Abstracts
A very large pterosaur (MN 6594-V) from the Romualdo Formation (Aptian/Albian), Santana Group, Araripe Basin, is described. The specimen is referred to Tropeognathus cf. T. mesembrinus mainly due to the presence of a low and blunt frontoparietal crest, the comparatively low number of teeth and the inclined dorsal part of the occipital region. Two distinct wingspan measurements for pterosaurs are introduced: the maximized wingspan (maxws), which essentially consists of doubling the addition of all wing elements and the length of the scapula or the coracoid (the smaller of the two), and the normal wingspan (nws), which applies a reducing factor (rfc) to the maximized wingspan to account for the natural flexures of the wing. The rfc suggested for pteranodontoids is 5%. In the case of MN 6594-V, the maxws and nws are 8.70 m and 8.26 m, respectively, making it the largest pterosaur recovered from Gondwana so far. The distal end of a larger humerus (MCT 1838-R) and a partial wing (MPSC R 1395) are also described showing that large to giant flying reptiles formed a significant part of the pterosaur fauna from the Romualdo Formation. Lastly, some comments on the nomenclatural stability of the Santana deposits are presented.
Pterosauria; Anhangueridae; Tropeognathus ; Cretaceous; Gondwana
Um grande pterossauro (MN 6594-V) da Formação Romualdo (Aptiano/Albiano), Grupo Santana, Bacia do Araripe, é descrito. O espécime é referido a Tropeognathus cf. T. mesembrinus principalmente pela crista frontoparietal baixa e obtusa, por um número comparativamente baixo de dentes e pela inclinação da porção dorsal da região occipital. Duas distintas medidas de envergadura para pterossauros são introduzidas: a envergadura maximizada (maxws), que consiste essencialmente no dobro da soma de todos os elementos da asa e o comprimento da escápula ou do coracóide (o menor de ambos), e a envergadura normal (nws), que aplica um fator de redução (rdc) à envergadura maximizada para compensar as curvaturas naturais da asa. Para pteranodontóides o rfc sugerido é de 5%. No caso de MN 6594-V, maxws e nws correspondem a 8,70 m e 8,26 m, respectivamente, o que faz deste espécime o maior pterossauro coletado do Gondwana até hoje. A extremidade distal de um úmero de maior tamanho (MCT 1838-R) e uma asa parcial (MPSC R 1395) também são aqui descritos, mostrando que pterossauros grandes ou gigantescos formaram uma parcela significativa da fauna de pterossauros da Formação Romualdo. Finalmente, alguns comentários sobre a estabilidade nomenclatural para os depósitos de Santana são feitos.
Pterosauria; Anhangueridae; Tropeognathus ; Cretáceo; Gondwana
INTRODUCTION
The pterosaur fauna from the Araripe Basin is worldwide known for the comparatively large number of specimens with exceptional preservation (e.g., Price 1971Pons D, Berthou PY and Campos DA. 1990. Quelques observations sur la palynologie de l'Aptien supérieur et del'Albien do bassin d'Araripe (N-E du Brasil). In: Campos DA, Viana MSS, Brito PM, Beurlen G (Eds), Atas do Simpósio sobre a Bacia do Araripe e Bacias interiores do Nordeste, Crato, p. 142-252., Wellnhofer 1991aWellnhofer P. 1985. Neue Pterosaurier aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 187: 105-182., Kellner and Campos 2002Kellner AWA and Campos DA. 2002. The function of the cranial crest and jaws of a unique pterosaur from the Early Cretaceous of Brazil. Science 297: 389-392.), occasionally including soft tissue (e.g., Kellner 1996Kellner AWA. 1996. Reinterpretation of a remarkably well-preserved pterosaur soft tissue from the Early Cretaceous of Brazil. J Vert Paleont 16: 718-722., Frey et al. 2003bFrey E, Tischlinger H, Buchy MC and Martill DM. 2003b. New specimens of Pterosauria (Reptilia) with soft parts with implications for pterosaurian anatomy and locomotion. Geol Soc London, Spec Pub 217: 233-266.). As has been pointed out numerous times in the literature, two very distinct Lagerstätten of that basin have yielded pterosaur material, the Crato and the Romualdo Formations, whose ages are generally regarded as being Aptian and Aptian/Albian, respectively (e.g., Pons et al. 1990NOWAK RM. 1999. Walker's Mammals of the World, 6th ed., Baltimore and London: The John Hopkins University Press, 1936 p., Maisey 1991Maisey J. (Ed). 1991. Santana fossils: an illustrated Atlas. Neptune, New Jersey: T.F.H. Publications, 459 p., Kellner and Campos 1999Kellner AWA and Campos DA. 1999. Vertebrate Paleontology in Brazil – a review. Episodes 22: 238-251., Neumann and Cabrera 1999Martill DM and Unwin DM. 2012. The world's largest toothed pterosaur, NHMUK R481, an incomplete rostrum of Coloborhynchus capito (Seeley, 1870) from the Cambridge Greensand of England. Cretaceous Res 34: 1-9., Valença et al. 2003Small H. 1913. Geologia e suprimento de agua subterranea no Ceará e parte do Piaui. Insp. Obras Contra Secas, Serie Geologia 25: 1-180.).
Up to date, 27 species from these deposits were formally introduced (Campos and Kellner 1985Campos DA and Kellner AWA. 1985. Panorama of the Flying Reptiles Study in Brazil and South America. An Acad Bras Cienc 57: 453-466., Witton 2009Wellnhofer P. 1991a. Weitere Pterosaurierfunde aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 215: 43-101., Eck et al. 2011Eck K, Elgin RA and Frey E. 2011. On the osteology of Tapejara wellnhoferi Kellner 1989 and the first occurrence of multiple specimen assemblage from the Santana Formation, Araripe Basin, NE-Brazil. Swiss J Palaeontol 130: 277-296.), but the validity and the taxonomic assignment of some taxa have been subject of controversy (e.g., Fastnacht 2001Fastnacht M. 2001. First record of Coloborhynchus (Pterosauria) from the Santana Formation (Lower Cretaceous) of the Chapada do Araripe, Brazil. Paläontol Z 75: 3-36., Veldmeijer 2003Valença LMM, Neumann VH and Mabesoone JM. 2003. An overview on Callovian-Cenomanian intracratonic basins of Northeast Brazil: onshore stratrigraphic record of the opening of the southern Atlantic. Geol Acta 1: 261-275., Rodrigues and Kellner 2008Price LI. 1971. A presença de Pterosauria no Cretáceo Inferior da Chapada do Araripe, Brasil. An Acad Bras Cienc 43(Supl): 451-461., Martill and Unwin 2012Martill DM, Frey E, Sadaqah RM and Khoury HN. 1998. Discovery of the holotype of the giant pterosaur Titanopteryx philadelphiae Arambourg 1959, and the status of Arambourgiania and Quetzalcoatlus. N Jb Geol Paläont Abh 207: 57-76.).
Here we describe a partial skeleton of a toothed pterosaur recovered from the Romualdo Formation. The material (MN 6594-V) includes several parts of the skull, lower jaw and postcranial elements that are referred to the anhanguerid pterodactyloid Tropeognathus cf. T. mesembrinus Wellnhofer, 1987Wang XL, Kellner AWA, Jiang SX, Cheng X, Meng X and Rodrigues T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. An Acad Bras Cienc 82: 1045-1062.. Until now this taxon was only known by a complete skull and mandible (Wellnhofer 1987Wang XL, Kellner AWA, Jiang SX, Cheng X, Meng X and Rodrigues T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. An Acad Bras Cienc 82: 1045-1062.) and one not fully prepared lower jaw (Veldmeijer 2002Suberbiola XP, Bardet N, Jouve S, Iarochène M, Bouya B and Amaghzaz M. 2003. A new azhdarchid pterosaur from the Late Cretaceous phosphates of Morocco. Geol Soc London, Spec Pub 217: 79-90.). MN 6594-V presents for the first time information on the postcranial skeleton of Tropeognathus. With an estimated wingspan exceeding 8 m, this specimen is the largest pterosaur that can be identified at the species level recovered from any Gondwanan deposit so far.
We also briefly describe an isolated distal part of a large anhanguerid humerus (MCT 1838-R) from the State of Pernambuco and another large individual (MPSC R 1395) presently regarded as Anhangueridae indet., which was recently collected in a controlled excavation made at the northwestern portion of the Araripe Basin in the State of Ceará.
Anatomical abbreviations: ac - acetabulum, afc - articular facet for coracoid, afs - articular facet for scapula, bs - basisphenoid, car - carpals, cor - coracoid, cs - cristospine, cv - cervical vertebrae, dpc - deltopectoral crest of the humerus, dv - dorsal vertebrae, efs - external fundamental system, fe - femur, fopn - foramen pneumaticum, glfo - glenoid fossa, hu - humerus, il- ilium, j - jugal, j.rid - jugal ridge, l - left, LAGs - lines of arrested growth, ltf - lower temporal fenestra, m - maxilla, maxws - maximized wingspan, MC - medullar cavity, mcIV - metacarpal IV, naof - nasoantorbital fenestra, not - notarium, ns - neural spine, nws - normal wingspan, or - orbit, pcr - parietal crest, pel - pelvis, ph1d4 - first phalanx of manual digit IV, ph2d4 - second phalanx of manual digit IV, ph3d4 - third phalanx of manual digit IV, ph4d4 - fourth phalanx of manual digit IV, pmcr - premaxillary crest, po - postorbital, prcor - processus coracoidalis, prid - palatal ridge, prsca - processus scapularis, q - quadrate, qfl - quadrate flange, r - right, rfc - reducing factor, RZ - resorptive zone, sca - scapula, scp - supracondylar process of the humerus, stk - sternum keel, stp - sternal plate, sv - sacral vertebrae, te - tooth, ti - tibia, tpr - transverse process, tub - tubercle, ul - ulna, utf - upper temporal fenestra.
Institutional abbreviations: AMNH - American Museum of Natural History, New York, USA, MCT - Museu de Ciências da Terra, DNPM, Rio de Janeiro, RJ, Brazil, MN - Museu Nacional/UFRJ, Rio de Janeiro, RJ, Brazil, MPSC - Museu de Paleontologia de Santana do Cariri, Santana do Cariri, CE, Brazil, NSM - Division of Vertebrate Paleontology, National Science Museum, Tokyo, Japan.
GEOLOGICAL SETTING
The stratigraphy of the Araripe Basin is controversial, particularly concerning what is commonly known as the Santana Formation (e.g., Maisey 1991Maisey J. (Ed). 1991. Santana fossils: an illustrated Atlas. Neptune, New Jersey: T.F.H. Publications, 459 p., Martill 2007Martill DM. 2007. The age of the Cretaceous Santana Formation fossil Konservat Lagerstätte of north-east Brazil: a historical review and an appraisal on the biochronostratigraphic utility of its paleobiota. Cretaceous Res 28: 895-920., Assine 2007Assine ML. 2007. Bacia do Araripe. Boletim de Geociências da Petrobras 15: 371-389.). Although the full revision of the nomenclatural problems regarding the Santana deposits is beyond the scope of this paper, some comments are necessary to avoid the spread of erroneous notions that can produce nomenclatural instability.
As pointed out before (e.g., Beurlen 1971Beurlen K. 1971. As condições ecológicas e faciológicas da Formação Santana na Chapada do Araripe (Nordeste do Brasil). An Acad Bras Cienc 43: 411-415.), the first author who studied the stratigraphic sequence of the Araripe Basin was Small (1913)Silva HP and Kellner AWA. 2006. A combinação de técnicas na preparação de vertebrados fósseis: o exemplo de Thalassodromeus sethi (Pterosauria, Tapejaridae). In: GALLO V ET AL. (Eds), Paleontologia de Vertebrados: Grandes Temas e Contribuições Científicas. Rio de Janeiro: Interciência, p. 193-301.. This author designated as “calcareo de Sant'Anna” the laminated limestone layers presently known as the Crato deposit (e.g., Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.). Later, Beurlen (1962)Beurlen K. 1962. Geologia da Chapada do Araripe. An Acad Bras Cienc 34: 365-370. formally established the Santana Formation, including not only the laminated calcareous layers of Small (1913)Silva HP and Kellner AWA. 2006. A combinação de técnicas na preparação de vertebrados fósseis: o exemplo de Thalassodromeus sethi (Pterosauria, Tapejaridae). In: GALLO V ET AL. (Eds), Paleontologia de Vertebrados: Grandes Temas e Contribuições Científicas. Rio de Janeiro: Interciência, p. 193-301., but also the overlying evaporitic and siliciclastic deposits, the latter incorporating the levels of extremely fossiliferous calcareous nodules embedded in laminated shales (e.g., Fara et al. 2005Fara E, Saraiva AAF, Campos DA, Moreira JKR, Siebra DC and Kellner AWA. 2005. Controlled excavations in the Romualdo Member of the Santana Formation (Early Cretaceous, Araripe Basin, northeastern Brazil): stratigraphic, palaeoenvironmental and palaeoecological implications. Palaeogeogr Palaeoclimatol Palaeoecol 218: 145-160., Vila Nova et al. 2011).
In the following year, Beurlen (1963)Beurlen K. 1963. Geologia e Estratigrafia da Chapada do Araripe. 17 Congresso Brasileiro de Geologia Publ. SUDENTE: 1-47. separated the laminated calcareous deposits of Small (1913)Silva HP and Kellner AWA. 2006. A combinação de técnicas na preparação de vertebrados fósseis: o exemplo de Thalassodromeus sethi (Pterosauria, Tapejaridae). In: GALLO V ET AL. (Eds), Paleontologia de Vertebrados: Grandes Temas e Contribuições Científicas. Rio de Janeiro: Interciência, p. 193-301. in a new stratigraphic unit, the Crato Formation. He further restricted the Santana Formation to the evaporites and to the overlying siliciclastic deposits that he named Ipubi and Romualdo members, respectively. However, this contradicts the original designation of Small (1913)Silva HP and Kellner AWA. 2006. A combinação de técnicas na preparação de vertebrados fósseis: o exemplo de Thalassodromeus sethi (Pterosauria, Tapejaridae). In: GALLO V ET AL. (Eds), Paleontologia de Vertebrados: Grandes Temas e Contribuições Científicas. Rio de Janeiro: Interciência, p. 193-301..
Some years later, in a very influential paper, Beurlen (1971)Beurlen K. 1971. As condições ecológicas e faciológicas da Formação Santana na Chapada do Araripe (Nordeste do Brasil). An Acad Bras Cienc 43: 411-415. revised the nomenclature of these deposits. The Santana Formation was maintained as he had introduced earlier (Beurlen 1962Beurlen K. 1962. Geologia da Chapada do Araripe. An Acad Bras Cienc 34: 365-370.), which had been followed by most authors at that time (e.g., Santos and Valença 1968Rodrigues T, Kellner AWA, Mader BJ and Russell D. 2011. New pterosaur specimens from the Kem Kem beds (Upper Cretaceous, Cenomanian) of Morocco. Riv Ital Paleontol S 117: 149-160.). He further subdivided the Santana Formation into three members, named from base to top Crato, Ipubi and Romualdo (Beurlen 1971Beurlen K. 1971. As condições ecológicas e faciológicas da Formação Santana na Chapada do Araripe (Nordeste do Brasil). An Acad Bras Cienc 43: 411-415.).
In a detailed stratigraphic review of the Santana deposits, Neumann and Cabrera (1999)Martill DM and Unwin DM. 2012. The world's largest toothed pterosaur, NHMUK R481, an incomplete rostrum of Coloborhynchus capito (Seeley, 1870) from the Cambridge Greensand of England. Cretaceous Res 34: 1-9. elevated the Santana Formation to the status of group, and the Crato, Ipubi and Romualdo members were then considered formations. Martill (2007)Martill DM. 2007. The age of the Cretaceous Santana Formation fossil Konservat Lagerstätte of north-east Brazil: a historical review and an appraisal on the biochronostratigraphic utility of its paleobiota. Cretaceous Res 28: 895-920. basically accepted Neumann and Cabrera's proposition, but challenged the name Romualdo, using Santana Formation instead. However, the International Stratigraphic Guide states that “if a unit is divided into two or more formal component units, the geographic name of the original unit should not be employed for any of the subdivisions” (International Subcommission on Stratigraphic Classification 1999International Subcommission on Stratigraphic Classification. 1999. International Stratigraphic Guide - An abridged edition. In: MURPHY MA AND SALVADOR A (Eds), Episodes 22: 255-271.: 258, Names for subdivisions of Stratigraphic Units). The Brazilian Code of Stratigraphic Nomenclature also has a similar guidance, stating that when a unit is divided and elevated to a more inclusive category, the geographic name is to be used for this unit, instead of restricitng its name to one of its subdivisions (Comissão Especial de Nomenclatura Estratigráfica - SBG 1986: 374). Therefore, we use the stratigraphic scheme adopted by Neumann and Cabrera (1999)Martill DM and Unwin DM. 2012. The world's largest toothed pterosaur, NHMUK R481, an incomplete rostrum of Coloborhynchus capito (Seeley, 1870) from the Cambridge Greensand of England. Cretaceous Res 34: 1-9. instead of a procedure that, in our opinion, introduces nomenclatural instability.
The main pterosaur material described here (MN 6594-V) was collected about three decades ago and the exact locality from where it comes is unknown. It was found by local inhabitants and broken into several pieces that ended up donated to the Museu Nacional/UFRJ. The bones are preserved in parts of what appears to have been one large calcareous nodule, typical of the Romualdo Formation, and can be confidently assigned to this stratigraphic unit. Some bones are partially eroded, indicating that parts of MN 6594-V were exposed for some time prior to collection.
The exact locality where the distal end of the large humerus MCT 1838-R, also described in this paper, was unearthed is unknown too, but based on the available information, it comes from an outcrop of the Romualdo Formation in the State of Pernambuco.
The last specimen (MPSC R 1395) addressed in the present study was collected in the locality known as Sítio Baixa Grande, within the municipality Araripe, at the northwestern portion of the Araripe basin in the State of Ceará. It was recovered in one of the few controlled excavations done in the Araripe Basin and belongs to a partial wing of a large pterosaur that was found along with hundreds of other fossils, mostly fishes, coprolites, mollusks and plant material.
SYSTEMATIC PALEONTOLOGY
Pterosauria Kaup, 1834
Pterodactyloidea Plieninger, 1901
Pteranodontoidea Marsh, 1876
Anhangueridae Campos & Kellner, 1985Campos DA and Kellner AWA. 1985. Panorama of the Flying Reptiles Study in Brazil and South America. An Acad Bras Cienc 57: 453-466.
Tropeognathus cf. T. mesembrinus Wellnhofer, 1987Wang XL, Kellner AWA, Jiang SX, Cheng X, Meng X and Rodrigues T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. An Acad Bras Cienc 82: 1045-1062.
Material: partial skeleton consisting of an incomplete skull (including the braincase) and lower jaw, five cervical vertebrae (including part of the axis), twelve dorsal vertebrae (five fused into a notarium), sacrum, incomplete scapula and coracoid from both sides, sternum, pelvic region and wing elements from both sides, housed in the Paleovertebrate Sector of the Department of Geology and Paleontology, Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro (MN 6594-V, Figs. 1-8).
Skull elements of Tropeognathus cf. T. mesembrinus (MN 6594-V). (a) Incomplete rostrum in right lateral view. (b) Incomplete rostrum in anterior view. (c) Posterior lower portion in right lateral view. (d) Posterior lower portion in posterior view. (e) Braincase in left lateral view. (f) Mandible in dorsal view. (g) Fragment of lower jaw in lateral view. (h) Fragment of lower jaw in medial view. Arrows indicate alveoli and teeth. See text for abbreviations. Scale bar 50 mm.
Reconstruction of the skull of Tropeognathus cf. T. mesembrinus (MN 6594-V) with the preserved parts indicated in grey.
Incomplete rostrum of Tropeognathus cf. T. mesembrinus (MN 6594-V) in left lateral view. Arrows indicate channels, regarded as impressions of blood vessels. Scale bar 50 mm.
Incomplete rostrum of Tropeognathus cf. T. mesembrinus (MN 6594-V) in right lateral view. Arrows indicate channels, regarded as impressions of blood vessels. Scale bar 50 mm.
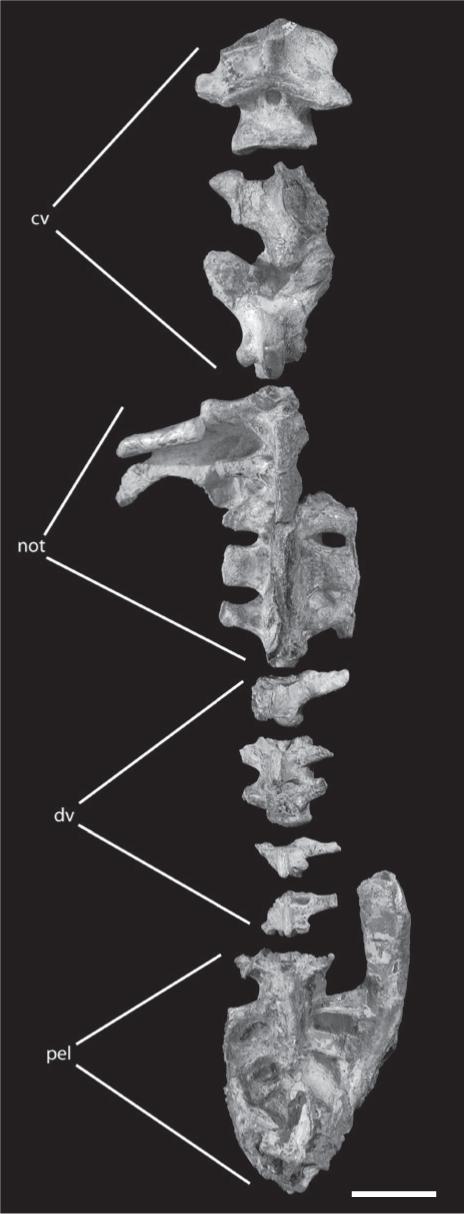
Vertebral column of Tropeognathus cf. T. mesembrinus (MN 6594-V) in dorsal view. See text for abbreviations. Scale bar 50 mm.
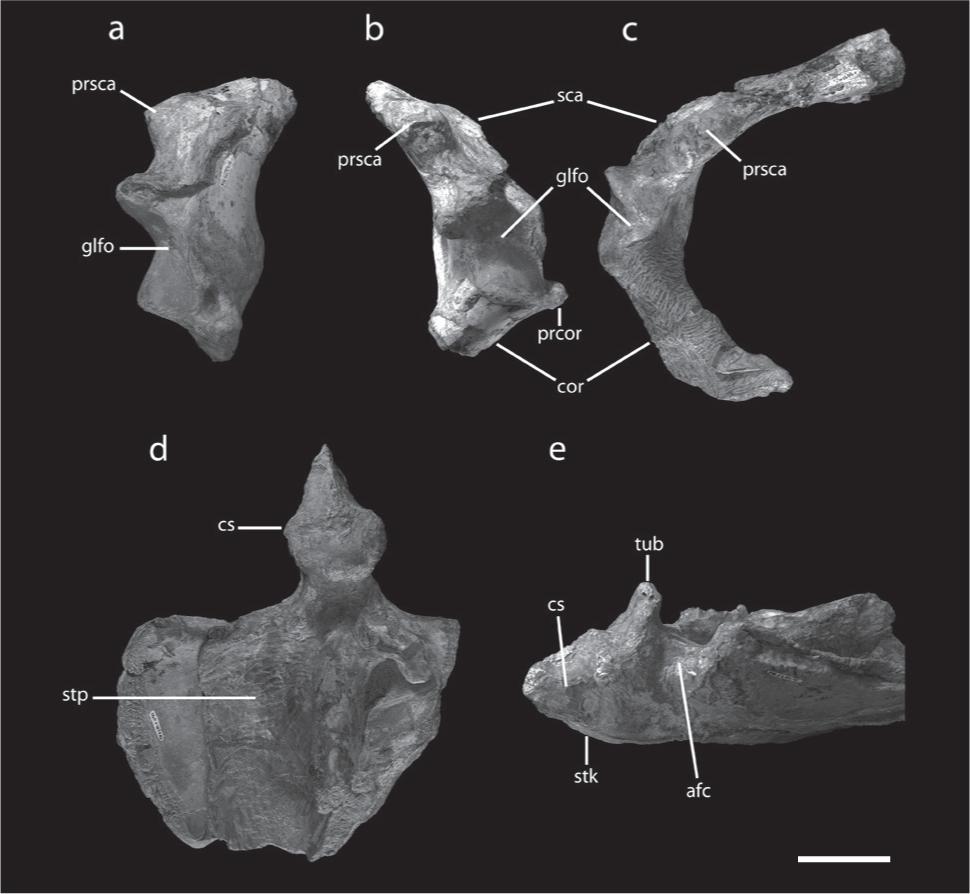
Postcranial elements of Tropeognathus cf. T. mesembrinus (MN 6594-V). (a) Right scapulocoracoid in anterior view. (b) Right scapulocoracoid in posterior view. (c) Left scapulocoracoid in posterior view. (d) Sternum in ventral view. (e) Anterior portion of the sternum in left lateral view. See text for abbreviations. Scale bar 50 mm.
Postcranial elements of Tropeognathus cf. T. mesembrinus (MN 6594-V). (a) Proximal portion of the right humerus in medial view. (b) Left humerus in medial view. (c) Distal portion of the right humerus in ventral view. (d) Notarium in left lateral view. See text for abbreviations. Scale bar 50 mm.
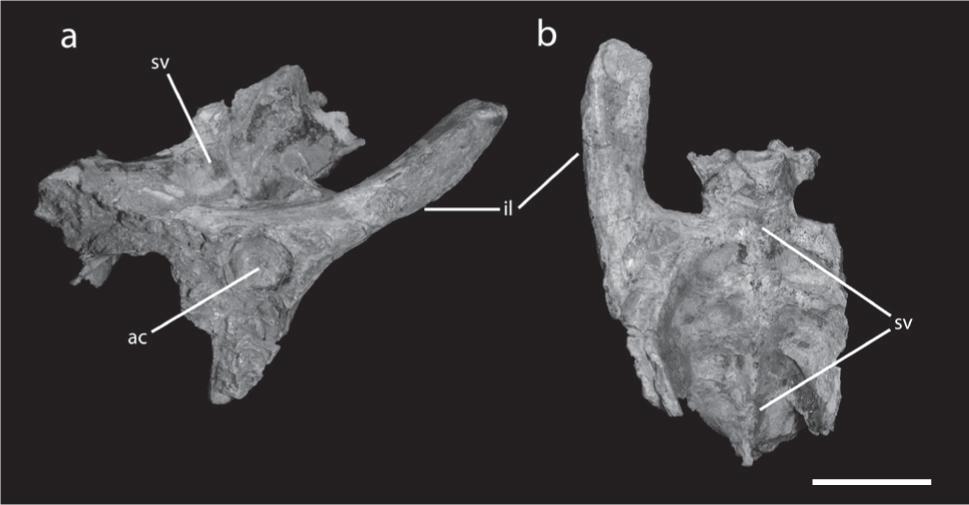
Pelvis of Tropeognathus cf. T. mesembrinus (MN 6594-V). (a) Right lateral view. (b) Dorsal view. See text for abbreviations. Scale bar 50 mm.
Locality and horizon: Romualdo Formation, Early Cretaceous (Aptian-Albian), Araripe Basin, Northeastern Brazil. Although the exact locality of collection is unknown, it probably came from the outcrops around Santana do Cariri, Ceará State.
Anhangueridae indet. 1
Material: distal part of a left humerus housed in the Museu de Ciências da Terra, Departamento Nacional de Produção Mineral/Companhia de Pesquisa de Recursos Minerais, Rio de Janeiro (MCT 1838-R, Fig. 9).
MCT 1838-R right humerus. (a) Ventral view. (b) Transverse section, showing the bony trabeculae covered mainly by calcite. (c) Distal view. Arrows indicate the cortical bone. See text for abbreviations. Scale bar 50 mm.
Locality and horizon: Romualdo Formation, Early Cretaceous (Aptian-Albian), Araripe Basin, Northeastern Brazil. This specimen was collected in the State of Pernambuco, close to the border with Ceará.
Anhangueridae indet. 2
Material: partial left wing, including humerus, wing metacarpal and manual digits with the complete wing finger, housed in the Museu de Paleontologia de Santana do Cariri (Universidade Regional do Cariri/URCA), Ceará (MPSC R 1395, Fig. 10).
Locality and horizon: Romualdo Formation, Early Cretaceous (Aptian-Albian), Araripe Basin, Northeastern Brazil. Locality known as Sítio Baixa Grande, Araripe Municipality, State of Ceará.
DESCRIPTION AND COMPARISON
MN 6594-V. The most complete specimen described here is divided into several parts ( Figs. 1-2). Most bones were isolated using a combination of chemical and mechanic preparation techniques that was already applied with success in other pterosaurs from the Romualdo Formation before (Silva and Kellner 2006Schweigert G. 2007. Ammonite biostratigraphy as a tool for dating Upper Jurassic lithographic limestones from South Germany - first results and open questions. N Jb Geol Paläont Abh 245:117-125.). Several elements show clear signs of erosion (e.g., braincase, both humeri, pelvis, part of the notarium) indicating that this nodule was at least partially broken due to weathering and long time exposed before being collected. It is also possible that some broken parts ended up separated from the main material and might come to light in the future.
From the taphonomic point of view, although most elements are three-dimensionally preserved, some are distorted and show different degrees of compression. The most remarkable feature regarding preservation, however, is the extensive presence of calcite veins cutting and deforming the bones in several parts. The skull, for example, had some of its areas displaced from their original position at the exact point of the fracture line of a calcite vein (e.g., Fig. 1a).
The skull is incomplete and divided into three parts: the incomplete rostrum (lacking the anterior end) until the anterior margin of the nasoantorbital fenestra, the posterior lower part formed essentially by the jugal and quadrate (including the basisphenoid) and the braincase. All elements are fused and no clear sutures can be distinguished.
None of the cranial openings are complete. Based on the dorsal and ventral margins of the right orbit, this opening is inferred to have been elongated as in most pteranodontoids such as Anhanguera, Tropeognathus mesembrinus, Pteranodon and Dawndraco (Campos and Kellner 1985Campos DA and Kellner AWA. 1985. Panorama of the Flying Reptiles Study in Brazil and South America. An Acad Bras Cienc 57: 453-466., Wellnhofer 1985Wang X, Kellner AWA, Jiang S and Cheng X. 2012. New toothed flying reptile from Asia: close similarities between early Cretaceous pterosaur faunas from China and Brazil. Naturwissenschaften 99: 249-257, 1987Wang XL, Kellner AWA, Jiang SX, Cheng X, Meng X and Rodrigues T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. An Acad Bras Cienc 82: 1045-1062., Bennett 2001aBennett SC. 2001a. The osteology and functional morphology of the Late Cretaceous Pterosaur Pteranodon Part 1 - General description and osteology. Palaeont Abt A 260: 1-112., Kellner 2010Kellner AWA. 2010. Comments on the Pteranodontidae (Pterosauria, Pterodactyloidea) with the description of two new species. An Acad Bras Cienc 82: 1063-1084.) and not rounded as in Ludodactylus and Guidraco (Frey et al. 2003aFrey E, Martill DM and Buchy MC. 2003a. A new crested ornithocheirid from the Lower Cretaceous of northeastern Brazil and the unusual death of an unusual pterosaur. Geol Soc London, Spec Pub 217: 55-63., Wang et al. 2012Vila Nova BC and Sayão JM. 2012. On wing disparity and morphological variation of the Santana Group pterosaurs. Hist Biol 24: 567-574.). The upper temporal fenestra is rounded and opens dorsally. The ventral margin of the lower temporal fenestra indicates that this opening is rather large and piriform, not compressed and elongated as in Pteranodon and Dawndraco (Bennett 2001aBennett SC. 2001a. The osteology and functional morphology of the Late Cretaceous Pterosaur Pteranodon Part 1 - General description and osteology. Palaeont Abt A 260: 1-112., Kellner 2010Kellner AWA. 2010. Comments on the Pteranodontidae (Pterosauria, Pterodactyloidea) with the description of two new species. An Acad Bras Cienc 82: 1063-1084.). Only the anterior margin and part of the lower posterior margin of the nasoantorbital fenestra are preserved. At the distal end of the nasoantorbital fenestra, the portion dorsal to the fenestra, formed by the premaxilla, is much thinner than the part ventral to it, formed by the maxilla. The holotype of Tropeognathus mesembrinus (cast MN 4736-V) also shows this condition, but this difference is more pronounced in MN 6594-V. In other anhanguerids (such as Anhanguera and Liaoningopterus), the anteriormost point of the nasoantorbital fenestra is approximately in the middle of the rostrum's height.
The main feature of the premaxilla is the presence of a very large and high sagittal crest ( Figs. 3-4). This crest does not reach the anterior margin of the nasoantorbital fenestra and has the posterior margin angled at about 60° in respect to the ventral border of the skull. This is steeper than in any other anhanguerid, including the holotype of Tropeognathus mesembrinus (36°).
Among the interesting features of this crest is the presence of several shallow channels ( Figs. 3-4). On the right lateral side there is one central channel that runs sub parallel to the ventral border of the skull. Right after the anterior margin of the nasoantorbital fenestra one slightly smaller channel branches off following the posterior margin of the premaxillary crest. More anteriorly, several smaller channels branch off and run towards the dorsal margin of the sagittal crest in an irregular pattern. These channels are interpreted as the impressions of blood vessels and resemble a similar structure first described in the tapejarid Thalassodromeus (Kellner and Campos 2002Kellner AWA and Campos DA. 2002. The function of the cranial crest and jaws of a unique pterosaur from the Early Cretaceous of Brazil. Science 297: 389-392.). It likely also had the same function, serving as a cooling device for this pterosaur. On the dorsal part of the premaxillary crest there are several sub parallel channels.
The maxilla forms the lower margin of the nasoantorbital fenestra. The portion ventral to this margin is very developed, more than in any other pterodactyloid, being about two times broader than the corresponding part of the premaxilla dorsal to the upper margin of the nasoantorbital fenestra. This is quite unusual and so far unique of MN 6594-V. The number of alveoli is rather reduced, indicating that this specimen had fewer teeth than Anhanguera, being more similar to Tropeognathus mesembrinus.
The frontal and parietal are partially preserved. They bear a low and blunt frontoparietal crest that is regarded to be a potential anhanguerid synapomorphy (Kellner 2003Kellner AWA. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. Geol Soc London, Spec Pub 217: 105-137.). This crest is proportionally larger and broader than in Anhanguera and similar to Tropeognathus, albeit more developed. This crest extends slightly posteriorly as in Tropeognathus, differing from other anhanguerids. A marked lateral ridge can be found on the lateral surface of the parietal, dorsal to the upper temporal fenestra.
The postorbital is small and triangular (Fig. 1e).
The jugal is incomplete in both sides, with the left one being displaced (along with the quadrate) inside the skull (Fig. 1c). As typical for pterodactyloids, it is tetraradiate, forming the posteroventral margin of the nasoantorbital fenestra and the anteroventral margin of the orbit. It bears a well-developed jugal ridge. On the right side there is a splint-like bone covering the maxilla that is likely the maxillary process of the jugal (Fig. 1a). This indicates that this bone extends slightly anteriorly regarding the nasoantorbital fenestra.
The quadrate is a strong and elongated bone similar to other anhanguerids (e.g., Wellnhofer 1985Wang X, Kellner AWA, Jiang S and Cheng X. 2012. New toothed flying reptile from Asia: close similarities between early Cretaceous pterosaur faunas from China and Brazil. Naturwissenschaften 99: 249-257). It is inclined at about 120°, a typical pterodactyloid feature (Fig. 1c). The cranioquadrate articulation is wide and shows the typical helical jaw joint that has been observed in several derived pterodactyloid taxa (e.g., Eaton 1910Eaton GF. 1910. Osteology of Pteranodon. Memoirs of the Connecticut Academy of Arts and Sciences 1: 1-38., Kellner and Langston 1996Kellner AWA and Langston WJR. 1996. Cranial remains of Quetzalcoatlus (Pterosauria, Azhdarchidae) from the Late Cretaceous Sediments of Big Bend National Park, Texas. J Vert Paleont 16: 222-231.), including anhanguerids (e.g., Campos and Kellner 1985Campos DA and Kellner AWA. 1985. Panorama of the Flying Reptiles Study in Brazil and South America. An Acad Bras Cienc 57: 453-466., Wellnhofer 1991aWellnhofer P. 1985. Neue Pterosaurier aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 187: 105-182.). The quadrate flange is present as reported in several pteranodontoids (e.g., Wang et al. 2008Vila Nova BC, Saraiva AAF, Moreira JKR and Sayão JM. 2011. Controlled excavations in the Romualdo Formation lagerstätte (Araripe Basin, Brazil) and pterosaur diversity: remarks based on new findings. Palaios 26: 173-179., Bennett 2001aBennett SC. 2001a. The osteology and functional morphology of the Late Cretaceous Pterosaur Pteranodon Part 1 - General description and osteology. Palaeont Abt A 260: 1-112., Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.), but not very well-developed (Fig. 1d).
The palatal region is compressed and very few bones are observed. The choanae are separated by a comparatively strong vomer. The palatine is separated from the maxillae by a lateral sulcus as has been reported several times in anhanguerids (e.g., Campos and Kellner 1985Campos DA and Kellner AWA. 1985. Panorama of the Flying Reptiles Study in Brazil and South America. An Acad Bras Cienc 57: 453-466., Wellnhofer 1985Wang X, Kellner AWA, Jiang S and Cheng X. 2012. New toothed flying reptile from Asia: close similarities between early Cretaceous pterosaur faunas from China and Brazil. Naturwissenschaften 99: 249-257, 1987Wang XL, Kellner AWA, Jiang SX, Cheng X, Meng X and Rodrigues T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. An Acad Bras Cienc 82: 1045-1062.).
Very little from the occipital region is observable. This part of the skull was either broken off during the collecting process or exposed to recent weathering, showing the spongy internal bone structure. The posttemporal fenestra is small and can be observed on the left side. The dorsal portion of the occiput is more inclined and concave than in Anhanguera (e.g., Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.) and similar to Tropeognathus mesembrinus (Wellnhofer 1987Wang XL, Kellner AWA, Jiang SX, Cheng X, Meng X and Rodrigues T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. An Acad Bras Cienc 82: 1045-1062.). The basisphenoid is a broad bone (Fig. 1d), similar to Tropeognathus (Wellnhofer 1987Wang XL, Kellner AWA, Jiang SX, Cheng X, Meng X and Rodrigues T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. An Acad Bras Cienc 82: 1045-1062.) and contrasting with the one of Pteranodon (e.g., Bennett 2001aBennett SC. 2001a. The osteology and functional morphology of the Late Cretaceous Pterosaur Pteranodon Part 1 - General description and osteology. Palaeont Abt A 260: 1-112.).
The lower jaw is incomplete and shows the posterior end of the mandibular symphysis and part of both mandibular rami, with the right one more complete than the left one (Fig. 1f). The ventral part is not preserved and was likely broken off during the collecting process. Only the dentary and part of the splenial are preserved. Four alveoli are present on the right and three on the left mandibular ramus, respectively, some bearing teeth. Compared to other lower jaws of anhanguerid pterosaurs, the number of teeth in MN 6594-V in the preserved portions is the same as in Tropeognathus and less than in Anhanguera (Wellnhofer 1985Wang X, Kellner AWA, Jiang S and Cheng X. 2012. New toothed flying reptile from Asia: close similarities between early Cretaceous pterosaur faunas from China and Brazil. Naturwissenschaften 99: 249-257, 1987Wang XL, Kellner AWA, Jiang SX, Cheng X, Meng X and Rodrigues T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. An Acad Bras Cienc 82: 1045-1062., Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.). Part of a small and rounded Meckelian fossa is observed. All preserved teeth are comparatively small and vertically oriented.
A fragmentary portion of a jaw with remains of two alveoli was also found and is interpreted as part of the mandibular symphysis. The preserved teeth tend to be very small. The deepness of this element suggests the presence of a dentary sagittal crest (Figs. 1g, h).
The postcranial elements of MN 6594-V represent the main parts of the skeleton, including cervical, dorsal and sacral vertebrae, scapula and coracoid, sternum, all main parts of the wing, partial pelvis and one femur ( Figs. 5-8). The preservation varies from complete and lacking distortions (e.g., some vertebrae) to partial, compacted and broken (e.g., middle portion of radius and ulna). All postcranial elements are very similar to the ones of anhanguerids (e.g., Wellnhofer 1991aWellnhofer P. 1985. Neue Pterosaurier aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 187: 105-182., Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135., Veldmeijer 2003Valença LMM, Neumann VH and Mabesoone JM. 2003. An overview on Callovian-Cenomanian intracratonic basins of Northeast Brazil: onshore stratrigraphic record of the opening of the southern Atlantic. Geol Acta 1: 261-275.).
The axis is incomplete, lacking the anterior part. It bears a tall neural spine and blunt postzygapophyses that are stouter than in Anhanguera piscator. At the contact between neural arch and centrum there is a well-developed lateral pneumatic foramen. The remaining cervical vertebrae belong to the posterior part of the neck (Fig. 5). All are well preserved and show a spike-like neural spine. They show a lateral pneumatic foramen as observed in Anhanguera santanae and Anhanguera piscator (Wellnhofer 1991aWellnhofer P. 1985. Neue Pterosaurier aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 187: 105-182., Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.). There is no additional foramen or opening around the neural canal. The last cervical is partially dorsalized and is interpreted as the 8th. All bear a well-developed hypapophysis. The main differences between the cervical vertebrae of MN 6594-V and Anhanguera's are the stronger zygapophyses in the former.
There are 12 dorsal vertebrae, the first five fused into a notarium (Figs. 5, 7d). The fourth dorsal element is the longest one and bears the articulation surface for the scapula. This vertebra also shows stronger transverse processes and a shallow depression at the base of the neural spine. The centra of the first five dorsal vertebrae are fused, although the neural spine of some was not completely fused to each other yet. There is one rib connected with the first dorsal vertebra, but not fused to it. The notarium of MN 6594-V differs from that observed in Anhanguera spielbergi, where this structure, formed by six elements, is composed by the ninth cervical and the first five dorsal vertebrae (Veldmeijer 2003Valença LMM, Neumann VH and Mabesoone JM. 2003. An overview on Callovian-Cenomanian intracratonic basins of Northeast Brazil: onshore stratrigraphic record of the opening of the southern Atlantic. Geol Acta 1: 261-275.). Although possible, it is not clear if these differences are ontogenetic.
There are five sacral vertebrae, all fused to each other and to the ilium, forming a synsacrum (Fig. 5). Although unfused, Anhanguera piscator also has five sacral vertebrae (Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.) while Anhanguera spielbergi bears six (Veldmeijer 2003Valença LMM, Neumann VH and Mabesoone JM. 2003. An overview on Callovian-Cenomanian intracratonic basins of Northeast Brazil: onshore stratrigraphic record of the opening of the southern Atlantic. Geol Acta 1: 261-275.), the last one being an incorporated caudal. Sacral vertebrae three and four in MN 6594-V bear a keeled ventral surface, an unusual feature never before recorded in Pterosauria.
The sternum is comparatively large and shows a blunt, short and deep cristospine (Figs. 6 d, e), similar to the one of Anhanguera piscator (Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.). The sternocoracoid articulations are symmetrical.
Both scapulae and coracoids are preserved and fused into scapulocoracoids (Figs. 6 a, b). The left scapula is almost complete, with an estimated length of 195 mm. Besides being stronger, the scapula is similar to that of Anhanguera (Wellnhofer 1991aWellnhofer P. 1985. Neue Pterosaurier aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 187: 105-182., Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.).
Regarding the wing bones, both humeri are incomplete, with the right one having the proximal and distal parts preserved (Figs. 7a-c). It shows a developed supracondylar process and a warped deltopectoral crest. The radius and ulna are known from both sides, particularly their proximal and distal articulations. Unfortunately the shaft is not complete, but the preserved parts show that the diameter of the radius is less than half of that of the ulna, as has been observed in other anhanguerids (e.g., Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135., Veldmeijer 2003Valença LMM, Neumann VH and Mabesoone JM. 2003. An overview on Callovian-Cenomanian intracratonic basins of Northeast Brazil: onshore stratrigraphic record of the opening of the southern Atlantic. Geol Acta 1: 261-275.). The carpus is incomplete, but the proximal and distal carpal series are fused. Only the proximal articulation of the wing metacarpal is observed. Metacarpal I articulates with the carpus and metacarpals II and III are apparently reduced. The wing finger is incomplete and its main preserved part is the distal articulation of what we interpret as being the first wing finger phalanx. A histological section of this bone was made (see below).
The pelvic girdle from the right side is preserved (Fig. 8). The ilium is elongated, with the preacetabular part longer than the postacetabular portion. The preacetabular part is strongly deflected dorsally, similar to Anhanguera santanae and Anhanguera spielbergi (Wellnhofer 1991aWellnhofer P. 1985. Neue Pterosaurier aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 187: 105-182., Veldmeijer 2003Valença LMM, Neumann VH and Mabesoone JM. 2003. An overview on Callovian-Cenomanian intracratonic basins of Northeast Brazil: onshore stratrigraphic record of the opening of the southern Atlantic. Geol Acta 1: 261-275.). Both pubis and ischium are very incomplete in this specimen.
The only preserved bone of the hindlimb is the femur, which is long and slender. Its rounded head has a constricted neck and is set apart from the shaft. As observed in other anhanguerids (e.g., Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.), the femur is not very bowed.
MCT 1838-R. The second specimen studied here is formed by an isolated distal part of a right humerus that was prepared using chemical methods (Silva and Kellner 2006Schweigert G. 2007. Ammonite biostratigraphy as a tool for dating Upper Jurassic lithographic limestones from South Germany - first results and open questions. N Jb Geol Paläont Abh 245:117-125.). It is quite large, with the distal end about 127 mm wide (Fig. 9). The distal articulation surface is triangular, a trait found in pteranodontoids (Fig. 9c). The supracondylar process is ridge-like and positioned close to the distal articulation. There are two pneumatic foramina, one on the anterior surface and another on the distal articulation.
Despite its size, the cortical bone is very thin, less than 2 mm wide in most parts. The bone is very hollow, reinforced internally with well-developed trabeculae (Fig. 9b). In this specimen, most trabeculae are covered with a brownish layer that is composed of diagenetic carbonates, mainly calcite.
MPSC R 1395. Another specimen only briefly described here is an almost complete left wing formed by the humerus, wing metacarpal and the complete wing finger. This material is still being prepared by one of us (JSF) using mechanical methods (Silva and Kellner 2006Schweigert G. 2007. Ammonite biostratigraphy as a tool for dating Upper Jurassic lithographic limestones from South Germany - first results and open questions. N Jb Geol Paläont Abh 245:117-125.) and so far the humerus, metacarpal IV and the first wing phalanx are partially freed from the calcareous matrix (Fig. 10). The specimen is extremely well preserved, showing no detectable distortion.
The humerus lacks the middle part of the shaft. It has a well-developed warped deltopectoral crest that shows the same morphology of other anhanguerids from the Romualdo Formation (e.g., Anhanguera piscator). Based on the proportions of other anhanguerids, in which the humerus has almost the same length of the wing metacarpal IV (e.g., Vila Nova and Sayão 2012), it is estimated to be 262 mm long.
The proximal articulation of the wing metacarpal IV shows a developed process that fits into a fovea on the distal carpal series (not preserved in this specimen). The distal articulation shows the typical rounded surface to receive the first wing phalanx.
The first wing phalanx is a large element, a typical feature of most pteranodontoids. Its general morphology agrees well with the corresponding elements referred to Anhangueridae. The extensor tendon process is not fused, suggesting that this specimen was a young individual despite its large size.
BONE HISTOLOGY OF MN 6594-V
We have also examined one histological section of the distal part of a wing phalanx of MN 6594-V, regarded to be the first phalanx of the wing finger. Despite the three-dimensional preservation of this bone, histological details in some parts of the thin section are not clearly discernible because of the extensive calcite permineralization. Where the structures are better preserved, the outermost periosteal layer frequently constitutes the external fundamental system (efs) of pseudolamellar tissue with coarsely parallel-fibers. Predominantly circumferentially oriented vascular channels compose the cortex (Fig. 11). In a small amount they are arranged in lines with only some isolated occurrences of secondary osteons. MN 6594-V presents a well-developed layer of inner circumferential lamellar bone that lines the medullary cavity. It bears three reabsorption zones filled by well-developed calcite crystals.
Bone microstructure of Tropeognathus cf. T. mesembrinus. (A) Transverse section of the first wing phalanx showing a resorptive cavity in the medullary surface. Scale bar 50 µ. (B) Detail of the wing phalanx section, arrows indicate periodical interruption in growth. See text for abbreviations. Scale bar 100 µ.
Three cycles of zones and annuli are observed, resulting in the formation of lines of arrested growth (LAGs). As pointed out before in the literature, when increasing ontogeny with the formation of LAGs, the bone deposited in the zones changes to a parallel-fibered bone tissue with fewer channels for vascularization, suggesting an overall slower rate of bone formation when compared with an early growth stage (Sayão 2003Santos RS and Valença JG. 1968. A Formação Santana e sua paleoictiofauna do Cretáceo do Brasil. An Acad Bras Cienc 40: 339-360., Chinsamy et al. 2008Chinsamy A, Codorniú L and Chiappe L. 2008. Developmental growth patterns of the filter-feeder pterosaur, Pterodaustro guinazui. Biol Lett 4: 282-285.). Later in ontogeny, the spacing of the LAGs tends to become visibly closer as a result of less bone being appositionally (Chinsamy et al. 2008Chinsamy A, Codorniú L and Chiappe L. 2008. Developmental growth patterns of the filter-feeder pterosaur, Pterodaustro guinazui. Biol Lett 4: 282-285.) as the pattern observed here. Based on the bone histology of Pterodaustro, Chinsamy et al. (2009)Chinsamy A, Codorniú L and Chiappe L. 2009. Palaeobiological Implications of the Bone Histology of Pterodaustro guinazui. Anat Rec 292: 1462-1477. established two phases of posthatching growth. The first one is an early phase of relative rapid growth cycles composed of zonal fibro-lamellar bone with annuli. This would be replaced by a later phase of relative slower growth, with cycles of parallel-fibred to lamellar bone tissue interrupted by LAGs (Chinsamy et al. 2009Chinsamy A, Codorniú L and Chiappe L. 2009. Palaeobiological Implications of the Bone Histology of Pterodaustro guinazui. Anat Rec 292: 1462-1477.). The close spacing of the growth lines in MN 6594-V, as well as its general histological features such as the presence of an external fundamental system, suggests that the overall growth had ceased and that it represents an adult individual (see Horner et al. 1999Horner RJ, Padian K and Ricqles A. 1999. Variation in dinosaur skeletochronology indicators: implications for age assessment and physiology. Paleobiology 25(3): 295-304. for a review). These features agree with the anatomical observation in which essentially all elements that tend to be unfused in ontogenetically immature individuals (e.g., Bennett 1993Bennett SC. 1993. The ontogeny of Pteranodon and other pterosaurs. Paleobiology 19: 92-106., Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.) are fused. Despite these observations, it is not totally clear if this individual could not have grown a little further, mainly because of the unfused neural spines of the notarium, as well as the ribs and the dorsal elements. It is possible that even if the appendicular elements completely cease to grow, the animal still tends to finish its fusion on the axial skeleton in order to allow greater stability during the flight of more mature individuals.
TAXONOMIC IDENTIFICATION
The specimen MN 6594-V can be referred to the Anhangueridae particularly due to the presence of a premaxillary sagittal crest confined to the anterior portion of the skull that does not reach the nasoantorbital fenestra. It also has a blunt frontoparietal crest, another potential anhanguerid trait (Kellner 2003Kellner AWA. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. Geol Soc London, Spec Pub 217: 105-137.). Unfortunately MN 6594-V lacks the rostral end and therefore it is unclear if it also has the distal end of the jaws expanded. However, the fragment of the jaw that is here interpreted as part of the mandibular symphysis suggests the presence of dentary sagittal crest, which is another anhanguerid feature.
Within the Anhangueridae, only two genera are present in the Romualdo Formation: Anhanguera, known by several species (e.g., Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135., Veldmeijer 2003Valença LMM, Neumann VH and Mabesoone JM. 2003. An overview on Callovian-Cenomanian intracratonic basins of Northeast Brazil: onshore stratrigraphic record of the opening of the southern Atlantic. Geol Acta 1: 261-275., Kellner 2006Kellner AWA. 2006. Pterossauros - os senhores do céu do Brasil. Rio de Janeiro: Vieira & Lent, 176 p.), and Tropeognathus, which is monospecific. Tropeognathus differs from Anhanguera mainly by having considerably fewer teeth (in both upper and lower jaws), a broader and blunter frontoparietal crest, a more developed palatal ridge, the dorsal portion of the occiput more inclined posteriorly and a shorter mandibular symphysis. Although the total amount of teeth in MN 6594-V cannot be established, the preserved parts clearly indicate that the number of teeth was less than in Anhanguera and the same as in Tropeognathus mesembrinus. Furthermore, although not complete, the frontoparietal crest of MN 6594-V is blunt and more massive than in Anhanguera and similar (albeit slightly more developed) to Tropeognathus mesembrinus. Other features that MN 6594-V shares with this taxon are the strong palatal ridge and the more inclined dorsal portion of the occipital region. Unfortunately the extension of the mandibular symphysis cannot be established in MN 6594-V, but based on the preserved portion it was shorter than in Anhanguera. The comparisons above indicate that MN 6594-V represents a taxon that shares more features with Tropeognathus than with Anhanguera.
However, there are some differences between MN 6594-V and the holotype of Tropeognathus mesembrinus. The main one is size, with the estimated length of the skull of MN 6594-V (based on the proportions of the postcranial elements, including the humerus) of approximately 979 mm, about 55% longer than the holotype of Tropeognathus mesembrinus (630 mm, see Wellnhofer 1987Wang XL, Kellner AWA, Jiang SX, Cheng X, Meng X and Rodrigues T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. An Acad Bras Cienc 82: 1045-1062.). Furthermore, the posterior margin of the premaxillary sagittal crest is much steeper than in Tropeognathus mesembrinus, being about 60° and 36° relative to the ventral margin of the skull, respectively. Another difference is observed in the bones that compose the dorsal and ventral portions of the nasoantorbital fenestra. In MN 6594-V, the lower bony bar, formed by the maxilla, is much larger than the dorsal one, formed by the premaxilla.
These anatomical differences lead to the question if MN 6594-V represents an individual of the species Tropeognathus mesembrinus or a new taxon. The main difficulty for a definitive answer is the limited information (if any) available about individual variation and the lack of a reliable notion concerning anatomical changes due to ontogeny in anhanguerids and in pterosaurs in general. In order to address these kinds of questions, there is a need of a large number of fairly complete and articulated specimens that could be reliably regarded as part of one population. To our knowledge, no deposit known so far fulfills this condition. This is true even for those Lagerstätten that have yielded a large number of specimens such as the Yixian (Barremian) and Jiufotang (Aptian) Formations in China, the Romualdo Formation (Aptian-Albian) in Brazil, the Cambridge Greensand (Cenomanian) in England, the Smoky Hill Chalk Member of the Niobrara Formation (Coniacian-Early Campanian) and the Sharon Springs Formation (Campanian) in North America. Another deposit where a large number of pterosaur specimens are found is the Kimmeridgian-Tithonian Solnhofen Limestone and other lithographic deposits in south Germany, recently regarded as having been formed in a time span of 4 million years (Schweigert 2007Sayão JM. 2003. Histovariability in bones of two pterodactyloid pterosaurs from the Santana Formation, Araripe Basin, Brazil: preliminary results. Geol Soc London, Spec Pub 217: 335-342.). Unfortunately, these deposits have also not provided specimens that could be definitely regarded as part of one population. The lack of stratigraphic control regarding most of the pterosaur material collected in all these deposits is part of the problem and could be solved by controlled excavations.
One of the few examples in which a species is known from a great number of individuals is the archaeopterodactyloid Pterodaustro guinazui. Although hundreds of specimens have been reported, the vast majority is formed by unarticulated material and more complete individuals are very rare (Codorniú and Chiappe 2004Codorniú L and Chiappe LM. 2004. Early juveline pterosaurs (Pterodactyloidea: Pterodaustro guinazui) from the Lower Cretaceous of central Argentina. Can J Earth Sci 41: 9-18.). Unfortunately, due to the nature of the material (very compacted), little is known or has been reported regarding individual variations.
Perhaps the best example of a certain number of pterosaurs potentially belonging to the same population is Quetzalcoatlus sp. from the Javelina Formation (Maastrichtian) at the Big Bend National Park (Texas, USA, see Kellner 1994Kellner AWA. 1994. Remarks on pterosaur taphonomy and paleoecology. Acta Geol Leopol 39: 175-189.). However, even there the total sample is limited to seven individuals (minimum number) whose size difference is less than 10% and that lack any significant morphological variation (A.W.A. Kellner, unpublished data).
Regarding ontogeny, both macroscopic (closed cranial sutures, fusion of several elements such as the proximal syncarpals and the scapula and coracoid) and microscopic (seen in the histological section of the first wing phalanx) features indicate that MN 6594-V represents an adult specimen that has essentially finished its ontogenetic development, ceasing or about to cease growth. The cranial sutures of the holotype of Tropeognathus mesembrinus are also closed and it was therefore regarded as likely representing an adult individual (Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.). Unfortunately there are no postcranial elements known from that individual that would allow further comparisons, including histological sections. Therefore, it is unclear if the holotype of Tropeognathus mesembrinus represents the maximum size reached by this species.
Since the main differences of the new specimen are size and the steeper inclination of the posterior margin of the premaxillary crest, we refer MN 6594-V to Tropeognathus cf. T. mesembrinus.
The two other specimens described here are not complete enough to allow a classification at the generic or specific level. The distal portion of the very large humerus (MCT 1838-R) has a triangular distal articulation, a feature found in pteranodontoids, including anhanguerids. Since the members of the main other clade represented in the Romualdo Formation, the Tapejaridae, possess a quadrangular distal end of the humerus (e.g., Kellner 2003Kellner AWA. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. Geol Soc London, Spec Pub 217: 105-137.), this specimen is tentatively referred to the Anhangueridae. If it represents the same taxon as MN 6594-V is presently unknown.
MPSC R 1395 is composed by a partial wing and lacks any cranial material. The humerus shows a warped deltopectoral crest, which is a typical pteranodontoid feature (e.g., Bennett 2001aBennett SC. 2001a. The osteology and functional morphology of the Late Cretaceous Pterosaur Pteranodon Part 1 - General description and osteology. Palaeont Abt A 260: 1-112., Kellner 2003Kellner AWA. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. Geol Soc London, Spec Pub 217: 105-137.). The proportional size of the wing metacarpal is also consistent with pteranodontoids, particularly the Anhangueridae (Vila Nova & Sayão 2012), to which this specimen is referred.
MAXIMIZED AND NORMAL WINGSPANS
Establishing the wingspan of a pterosaur is always a complex issue and can get even more complicated when the specimen is only known from incomplete bones. Among the problems, there is how one should deal with the natural flexures of several parts of the wing, particularly the elbow and the phalanges of the wing finger. Also the degree of the natural bending of the wing elements during flight influences the wingspan.
In order to establish the wingspan of Pteranodon specimens, Bennett (2001b)Bennett SC. 2001b. The osteology and functional morphology of the Late Cretaceous Pterosaur Pteranodon Part II - Size and Functional Morphology. Palaeont Abt A 260: 113-153. disregarded the lengths of the carpals and the scapulocoracoid, added the lengths of the humerus through the last wing phalanx, and multiplied the final number by two. Other researchers tried to find some compromise that might take those flexures into account or have essentially added the lengths of all those bones (including carpus and the distance between the scapulocoracoid and the body) and multiplied the value by two (e.g., Eaton 1910Eaton GF. 1910. Osteology of Pteranodon. Memoirs of the Connecticut Academy of Arts and Sciences 1: 1-38., Wang et al. 2010Wang X, Campos DA, Zhou Z and Kellner AWA. 2008. A primitive istiodactylid pterosaur (Pterodactyloidea) from the Jiufotang Formation (Early Cretaceous), northeast China. Zootaxa 1813: 1-18.).
Using the reconstruction provided by Wellnhofer (1991a)Wellnhofer P. 1985. Neue Pterosaurier aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 187: 105-182. for AMNH 22552, the ratio between the estimated wingspan (3,270 mm) and the total sum of the lengths of individual wing bones (including the estimated length between the glenoid articulation of the scapulocoracoid and the body, plus the length of the fourth wing phalanx, totalizing 3,445 mm) is about 95%. Based on the reconstructions of Kellner and Tomida (2000)Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135. for Anhanguera piscator and of Bennett (2001a)Bennett SC. 2001a. The osteology and functional morphology of the Late Cretaceous Pterosaur Pteranodon Part 1 - General description and osteology. Palaeont Abt A 260: 1-112. for a general Pteranodon, the values are similar (96-97%), although the natural bending of the wing in the last two cases was not taken into account.
We suggest that, whenever possible, two values of pterosaur wingspan should be presented. The first can be considered the maximized wingspan (maxws) and is essentially the addition of the maximum length of all wing elements, including carpals and the length of the scapula or coracoid (always the smaller of the two). This is a straightforward measure and offers a good basis for comparisons between the wingspans of different taxa or specimens since it shows less subjectivity other than the estimates of the length of bones that might be incomplete. A second value can be considered the normal wingspan (nws), defined as the maximized wingspan reduced by a certain reducing factor (rfc) that should account for the dorsoventral bending and the natural flexures between different parts of the wing. Since proportions of the wing elements vary among pterosaurs (see Kellner 2003Kellner AWA. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. Geol Soc London, Spec Pub 217: 105-137., Vila Nova and Sayão 2012), this reduction is likely to change depending on the clade. For pteranodontoids, we suggest that this reducing factor is in the magnitude of 5%, which is based on the ratio of the estimated wingspan and the total sum of the lengths of individual wing bones taking into account the data and reconstructions provided by Wellnhofer (1991a)Wellnhofer P. 1985. Neue Pterosaurier aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 187: 105-182., Kellner and Tomida (2000)Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135., and Bennett (2001a)Bennett SC. 2001a. The osteology and functional morphology of the Late Cretaceous Pterosaur Pteranodon Part 1 - General description and osteology. Palaeont Abt A 260: 1-112..
The calculation of the wingspan of MN 6594-V was made based on comparisons with Anhanguera piscator and AMNH 22552. The latter has been regarded by Wellnhofer (1991a)Wellnhofer P. 1985. Neue Pterosaurier aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 187: 105-182. to represent Santanadactylus pricei, but this assignment is problematic (see Kellner and Tomida 2000Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.). With the measurement of the distal width of the humerus of MN 6594-V (112 mm), the total length of this bone was estimated using the proportions of the humerus length relative to the distal width based on the measurements available for Anhanguera piscator and AMNH 22552 (Table I). Since the ratios vary, a mean value was obtained (408 mm). This value and the ratios obtained from Anhanguera piscator and AMNH 22552 were used to estimate the lengths of all other bones (Tables II and III). The maximized wingspan of MN 6594-V is 8.70 m, with a normal wingspan of 8.26 m.
Anhanguera piscator (NSM-PV 19892) and AMNH 22552 ratios based, respectively on the measurements presented by Kellner and Tomida (2000)Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135. and Wellnhofer (1991a)Wellnhofer P. 1985. Neue Pterosaurier aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 187: 105-182..
Estimated lengths (in mm) of the forelimb and hind limb elements of Tropeognathus cf. T. mesembrinus (MN 6594-V) based on the bone ratios of Anhanguera piscator (NSM-PV 19892) and AMNH 22552 (Table 2). The length of scapula and carpals were kept constant since they were measured directly on the specimen. Abbreviations: car - carpals, fe - femur, hu - humerus, maxws - maximized wingspan, mcIV - metacarpal IV, nws - normal wingspan, ph1d4 - phalanx 1 of manual digit 4, ph2d4 - phalanx 2 of manual digit 4, ph3d4 - phalanx 3 of manual digit 4, ph4d4 - phalanx 4 of manual digit 4, sca - scapula, ti - tibia ul - ulna. The maxws was obtained by the addition of the lengths of sca, hu, ul, car, mcIV, ph1d4, ph2d4, ph3d4 and ph4d4 multiplied by 2, the nws was calculated by multiplying the maxws by 0.95.
It should be noted that the distal articulation of MCT 1838-R is larger (127 mm) and that it represents an animal with a maximized and a normal wingspan of about 9 m. These two specimens show that large pterosaurs, some reaching gigantic sizes, have formed a considerable part of the pterosaur fauna from the Romualdo Formation.
The estimated length of the humerus and the wing metacarpal of MPSC R 1395 indicates an individual with a maximized wingspan of 5.5 m, with a normal wingspan of around 5.2 m and skull length around 630 mm. It is important to point out that the extensor tendon process of the first wing phalanx and the epiphysis of the humerus are both unfused, clearly indicating that this specimen represents a young animal. If it attained gigantic size after completing its ontogeny, a strong possibility, cannot be conclusively established at present.
GIANT PTEROSAURS - DEFINITION
Still concerning pterosaur wingspans, some individuals have been occasionally regarded as giants. However, there is no definition about which size a species of those volant archosaurs should have to fall into this category (e.g., Averianov et al. 2005Averianov AO, Arkhangelsky MS, Pervushov EM and Ivanov AV. 2005. A new record of an Azhdarchid (Pterosauria: Azhdarchidae) from the Upper Cretaceous of the Volga Region. Paleont J 40: 91-97., Witton and Naish 2008Witton M. 2009. A new species of Tupuxuara (Thalassodromidae, Azhdarchoidea) from the Lower Cretaceous Santana Formation of Brazil, with a note on the nomenclature of Thalassodromidae. Cretaceous Res 30: 1293-1300., Rodrigues et al. 2011Rodrigues T and Kellner AWA. 2008. Review of the pterodactyloid Coloborhynchus. Zitteliana B 28: 210-228., Novas et al. 2012Neumann VH and Cabrera L. 1999. Una nueva propuesta estratigráfica para la tecnosecuencia post-rifte de la cuenca de Araripe, Noreste de Brasil. In: Boletim do 5° Simpósio sobre o Cretáceo do Brasil, Serra Negra, p. 279-285.). Witton and Habib (2010)Wellnhofer P. 1991b. The illustrated encyclopedia of pterosaurs. London: Salamander Books, 192 p., for example, restrict the term giant pterosaur to Pteranodon and the largest azhdarchids, implying that all pterosaurs with a wingspan over 7 m fall into this category, without further discussion.
Ideally, in order to establish the size variation within a particular group, even such a diverse one as pterosaurs, it would be important to have data showing the wingspan considering the whole spectrum of these animals. However, the available information about pterosaur diversity is strongly biased towards the occurrences of species that lived close to or at shorelines and concentrated in very few deposits (Kellner 1994Kellner AWA. 1994. Remarks on pterosaur taphonomy and paleoecology. Acta Geol Leopol 39: 175-189.). Furthermore, most species are based on incomplete specimens (e.g., Wellnhofer 1991bWellnhofer P. 1987. New crested pterosaurs from the Lower Cretaceous of Brazil. Mitt Bayer Staatssg Paläontol Hist Geol 27: 175-186.), making the evaluation of their wingspan problematic.
Considering living flying vertebrates, one of the largest bats is the giant golden-crowned flying fox (Aceron jubatus) that achieves a wingspan of around 1.8 m (Nowak 1999Novas FE, Kundrat M, Agnolín FL, Ezcurra MD, Ahlberg PE, Isasi MP, Arriagada A and Chafrat P. 2012. A new large pterosaur from the Late Cretacous of Patagonia. J Vert Paleont 32: 1147-1452.). Regarding birds, the species with the largest wingspan recorded so far is Argentavis magnificens recovered from Miocene deposits of Argentina, which is estimated around 7 m (Campbell and Tonni 1980Campbell KE and Tonni EP. 1980. A new genus of teratorn from the Huayquerian of Argentina (Aves: Teratornithidae). Cont. in Science, Nat Hist Mus Los Angeles County 330: 59-68.). Among the living birds the two taxa that share the largest wingspans are the albatross Diomedea exulans Linnaeus, 1758 (3.7 m) and the marabu Leptoptilos crumeniferus Lesson, 1831 (3.5 m). Although admittedly arbitrary, we suggest that a pterosaur whose wingspan is larger than 7.4 m (double that of an albatross) should fall into the category of a giant.
Up to date, the largest toothed pterosaur reported is represented by a fragment of a skull (NHMUK R481) assigned by Martill and Unwin (2012)Martill DM, Frey E, Sadaqah RM and Khoury HN. 1998. Discovery of the holotype of the giant pterosaur Titanopteryx philadelphiae Arambourg 1959, and the status of Arambourgiania and Quetzalcoatlus. N Jb Geol Paläont Abh 207: 57-76. to Coloborhynchus capito (Seeley, 1870). Although this specimen is undoubtedly large, its taxonomy is questionable. The holotype of “Coloborhynchus” capito (CAMSM B 54625, whose assignment to this genus is also disputed, see Rodrigues and Kellner 2008Price LI. 1971. A presença de Pterosauria no Cretáceo Inferior da Chapada do Araripe, Brasil. An Acad Bras Cienc 43(Supl): 451-461.) is a very fragmentary specimen whose most prominent and diagnostic feature is the presence of a premaxillary crest that is concave in lateral view. NHMUK R481, being preserved basically as an inch-thick transverse section, does not show this feature, and thus cannot be confidently referred to this species. Although aware about the problematic to establish the wingspan of NHMUK R481, Martill and Unwin (2012)Martill DM, Frey E, Sadaqah RM and Khoury HN. 1998. Discovery of the holotype of the giant pterosaur Titanopteryx philadelphiae Arambourg 1959, and the status of Arambourgiania and Quetzalcoatlus. N Jb Geol Paläont Abh 207: 57-76. tentatively regarded this specimen as representing an animal with a wingspan up to 7 m. Those authors also set this as the upper size limit for toothed pterosaurs, but did not explain why this would be the case. Anyhow, MN 6594-V and MCT 1838-R clearly show that the wingspan of toothed pterosaurs could exceed 7 m.
The occurrence of large and, according to our definition, giant pterosaurs in the Romualdo Formation has been suggested before by Dalla Vecchia and Ligabue (1993) on the basis of a large isolated distal phalanx of a pterodactyloid. Martill and Unwin (2012)Martill DM, Frey E, Sadaqah RM and Khoury HN. 1998. Discovery of the holotype of the giant pterosaur Titanopteryx philadelphiae Arambourg 1959, and the status of Arambourgiania and Quetzalcoatlus. N Jb Geol Paläont Abh 207: 57-76. accepted the large size represented by this specimen, but stated that this bone likely belonged to an azhdarchoid, suggesting that it might represent a large individual of Thalassodromeus or Tupuxuara. Although the identity of the isolated phalanx described by Dalla Vecchia and Ligabue (1993) cannot be assessed with confidence, it should be noted that there is a considerable difference in the proportions of the wing elements of azhdarchoids (including thalassodromines) and pteranodontoids (including anhanguerids). While pteranodontoids have wings comparatively large relative to the body (e.g., Bennett 2001aBennett SC. 2001a. The osteology and functional morphology of the Late Cretaceous Pterosaur Pteranodon Part 1 - General description and osteology. Palaeont Abt A 260: 1-112., Kellner 2003Kellner AWA. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. Geol Soc London, Spec Pub 217: 105-137., Vila Nova and Sayão 2012), azhdarchoids are short winged (e.g., Kellner 2003Kellner AWA. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. Geol Soc London, Spec Pub 217: 105-137.). Therefore, the largest flying creatures in the Romualdo Formation are pteranodontoids (including anhanguerids) and not azhdarchoids (including thalassodromines). We suspect that this might have also been the case in most pterosaur faunas where these clades have been recorded (e.g., Jiufotang Formation).
Still regarding the isolated phalanx described by Dalla Vecchia and Ligabue (1993), if it indeed represents an azhdarchoid, it must have belonged to a pterosaur with a wingspan considerably smaller than MN 6594-V and MCT 1838-R. However, if it represents a pteranodontoid, as suspected here, it belonged to an individual larger than MN 6594-V and perhaps with the same size or slightly smaller than MCT 1838-R. In any case, these three specimens (MN 6594-V, MPSC R 1395 and MCT 1838-R) show that large and gigantic pterosaurs formed an expressive part of the pterosaur fauna from the Romualdo Formation. Tropeognathus cf. T. mesembrinus is presently the largest flying vertebrate recovered from any Gondwanan deposit. The sole other species that could potentially be larger is Arambourgiania philadelphiae (Arambourg, 1959), known from an incomplete isolated cervical vertebra found in Maastrichtian deposits of Jordan. Martill et al. (1998) tentatively estimated the wingspan of this pterosaur as 12 m, but Suberbiola et al. (2003) suggested 7 m instead. MCT 1838-R that may or may not belong to the same species as MN 6594-V, represents still a larger animal, and at least also potentially the isolated distal end of the wing phalanx described by Dalla Vecchia and Ligabue (1993). Since the age of the Romualdo Formation is regarded as Aptian/Albian, these specimens show that pterosaurs have developed large and some gigantic sizes at least as early as the middle Cretaceous. Lastly, MN 6594-V provides the first information on the postcranial skeleton of Tropeognathus, which is very similar to Anhanguera, suggesting limited postcranial variation within the Anhangueridae.
CONCLUSION
MN 6594-V is referred to Tropeognathus cf. T. mesembrinus based mainly on the comparatively reduced number of teeth, the presence of a broad and blunt frontoparietal crest, the developed palatal ridge and the dorsal portion of the occiput more inclined posteriorly. Among others, MN 6594-V differs from the holotype of Tropeognathus mesembrinus by being about 55% longer and showing a steeper posterior margin of the sagittal crest. Tropeognathus cf. T. mesembrinus is the largest pterosaur recovered from Gondwanan deposits so far and, along with MPSC R 1395 and MCT 1838-R, shows that large taxa represented a significant part of the pterosaur fauna from the Aptian/Albian Romualdo Formation. Furthermore, these and other specimens (e.g., Dalla Vecchia and Ligabue 1993) show that pterosaurs that developed large and gigantic sizes (here defined as specimens with wingspans larger than 7.4 m) evolved at least as early as the middle Cretaceous. MN 6594-V also provides the first information on the postcranial skeleton of Tropeognathus, indicating little variation within anhanguerids.
ACKNOWLEDGMENTS
We have profited from discussions with Fabio Dalla Vecchia (Institut Català de Paleontologia ‘Miquel Crusafont’, Sabadell, Spain) and Renan Bantim (Universidade Federal de Pernambuco) for comments on earlier version of the manuscript. This study was partially funded by the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ #E-26/111.273/2010 and E-26/102.737/2012 to AWAK, #E-26/100.458/2012 to FRC), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq #307276/ 2009-0 to AWAK and CNPq #401787/2010-9 to JMS) and Fundação de Amparo à Pesquisa do Espírito Santo (FAPES #52986870/2011 to TR).
REFERENCES
- Assine ML. 2007. Bacia do Araripe. Boletim de Geociências da Petrobras 15: 371-389.
- Averianov AO, Arkhangelsky MS, Pervushov EM and Ivanov AV. 2005. A new record of an Azhdarchid (Pterosauria: Azhdarchidae) from the Upper Cretaceous of the Volga Region. Paleont J 40: 91-97.
- Bennett SC. 1993. The ontogeny of Pteranodon and other pterosaurs. Paleobiology 19: 92-106.
- Bennett SC. 2001a. The osteology and functional morphology of the Late Cretaceous Pterosaur Pteranodon Part 1 - General description and osteology. Palaeont Abt A 260: 1-112.
- Bennett SC. 2001b. The osteology and functional morphology of the Late Cretaceous Pterosaur Pteranodon Part II - Size and Functional Morphology. Palaeont Abt A 260: 113-153.
- Beurlen K. 1962. Geologia da Chapada do Araripe. An Acad Bras Cienc 34: 365-370.
- Beurlen K. 1963. Geologia e Estratigrafia da Chapada do Araripe. 17 Congresso Brasileiro de Geologia Publ. SUDENTE: 1-47.
- Beurlen K. 1971. As condições ecológicas e faciológicas da Formação Santana na Chapada do Araripe (Nordeste do Brasil). An Acad Bras Cienc 43: 411-415.
- Campbell KE and Tonni EP. 1980. A new genus of teratorn from the Huayquerian of Argentina (Aves: Teratornithidae). Cont. in Science, Nat Hist Mus Los Angeles County 330: 59-68.
- Campos DA and Kellner AWA. 1985. Panorama of the Flying Reptiles Study in Brazil and South America. An Acad Bras Cienc 57: 453-466.
- Chinsamy A, Codorniú L and Chiappe L. 2008. Developmental growth patterns of the filter-feeder pterosaur, Pterodaustro guinazui. Biol Lett 4: 282-285.
- Chinsamy A, Codorniú L and Chiappe L. 2009. Palaeobiological Implications of the Bone Histology of Pterodaustro guinazui. Anat Rec 292: 1462-1477.
- Codorniú L and Chiappe LM. 2004. Early juveline pterosaurs (Pterodactyloidea: Pterodaustro guinazui) from the Lower Cretaceous of central Argentina. Can J Earth Sci 41: 9-18.
- Comissão Especial de Nomenclatura Estratigráfica. 1986. Código Brasileiro de Nomenclatura Estratigráfica - Guia de Nomenclatura Estratigráfica. Rev Bras Geo 16: 370-415.
- Dalla Vecchia FM and Ligabue G. 1993. On the presence of a giant pterosaur in the Lower Cretaceous (Aptian) of Chapada do Araripe (northeastern Brazil). Boll Soc Paleont Italiana 32: 131-136.
- Eaton GF. 1910. Osteology of Pteranodon. Memoirs of the Connecticut Academy of Arts and Sciences 1: 1-38.
- Eck K, Elgin RA and Frey E. 2011. On the osteology of Tapejara wellnhoferi Kellner 1989 and the first occurrence of multiple specimen assemblage from the Santana Formation, Araripe Basin, NE-Brazil. Swiss J Palaeontol 130: 277-296.
- Fara E, Saraiva AAF, Campos DA, Moreira JKR, Siebra DC and Kellner AWA. 2005. Controlled excavations in the Romualdo Member of the Santana Formation (Early Cretaceous, Araripe Basin, northeastern Brazil): stratigraphic, palaeoenvironmental and palaeoecological implications. Palaeogeogr Palaeoclimatol Palaeoecol 218: 145-160.
- Fastnacht M. 2001. First record of Coloborhynchus (Pterosauria) from the Santana Formation (Lower Cretaceous) of the Chapada do Araripe, Brazil. Paläontol Z 75: 3-36.
- Frey E, Martill DM and Buchy MC. 2003a. A new crested ornithocheirid from the Lower Cretaceous of northeastern Brazil and the unusual death of an unusual pterosaur. Geol Soc London, Spec Pub 217: 55-63.
- Frey E, Tischlinger H, Buchy MC and Martill DM. 2003b. New specimens of Pterosauria (Reptilia) with soft parts with implications for pterosaurian anatomy and locomotion. Geol Soc London, Spec Pub 217: 233-266.
- Horner RJ, Padian K and Ricqles A. 1999. Variation in dinosaur skeletochronology indicators: implications for age assessment and physiology. Paleobiology 25(3): 295-304.
- International Subcommission on Stratigraphic Classification. 1999. International Stratigraphic Guide - An abridged edition. In: MURPHY MA AND SALVADOR A (Eds), Episodes 22: 255-271.
- Kellner AWA. 1994. Remarks on pterosaur taphonomy and paleoecology. Acta Geol Leopol 39: 175-189.
- Kellner AWA. 1996. Reinterpretation of a remarkably well-preserved pterosaur soft tissue from the Early Cretaceous of Brazil. J Vert Paleont 16: 718-722.
- Kellner AWA. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. Geol Soc London, Spec Pub 217: 105-137.
- Kellner AWA. 2006. Pterossauros - os senhores do céu do Brasil. Rio de Janeiro: Vieira & Lent, 176 p.
- Kellner AWA. 2010. Comments on the Pteranodontidae (Pterosauria, Pterodactyloidea) with the description of two new species. An Acad Bras Cienc 82: 1063-1084.
- Kellner AWA and Campos DA. 1999. Vertebrate Paleontology in Brazil – a review. Episodes 22: 238-251.
- Kellner AWA and Campos DA. 2002. The function of the cranial crest and jaws of a unique pterosaur from the Early Cretaceous of Brazil. Science 297: 389-392.
- Kellner AWA and Langston WJR. 1996. Cranial remains of Quetzalcoatlus (Pterosauria, Azhdarchidae) from the Late Cretaceous Sediments of Big Bend National Park, Texas. J Vert Paleont 16: 222-231.
- Kellner AWA and Tomida Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum Monographs 17: 1-135.
- Maisey J. (Ed). 1991. Santana fossils: an illustrated Atlas. Neptune, New Jersey: T.F.H. Publications, 459 p.
- Martill DM. 2007. The age of the Cretaceous Santana Formation fossil Konservat Lagerstätte of north-east Brazil: a historical review and an appraisal on the biochronostratigraphic utility of its paleobiota. Cretaceous Res 28: 895-920.
- Martill DM, Frey E, Sadaqah RM and Khoury HN. 1998. Discovery of the holotype of the giant pterosaur Titanopteryx philadelphiae Arambourg 1959, and the status of Arambourgiania and Quetzalcoatlus. N Jb Geol Paläont Abh 207: 57-76.
- Martill DM and Unwin DM. 2012. The world's largest toothed pterosaur, NHMUK R481, an incomplete rostrum of Coloborhynchus capito (Seeley, 1870) from the Cambridge Greensand of England. Cretaceous Res 34: 1-9.
- Neumann VH and Cabrera L. 1999. Una nueva propuesta estratigráfica para la tecnosecuencia post-rifte de la cuenca de Araripe, Noreste de Brasil. In: Boletim do 5° Simpósio sobre o Cretáceo do Brasil, Serra Negra, p. 279-285.
- Novas FE, Kundrat M, Agnolín FL, Ezcurra MD, Ahlberg PE, Isasi MP, Arriagada A and Chafrat P. 2012. A new large pterosaur from the Late Cretacous of Patagonia. J Vert Paleont 32: 1147-1452.
- NOWAK RM. 1999. Walker's Mammals of the World, 6th ed., Baltimore and London: The John Hopkins University Press, 1936 p.
- Pons D, Berthou PY and Campos DA. 1990. Quelques observations sur la palynologie de l'Aptien supérieur et del'Albien do bassin d'Araripe (N-E du Brasil). In: Campos DA, Viana MSS, Brito PM, Beurlen G (Eds), Atas do Simpósio sobre a Bacia do Araripe e Bacias interiores do Nordeste, Crato, p. 142-252.
- Price LI. 1971. A presença de Pterosauria no Cretáceo Inferior da Chapada do Araripe, Brasil. An Acad Bras Cienc 43(Supl): 451-461.
- Rodrigues T and Kellner AWA. 2008. Review of the pterodactyloid Coloborhynchus. Zitteliana B 28: 210-228.
- Rodrigues T, Kellner AWA, Mader BJ and Russell D. 2011. New pterosaur specimens from the Kem Kem beds (Upper Cretaceous, Cenomanian) of Morocco. Riv Ital Paleontol S 117: 149-160.
- Santos RS and Valença JG. 1968. A Formação Santana e sua paleoictiofauna do Cretáceo do Brasil. An Acad Bras Cienc 40: 339-360.
- Sayão JM. 2003. Histovariability in bones of two pterodactyloid pterosaurs from the Santana Formation, Araripe Basin, Brazil: preliminary results. Geol Soc London, Spec Pub 217: 335-342.
- Schweigert G. 2007. Ammonite biostratigraphy as a tool for dating Upper Jurassic lithographic limestones from South Germany - first results and open questions. N Jb Geol Paläont Abh 245:117-125.
- Silva HP and Kellner AWA. 2006. A combinação de técnicas na preparação de vertebrados fósseis: o exemplo de Thalassodromeus sethi (Pterosauria, Tapejaridae). In: GALLO V ET AL. (Eds), Paleontologia de Vertebrados: Grandes Temas e Contribuições Científicas. Rio de Janeiro: Interciência, p. 193-301.
- Small H. 1913. Geologia e suprimento de agua subterranea no Ceará e parte do Piaui. Insp. Obras Contra Secas, Serie Geologia 25: 1-180.
- Suberbiola XP, Bardet N, Jouve S, Iarochène M, Bouya B and Amaghzaz M. 2003. A new azhdarchid pterosaur from the Late Cretaceous phosphates of Morocco. Geol Soc London, Spec Pub 217: 79-90.
- Valença LMM, Neumann VH and Mabesoone JM. 2003. An overview on Callovian-Cenomanian intracratonic basins of Northeast Brazil: onshore stratrigraphic record of the opening of the southern Atlantic. Geol Acta 1: 261-275.
- Veldmeijer AJ. 2002. Pterosaurs from the Lower Cretaceous of Brazil in the Stuttgart collection. Stuttgarter Beitr Naturk (B) 327: 1-27.
- Veldmeijer AJ. 2003. Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil. Scripta Geol 125: 35-139.
- Vila Nova BC, Saraiva AAF, Moreira JKR and Sayão JM. 2011. Controlled excavations in the Romualdo Formation lagerstätte (Araripe Basin, Brazil) and pterosaur diversity: remarks based on new findings. Palaios 26: 173-179.
- Vila Nova BC and Sayão JM. 2012. On wing disparity and morphological variation of the Santana Group pterosaurs. Hist Biol 24: 567-574.
- Wang X, Campos DA, Zhou Z and Kellner AWA. 2008. A primitive istiodactylid pterosaur (Pterodactyloidea) from the Jiufotang Formation (Early Cretaceous), northeast China. Zootaxa 1813: 1-18.
- Wang X, Kellner AWA, Jiang S and Cheng X. 2012. New toothed flying reptile from Asia: close similarities between early Cretaceous pterosaur faunas from China and Brazil. Naturwissenschaften 99: 249-257
- Wang XL, Kellner AWA, Jiang SX, Cheng X, Meng X and Rodrigues T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. An Acad Bras Cienc 82: 1045-1062.
- Wellnhofer P. 1985. Neue Pterosaurier aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 187: 105-182.
- Wellnhofer P. 1987. New crested pterosaurs from the Lower Cretaceous of Brazil. Mitt Bayer Staatssg Paläontol Hist Geol 27: 175-186.
- Wellnhofer P. 1991a. Weitere Pterosaurierfunde aus der Santana Formation (Apt) der Chapada do Araripe. Palaeont A 215: 43-101.
- Wellnhofer P. 1991b. The illustrated encyclopedia of pterosaurs. London: Salamander Books, 192 p.
- Witton M. 2009. A new species of Tupuxuara (Thalassodromidae, Azhdarchoidea) from the Lower Cretaceous Santana Formation of Brazil, with a note on the nomenclature of Thalassodromidae. Cretaceous Res 30: 1293-1300.
- Witton MP and Habib MB. 2010. On the size and flight diversity of giant pterosaurs, the use of birds as pterosaur analogues and comments on the pterosaur flightlessness. PLoS ONE 5(11): e13982
- Witton MP and Naish D. 2008. A reappraisal of azhdarchid pterosaur functional morphology and paleoecology. PLoS ONE 3: e2271.
Publication Dates
-
Publication in this collection
Jan-Mar 2013
History
-
Received
4 Jan 2013 -
Accepted
19 Feb 2013