASISEKHARAN R, SHRIVER Z, VENKATARAMAN G AND NARAYANASAMI U. 2002. Roles of heparan-sulphate glycosaminoglycans in cancer. Nat Rev Cancer 2: 521-528.
- SCHNEPP A, KOMP LINDGREN P, HULSMANN H, KROGER S, PAULSSON M AND HARTMANN U. 2005. Mouse testican- 2. Expression, glycosylation, and effects on neurite outgrowth. J Biol Chem 280: 11274-11280.
- SCHWALL RH, CHANG LY, GODOWSKI PJ, KAHN DW, HILLAN KJ, BAUER KD AND ZIONCHECK TF. 1996. Heparin induces dimerization and confers proliferative activity onto the hepatocyte growth factor antagonists NK1 and NK2. J Cell Biol 133: 709-718.
- SHINJO SK, TERSARIOL IL, OLIVEIRA V, NAKAIE CR, OSHIRO ME, FERREIRA AT, SANTOS IA, DIETRICH CP AND NADER HB. 2002. Heparin and heparan sulfate disaccharides bind to the exchanger inhibitor peptide region of Na+/Ca2+ exchanger and reduce the cytosolic calcium of smooth muscle cell lines. Requirement of C4- C5 unsaturation and 1 → 4 glycosidic linkage for activity. J Biol Chem 277: 48227-48233.
- SMIRNOV SP, BARZAGHI P, MCKEE KK, RUEGG MA AND YURCHENCO PD. 2005. Conjugation of LG domains of agrins and perlecan to polymerizing laminin-2 promotes acetylcholine receptor clustering. J Biol Chem 280: 41449-41457.
- SONCIN F, STRYDOM DJ AND SHAPIRO R. 1997. Interaction of heparin with human angiogenin. J Biol Chem 272: 9818-9824.
- SONG HH, SHI W, XIANG YY AND FILMUS J. 2005. The loss of glypican-3 induces alterations in Wnt signaling. J Biol Chem 280: 2116-2125.
- SPIVAK-KROIZMAN T, LEMMON MA, DIKIC I, LADBURY JE, PINCHASI D, HUANG J, JAYE M, CRUMLEY G, SCHLESSINGER J AND LAX I. 1994. Heparin-induced oligomerization of FGF molecules is responsible for FGF receptor dimerization, activation, and cell proliferation. Cell 79: 1015-1024.
- STIGLIANO I, PURICELLI L, FILMUS J, SOGAYAR MC, BAL DE KIER JOFFE E AND PETERS MG. 2009. Glypican-3 regulates migration, adhesion and actin cytoskeleton organization in mammary tumor cells through Wnt signaling modulation. Breast Cancer Res Treat 114(2): 251-262.
- STRAUS AH, NADER HB, TAKAHASHI HK AND DIETRICH CP. 1982. Ontogeny of heparin in mammals: a correlation with the appearance of mast cells in tissues. An Acad Bras Cienc 54: 439-448.
- STRINGER SE. 2006. The role of heparan sulphate proteoglycans in angiogenesis. Biochem Soc Trans 34: 451-453.
- SU G, BLAINE SA, QIAO D AND FRIEDL A. 2007. Shedding of syndecan-1 by stromal fibroblasts stimulates human breast cancer cell proliferation via FGF2 activation. J Biol Chem 282: 14906-14915.
- SUAREZ ER, NOHARA AS, MATAVELI FD, DE MATOS LL, NADER HB AND PINHAL MA. 2007. Glycosaminoglycan synthesis and shedding induced by growth factors are cell and compound specific. Growth Factors 25: 50-59.
- SUGAYA N, HABUCHI H, NAGAI N, ASHIKARI-HADA S AND KIMATA K. 2008. 6-O-sulfation of heparan sulfate differentially regulates various fibroblast growth factordependent signalings in culture. J Biol Chem 283: 10366-10376.
- SULPICE E, BRYCKAERT M, LACOUR J, CONTRERES JO AND TOBELEM G. 2002. Platelet factor 4 inhibits FGF2- induced endothelial cell proliferation via the extracellular signal-regulated kinase pathway but not by the phosphatidylinositol 3-kinase pathway. Blood 100: 3087-3094.
- TANTRAVAHI RV, STEVENS RL, AUSTEN KF AND WEIS JH. 1986. A single gene in mast cells encodes the core peptides of heparin and chondroitin sulfate proteoglycans. Proc Natl Acad Sci USA 83: 9207-9210.
- TAYLOR KR AND GALLO RL. 2006. Glycosaminoglycans and their proteoglycans: host-associated molecular patterns for initiation and modulation of inflammation. Faseb J 20: 9-22.
- TELCI D, WANG Z, LI X, VERDERIO EA, HUMPHRIES MJ, BACCARINI M, BASAGA H AND GRIFFIN M. 2008. Fibronectin-Tissue Transglutaminase Matrix Rescues RGD-impaired Cell Adhesion through Syndecan-4 and beta1 Integrin Co-signaling. J BiolChem283: 20937-20947.
- TERSARIOL IL, DIETRICH CP AND NADER HB. 1992. Interaction of heparin with myosin ATPase: possible involvement with the hemorrhagic activity and a correlation with antithrombin III high affinity-heparin molecules. Thromb Res 68: 247-258.
- TERSARIOL IL, FERREIRA TM, MEDEIROS MG, PORCIONATTO MA, MORAES CT, ABREU LR, NADER HB AND DIETRICH CP. 1994. Sequencing of heparan sulfate proteoglycans: identification of variable and constant oligosaccharide regions in eight heparan sulfate proteoglycans of different origins. Braz J Med Biol Res 27: 2097-2102.
- TKACHENKO E, LUTGENS E, STAN RV AND SIMONS M. 2004. Fibroblast growth factor 2 endocytosis in endothelial cells proceed via syndecan-4-dependent activation of Rac1 and a Cdc42-dependent macropinocytic pathway. J Cell Sci 117: 3189-3199.
- TKACHENKO E, RHODES JM AND SIMONS M. 2005. Syndecans: new kids on the signaling block. Circ Res 96: 488-500.
- TOLEDO OM AND DIETRICH CP. 1977. Tissue specific distribution of sulfated mucopolysaccharides in mammals. Biochim Biophys Acta 498: 114-122.
- TOUROVSKAIA A, LI N AND FOLCH A. 2008. Localized acetylcholine receptor clustering dynamics in response to microfluidic focal stimulation with agrin. Biophys J 95: 3009-3016.
- TRAISTER A, SHI W AND FILMUS J. 2008. Mammalian Notum induces the release of glypicans and other GPIanchored proteins from the cell surface. Biochem J 410: 503-511.
- UTANI A, NOMIZU M, MATSUURA H, KATO K, KOBAYASHI T, TAKEDA U, AOTA S, NIELSEN PK AND SHINKAI H. 2001. A unique sequence of the laminin alpha 3 G domain binds to heparin and promotes cell adhesion through syndecan-2 and -4. J Biol Chem 276: 28779-28788.
- VEIGA SS, ELIAS M, GREMSKI W, PORCIONATTO MA, DA SILVA R, NADER HB AND BRENTANI RR. 1997. Post-translational modifications of alpha5beta1 integrin by glycosaminoglycan chains. The alpha5beta1 integrin is a facultative proteoglycan. J Biol Chem 272: 12529-12535.
- VEUGELERS M, VERMEESCH J, REEKMANS G, STEINFELD R, MARYNEN P AND DAVID G. 1997. Characterization of glypican-5 and chromosomal localization of human GPC5, a new member of the glypican gene family. Genomics 40: 24-30.
- VEUGELERS M, DE CAT B, CEULEMANS H, BRUYSTENS AM, COOMANS C, DURR J, VERMEESCH J, MARYNEN P AND DAVID G. 1999. Glypican-6, a new member of the glypican family of cell surface heparan sulfate proteoglycans. J Biol Chem 274: 26968-26977.
- VIVES RR, CRUBLET E, ANDRIEU JP, GAGNON J, ROUSSELLE P AND LORTAT-JACOB H. 2004. A novel strategy for defining critical amino acid residues involved in protein/glycosaminoglycan interactions. J Biol Chem 279: 54327-54333.
- VLODAVSKY I, GOLDSHMIDT O, ZCHARIA E, ATZMON R, RANGINI-GUATTA Z, ELKIN M, PERETZ T AND FRIEDMANN Y. 2002. Mammalian heparanase: involvement in cancer metastasis, angiogenesis and normal development. Semin Cancer Biol 12: 121-129.
- WALKER A, TURNBULL JE AND GALLAGHER JT. 1994. Specific heparan sulfate saccharides mediate the activity of basic fibroblast growth factor. J Biol Chem 269: 931-935.
- WATT SM, WILLIAMSON J, GENEVIER H, FAWCETT J, SIMMONS DL, HATZFELD A, NESBITT SA AND COOMBE DR. 1993. The heparin binding PECAM-1 adhesion molecule is expressed by CD34+ hematopoietic precursor cells with early myeloid and B-lymphoid cell phenotypes. Blood 82: 2649-2663.
- WHITELOCK JM, MURDOCH AD, IOZZO RV AND UNDERWOOD PA. 1996. The degradation of human endothelial cell-derived perlecan and release of bound basic fibroblast growth factor by stromelysin, collagenase, plasmin, and heparanases. J Biol Chem 271: 10079-10086.
- WIEDLOCHA A AND SORENSEN V. 2004. Signaling, internalization, and intracellular activity of fibroblast growth factor. Curr Top Microbiol Immunol 286: 45-79.
- WILCOX-ADELMAN SA, DENHEZ F AND GOETINCK PF. 2002. Syndecan-4 modulates focal adhesion kinase phosphorylation. J Biol Chem 277: 32970-32977.
- WILKINS-PORT CE AND MCKEOWN-LONGO PJ. 1996. Heparan sulfate proteoglycans function in the binding and degradation of vitronectin by fibroblast monolayers. Biochem Cell Biol 74: 887-897.
- WILLIAMS S, RYAN C AND JACOBSON C. 2008. Agrin and neuregulin, expanding roles and implications for therapeutics. Biotechnol Adv 26: 187-201.
- WOJTAL KA, HOEKSTRA D AND VAN IJZENDOORN SC. 2008. cAMP-dependent protein kinase A and the dynamics of epithelial cell surface domains: moving membranes to keep in shape. Bioessays 30: 146-155.
- WOODS A AND COUCHMAN JR. 2001. Syndecan-4 and focal adhesion function. Curr Opin Cell Biol 13: 578-583.
- WU DQ, KAN MK, SATO GH, OKAMOTO T AND SATO JD. 1991. Characterization and molecular cloning of a putative binding protein for heparin-binding growth factors. J Biol Chem 266: 16778-16785.
- YATES EA, GUIMOND SE AND TURNBULL JE. 2004. Highly diverse heparan sulfate analogue libraries: providing access to expanded areas of sequence space for bioactivity screening. J Med Chem 47: 277-280.
- YAYON A, KLAGSBRUN M, ESKO JD, LEDER P AND ORNITZ DM. 1991. Cell surface, heparin-like molecules are required for binding of basic fibroblast growth factor to its high affinity receptor. Cell 64: 841-848.
- YU WH AND WOESSNER JF JR. 2000. Heparan sulfate proteoglycans as extracellular docking molecules for matrilysin (matrix metalloproteinase 7). J Biol Chem 275: 4183-4191.
- YU WH, YU S, MENG Q, BREW K AND WOESSNER JF JR. 2000. TIMP-3 binds to sulfated glycosaminoglycans of the extracellular matrix. J Biol Chem 275: 31226-31232.
- ZHANG F, MCLELLAN JS, AYALA AM, LEAHY DJ AND LINHARDT RJ. 2007. Kinetic and structural studies on interactions between heparin or heparan sulfate and proteins of the hedgehog signaling pathway. Biochemistry 46: 3933-3941.
Publication Dates
-
Publication in this collection
19 Aug 2009 -
Date of issue
Sept 2009
History
-
Received
26 Aug 2008 -
Accepted
08 Oct 2008