Abstract
The appropriate choice of tree species and planting arrangement for use in agroforestry systems is very important in order to maximize productivity and sustainability. The aim of this study was to evaluate the growth of five tree species and its effect on sugarcane yields in different planting arrays in agroforestry systems. The study was conducted from 2007 to 2011 in the city of Frederico Westphalen, RS, Brazil. The five studied tree species were: guapuruvu (Schizolobium parahybae), bracatinga (Mimosa scabrella), canafístula (Peltophorum dubium), angico-vermelho (Parapiptadenia rigida) and eucalypt (Eucalyptus urophylla S.T. Blake x Eucalyptus grandis Hill ex Maiden). Root collar diameter, diameter at breast height, plant height and mean diameter of crown were evaluated for four years with an interval of 90 days between assessments. Yield data for sugarcane were collected in June 2009, 2010, and 2011. Eucalypt presented the greatest root collar diameter values, diameter at breast height, crown development, and plant height. Angico-vermelho, canafístula and eucalypt presented a higher growth in the strip system, while guapuruvu presented the greater growth in the line system. Bracatinga was not influenced by an arrangement in the agroforestry systems. Sugarcane presented greater production in the understory of angico and bracatinga.
Key words
array of plants; competition; crown diameter; height
INTRODUCTION
Agroforestry systems consist of integrated land use for forestry, crops, and/or animal purposes. This integration often results in socio-economic, environmental and production benefits such as the recovery of degraded areas, reduction of production costs, and an increase input of organic matter which has been shown to improve the chemical, physical and biological properties of the soil (Tracy and Zhang 2008TRACY BF and ZHANG Y. 2008. Soil compaction, corn yield response, and soil nutrient pool dynamics within an integrated crop-livestock system in Illinois. Crop Sci 48: 1211-1218., Neves et al. 2009NEVES CMN, SILVA MLN, CURI N, MACEDO RLG, MOREIRA FDS and D’ANDRÉA AF. 2009. Indicadores biológicos da qualidade do solo em sistema agrossilvipastoril no noroeste do Estado de Minas Gerais. Ciên Agrotec 33: 105-112., Xavier et al. 2014XAVIER DF, LÉDO FJS, PACIULLO DSC, URQUIAGA S, ALVES BJR and BODDEY RM. 2014. Nitrogen cycling in a Brachiaria-based silvopastoral system in the Atlantic forest region of Minas Gerais, Brazil. Nutr Cycl Agroecosyst 99: 45-62.).
To analyze the sustainability of an agroforestry system, it is important to understand factors and interactions between all species within it (Berlyn and Cho 2000BERLYN GP and CHO J. 2000. Light, moisture and nutrient use by plants. In: Ashton MS and Montagnini F (Eds), The silvicultural basis for agroforestry systems. CRC Press, Boca Raton, p. 9-39., Ong et al. 2000ONG CK, BLACK CR, WALLACE JS, KHAN AAH, LOTT JE, JACKSON NA, HOWARD SB and SMITH DM. 2000. Productivity, microclimate and water use in Grevillea Robusta-based agroforestry systems on hill slopes in semi-arid Kenya agriculture. Agric Ecosyst Environ 80: 121-141.), and therefore the choice of species which compose a system is a consideration of great importance (Bayala and Wallace 2015BAYALA J and WALLACE JS. 2015. The water balance of mixed tree crop systems. In: Ong CK, Black C and Wilson J (Eds), Tree-crop interactions, 2nd edition: agroforestry in a changing climate. CAB International, Wallingford, p. 146-190., Ong and Kho 2015ONG CK and KHO RM. 2015. A framework for quantifying the various effects of tree-crop interactions. In: Ong CK, Black C and Wilson J (Eds), Tree-crop interactions, 2nd edition: agroforestry in a changing climate. CAB International, Wallingford, p. 1-23.). The growth and development of different types of plants (e.g. crops and tree species) in the same area presupposes the existence of dynamic system interactions and change over time; this is especially true in areas where there are tree components, due to continued growth in height, crown projection, and leaf area of tree species. These factors can affect the distribution of existing resources which in turn can cause a constant change in the productivity of species in a system (Jose et al. 2004JOSE S, GILLESPIE AR and PALLARDY SG. 2004. Interespecific interactions in temperate agroforestry. Agrofor Syst 61: 237-255.).
The study of the vegetative growth of woody plants in mixed systems is important when considering management practices; for example, the definition of plant spacing and planting arrangements. Similarly, meteorological elements directly influence morphological and physiological characteristics of the trees as well as the nutritional factors which can affect the growth of plants in the understory (Paciullo et al. 2011PACIULLO DS, GOMIDE CA, CASTRO CR, FERNANDES PB, MÜLLER MD, PIRES MF, FERNANDES EM and XAVIER DF. 2011. Características produtivas e nutricionais do pasto em sistema agrossilvipastoril, conforme a distância das árvores. Pesq Agropec Bras 46: 1173-1186., Mendes et al. 2013MENDES MMS, LACERDA CF, CAVALCANTE ACR, FERNANDES FÉP and OLIVEIRA TS. 2013. Desenvolvimento do milho sob influência de árvores de pau branco em sistema agrossilvipastoril. Pesq Agropec Bras 48: 1342-1350.,Machado et al. 2014MACHADO SA, ZAMIN NT, NASCIMENTO RGM and SANTOS AAP. 2014. Efeito de variáveis climáticas no crescimento mensal de Pinus taeda e Araucaria angustifolia em fase juvenil. Floresta Ambient 21: 170-181. ). The viability of sugarcane in agroforestry systems in Brazil has been demonstrated by Pinto et al. (2003)PINTO LFG, BERNARDES MS and SPAROVEK G. 2003. Feasibility of cultivation of sugarcane in agroforestry systems. Sci Agric 60: 489-493. and Elli et al. (2016)ELLI EF, CARON BO, PAULA GM, ELOY E, SCHWERZ F and SCHMIDT D. 2016. Ecofisiologia da cana-de-açúcar no sub-bosque de canafístula em arranjos de sistema agroflorestal. Comunicata Scientiae 7: 464-472.. Several studies in Brazil and around the world demonstrate the viability of using annual crops such as maize and soybeans (Peng et al. 2009PENG X, ZHANG Y, CAI J, JIANG Z and ZHANG S. 2009. Photosynthesis, growth and yield of soybean and maize in a tree-based agroforestry intercropping system on the Loess Plateau. Agrofor Syst 76: 569-577.), wheat (Rivest et al. 2013RIVEST D, LORENTE M, OLIVIER A and MESSIER C. 2013. Soil biochemical properties and microbial resilience in agroforestry systems: effects on wheat growth under controlled drought and flooding conditions. Sci Total Environ 463: 51-60.), oat (Deiss et al. 2016DEISS L, MORAES AD, PELISSARI A, SILVA VPD and ANDREOLLA VRM. 2016. Oat grain yield under nitrogen uses in an eucalyptus intercropping system in Subtropical Brazil. Rev Cienc Agron 47: 462-470.) and ryegrass (Pilau et al. 2015PILAU J, ELLI EF, NARDINO M, KORCELSKI C, SCHMIDT D and CARON BO. 2015. Desenvolvimento e qualidade do azevém no sub-bosque de angico-vermelho em sistema silvipastoril. Comunicata Scientiae 6: 437-444.) in agroforestry systems. There is currently a lack of information regarding tree species and the performance of plants in agroforestry system when placed in different arrangements. Research needs to be conducted in order to examine the interactions between species and crops of interest, which in this case is sugarcane.
In addressing this lack of information, the proposed hypothesis is that tree species react differently depending on their arrangement. In order to test this hypothesis, the aim of this study was to evaluate the growth of the tree species: Schizolobium parahybae (Vell.) Blake (guapuruvu), Mimosa scabrella Benth. (bracatinga), Peltophorum dubium (Spr.) Taubert (canafístula), Parapiptadenia rigida (Benth.) Brenan (angico-vermelho) and Eucalyptus urophylla S.T. Blake x Eucalyptus grandis Hill ex Maiden (eucalypt), and its effect on the sugarcane (Saccharum spp.) yield in different planting arrays in agroforestry systems.
MATERIALS AND METHODS
STUDY AREA
The study was conducted in the city of Frederico Westphalen, RS (27°22’S, 53°25’W) at an altitude of 480 m. According to the Köppen climate classification, the climate of the region is Cfa, i.e. humid subtropical with an average annual temperature of 19.1 °C, varying with maximum 38 °C and minimum of 0 °C, presenting an average annual precipitation of 1606 mm. The soil of the area is classified as a typical Entisol Orthents (Cunha et al. 2011CUNHA NG SILVEIRA RDC, KOESTER E, OLIVEIRA LD, FILIPPINI ALBA JM, TERRES V and LOPES R. 2011. Estudos de Solos do Município de Frederico Westphalen, RS. EMBRAPA, Pelotas, RS. Brazil. (Circular Técnica 116).).
DESIGN AND EXPERIMENTAL UNITS
The experimental design was composed of a complete randomized block, and study objects were characterized by a factorial scheme of 2x5x16, i.e., two planting arrays (strip and line), five tree species (angico, bracatinga, canafístula, eucalypt and guapuruvu) and three blocks were studied. Each species was measured 16 times in total, beginning 159 to 1509 days after planting (DAP) with an interval of 90 days between assessments and three replications. Ten experimental units were included in each block, and each unit was randomly distributed in the agroforestry system.
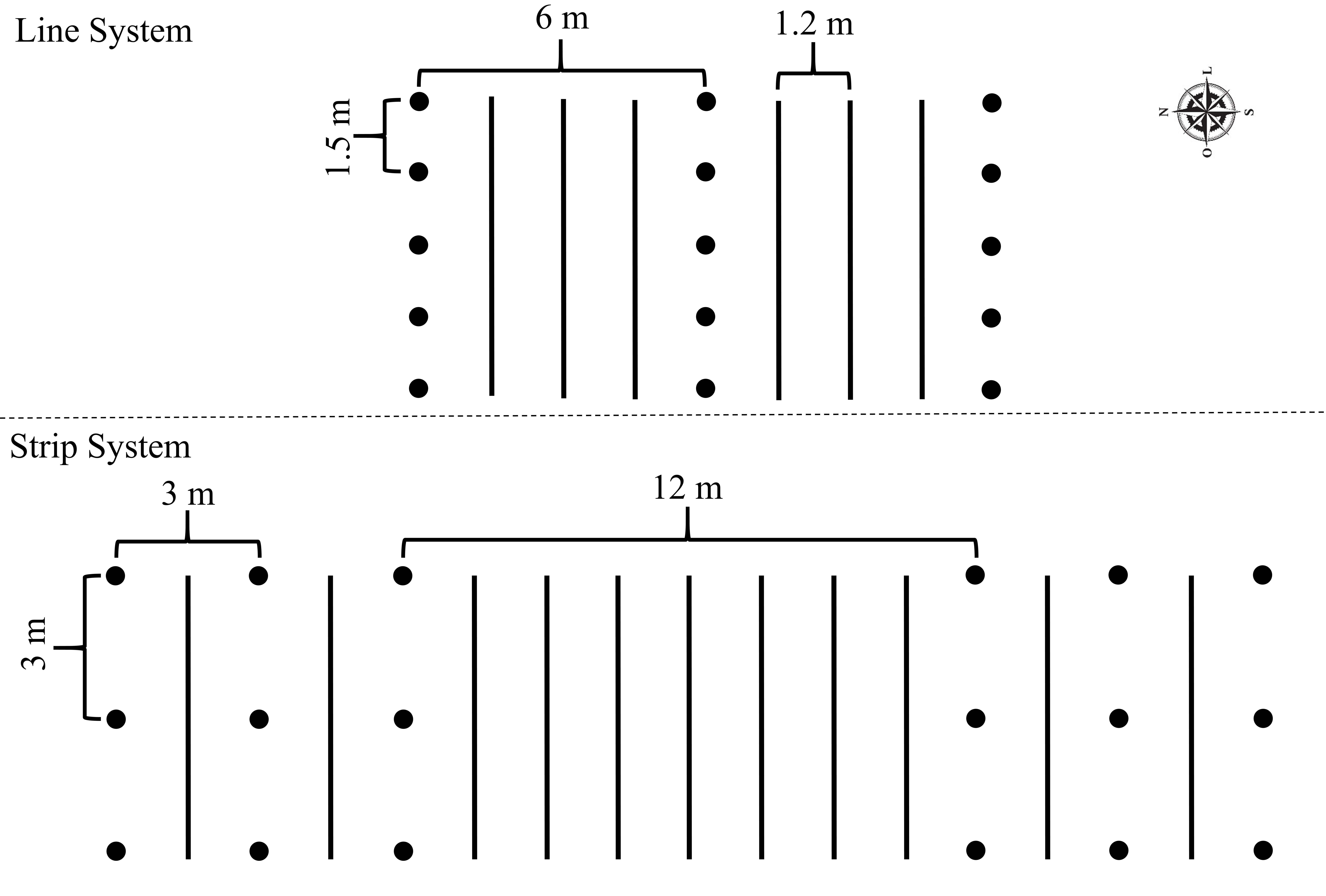
The arrangement of trees and sugarcane are shown in Figure 1. In the strip system, the forest species were divided into separate strips by 12m. Each strip was composed of three rows. Each row was composed of three trees which were spaced at 3x3m. Sugarcane (Saccharum spp.) was distributed in six rows (between strips, within 12 m) and two rows between lines of trees. In the line system, forestry species were distributed with 6x1.5 m spacing or 6 m between lines and 1.5 m between plants in line. Each line was composed of five trees. The sugarcane crops were distributed in three rows (between the lines of trees). The species were arranged in sequence, and not in a mixed distribution. Each block was composed of two systems and the five species, resulting in 165 total trees. In both systems, sugarcane was distributed in a spacing of 1.20 m and was an equal distance from the trees in this spacing. Both rows of sugarcane, as well as trees, were oriented in an East-West direction.
A sketch of an experimental unit of line and strip systems. Black circles represent the trees; continuous lines indicate where the sugarcane was planted. Each species presented three experimental units in each system.
The field experiment began in Sep. 2007 with the manual planting of seedlings after subsoiling and harrowing of the area was completed. Fertilization was then performed according to recommendations for each species (CQFS 2004CQFS - COMISSÃO DE QUÍMICA E FERTILIDADE DO SOLO. 2004. Manual de adubação e calagem para os estados do Rio Grande do Sul e Santa Catarina. Porto Alegre: Sociedade Brasileira de Ciência do Solo, RS, Brasil, 400 p.).
Evaluations began on March 21, 2008, 159 days after planting (DAP) in the early autumn season. From this point on, assessments were performed quarterly at the beginning of each season until the summer of 2011, at 1509 DAP, totaling 16 instances of measurement.
ASSESSED TRAITS
The root collar diameter (RCD) was sampled immediately above the soil surface and diameter at breast height (DBH) measurements were obtained 1.30 m above the soil. Both variables were determined with the aid of a digital caliper until the values reached 6.00 cm, later a graduated diameter tape in millimeters was used to take measurements. The caliper was used to make two orthogonal measures and the arithmetic means of these measurements was recorded.
For measurement of plant height (H), a graduated diameter tape measure was used until the values reached 2.00 m, at which point a Vertex Hypsometer III was used to make measurements greater than 2.00 m. This device was used to determine the distance from the ground surface to the highest leaf axils. The mean crown diameter (MCD) was considered to be the distance between the projection lines of the outermost points of the crown. Data were obtained by taking two orthogonal measurements with the aid of a graduated tape and later calculating the arithmetic mean.
Yield data for the sugarcane was collected in June 2009, 2010 and 2011, constituting the three years of sugarcane evaluation. In each experimental unit, six medium stalks for strip system and eight stalks for line system were collected and taken to the laboratory for evaluation. For the existing population in the experimental units, values were extrapolated for one hectare (ha). Stalk weight (SW, t ha-1) was determined with the aid of a digital scale.
Data for Photosynthetically active radiation (PAR) in the understory of the agroforestry systems were obtained during the harvest of sugarcane over the three years of evaluation, with a quantum sensor LI-190-1 with a spectral strip of 400-700 nm coupled to a porometer dynamic balance LICOR-LI1600 model. The interception of photosynthetically active radiation (IPAR) was determined for canopy tree species, according to the equation used by Caron et al. (2012CARON BO, LAMEGO FP, SOUZA VQ, COSTA EC, ELOY E, BEHLING A and TREVISAN R. 2012. Interceptação de radiação luminosa pelo dossel de espécies florestais e sua relação com o manejo de plantas daninhas. Cienc Rural 42: 75-82.):
Where Rin = photosynthetically active radiation inside the canopy of the tree species; Rabo = photosynthetically active radiation above the canopy of the tree species.
DATA ANALYSIS
The data were statistically analyzed with the software “Statistical Analysis System” (SAS 2003SAS - STATISTICAL ANALYSIS SYSTEM. 2003. Getting started with the SAS Learning Edition. Cary, 200 p.), and results were obtained through an analysis of variance, F test, and Tukey test (p > 0.05). The Bartlett test was used to verify the homogeneity of variances. In the strip system, a test of contrast was conducted between plants of the central and external lines, taking into account differing environmental systems. We plotted graphics demonstrating the variation of sugarcane production and the interception of photosynthetically active radiation of tree species throughout the years. We plotted the growth of tree species against evaluated periods and the mean annual increment (MAI) of the variables for each evaluation year; for the sugarcane, we plotted the yield and the IPAR against harvest years (2009, 2010, 2011).
CHARACTERIZATION OF WEATHER
Values for the climatic factors which occurred during the study were obtained through the weather station of National Institute of Meteorological, located at 1500 m from the study site, at coordinates 27°39’S and 53°43’W. Average daily global solar radiation, average monthly maximum and minimum temperature, rainfall and sequential water balance during the years 2008, 2009, 2010 and 2011 is described in Figure 2. The water balance followed Thornthwaite and Mather’s (1955)THORNTHWAITE CW and MATHER JR. 1955. The Water Balance. Centerton, Drexel Institute of Technology, Laboratory of Climatology, Publications in climatology, 104 p. approach, which has inputs rainfall, references evapotranspiration and water holding capacity. We estimated reference evapotranspiration by Penman-Monteith FAO 56 method (Allen et al. 1998ALLEN RG, PEREIRA LS, RAES D and SMITH M. 1998. Crop evapotranspiration: guidelines for computing crop water requirements. Rome: FAO, 300 p. (FAO Irrigation and drainage paper, 56).) and considered a water holding capacity of 150 mm.
Average daily global solar radiation (a), average monthly maximum and minimum temperature (b), rainfall and sequential water balance (c) during the years 2008, 2009, 2010 and 2011 in Frederico Westphalen, RS, Brazil. Bars indicate the standard deviation for each month.
RESULTS
The flux of global solar radiation during the years of evaluation was 16.70 MJ m-2 day-1 on average, with a variation of 8.7 to 25.5 MJ m-2 day-1 (Figure 2). The average monthly temperature ranged from 8.1 to 31.4 ºC. We observed negative minimum absolute temperatures (-1.54; -2.24; -1.13 and -1.76 in 2008, 2009, 2010 and 2011, respectively), which resulted in the occurrence of frost in all years. Considering water balance, there were periods with water deficits in all years of evaluation. The accumulated annual water deficits were 186.69; 175.11; 28.43 and 93.27 mm in 2008, 2009, 2010 and 2011, respectively.
An analysis of variance revealed a difference in RCD for interaction between agroforestry systems x tree species x DAP, for all variables of tree species x DAP and H, MCD and DBH in the interaction between tree species x agroforestry system (Table I). In the strip system, based on the analysis of contrast made between plants of the central and external lines, no differences were observed, therefore, tree lines were not analyzed separately in the interactions.
Analysis of variance of root collar diameter (RCD), diameter at breast height (DBH), plant height (H) and mean cro wn diameter (MCD) of five tree species evaluated in 16 periods after planting in two agroforestry systems, in the city of Frederico Westphalen – RS.
At 249 DAP, species began to show a stratification of RCD means (Figure 3). This became more evident at 429 DAP due to averages of eucalypt which were greater than all other species in the strip system. In the line system, this trend was observed at 609 DAP, and remained until the last assessment (1509 DAP). Canafístula and guapuruvu presented RCD values greater than bracatinga and angico. This was mainly observed in the strip system at 1059 DAP. At 1509 DAP, these two species had the lowest values in the strip system, but this was only observed for angico in the line system.
Growth and mean annual increment (MAI) of root collar diameter (RCD) of tree species angico (Ang), bracatinga (Brac), canafístula (Can), eucalyptus (Euc) and guapuruvu (Guap) in agroforestry systems strip (a and b) and line (c and d), from 159 to 1509 days after planting (DAP) of the seedlings in Frederico Westphalen - RS.
Comparing agroforestry systems, the superiority of RCD was observed in the strip system for eucalypt from 429 DAP; this difference between systems remained until the last evaluation period. The evolution of RCD forest species studied over time in both agroforestry systems can be seen in Figure 3. In strip system (Figure 3a), one can see the eucalypt species more pronounced rate of growth when compared to other species. In the line system (Figure 3b) this difference was less evident.
Eucalypt showed the greater IMA for all evaluated years in both agroforestry systems (5.85; 6.01; 6.64 and 6.14 cm year-1 in the strip system; 4.51; 4.41; 4.92 and 5.00 cm year-1 in the line system, in the years 2008, 2009, 2010 and 2011, respectively), compared to other species. Comparing the agroforestry systems, strip system provided greater MAI of the angico and eucalypt in all the evaluated years. Bracatinga and guapuruvu had greater MAI of RCD in the line system starting with the second year while canafístula did not show a trend in relation to the agroforestry system for this variable (Figure 3).
In general, the DBH values were higher for eucalypt (Figure 4a), followed by guapuruvu and later by the other three species (angico-vermelho, bracatinga and canafístula). The MAI of DBH were greater for eucalypt, reaching the highest values at three and four years of age (4.18 and 4.17 cm year-1, respectively). Regarding H, eucalypt demonstrated greater values than other species from 339 DAP. This trend continued through the last evaluation (Figure 4b). The MAI of H, on average, was higher for eucalypt, reaching its greatest values at 3 years of age (3.93 m year-1), followed by bracatinga, and then by the other three species.
Growth and mean annual increment (MAI) of diameter at breast height (DBH; a and b), height (H; c and d) and mean crown diameter (MCD; e and f) of tree species angico (Ang), bracatinga (Brac), canafístula (Can), eucalypt (Euc) and guapuruvu (Guap), from 159 to 1509 days after planting in Frederico Westphalen - RS. Vertical arrows indicate damage related to frost.
Higher values were found for the MCD from eucalypt DAP 339, and remained superior until the last evaluation. The MAI of MCD was higher in the first evaluation year for all species. In this period, eucalypt showed the greater MAI (2.23 m year-1), followed by bracatinga (1.15 m year-1), guapuruvu (0.84 m year-1), canafístula (0.79 m year-1) and angico (0.64 m year-1). Except for eucalypt trees, guapuruvu demonstrated higher MCD than the other species; however, likely due to frost damage, this superiority of guapuruvu MCD was not observed in spring months after the occurrence of winter frosts (Figure 4c). Between winter and spring, the reduction of guapuruvu MCD was 1.06; 1.44; 0.81 and 0.61 m for the years 2008, 2009, 2010 and 2011, respectively. One can identify a stagnant growth of bracatinga, observed in respective MCD values, from the DAP 699.
The strip system demonstrated the highest values for all variables for eucalypt, of DBH and MCD for angico, and only DBH for canafístula (Table II). Guapuruvu presented an inverse behavior, where it benefited from the line system and had a greater average DBH and H, while bracatinga showed similar characteristics in both systems for all variables.
Diameter at breast height (DBH), plant height (H) and mean diameter of crown (MCD) of tree species angico, bracatinga, canafístula, eucalypt and guapuruvu in strip and line agroforestry systems in the city of Frederico Westphalen – RS.
The growth of MCD (Table II) of forest species along the time reduced the stalk weight of sugarcane (Figure 5a), due to an increase of intercepted photosynthetically active radiation (IPAR) by tree components (Figure 5b). The stalk weight in the understory of eucalypt was lower from the second evaluation year of sugarcane (2010), compared to the understory of other species. Eucalypt was also responsible for higher IPAR in 2009 and 2011 (56.41 and 89.74%, respectively) and had a lesser amount of available photosynthetically active radiation for the sugarcane. Sugarcane yield and IPAR by the crown of angico were, on average, 178% greater and 54% lower than the sugarcane yield, and the IPAR by the crown of eucalypt.
Stalk weight (a) and Interception of photosynthetically active radiation – IPAR (b), of sugarcane grown in the understory of various tree species from 2009 to 2011, in Frederico Westphalen - RS. Bars indicate the standard deviation for each evaluation year.
DISCUSSION
The superiority of RCD observed in the strip system for eucalypt from 429 DAP and the superiority of the IMA in all evaluation years (Figure 3) is justified by the greater plant spacing of this system (3 x 3 + 12). In the line system, reduced spacing between plants in the row system (1.5 m) leads to competition for water and nutrients (Stape et al. 2010STAPE JL et al. 2010. The Brazil eucalyptus potential productivity project: influence of water, nutrients and stand uniformity on wood production. For Ecol Manage 259: 1684-1694.). The mean annual increments of RCD found in this study are higher than the results found by Nicodemo et al. (2016)NICODEMO MLF, MULLER MD, PORFÍRIO-DA-SILVA V, CARPANEZZI AA, PEZZOPANE JRM and BARIONI JÚNIOR W. 2016. Growth of native trees in two agroforestry systems. Rev Árvore 40: 639-648. who studied the growth of native species in agroforestry systems in Brazil and found a growth rate of 0.10 cm month-1 (1.2 cm year-1) for canafístula at four years of age.
Regarding DBH (Figure 4a), our results are higher than those found by Ferreira et al. (2014)FERREIRA DHAA, SANTOS LELES PS, MACHADO EC, ABREU AHM and ABÍLIO FM. 2014. Crescimento de clone de Eucalyptus urophylla x E. grandis em diferentes espaçamentos. Rev Árvore 44: 431-440., who analyzed the growth of clones Eucalyptus urophylla x Eucalyptus grandis in Avaré/SP, and found a mean DBH of 13.8 cm four years after planting using a spacing of 3.0 x 2.5 m. The highest values found in the present study were on 1509 DAP, four years after transplantation, but smaller than what was observed by Ferreira (2014)FERREIRA DHAA, SANTOS LELES PS, MACHADO EC, ABREU AHM and ABÍLIO FM. 2014. Crescimento de clone de Eucalyptus urophylla x E. grandis em diferentes espaçamentos. Rev Árvore 44: 431-440.. The differences in RCD and DBH can be explained by different climate conditions between locations and the spatial distribution of species in the field (Nascimento et al. 2012NASCIMENTO DF, LELES PSDS, OLIVEIRA NETO SND, MOREIRA RTS and ALONSO JM. 2012. Crescimento inicial de seis espécies florestais em diferentes espaçamentos. Cerne 18: 59-165.). Oliveira et al. (2015OLIVEIRA FLRD, CABACINHA CD, SANTOS LDT, BARROSO DG, SANTOS JÚNIOR AD, BRANT MC and SAMPAIO RA. 2015. Crescimento inicial de eucalipto e acácia, em diferentes arranjos de integração lavoura-pecuária-floresta. Cerne 21: 227-233.), evaluating the growth of eucalypt in agroforestry systems, found DBH values around 9.5 cm at 25 months of age. Lopes et al. (2017)LOPES ED, LAIA ML, SANTOS AS, SOARES GM, LEITE RWP and SOUZA MARTINS N. 2017. Influência do espaçamento de plantio na produção energética de clones de Corymbia e Eucalyptus. Floresta 47: 95-104. found values of DBH ranging from 3.28 to 4.57 cm (in different clones) in the spacing of 3.0 x 1.5 m at 12 months of age. These results are in agreement with the results obtained in the present study.
Regarding H (Figure 4b), our results supported similar findings by Nascimento et al. (2012)NASCIMENTO DF, LELES PSDS, OLIVEIRA NETO SND, MOREIRA RTS and ALONSO JM. 2012. Crescimento inicial de seis espécies florestais em diferentes espaçamentos. Cerne 18: 59-165., who also found different H values between tree species. The results of their study in the spacing of 3.0 x 2.0 m at 22 months of age are as follows: 2.8 m for the angico-vermelho (Anadenanthera macrocarpa); 4.1 m for guapuruvu (Schizolobium parahybae); 2.7 m in arroeira (Schinus terebinthifolius); 4.1 m in aloe (Cordia sp.); 1.5 m for inga (Inga marginata) and 3.2 m for cotton tree (Chorisia speciosa).
The occurrence of frost resulted in a severe reduction of MCD of guapuruvu (vertical arrows in Figure 4c). These results are in agreement with Souza et al. (2011)SOUZA VQ, CARON BO, SCHMIDT D, BEHLING A, BAMBERG R AND VIAN AL. 2011. Resistência de espécies arbóreas submetidas a extremos climáticos de geada em diferentes sistemas agroflorestais. Cienc Rural 41: 972-977. who classified this species as highly sensitive to frost. Despite the high degree of frost damage, this species presented a high degree of regrowth. Augspurger (2011)AUGSPURGER CK. 2011. Frost damage and its cascading negative effects on Aesculus glabra. Plant Ecol 212: 1193-1203. found that the death of branches reduced the canopy percentage and contributed to early senescence and reduced growth, and led to the death of Aesculus glabra buds. The freezing of extracellular solutions determines the senescence of leaves and stems and result in an imbalance between the chemical potential of intra and extra-cellular water. This leads to outward water transfer and cellular dehydration, decrease cell turgor, and lad to solute accumulation and the eventual rupture of the plasma membrane (Heber and Santarius 1973HEBER U and SANTARIUS KA. 1973. Cell death by cold and heat and resistance to extreme temperatures: mechanisms of hardening and dehardening. In: Precht H, Christophersen J, Hensel H and Larcher W (Eds), Temperature and life. Springer-Verlag, Berlin, Germany, p. 232-292.).
The superiority of DBH in strip system in the most of forest species (Table II) agrees with the results found by several authors, which concluded that a large degree of spacing promotes greater diameter growth of trees (Clark et al. 2008CLARK A, JORDAN L, SCHIMLECK L and DANIELS RF. 2008. Effect of initial planting spacing on wood properties of unthinned loblolly pine at age 21. For Prod J 58: 78-83., Harrington et al. 2009HARRINGTON TB, HARRINGTON CA and DEBELL DS. 2009. Effects of planting spacing and site quality on 25-year growth and mortality relationships of Douglas-fir (Pseudotsuga menziesii var. menziesii). For Ecol Manage 258: 18-25.). Another key factor in the growth of forest species is the arrangement in which trees are planted. The equidistance spacing of trees in center rows and the larger area of outer rows in strip systems contributed to the greater availability of resources thus allowing enhanced plant growth.
While eucalypt was observed to have higher growth rates compared to other tree species, less photosynthetically active radiation in its understory was available for the sugarcane (Figure 5b). This tree species, in the last cultivation year of the sugarcane (2011), intercepted more than 80% of photosynthetically active radiation and resulted in a reduction of 50% sugarcane stalk weight (Figure 5a). This is a result of interspecific competition, primarily of solar radiation between tree species and the species growing in its understory, as also examined by Monteith et al. (1991)MONTEITH JL, ONG CK and CORLETT JE. 1991. Microclimate interactions in agroforestry systems. Forest Ecol Manag 45: 31-44., Knowles et. al. (1999)KNOWLES RL, HORVATH GC, CARTER MA and HAWKE MF. 1999. Developing canopy closure model to predict overstorey/understorey relationships in Pinus radiata silvopastoral systems. Agroforest Syst 43: 109-119., Gillespie et al. (2000)GILLESPIE AR, JOSE S, MENGEL DB, HOOVER WL, POPE PE, SEIFERT JR, BIEHLE DJ, STALL T and BENJAMIN TJ. 2000. Defining competition vectors in a temperate alley cropping system in the midwestern USA. 1. Production physiology. Agroforest Syst 48: 25-40. and Jose et al. (2004)JOSE S, GILLESPIE AR and PALLARDY SG. 2004. Interespecific interactions in temperate agroforestry. Agrofor Syst 61: 237-255..
The observed results are in agreement with those of Hardy et al. (2004)HARDY JP, MELLOH R, KOENIG G, MARKS D, WINSTRAL A, POMEROY JW and LINK T. 2004. Solar radiation transmission through conifer canopies. Agr Forest Meteorol 126: 257-270., who reported that the amount of radiation intercepted by the forestry species and consequently the radiation that reaches the ground level is determined by canopy characteristics, such as the MCD, which can be affected by plant arrangement.
IPAR values in this study are similar to those found by Caron et al. (2012)CARON BO, LAMEGO FP, SOUZA VQ, COSTA EC, ELOY E, BEHLING A and TREVISAN R. 2012. Interceptação de radiação luminosa pelo dossel de espécies florestais e sua relação com o manejo de plantas daninhas. Cienc Rural 42: 75-82. between forest planting lines from 42.3% in black wattle, 83.2% in bracatinga and 89.1% in one-year-old eucalypt trees with one year old. Our results agree with several studies (Keating et al. 1999KEATING BA, ROBERTSON MJ, MUCHOW RC and HUTH NI. 1999. Modelling sugarcane production systems. I. Development and performance of the sugarcane module. Field Crops Res 48: 27-36., Inman-Bamber et al. 2010INMAN-BAMBER NG, BONNETT GD, SPILLMAN MF, HEWITT MH and GLASSOP D. 2010. Sucrose accumulation in sugarcane is influenced by temperature and genotype through the carbon source-sink balance. Crop Pasture Sci 61: 111-121., Cardozo and Sentelhas 2013CARDOZO NP and SENTELHAS PC. 2013. Climatic effects on sugarcane ripening under the influence of cultivars and crop age. Sci Agric 70: 449-456.), which show that the climate variables are the primary factors responsible for the yield and quality of sugarcane. A greater amount of intercepted solar radiation resulted in higher photosynthetic rates (Ribeiro et al. 2017RIBEIRO RV, MACHADO EC, MAGALHÃES FILHO JR, LOBO AKM, MARTINS MO, SILVEIRA JA, YIND X and STRUIK PC. 2017. Increased sink strength offsets the inhibitory effect of sucrose on sugarcane photosynthesis. J Plant Physiol 208: 61-69.), and consequently, a greater production of assimilates which can help determine the productive responses of sugarcane plants. Additionally, Elli et al. (2016)ELLI EF, CARON BO, PAULA GM, ELOY E, SCHWERZ F and SCHMIDT D. 2016. Ecofisiologia da cana-de-açúcar no sub-bosque de canafístula em arranjos de sistema agroflorestal. Comunicata Scientiae 7: 464-472. found that a reduction of solar radiation in agroforestry systems leads to greater stability in the metabolic activity and sucrose quantity of sugarcane throughout its cycle.
CONCLUSIONS
Among the studied species, eucalypt presented the greatest root collar diameter values, diameter at breast height, crown development, and plant height.
Angico, canafístula and eucalypt presented a greater growth in the strip system while guapuruvu presented greater growth in the line system. Bracatinga was not influenced by the agroforestry system. Sugarcane presented the greater production in the understory of angico and bracatinga.
ACKNOWLEGMENTS
The authors wish to acknowledge the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq – Brazil) and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES – Brazil) for their financial support.
REFERENCES
- ALLEN RG, PEREIRA LS, RAES D and SMITH M. 1998. Crop evapotranspiration: guidelines for computing crop water requirements. Rome: FAO, 300 p. (FAO Irrigation and drainage paper, 56).
- AUGSPURGER CK. 2011. Frost damage and its cascading negative effects on Aesculus glabra. Plant Ecol 212: 1193-1203.
- BAYALA J and WALLACE JS. 2015. The water balance of mixed tree crop systems. In: Ong CK, Black C and Wilson J (Eds), Tree-crop interactions, 2nd edition: agroforestry in a changing climate. CAB International, Wallingford, p. 146-190.
- BERLYN GP and CHO J. 2000. Light, moisture and nutrient use by plants. In: Ashton MS and Montagnini F (Eds), The silvicultural basis for agroforestry systems. CRC Press, Boca Raton, p. 9-39.
- CARDOZO NP and SENTELHAS PC. 2013. Climatic effects on sugarcane ripening under the influence of cultivars and crop age. Sci Agric 70: 449-456.
- CARON BO, LAMEGO FP, SOUZA VQ, COSTA EC, ELOY E, BEHLING A and TREVISAN R. 2012. Interceptação de radiação luminosa pelo dossel de espécies florestais e sua relação com o manejo de plantas daninhas. Cienc Rural 42: 75-82.
- CLARK A, JORDAN L, SCHIMLECK L and DANIELS RF. 2008. Effect of initial planting spacing on wood properties of unthinned loblolly pine at age 21. For Prod J 58: 78-83.
- CQFS - COMISSÃO DE QUÍMICA E FERTILIDADE DO SOLO. 2004. Manual de adubação e calagem para os estados do Rio Grande do Sul e Santa Catarina. Porto Alegre: Sociedade Brasileira de Ciência do Solo, RS, Brasil, 400 p.
- CUNHA NG SILVEIRA RDC, KOESTER E, OLIVEIRA LD, FILIPPINI ALBA JM, TERRES V and LOPES R. 2011. Estudos de Solos do Município de Frederico Westphalen, RS. EMBRAPA, Pelotas, RS. Brazil. (Circular Técnica 116).
- DEISS L, MORAES AD, PELISSARI A, SILVA VPD and ANDREOLLA VRM. 2016. Oat grain yield under nitrogen uses in an eucalyptus intercropping system in Subtropical Brazil. Rev Cienc Agron 47: 462-470.
- ELLI EF, CARON BO, PAULA GM, ELOY E, SCHWERZ F and SCHMIDT D. 2016. Ecofisiologia da cana-de-açúcar no sub-bosque de canafístula em arranjos de sistema agroflorestal. Comunicata Scientiae 7: 464-472.
- FERREIRA DHAA, SANTOS LELES PS, MACHADO EC, ABREU AHM and ABÍLIO FM. 2014. Crescimento de clone de Eucalyptus urophylla x E. grandis em diferentes espaçamentos. Rev Árvore 44: 431-440.
- GILLESPIE AR, JOSE S, MENGEL DB, HOOVER WL, POPE PE, SEIFERT JR, BIEHLE DJ, STALL T and BENJAMIN TJ. 2000. Defining competition vectors in a temperate alley cropping system in the midwestern USA. 1. Production physiology. Agroforest Syst 48: 25-40.
- HARDY JP, MELLOH R, KOENIG G, MARKS D, WINSTRAL A, POMEROY JW and LINK T. 2004. Solar radiation transmission through conifer canopies. Agr Forest Meteorol 126: 257-270.
- HARRINGTON TB, HARRINGTON CA and DEBELL DS. 2009. Effects of planting spacing and site quality on 25-year growth and mortality relationships of Douglas-fir (Pseudotsuga menziesii var. menziesii). For Ecol Manage 258: 18-25.
- HEBER U and SANTARIUS KA. 1973. Cell death by cold and heat and resistance to extreme temperatures: mechanisms of hardening and dehardening. In: Precht H, Christophersen J, Hensel H and Larcher W (Eds), Temperature and life. Springer-Verlag, Berlin, Germany, p. 232-292.
- INMAN-BAMBER NG, BONNETT GD, SPILLMAN MF, HEWITT MH and GLASSOP D. 2010. Sucrose accumulation in sugarcane is influenced by temperature and genotype through the carbon source-sink balance. Crop Pasture Sci 61: 111-121.
- JOSE S, GILLESPIE AR and PALLARDY SG. 2004. Interespecific interactions in temperate agroforestry. Agrofor Syst 61: 237-255.
- KEATING BA, ROBERTSON MJ, MUCHOW RC and HUTH NI. 1999. Modelling sugarcane production systems. I. Development and performance of the sugarcane module. Field Crops Res 48: 27-36.
- KNOWLES RL, HORVATH GC, CARTER MA and HAWKE MF. 1999. Developing canopy closure model to predict overstorey/understorey relationships in Pinus radiata silvopastoral systems. Agroforest Syst 43: 109-119.
- LOPES ED, LAIA ML, SANTOS AS, SOARES GM, LEITE RWP and SOUZA MARTINS N. 2017. Influência do espaçamento de plantio na produção energética de clones de Corymbia e Eucalyptus. Floresta 47: 95-104.
- MACHADO SA, ZAMIN NT, NASCIMENTO RGM and SANTOS AAP. 2014. Efeito de variáveis climáticas no crescimento mensal de Pinus taeda e Araucaria angustifolia em fase juvenil. Floresta Ambient 21: 170-181.
- MENDES MMS, LACERDA CF, CAVALCANTE ACR, FERNANDES FÉP and OLIVEIRA TS. 2013. Desenvolvimento do milho sob influência de árvores de pau branco em sistema agrossilvipastoril. Pesq Agropec Bras 48: 1342-1350.
- MONTEITH JL, ONG CK and CORLETT JE. 1991. Microclimate interactions in agroforestry systems. Forest Ecol Manag 45: 31-44.
- NASCIMENTO DF, LELES PSDS, OLIVEIRA NETO SND, MOREIRA RTS and ALONSO JM. 2012. Crescimento inicial de seis espécies florestais em diferentes espaçamentos. Cerne 18: 59-165.
- NEVES CMN, SILVA MLN, CURI N, MACEDO RLG, MOREIRA FDS and D’ANDRÉA AF. 2009. Indicadores biológicos da qualidade do solo em sistema agrossilvipastoril no noroeste do Estado de Minas Gerais. Ciên Agrotec 33: 105-112.
- NICODEMO MLF, MULLER MD, PORFÍRIO-DA-SILVA V, CARPANEZZI AA, PEZZOPANE JRM and BARIONI JÚNIOR W. 2016. Growth of native trees in two agroforestry systems. Rev Árvore 40: 639-648.
- OLIVEIRA FLRD, CABACINHA CD, SANTOS LDT, BARROSO DG, SANTOS JÚNIOR AD, BRANT MC and SAMPAIO RA. 2015. Crescimento inicial de eucalipto e acácia, em diferentes arranjos de integração lavoura-pecuária-floresta. Cerne 21: 227-233.
- ONG CK, BLACK CR, WALLACE JS, KHAN AAH, LOTT JE, JACKSON NA, HOWARD SB and SMITH DM. 2000. Productivity, microclimate and water use in Grevillea Robusta-based agroforestry systems on hill slopes in semi-arid Kenya agriculture. Agric Ecosyst Environ 80: 121-141.
- ONG CK and KHO RM. 2015. A framework for quantifying the various effects of tree-crop interactions. In: Ong CK, Black C and Wilson J (Eds), Tree-crop interactions, 2nd edition: agroforestry in a changing climate. CAB International, Wallingford, p. 1-23.
- PACIULLO DS, GOMIDE CA, CASTRO CR, FERNANDES PB, MÜLLER MD, PIRES MF, FERNANDES EM and XAVIER DF. 2011. Características produtivas e nutricionais do pasto em sistema agrossilvipastoril, conforme a distância das árvores. Pesq Agropec Bras 46: 1173-1186.
- PENG X, ZHANG Y, CAI J, JIANG Z and ZHANG S. 2009. Photosynthesis, growth and yield of soybean and maize in a tree-based agroforestry intercropping system on the Loess Plateau. Agrofor Syst 76: 569-577.
- PILAU J, ELLI EF, NARDINO M, KORCELSKI C, SCHMIDT D and CARON BO. 2015. Desenvolvimento e qualidade do azevém no sub-bosque de angico-vermelho em sistema silvipastoril. Comunicata Scientiae 6: 437-444.
- PINTO LFG, BERNARDES MS and SPAROVEK G. 2003. Feasibility of cultivation of sugarcane in agroforestry systems. Sci Agric 60: 489-493.
- RIBEIRO RV, MACHADO EC, MAGALHÃES FILHO JR, LOBO AKM, MARTINS MO, SILVEIRA JA, YIND X and STRUIK PC. 2017. Increased sink strength offsets the inhibitory effect of sucrose on sugarcane photosynthesis. J Plant Physiol 208: 61-69.
- RIVEST D, LORENTE M, OLIVIER A and MESSIER C. 2013. Soil biochemical properties and microbial resilience in agroforestry systems: effects on wheat growth under controlled drought and flooding conditions. Sci Total Environ 463: 51-60.
- SAS - STATISTICAL ANALYSIS SYSTEM. 2003. Getting started with the SAS Learning Edition. Cary, 200 p.
- SOUZA VQ, CARON BO, SCHMIDT D, BEHLING A, BAMBERG R AND VIAN AL. 2011. Resistência de espécies arbóreas submetidas a extremos climáticos de geada em diferentes sistemas agroflorestais. Cienc Rural 41: 972-977.
- STAPE JL et al. 2010. The Brazil eucalyptus potential productivity project: influence of water, nutrients and stand uniformity on wood production. For Ecol Manage 259: 1684-1694.
- THORNTHWAITE CW and MATHER JR. 1955. The Water Balance. Centerton, Drexel Institute of Technology, Laboratory of Climatology, Publications in climatology, 104 p.
- TRACY BF and ZHANG Y. 2008. Soil compaction, corn yield response, and soil nutrient pool dynamics within an integrated crop-livestock system in Illinois. Crop Sci 48: 1211-1218.
- XAVIER DF, LÉDO FJS, PACIULLO DSC, URQUIAGA S, ALVES BJR and BODDEY RM. 2014. Nitrogen cycling in a Brachiaria-based silvopastoral system in the Atlantic forest region of Minas Gerais, Brazil. Nutr Cycl Agroecosyst 99: 45-62.
Publication Dates
-
Publication in this collection
26 July 2018 -
Date of issue
Aug 2018
History
-
Received
2 May 2017 -
Accepted
26 July 2017