ABSTRACT
The intercropping is an important cultural practice commonly used in pest management. It is based on the principle that increased plant diversity in the agro-ecosystem can lead to reductions of pest populations in the crop. The current study aimed to assess the impact the colored fiber cotton-cowpea intercropped systems on Aphis gossypii and Aphis craccivora and on their predator Cycloneda sanguinea and the losses and the dispersion behavior of these aphids and their predator in these cropping systems. The experiment had a randomized block experimental design with two bioassays and four treatments. The number of apterous and alate aphids (A. gossypii) per cotton plant was 1.46 and 1.73 or 1.97 and 2.19 times highest in the solid cotton system than that found in the cotton-cowpea intercropped systems (S1) and (S2), respectively. On the other hand, the cotton-cowpea intercropped systems (S1 and S2) reduced, respectively, in 43% and 31% the number of apterousA. gossypiiper cotton plant compared to the control. Implementing cotton-cowpea intercropped system in the S1 scheme reduced A. gossypii infestation, favored the multiplication of C. sanguinea, and allowed obtaining heavier open bolls.
Key words:
aphids; cowpea; cotton; behavior; intercropping
INTRODUCTION
Cotton (Gossypium hirsutum L.) cultivars with naturally colored fibers now have worldwide demand in the textile industry and have added value to the crop for small farmers (Fernandes et al. 2012aFERNANDES FS, RAMALHO FS, NASCIMENTO ARB, MALAQUIAS JB, SILVA CAD AND ZANUNCIO JC. 2012a. Within-plant distribution of cotton aphid (Hemiptera: Aphididae), in cotton with colored fibers and cotton-fennel intercropping system. Ann Entomol Soc Am 105: 599-607., Ramalho et al. 2012aRamalho FS, Fernandes FS, Nascimento ARB, Nascimento Júnior JL, Malaquias JB AND Silva CAD. 2012a. Assessment of fennel aphids (Hemiptera: Aphididae) and their predators in fennel intercropped with cotton with colored fibers. J Econ Entomol 105: 113-119.). Cowpea (Vigna unguiculata L. Walp.) fixs nitrogen to the soil and it is a protein supply for people in northeastern Brazil (Braga et al. 2007BRAGA YFB, GRANGEIRO TB, FREIRE EA, LOPES HL, BEZERRA JNS, ANDRADE-NETO M AND LIMA MAS. 2007. Insecticidal activity of 2-tridecanone against the cowpea weevil Callosobruchus maculatus (Coleoptera: Bruchidae). An Acad Bras Cienc 79: 35-39., Frota et al. 2008Frota KMG, Soares RAM AND Arêas JAG. 2008. Composição química do feijão caupi (Vigna unguiculata L. Walp ), cultivar BRS-Milênio. Food Sci Technol 28: 470-476.). These agricultural products contribute to employment and income of farmers in this Brazilian region. However, Aphis gossypii Glov. (Fernandes et al. 2012a) and Aphis craccivora K. (Hemiptera: Aphididae), respectively, have been reducing cotton cowpea production in Brazil. These aphids cause direct damage by sucking sap and indirect ones by virus transmittion and excreting honeydew that favors the proliferation of leaf stomata-obstructing fungi (Capnodium spp) (Kitajima et al. 2008Kitajima EW, Alcântara BK, Madureira PM, Alfenas-Zerbini P, Rezende JAM AND Zerbini FM. 2008. A mosaic of beach bean (Canavalia rosea) caused by an isolate of cowpea aphid-borne mosaic virus (CABMV). Arch Virol 153: 743-747., Fernandes et al. 2012a, 2013, Bachmann et al. 2014Bachmann AC, Nault BA AND Fleischer SJ. 2014. Alate aphid (Hemiptera : Aphididae) species composition and richness in northeastern USA snap beans and an update to historical lists. Fla Entomol 97: 979-994., Malaquias et al. 2014Malaquias JB, Ramalho FS, Lira ACS, Oliveira FQ, Fernandes FS, Godoy WAC AND Zanuncio JC. 2014. The biology and thermal requirements of the fennel aphid Hyadaphis foeniculi (Passerini) (Hemiptera: Aphididae). PLoS ONE 9(7):e100983.).
Aphis gossypii damages cotton plants during both vegetative and reproductive stages (Ramalho et al. 2012aRamalho FS, Fernandes FS, Nascimento ARB, Nascimento Júnior JL, Malaquias JB AND Silva CAD. 2012a. Assessment of fennel aphids (Hemiptera: Aphididae) and their predators in fennel intercropped with cotton with colored fibers. J Econ Entomol 105: 113-119., Fernandes et al. 2012bFERNANDES FS, RAMALHO FS, NASCIMENTO JL, MALAQUIAS JL, NASCIMENTO ARB, SILVA CAD AND ZANUNCIO JC. 2012b. Within-plant distribution of cotton aphids, Aphis gossypii Glover (Hemiptera: Aphididae), in Bt and non-Bt cotton fields. Bul Entomol Res 102: 79-87. ) and many plant species including eggplants and okra (Leite et al. 2006LEITE GLD, PICANÇO M, ZANUNCIO JC AND ECOLE CC. 2006. Factors affecting herbivory of Thrips palmi (Thysanoptera: Thripidae) and Aphis gossypii (Homoptera: Aphididae) on the eggplant (Solanum melongena). Braz Arch Biol Techn 49: 361-369. , 2007). Infestation in vegetative stages turn leaves shriveled, whereas infestation in reproductive stages, especially at fruit opening, fouls and reduces fiber quality (Almeida 2001Almeida RP. 2001. Effect of the population levels of Aphis gossypii on cotton agronomic traits and fibre quality. Proc Sec Exp and Appl Entomol 12: 97-100.). Severe attacks by A. gossypii may also reduce leaf area and biomass, besides branching and plant height, harming the crop and results in economic losses (Sarwar et al. 2014Sarwar MK, Azam I, Iram N, Iqbal W, Rashda A, Anwer F, Atta K AND Ali R. 2014. Cotton aphid Aphis gossypii L. (Homoptera; Aphididae); a challenging pest; biology and control strategies: a review. Int J Appl Biol Pharm Technol 5: 288-294.).
Aphis craccivora initially infests cowpea seedlings, but as this plant develops, it may infest flowers and pods (Berberet et al. 2009Berberet RC, Giles KL, Zarrabi AA AND Payton ME. 2009. Development, reproduction, and within-plant infestation patterns of Aphis craccivora (Homoptera: Aphididae) on alfafa. Environ Entomol 38: 1765-1771.). Its feeding causes leaf shriveling and bud deformation and as the population increases, plant infested become weaked due to loss of sap and toxins injected (Silva et al. 2005Silva PHS, Carneiro JS AND Quinderé MAW. 2005. Pragas. In: Freire Filho FR, Lima JAA and Ribeiro VQ (Eds), Feijão-caupi: avanços tecnológicos. Brasília: Embrapa Informação Tecnológica, Teresina: Embrapa Meio Norte, p. 369-402.). Aphis craccivora is an efficient virus vector, transmitting Cowpea aphidborne mosaic virus (CABMV) (Kitajima et al. 2008Kitajima EW, Alcântara BK, Madureira PM, Alfenas-Zerbini P, Rezende JAM AND Zerbini FM. 2008. A mosaic of beach bean (Canavalia rosea) caused by an isolate of cowpea aphid-borne mosaic virus (CABMV). Arch Virol 153: 743-747.), and Blackeye mosaic virus (BICpMV) (Lima et al. 1981Lima JAA, Oliveira FMEW, Kitajima EW AND Lima MGA. 1981. Propriedades biológicas sorológicas, citológicas e sorológicas de um potyvirus isolado de feijão-de-corda no Ceará. Fitopatol Bras 6: 205-216.). Aphis craccivora can decrease seed quality and plant productivity in commercial bean plants (Obopile 2006Obopile M. 2006. Economic threshold and injury levels for control of cowpea aphid, Aphis crassivora Linnaeus (Homoptera: Aphididae), on cowpea. Afr Plant Prot 12: 111-115., Laamari et al. 2008Laamari M, Khelfa L AND Coer d’Acier A. 2008. Resistance source to cowpea aphid (Aphis craccivora Koch) in broad bean (Vicia faba L.) Algerian landrace collection. Afr J Biotechnol 7: 2486-2490.) with yield losses as high as 50% (Berlandier and Sweetingham 2003Berlandier FA AND Sweetingham MW. 2003. Aphid feeding damage causes large losses in susceptible lupin cultivars. Aust J Exp Agr 43: 1357-1362., Obopile 2006).
In Northeastern Brazil, aphid are usually controlled with non-selective synthetic insecticides that are effective against pests but often cause mortality in beneficial insect (Ullah and Paul 1985Ullah K AND Paul P. 1985. Chemical control of cotton aphid. Pakistan J Agric Res 6: 213-215., Leite et al. 2010Leite MIS, Carvalho GA, Maia JB, Makiyama L AND Vilela M. 2010. Ação residual de inseticidas para larvas e adultos do predador Cycloneda sanguinea Linnaeus, 1763 (Coleoptera: Coccinellidae ). Arq Inst Biol 77: 275-282.). Agro-ecosystem diversification through intercropping may reduce insect pest infestations without the negative impacts of chemical measures (Gonzaga et al. 1991Gonzaga JV, Ramalho FS AND Santos JW. 1991. Distribuição do Aphis gossypii no algodoeiro nos sistemas de plantio solteiro e consorciado. Pesqui Agropecu Bras 26: 1839-1844., Medeiros et al. 2009Medeiros MA, Sujii ER AND Morais HC. 2009. Effect of plant diversification on abundance of South American tomato pinworm and predators in two cropping systems. Hort Bras 27: 300-306., Fernandes et al. 2012cFernandes FS, Ramalho FS, MALAQUIAS JB, Nascimento JUNIOR JL, CORREIA ET AND Zanuncio JC. 2012c. Within-plant distribution of cotton aphid (Hemiptera: Aphididae) in cotton cultivars with colored fibers. An Acad Bras Cienc 84: 707-719. , Ramalho et al. 2012aRamalho FS, Fernandes FS, Nascimento ARB, Nascimento Júnior JL, Malaquias JB AND Silva CAD. 2012a. Assessment of fennel aphids (Hemiptera: Aphididae) and their predators in fennel intercropped with cotton with colored fibers. J Econ Entomol 105: 113-119., b). The possibility of intercropping colored fiber cotton with fennel (Foeniculum vulgare Mill.) was studied (Ramalho et al. 2015) but the use of cowpea for the management of A. gossypii and A. craccivora populations has not been explored. Naturally colored cotton is ecologically friendly because it eliminates the dyeing process in industrial production with frequently (and perhaps incorrectly) chemicals use dangerous to humans (Horstmann 1995Horstmann G. 1995. Dyeing as a new environmental challenge. J Soc Dyers Colour 111: 182-184.).
Well-structured and connected habitat may decreases interspecific competition by allowing species to segregate spatially (Boeye et al. 2014Boeye J, Kubisch A AND Bonte D. 2014. Habitat structure mediates spatial segregation and therefore coexistence. Landscape Ecol 29: 593-604.). In contrast, different plant species in intercropped may change arthropod communities favoring the natural enemies (predators and parasitoids) and reducing pest populations (Gonzaga et al. 1991Gonzaga JV, Ramalho FS AND Santos JW. 1991. Distribuição do Aphis gossypii no algodoeiro nos sistemas de plantio solteiro e consorciado. Pesqui Agropecu Bras 26: 1839-1844.). Intercropping can also increase the productivity of agricultural crops (Ramalho et al. 2012aRamalho FS, Fernandes FS, Nascimento ARB, Nascimento Júnior JL, Malaquias JB AND Silva CAD. 2012a. Assessment of fennel aphids (Hemiptera: Aphididae) and their predators in fennel intercropped with cotton with colored fibers. J Econ Entomol 105: 113-119., b).
Intercropped plants may repel pests or attract natural enemies (Kadam et al. 2014Kadam D, Kadam D AND Lekurwale R. 2014. Seasonal incidence of sucking pests of Bt cotton in different intercropping systems and their natural enemies. J Plant Pest Sci 1: 29-34.). Predatory lady beetles (Coleoptera: Coccinellidae) may move between crops (Bastola et al. 2014Bastola A, Parajulee MN, Porter RP, Shrestha RB, Chen FJ AND Carroll SC. 2014. Intercrop movement of convergent lady beetle, Hippodamia convergens (Coleoptera: Coccinellidae), between adjacent cotton and alfalfa. Insect Sci 00: 1-12.). Coccinellid may be attracted to particular crops due to prey availability and plant features such as the resource provision; shelter, protection and feeding sites (Resende et al. 2012Resende ALS, Haro MM, Silva VF, Souza B AND Silveira LCP. 2012. Diversidade de predadores em coentro, endro e funcho sob manejo orgânico. Arq Inst Biol 79: 193-199.), and plant odors can attract predators in intercropped systems (Ninkovic and Pettersson 2003Ninkovic V AND Pettersson J. 2003. Searching behaviour of the sevenspotted ladybird, Coccinella septempunctata - effects of plant-plant odour interaction. Oikos 100: 65-70.). The impact of intercropping colored fiber cotton with cowpea on A. gossypii and A. craccivora populations and on their predator Cycloneda sanguinea (L.) (Coleoptera: Coccinellidae) and the dispersal of these insects among cropping systems were evaluated. The hypothesis were that: a) cotton intercropped with cowpea reduces A. gossypii populations in cotton, and that b) cotton intercropped with cowpea increases C. sanguinea abundance in the cotton crop.
MATERIALS AND METHODS
STUDY LOCATION AND COTTON AND COWPEA CULTIVARS
The study was conducted in a greenhouse at the Entomology Department of Luiz de Queiroz Agriculture College (ESALQ - Escola Superior de Agricultura Luiz de Queiroz/USP), Piracicaba, São Paulo, Brazil. Cotton plants with naturally colored fibers (cv. BRS Safira) and cowpea (cv. BRS Itaim : (upright)) were cultivated in plastic bags with dimensions 40 x 40 x 30 cm, irrigated daily and placed in a greenhouse at 30 °C, 70 ± 10% R.H, and 12 h photophase.
APHIDS AND PREDATORS
Apterous A. gossypii and A. craccivora and one predator species (C. sanguinea) were used. The A. gossypii and C. sanguinea specimens were collected in a cotton field near the Entomology Department at ESALQ - USP, Piracicaba, São Paulo State, Brazil and A. craccivora specimens on cowpea plants in an experimental area of Embrapa Algodão (Embrapa Cotton), Lagoa Seca, Paraíba State, Brazil. Both aphid species were grown on their host plants, i.e. cotton and cowpea, respectively in cages coated with anti-aphid plastic screening at 30 ± 1 °C, 70 ± 10% R.H. and 12 h photophase. The aphids were monitored daily on the plants, and individuals were separated whenever it was necessary by taking into account their life cycle stage (nymphs and adults). The predator was reared on its preys (A. gossypii or A. craccivora) under similar conditions to those used to rear aphids.
BIOASSAYS
Effects of cotton, cowpea and cotton intercropped with cowpea systems on aphid (A. gossypii and A. craccivora) populations
The current study used a randomized block experimental design with four treatments. The bioassay 1 had the following cropping systems: S1- two cotton plants: two cowpea plants within the row- each row began and ended with two cotton plants; S2- two cowpea plants: two cotton plants within the row, alternate rows starting and ending with two cowpea plants or two cotton plants; S3- a cotton row: one cowpea row; and S4- cotton (control) (Fig. 1). Bioassay 2, had the same treatments of bioassay 1, except for S4, which was named S5- cowpea (Fig. 1).
Layout of experimental units in the cotton-cowpea intercropping systems and monoculture. Cotton with colored fibers (closed circles) and cowpea (open circles).
The cotton-cowpea intercropped experimental units had rows with two cotton plants alternating with two cowpea plants within each row or of a cotton row alternating with a cowpea one (Fig. 1). Cotton and cowpea rows were spaced 40 cm from each other in both non-intercropped and intercropped plots. The spacing between plants within each row was 20 cm. The distance between the experimental units was 100 cm (Fig. 1). The experimental units were kept in a transparent plastic cage protected with white voal.
The cotton and cowpea seeds were planted in plastic pots (40 x 40 x 30 cm) and the plants were irrigated every other day.
Thirty-five days after plant emergence or at the end of the vegetative stage, 15 apterous aphids were released in a central plant randomly taken from cotton (S4) or cowpea (S5) plots (Fig. 1) in the non-intercropping systems. Thirty apterous aphids were released in the intercropping system plots (S1, S2 and S3) (Fig. 1), 15 A. gossypii and 15 A. craccivora, respectively.
Ladybugs dispersion and progenies in cotton, cowpea and cotton-cowpea intercropped systems
Three days after releasing the aphids (bioassays 1 and 2), 38 ladybug couples were marked (with corrective ink of different colors) and released in the crops. They were distributed in equal numbers in 15 distinct points, one in each plot. The plant with ladybug release differed from those with the aphids release within each plot.
Five colors were used to mark the forewing of each predator according to the treatment, i.e., a randomized block experimental design with five treatments (crop systems) (colors): black (S1), white (S2), red (S3), green (S4) and blue (S5). The distinction between blocks was done according to the scores each insect received. In the S1 cropping system, for instance, insects marked with black color scored 1, 2 and 3 and they were released in blocks I, II and III, respectively. Similar procedure was used for the other colors.
The dispersion of predators in the systems with only colored cotton (S4) and cowpea (S5) and in colored cotton-cowpea intercropped systems (S1, S2 and S3) was evaluated at three, 12 and 24 h after its release. All individuals captured were quantified and again released in three and 12 h evaluation intervals. Twenty-four hours after they were released, all the ladybugs were recaptured and discarded, except for the eggs deposited by couples during the movement bioassay. The aphids and the predator descendants remained on the plants. Other individuals of the predator were released after the first flowers emerged in the cowpea plant and after the first squares emerged in the cotton plant (approximately 65 days after the plant emergence). Adult predators marking, evaluation and disposal procedures were the same as those used in the vegetative stage.
Forty-nine days after the aphids were released (A. gossypii and A. craccivora) and 42 days after the adult predators were released and recaptured, the same-stage descendants were respectively quantified on 25 and 18 plants per plot.
Data analysis
The C. sanguinea records in all cropping systems were analyzed using PROC FREQ (Sas Institute 2006). The proportions of insects found in the different cropping systems were subjected to the Pearson χ2 test (Sas Institute 2006) to investigate the cropping system dependence for the predator dispersal in non-intercropped or in intercropped crops. The aphid and predator progenies and the cotton and cowpea plants’ reproductive parameters showed normality (Shapiro-Wilk test) and homogeneity (Hartley test). The data were subjected to variance analysis (ANOVA) and the means compared using the Student-Newman-Keuls test (SNK, P = 0.05).
Results
The number of apterous (F(3, 2) = 11.01; P = 0.0075) and alate (F(3, 2) = 5.49; P = 0.0372) A. gossypii found per plant differed among cropping systems (Table I). The highest number of aphids per cotton plant was recorded in the solid cropping system (S4) (Fig. 1) (799 apterous and 136 alates) than in the intercropped systems (S1) (Fig. 1) (545 apterous and 69 alates) (Fig. 2) and (S2) (Fig. 1) (461 apterous and 62 alates) (Fig. 2). However, the number of aphids per cotton plant quantified in the cotton-cowpea intercropped cropping system (S3) (Fig. 1) (656 apterous and 88 alates) (Fig. 2) was similar to that of the solid cotton system (S4) (Fig. 1) (798 apterous and 136 alates) (Fig. 2). The number of apterous and alate aphids (A. gossypii) per cotton plant was 1.46 and 1.73 or 1.97 and 2.19 times higher in the solid cotton system (S4) (Fig. 1) than in the cotton-cowpea intercropped systems (S1) and (S2) (Fig. 1). On the other hand, the cotton-cowpea intercropped systems (S1 and S2) reduced, respectively, in 43% and 31% the number of apterous A. gossypii per cotton plant (Fig. 3a). However, the reduction in the number of alate A. gossypii per cotton plant by the cotton-cowpea intercropped systems (S1 and S2) were, respectively, 53% and 47% (Fig. 3b). This indicates that the cotton-cowpea intercropped systems (S1 and S2) (Fig. 1) may be used to reduce A. gossypii populations.
Summarized model of the one-way analyses of variance (ANOVA) for the effects of crop systems (cotton or cowpea and cotton intercropped with cowpea) on the populations of apterous and alate Aphis gossypii and Aphis craccivora (Hemiptera: Aphididae).
Mean numbers (±SE) of apterous (a) and alate (b) Aphis gossypii (Hemiptera: Aphididae) in the cropping systems of cotton (S4) and cotton intercropped with cowpea systems (S1, S2, and S3). Each column represents the mean number of insects found in 25 plants. Error bars indicate SE. Within each figure, different letters indicate differences (SNK test; P = 0.05).
Mean reduction (±SE) (%) of apterous (a) and alate (b) Aphis gossypii (Hemiptera: Aphididae) in the cropping systems of cotton (S4) and cotton intercropped with cowpea systems (S1, S2, and S3). Error bars indicate SE. Within each figure, different letters indicate differences (SNK test; P = 0.05).
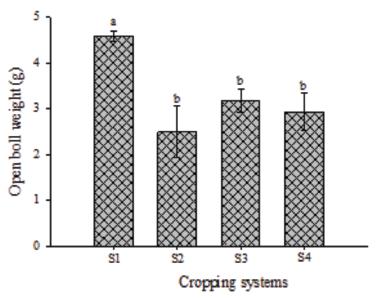
The cropping systems did not affect the mean number of cotton bolls per plant (F(3,2) = 4.46; P = 0.0569). However, they affected the mean weight of open bolls per plant (F(3,2) = 16; P = 0.0029). The S1 was the cropping system with the highest open boll weight (4.59 g) which was lowest in the S2 (2.49 g), S3 (3.19 g) and S4 (2.93 g) (Fig. 4). This indicates that cotton-cowpea intercropped system may be a good tactic to produce heavier open bolls and, therefore, it may help increase cotton yield.
Mean weight (±SE) open cotton bolls in the cropping systems of cotton (S4) or cotton intercropped with cowpea systems (S1, S2, and S3) infested by Aphis gossypii (Hemiptera: Aphididae). Each column represents the mean weight of cotton open bolls harvested in 12 plants. Error bars indicate SE. Different letters indicate differences (SNK test; P = 0.05).
The mean number of apterous (F(3,2) = 3.22; P = 0.1040) and alate (F(3, 2) = 2.05; P = 0.2088) A. craccivora per plant (Table I) and weight of seed/plant (F(3,2) = 0.35, P = 0.7897) showed that the cowpea did no affect these parameters. It indicates that the colored fiber cotton-cowpea intercropping systems did not affect the mean number of A. craccivora and the seed production per cowpea plant.
The predator movement in the vegetative and reproductive stages in all cropping systems showed that its dispersion occurred independently in the cotton-cowpea intercropped systems and in the solid cowpea system (Table II). On the other hand, the predator dispersion in the solid cotton system differed among crop phonological stages (Table II). The predator dispersion in the solid cotton system depended on the crop vegetative stage (χ2 (df=2) = 6.33; P = 0.0421); whereas its dispersion in the solid cotton system reproductive stage was independent (χ2 (df=2) = 0.30; P = 0.8623). Therefore, the predator dispersion in the cotton crop may vary depending on the cropping system and on its phenological stage.
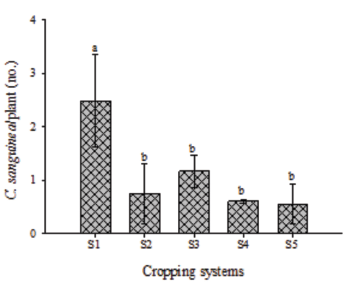
The number of offspring from predators released differed between cropping systems (F(4, 2) = 4.11; P = 0.0317). The mean number of C. sanguinea recovered per plant was higher in the S1 cropping system (2.48 individuals) than in the S2 (0.75 individuals), S3 (1.17 individuals), S4 (0.61 individuals), and S5 (0.56 individuals) cropping systems (Fig. 4). This indicates the S1 cropping system as the most suitable for this predator, probably due to better conditions and attraction to host plants.
Discussion
The intercropping of different plant species is an important cultural practice commonly used in pest management; it is based on the principle that the reduction of insect pest populations in the crop may occur due to increased agro-ecosystem diversity (Risch 2005Risch SJ. 2005. Intercropping as cultural pest control: prospects and limitations. Environ Entomol 7: 9-14.). Diversification practices, such as cotton intercropped with other crops, reduce damages by the insect pests (Ramalho et al. 2012aRamalho FS, Fernandes FS, Nascimento ARB, Nascimento Júnior JL, Malaquias JB AND Silva CAD. 2012a. Assessment of fennel aphids (Hemiptera: Aphididae) and their predators in fennel intercropped with cotton with colored fibers. J Econ Entomol 105: 113-119., b).
The implementing of cotton-cowpea intercropped system in the S1 scheme (Fig. 1) reduced A. gossypii infestation (Fig. 2, Fig. 3a, Fig. 3b) and favored multiplication of the C. sanguinea predator (Fig. 5), and consequently allowed obtaining the highest open boll weight (Fig. 4). The microclimate and plant dispositions in the plots helped to increase natural enemies population by better food and shelter and more adequate microclimate (Cividanes and Yamamoto 2002Cividanes FJ AND Yamamoto FT. 2002. Pragas e inimigos naturais na soja e no milho cultivados em sistemas diversificados. Sci Agr 59: 683-687., Barbosa et al. 2011Barbosa FS, Aguiar-Menezes EL, Arruda LN, Santos CLR AND Pereira MB. 2011. Potencial das flores na otimização do controle biológico de pragas para uma agricultura sustentável. Rev Bras Agroecol 6: 101-110., Manjula and Lakshmi 2014Manjula K AND Lakshmi RP. 2014. Positive influence of intercrops on coccinellid and spider fauna in groundnut. Int J Pl An Env Sci 4: 207-211.). Similar results were reported by Ramalho et al. (2012aRamalho FS, Fernandes FS, Nascimento ARB, Nascimento Júnior JL, Malaquias JB AND Silva CAD. 2012a. Assessment of fennel aphids (Hemiptera: Aphididae) and their predators in fennel intercropped with cotton with colored fibers. J Econ Entomol 105: 113-119., b), Fernandes et al. (2013Fernandes FS, Ramalho FS, Godoy WAC, Pachu JKS, Nascimento ARB, Malaquias JB AND Zanuncio JC. 2013. Within plant distribution and dynamics of Hyadaphis foeniculi (Hemiptera: Aphididae) in field fennel intercropped with naturally coloured cotton. Fla Entomol 96: 92-103.), and Mitiku et al. (2014Mitiku A, Chala A AND Beyene Y. 2014. Effect of intercropping on aphid vectors and yield of pepper (Capsicum annum L.) in southern part of Ethiopia. Int J Technol Enhanc Emerg Eng Res 2: 24-34. ), which reported the importance of using cotton cropping systems intercropped with other plant species to reduce the number of aphids in agricultural ecosystems. Intercrops like cotton with colored fibers and cowpea recorded higher populations of the predator C. sanguinea. This is an associational resistence, refering to the reduced herbivore attack experienced by a plant when they associated with taxonomically different species (Kaitaniemi and Piihimaki 2007Kaitaniemi P AND Piihimaki J. 2007. Experimental evidence for associational resistance against the European pine sawfly in mixed tree stands. Silva Fenn J 41: 259-268. ), and it may be true for the intercropping systems. Associational resistance is due to resource concentration, the natural enemy hypothesis, or both (Ramalho et al. 2012a, b).

Mean number (±SE) of Cycloneda sanguinea (Coleoptera: Coccinellidae) per cotton or cowpea plant on the cropping systems of cotton (S4) or cowpea (S5) and cotton intercropped with cowpea systems (S1, S2, and S3). Each column represents the mean number of C. sanguinea on 18 plants. Error bars indicate SE. Different letters indicate differences (SNK test; P = 0.05).
Choosing the intercropping system is very important to increase the density of natural enemies in the agro-ecosystem and to reduce the damages by aphids (Ramalho et al. 2012bRamalho FS, Fernandes FS, Nascimento ARB, Nascimento Júnior JL, Malaquias JB AND Silva CAD. 2012b. Feeding damage from cotton aphids, Aphis gossypii Glover (Hemiptera: Heteroptera: Aphididae), in cotton with colored fiber intercropped with fennel. Ann Entomol Soc Am 105: 20-27.). Cotton seed production was satisfactory when cotton was intercropped with sesame (Sesamum orientale L.) in the cultivation ratios of 3:1 and 2:1 (1148 kg ha-1 and 993 kg ha-1, respectively) (Aladakatti et al. 2011Aladakatti YR, Hallikeri SS, Nandagavi RH, Hugar AY AND Naveen NE. 2011. Effect of intercropping of oilseed crops on growth, yield and economics of cotton (Gossypium hirsutum) under rainfed conditions. Karnataka J Agric Sci 24: 280-282.). However, higher cotton production per plant was obtained with cotton-cassia (Cassia angustifolia L.) intercropped system (1094 kg ha-1) than with solid cotton system (687 kg ha-1) (Rathod et al. 2011Rathod PS, Biradar DP, Chimmad VP, Mantur SM, Patil VC AND Reddy SCV. 2011. Economic feasibility of senna (Cassia angustifolia Vahl) intercropping with cotton, pigeonpea and castor at different row proportions in dry land situations. Karnataka J Agric Sci 24: 444-447.).
The predator C. sanguinea movements in the vegetative and reproductive stages of the plants in all cropping systems showed dispersion in the cotton-cowpea intercropped systems and in the cowpea system independently. Predator movement between two habitats might be bidirectional, despite the approach involving its dispersion in the crops (Bastola et al. 2014Bastola A, Parajulee MN, Porter RP, Shrestha RB, Chen FJ AND Carroll SC. 2014. Intercrop movement of convergent lady beetle, Hippodamia convergens (Coleoptera: Coccinellidae), between adjacent cotton and alfalfa. Insect Sci 00: 1-12.). However, the literature shows no reports of similar crop settings to those used in the current study. When new crop combinations are set in a particular area, they may provide benefits to the agricultural system since the natural enemy populations found in adjacent areas can quickly colonize these new habitats (Togni 2014Togni PHB. 2014. Habitat manipulation for conservation biological control in organic vegetable crops. Doctor Thesis, UFV, Viçosa, MG. ). Intercropping systems with pest hosts and-non host plants may enable pest movements to new habitats (Togni 2014). This increases the pest vulnerability to natural enemies and the opportunity for predators to find and predate them (Straub et al. 2014Straub CR, Simasek NP, Dohm R, Gapinski MR, Aikens EO AND Nagy O. 2014. Plant diversity increases herbivore movement and vulnerability to predation. Basic Appl Ecol 15: 50-58.). On the other hand, predator actions may increase the pest dispersion in agricultural crops (Otsuki and Yano 2014Otsuki H And Yano S. 2014. Predation risk increases dispersal distance in prey. Naturwissenschaften 101: 513-516.).
The C. sanguinea dispersion showed that this predator has high probability to move to several intercropping habitats, i.e., cotton-cowpea intercropped systems (S1, S2, and S3) and cowpea system (S5). Cotton-cowpea intercropped systems (S1, S2, and S3) increased predator chances of finding food resources, i.e., prey besides colonization because prey will be more vulnerable in these conditions. Therefore, understanding the dispersion of predators can benefit insect conservation programs in agricultural habitats and its crucial for pest management in complex agro-ecosystems (Chailleux et al. 2014Chailleux A, Mohl EK, Alves MT, Messelink GJ AND Desneux N. 2014. Natural enemy-mediated indirect interactions among prey species: potential for enhancing biocontrol services in agroecosystems. Pest Manag Sci 70: 1769-1779., Choate and Lundgren 2014Choate BA AND Lundgren JG. 2014. Protein-marking-based assessment of infield predator dispersal. Biocontrol Sci Techn 24: 1183-1187.).
Natural enemy affect aphid populations by killing or inducing their dispersion (Duarte et al. 2014Duarte L, Pacheco R, Quiñones M, Martínez MA AND Bueno VHP. 2014. Nesidiocoris tenuis Reuter (Hemiptera: Miridae) and Cycloneda sanguinea limbifer (Casey) (Coleoptera: Coccinellidae): behaviour and predatory activity on Myzus persicae Zulzer (Hemiptera: Aphididae). Rev Protec Veg 29: 99-105.). This probably occurred with A. gossypii in cotton-cowpea intercropped systems (S1 and S2) (Fig. 1, Fig. 3a and Fig. 3b), but not with A. craccivora, because this last aphid had similar densities in cowpea plants in cotton-cowpea intercropped systems (S1, S2 and S3) and in solid cowpea system (S5) (Table I).
The cropping systems tested did not affect the weight of seeds/cowpea plant (F(3,2) = 0.35, P = 0.7897). However, they benefited cotton plants, especially by attracting C. sanguinea (Table II and Fig. 5) as reported for this predator in Brassica oleracea (L.) cultivars (Azeredo et al. 2004Azeredo EH, Perruso JC, Menezes EB AND Cassino PCR. 2004. Utilização de Brassica oleracea (L.) como planta atrativa simultânea de Myzus persicae (Sulzei, 1776) e Cycloneda sanguinea (Linnaeus, 1763) em área de cultivo de batata (Solanum tuberosum L.). Rev Univ Rural 24: 89-95.) and the presence of this predator might be linked to specific genotypes.
Plant odor attracts natural enemies (Boullis et al. 2015Boullis A, Francis F AND Verheggen FJ. 2015. Climate change and tritrophic interactions: will modifications to greenhouse gas emissions increase the vulnerability of herbivorous insects to natural enemies? Environ Entomol 10: 1-15.). Ladybugs can learn to associate the odors of plants infested by different aphid species even under restricted environmental condictions (Glinwood et al. 2011Glinwood R, Ahmed E, Qvarfordt E AND Ninkovic V. 2011. Olfactory learning of plant genotypes by a polyphagous insect predator. Oecologia 166: 637-647.). This behavior by ladybugs may explain why C. sanguinea was attracted to the cotton-cowpea intercropped systems.
We conclude that implementing cotton-cowpea intercropped system in the S1 scheme reduces A. gossypii infestation, favors the multiplication of C. sanguinea predator, and allows the obtaining of heavier open bolls.
Acknowledgments
Support for this research was provided by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP).
References
- Aladakatti YR, Hallikeri SS, Nandagavi RH, Hugar AY AND Naveen NE. 2011. Effect of intercropping of oilseed crops on growth, yield and economics of cotton (Gossypium hirsutum) under rainfed conditions. Karnataka J Agric Sci 24: 280-282.
- Almeida RP. 2001. Effect of the population levels of Aphis gossypii on cotton agronomic traits and fibre quality. Proc Sec Exp and Appl Entomol 12: 97-100.
- Azeredo EH, Perruso JC, Menezes EB AND Cassino PCR. 2004. Utilização de Brassica oleracea (L.) como planta atrativa simultânea de Myzus persicae (Sulzei, 1776) e Cycloneda sanguinea (Linnaeus, 1763) em área de cultivo de batata (Solanum tuberosum L.). Rev Univ Rural 24: 89-95.
- Bachmann AC, Nault BA AND Fleischer SJ. 2014. Alate aphid (Hemiptera : Aphididae) species composition and richness in northeastern USA snap beans and an update to historical lists. Fla Entomol 97: 979-994.
- Barbosa FS, Aguiar-Menezes EL, Arruda LN, Santos CLR AND Pereira MB. 2011. Potencial das flores na otimização do controle biológico de pragas para uma agricultura sustentável. Rev Bras Agroecol 6: 101-110.
- Bastola A, Parajulee MN, Porter RP, Shrestha RB, Chen FJ AND Carroll SC. 2014. Intercrop movement of convergent lady beetle, Hippodamia convergens (Coleoptera: Coccinellidae), between adjacent cotton and alfalfa. Insect Sci 00: 1-12.
- Berberet RC, Giles KL, Zarrabi AA AND Payton ME. 2009. Development, reproduction, and within-plant infestation patterns of Aphis craccivora (Homoptera: Aphididae) on alfafa. Environ Entomol 38: 1765-1771.
- Berlandier FA AND Sweetingham MW. 2003. Aphid feeding damage causes large losses in susceptible lupin cultivars. Aust J Exp Agr 43: 1357-1362.
- Boeye J, Kubisch A AND Bonte D. 2014. Habitat structure mediates spatial segregation and therefore coexistence. Landscape Ecol 29: 593-604.
- Boullis A, Francis F AND Verheggen FJ. 2015. Climate change and tritrophic interactions: will modifications to greenhouse gas emissions increase the vulnerability of herbivorous insects to natural enemies? Environ Entomol 10: 1-15.
- BRAGA YFB, GRANGEIRO TB, FREIRE EA, LOPES HL, BEZERRA JNS, ANDRADE-NETO M AND LIMA MAS. 2007. Insecticidal activity of 2-tridecanone against the cowpea weevil Callosobruchus maculatus (Coleoptera: Bruchidae). An Acad Bras Cienc 79: 35-39.
- Chailleux A, Mohl EK, Alves MT, Messelink GJ AND Desneux N. 2014. Natural enemy-mediated indirect interactions among prey species: potential for enhancing biocontrol services in agroecosystems. Pest Manag Sci 70: 1769-1779.
- Choate BA AND Lundgren JG. 2014. Protein-marking-based assessment of infield predator dispersal. Biocontrol Sci Techn 24: 1183-1187.
- Cividanes FJ AND Yamamoto FT. 2002. Pragas e inimigos naturais na soja e no milho cultivados em sistemas diversificados. Sci Agr 59: 683-687.
- Duarte L, Pacheco R, Quiñones M, Martínez MA AND Bueno VHP. 2014. Nesidiocoris tenuis Reuter (Hemiptera: Miridae) and Cycloneda sanguinea limbifer (Casey) (Coleoptera: Coccinellidae): behaviour and predatory activity on Myzus persicae Zulzer (Hemiptera: Aphididae). Rev Protec Veg 29: 99-105.
- Fernandes FS, Ramalho FS, Godoy WAC, Pachu JKS, Nascimento ARB, Malaquias JB AND Zanuncio JC. 2013. Within plant distribution and dynamics of Hyadaphis foeniculi (Hemiptera: Aphididae) in field fennel intercropped with naturally coloured cotton. Fla Entomol 96: 92-103.
- Fernandes FS, Ramalho FS, MALAQUIAS JB, Nascimento JUNIOR JL, CORREIA ET AND Zanuncio JC. 2012c. Within-plant distribution of cotton aphid (Hemiptera: Aphididae) in cotton cultivars with colored fibers. An Acad Bras Cienc 84: 707-719.
- FERNANDES FS, RAMALHO FS, NASCIMENTO ARB, MALAQUIAS JB, SILVA CAD AND ZANUNCIO JC. 2012a. Within-plant distribution of cotton aphid (Hemiptera: Aphididae), in cotton with colored fibers and cotton-fennel intercropping system. Ann Entomol Soc Am 105: 599-607.
- FERNANDES FS, RAMALHO FS, NASCIMENTO JL, MALAQUIAS JL, NASCIMENTO ARB, SILVA CAD AND ZANUNCIO JC. 2012b. Within-plant distribution of cotton aphids, Aphis gossypii Glover (Hemiptera: Aphididae), in Bt and non-Bt cotton fields. Bul Entomol Res 102: 79-87.
- Frota KMG, Soares RAM AND Arêas JAG. 2008. Composição química do feijão caupi (Vigna unguiculata L. Walp ), cultivar BRS-Milênio. Food Sci Technol 28: 470-476.
- Glinwood R, Ahmed E, Qvarfordt E AND Ninkovic V. 2011. Olfactory learning of plant genotypes by a polyphagous insect predator. Oecologia 166: 637-647.
- Gonzaga JV, Ramalho FS AND Santos JW. 1991. Distribuição do Aphis gossypii no algodoeiro nos sistemas de plantio solteiro e consorciado. Pesqui Agropecu Bras 26: 1839-1844.
- Horstmann G. 1995. Dyeing as a new environmental challenge. J Soc Dyers Colour 111: 182-184.
- Kadam D, Kadam D AND Lekurwale R. 2014. Seasonal incidence of sucking pests of Bt cotton in different intercropping systems and their natural enemies. J Plant Pest Sci 1: 29-34.
- Kaitaniemi P AND Piihimaki J. 2007. Experimental evidence for associational resistance against the European pine sawfly in mixed tree stands. Silva Fenn J 41: 259-268.
- Kitajima EW, Alcântara BK, Madureira PM, Alfenas-Zerbini P, Rezende JAM AND Zerbini FM. 2008. A mosaic of beach bean (Canavalia rosea) caused by an isolate of cowpea aphid-borne mosaic virus (CABMV). Arch Virol 153: 743-747.
- Laamari M, Khelfa L AND Coer d’Acier A. 2008. Resistance source to cowpea aphid (Aphis craccivora Koch) in broad bean (Vicia faba L.) Algerian landrace collection. Afr J Biotechnol 7: 2486-2490.
- LEITE GLD, PICANÇO M, ZANUNCIO JC AND ECOLE CC. 2006. Factors affecting herbivory of Thrips palmi (Thysanoptera: Thripidae) and Aphis gossypii (Homoptera: Aphididae) on the eggplant (Solanum melongena). Braz Arch Biol Techn 49: 361-369.
- LEITE GLD, PICANÇO M, ZANUNCIO JC AND GUSMÃO MR. 2007. Factors affecting colonization and abundance of Aphis gossypii Glover (Homoptera: Aphididae) on okra plantations. Cienc Agrotec 31(2): 337-343.
- Leite MIS, Carvalho GA, Maia JB, Makiyama L AND Vilela M. 2010. Ação residual de inseticidas para larvas e adultos do predador Cycloneda sanguinea Linnaeus, 1763 (Coleoptera: Coccinellidae ). Arq Inst Biol 77: 275-282.
- Lima JAA, Oliveira FMEW, Kitajima EW AND Lima MGA. 1981. Propriedades biológicas sorológicas, citológicas e sorológicas de um potyvirus isolado de feijão-de-corda no Ceará. Fitopatol Bras 6: 205-216.
- Malaquias JB, Ramalho FS, Lira ACS, Oliveira FQ, Fernandes FS, Godoy WAC AND Zanuncio JC. 2014. The biology and thermal requirements of the fennel aphid Hyadaphis foeniculi (Passerini) (Hemiptera: Aphididae). PLoS ONE 9(7):e100983.
- Manjula K AND Lakshmi RP. 2014. Positive influence of intercrops on coccinellid and spider fauna in groundnut. Int J Pl An Env Sci 4: 207-211.
- Medeiros MA, Sujii ER AND Morais HC. 2009. Effect of plant diversification on abundance of South American tomato pinworm and predators in two cropping systems. Hort Bras 27: 300-306.
- Mitiku A, Chala A AND Beyene Y. 2014. Effect of intercropping on aphid vectors and yield of pepper (Capsicum annum L.) in southern part of Ethiopia. Int J Technol Enhanc Emerg Eng Res 2: 24-34.
- Ninkovic V AND Pettersson J. 2003. Searching behaviour of the sevenspotted ladybird, Coccinella septempunctata - effects of plant-plant odour interaction. Oikos 100: 65-70.
- Obopile M. 2006. Economic threshold and injury levels for control of cowpea aphid, Aphis crassivora Linnaeus (Homoptera: Aphididae), on cowpea. Afr Plant Prot 12: 111-115.
- Otsuki H And Yano S. 2014. Predation risk increases dispersal distance in prey. Naturwissenschaften 101: 513-516.
- Ramalho FS, Fernandes FS, Nascimento ARB, Nascimento Júnior JL, Malaquias JB AND Silva CAD. 2012a. Assessment of fennel aphids (Hemiptera: Aphididae) and their predators in fennel intercropped with cotton with colored fibers. J Econ Entomol 105: 113-119.
- Ramalho FS, Fernandes FS, Nascimento ARB, Nascimento Júnior JL, Malaquias JB AND Silva CAD. 2012b. Feeding damage from cotton aphids, Aphis gossypii Glover (Hemiptera: Heteroptera: Aphididae), in cotton with colored fiber intercropped with fennel. Ann Entomol Soc Am 105: 20-27.
- RAMALHO FS, MALAQUIAS JB, BRITO BDS, FERNANDES FS AND ZANUNCIO JC. 2015. Assessment of the attack of Hyadaphis foeniculi (Passerini) (Hemiptera: Aphididae) on biomass, seed and oil in fennel intercropped with cotton with colored fibers. Ind Crops Prod 77: 511-515.
- Rathod PS, Biradar DP, Chimmad VP, Mantur SM, Patil VC AND Reddy SCV. 2011. Economic feasibility of senna (Cassia angustifolia Vahl) intercropping with cotton, pigeonpea and castor at different row proportions in dry land situations. Karnataka J Agric Sci 24: 444-447.
- Resende ALS, Haro MM, Silva VF, Souza B AND Silveira LCP. 2012. Diversidade de predadores em coentro, endro e funcho sob manejo orgânico. Arq Inst Biol 79: 193-199.
- Risch SJ. 2005. Intercropping as cultural pest control: prospects and limitations. Environ Entomol 7: 9-14.
- Sarwar MK, Azam I, Iram N, Iqbal W, Rashda A, Anwer F, Atta K AND Ali R. 2014. Cotton aphid Aphis gossypii L. (Homoptera; Aphididae); a challenging pest; biology and control strategies: a review. Int J Appl Biol Pharm Technol 5: 288-294.
- SAS Institute. 2006. Sas/Stat user’s guide. Sas Institute, Cary, NC.
- Silva PHS, Carneiro JS AND Quinderé MAW. 2005. Pragas. In: Freire Filho FR, Lima JAA and Ribeiro VQ (Eds), Feijão-caupi: avanços tecnológicos. Brasília: Embrapa Informação Tecnológica, Teresina: Embrapa Meio Norte, p. 369-402.
- Straub CR, Simasek NP, Dohm R, Gapinski MR, Aikens EO AND Nagy O. 2014. Plant diversity increases herbivore movement and vulnerability to predation. Basic Appl Ecol 15: 50-58.
- Togni PHB. 2014. Habitat manipulation for conservation biological control in organic vegetable crops. Doctor Thesis, UFV, Viçosa, MG.
- Ullah K AND Paul P. 1985. Chemical control of cotton aphid. Pakistan J Agric Res 6: 213-215.
Publication Dates
-
Publication in this collection
01 Feb 2018 -
Date of issue
Jan-Mar 2018
History
-
Received
14 Apr 2016 -
Accepted
26 May 2016