Abstract
Abstract:Genypterus brasiliensis, a demersal fish of large size (1.3 m of total length), occurs in the southwest Atlantic (23 to 38oS) and well appreciated due to the quality of its flesh. It was analyzed through longline and trawl fisheries, and the average length and weight of 724 specimens were significantly different between them, with the biggest and heaviest specimens caught by the former. The weight-length relationships were significantly different based upon sex: females Wt=4x10-7Lt3.3799; r2=0.97; n=262; p<0.001, and males: Wt=4x10-7Lt3.3886; r2=0.95; n=190; p<0.001. Using the otoliths readings for 153 specimens (APE of 2% and CV of 4.2%), the growth parameters estimates reveal statistical differences between sexes (p=0.05; r2=0.889). The fisheries were composed of individuals aged 1 to 13 (females) and 1 to 14 years old (males), with ring formation in late spring. The back calculated parameters (VBGF) were: Loo=1,452.38 mm, K=0.0996, t0=-0.2757 yr-1 (females) and Loo=983.07 mm, K=0.155, t0=-0.236 yr-1 (males), and longevity as 29.8 and 19.1 years old, respectively. The low biological production allied to the slow growth and a late complete maturation and its endemism, allow classify G. brasiliensis as a vulnerable resource, jeopardizing a sustainable fishery. Until reliable quantitative biological data are available, precautionary management must be considered.
Key words
population dynamics; endemism; fishes; fisheries
INTRODUCTION
Although considered “an unpatterned form” and a junior synonym for Genypterus blacodes (Bloch and Schneider 1801) (NielsenNIELSEN JG, COHEN DM, MARKIE DF and ROBINS CR. 1999. FAO Species Catalogue. Ophidiiform Fishes of the World. (Order Ophidiiformes). FAO Fish Synop 18: 1-178. et al. 1999), Genypterus brasiliensis (Regan, 1903) is recognized as a separate taxon (NakamuraNAKAMURA I. 1986. Family Ophidiidae. In: Nakamura I et al. (Eds), Important Fishes Trawled off Patagonia. Japan Marine Fisheries Resources Research Center, 369 p. 1986) and is accepted herein as an endemic groundfish species from the southwestern Atlantic (23o to 38oS) (FigueiredoFIGUEIREDO JL and MENEZES NA. 1978. Manual de Peixes Marinhos do Sudeste do Brasil. II. Teleostei (1). Museu de Zoologia, Universidade de São Paulo. São Paulo: Brasil, 110 p. and MenezesMENEZES NA, BUCKUP PA, FIGUEIREDO JL and MOURA RL. 2003. Catálogo das Espécies de Peixes Marinhos do Brasil. Museu de Zoologia. Universidade de São Paulo. São Paulo: Brasil, 160 p. 1978, Menezes et al. 2003). It is one of the most valuable species of bottom trawl and longline fisheries in southern Brazil, operating at depths between 50 and 510 m (TutuiTUTUI SLS, BASTOS GCC, TOMÁS ARG, TIAGO GG and ZAVALA-CAMIN LA. 2000. Species composition of the exploratory fisheries with bottom longline off southeastern Brazil. Cienc Cult 52(1): 55-58. et al. 2000, Graça-LopesGRAÇA-LOPES R, TOMÁS ARG, TUTUI SLS, SEVERINO-RODRIGUES E and PUZZI A. 2002. Fauna Acompanhante de Pesca Camaroeira no Litoral do Estado de São Paulo, Brasil. Bol Inst Pesca 28(2): 173-188. et al. 2002, PerezPEREZ JAA, WAHRLICH R, PEZZUTO PR, SCHWINGEL PR, LOPES FRA and RODRIGUES-RIBEIRO M. 2002. Deep-sea fishery off southern Brazil: recent trends of the Brazil fishing industry. J North Atlant Fish Sci 31: 1-18. et al. 2002, FrotaFROTA LO, COSTA PAS and BRAGA AC. 2004. Length-weight relationships of marine fishes from the central Brazilian coast. Naga 27(1-2): 20-26. et al. 2004, HaimoviciHAIMOVICI M, AVILA-DA-SILVA AO and ROSSI-WONGTSCHOWSKI CLDB. 2004. Prospecção Pesqueira de Espécies Demersais com Espinhel-de-fundo na Zona Econômica Exclusiva da Região Sudeste-Sul. Instituto Oceanográfico. Universidade de São Paulo. Série Documentos Revizee, 112 p. et al. 2004). The Brazilian fisheries statistics official series from 1995 to 2012 reported an average of 585.5 t (+/- 58.4 SEM) annual average catch of the species, with an increasing trend of the landings since 2004. This flesh, of an excellent quality, is of a great interest for the gastronomy, and it is sold in local markets as fillets, steaks or whole fish. Unfortunately, fishing effort data is not available, in general and in the present case, for Brazilian fisheries. Most of those catches were completed by trawling in the outer shelf and upper slope between 23o to 25oS, an area dominated by an intrusion of cold water mass (PiolaPIOLA AR, CAMPOS EJD, MOLLER JUNIOR O, CHARO M and MARTINEZ C. 2000. Subtropical shelf front off eastern South America. J Geophys Res 105(3): 6565-6578. et al. 2000, BelemBELEM AL, CASTELAO RM and ALBUQUERQUE AL. 2013. Controls of subsurface temperature variability in a western boundary upwelling system. Geophys Res Let 40(7): 1362-1366. et al. 2013). Meanwhile, early publications revealed catches in small numbers southwards, at the Subtropical Convergence System (Haimovici et al. 1994HAIMOVICI M, MARTINS AS, FIGUEIREDO JL and VIEIRA PC. 1994. Demersal bony fish of the outer shelf and upper slope of the southern Brazil Subtropical Convergence Ecosystem. Mar Ecol Prog Ser 108: 59-77.). In addition, the species lacks information concerning its life cycle and fisheries biology. This study aimed to describe the life history parameters of the species. Therefore, the growth size and weight for instantaneous rates of mortality were first investigated to determine the level of exploitation.
MATERIALS AND METHODS
Sampling was conducted by a survey of longline fisheries of the RV Orion in 1994 and 1995 (Tutui et al 2000), the RV Margus II during Revizee (a Brazilian research program of for Living Resources from Economic Exclusive Zone) in 1997 (Haimovici et al. 2004), and the RV Soloncy Moura otter trawl cruises during the winter and spring of 2001. All the surveys occurred in the outer shelf and upper slope (100 to 500 m depth) between 22°00’S to 34°40’S. Additional data were obtained from commercial fisheries from the double-rig trawl and pair trawl fleets in 2003-2004 and 2006-2007 at diverse fishery wharves in southeastern Brazil.
Total length (L, 1 mm) and weight (W, 0.01 g) were measured for each fish. The weight-length relationship was modeled using a power function (W=a*Lb) and tested for sex by an ANCOVA. Otoliths were removed, cleaned and stored dry. The intact right otoliths were measured with a caliper to the nearest 0.05 mm in length, and weighed to the nearest 0.001 g. Three to five pairs of otoliths from each size class were randomly chosen for cutting in the transverse plane into sections 0.3 mm in width with a Buhler Isomet™ low-speed saw. Two to three sections were cut at each otolith and mounted on glass microscope slides preserved with Canadian balsam. Digital images of the sections with reflected light and 16x magnification were obtained. For age estimation, 263 otoliths of both sexes and of all L sampled classes (1 cm) were used. The ages were estimated based on transitional counts of hyaline and opaque annuli (TurnerTURNER SC. 1986. Population dynamics of and impact of fishing on tilefish, Lopholatilus chamaeleonticeps, in the middle Atlantic-southern New England region during the 1970s and early 1980s. PhD thesis, New Brunswick Rutgers. University of New Jersey: NJ, USA, 289 p. 1986, BeckmanBECKMAN DW and WILSON CA. 1995. Seasonal timing of opaque zone formation in fish otoliths. In: Secor DH et al. (Eds), Recent Developments in Fish Otolith Research. Columbia, University of South Carolina Press, p. 27-43. and Wilson 1995), assuming one completed annuli as the presence of both one opaque and one hyaline rings. Two readings had to be equal to be accepted as legible. A coefficient of variation (CV%) and the Average Percent Error (APE) index (BeamishBEAMISH RJ and FOURNIER DA. 1981. A Method for Comparing the Precision of a Set of Age Determinations. Can J Fish Aquat Sc 38: 982-983. and Fournier 1981) were calculated. Otolith radius and the distance between otolith nucleus to the annuli (opaque ring, Rn) were determined using the software Image Tool (UTHSCSAUTHSCSA - UNIVERSITY OF TEXAS HEALTH SCIENCE CENTER AT SAN ANTONIO. 2008. Image Tool. Texas. 2008). ANCOVA was applied to test statistical differences between sexes in addition to Lt and total radius (Rt). The border readings (opaque or hyaline) were seasonally grouped (Spring: September to November, Summer: December to February, Autumn: March to May, Winter: June to August). To support the hypothesis of annual formation of growth increments (annuli), the data (separated by sex) were pooled for each season and by marginal increment analysis (MAI) expressed as: MI%=(Rt–Ru)/Rt, where Rt is the otolith radius and Ru the last annulus radius. All data were analyzed by the Kruskal-Wallis test (ZarZAR JH. 1999. Biostatistical Analysis. 4th ed., New Jersey: Prentice Hall, 663 p. 1999), as data was not normally distributed. If growth increments were deposited annually, a single peak could be expected in the seasonal frequency of each margin type (opaque/hyaline) over a year. Annual increments (annuli) were also validated by the spacing pattern between rings for each sex, the relationship among each radius and rings, and by the frequency distribution of the annulus radius; all were tested as proposed by LaiLAI HL, GALLUCCI VF, GUNDERSON DR and DONNELLY RF. 1996. Age determination in fisheries: methods and applications to stock assessment. In: Gallucci VF et al. (Eds), Stock Assessment Quantitative Methods and Applications for Small–scale Fisheries. CRC Press, Boca Raton: Florida, USA, p. 82-178. et al. (1996).
The back calculation was applied to verify the period of annuli formation by the body proportional hypothesis (BPH) function, Li=(a+bRi)/(a+bRt), where Li is the fish length obtained from back calculation for each ith ring, a and b are constants of the linear regression between Rt and Lt, Lt is the observed length, Ri the radius of each i ring, and Rt the total radius. The back calculated lengths in each age class were estimated using a non-linear least-squares regression to estimate the von Bertalanffy growth function (VBGF). The growth parameters L∞ (asymptotic length), K (intrinsic growth ratio) and t0 (theoretical age at length equal to 0) were estimated by the fitted function VBGF, and the growth performance index (Φ’=logK+2logL∞) (PaulyPAULY D and MUNRO JL. 1984. Once more on the comparison of growth in fish and invertebrates. Fishbyte 2(1): 21. and Munro 1984) was obtained for comparison with other species from the genus Genypterus.
The longevity of the stock was estimated as tmax=t0+2.996/K. The instantaneous natural mortality rate (M) was estimated by the empirical expression of PaulyPAULY D. 1980. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. ICES J Mar Sci 39(2): 175-192. (1980), considering 16 °C as the mean annual bottom water temperature at the fisheries’ grounds. With those value, the von Bertalanffy growth ratio M/K was also estimated.
All statistical analyses were completed with the SYSTAT package (WilkinsonWILKINSON P. 2000. Systat for Windows. Statistics ver. 10. SPSS, Evanston: IL, USA. 2000).
RESULTS
A total of 724 specimens in the length range of 235-1,100 mm were measured during the sampling. The average length and weight were significantly different by fisheries (ANCOVA: p<0.001), higher for the longline (Table I). From this total, 452 (62.4%) specimens were sexed, and the sex ratio was favorable for females (1:0.725).
Average length standard errors (Lmed± se) in mm, range and sampled number of the pink cusk eel Genypterus brasiliensis for distinct sample sources.
The weight-length relationship was significantly different between sexes, with females described by W=4x10-7L3.3799 (r2=0.97; n=262; sa =3.37; sb=0.033; p<0.001) and males by W=4x10-7L3.3886 (r2=0.95; n=190; sa=0.28; sb=0.004; p<0.001). For general purposes, a general weight-length relationship was also estimated (W=5x10-7L3.3712; r2=0.9719; n=557).
The ANOVA indicated no significant difference between the left and right otoliths with respect to mean fish length (n=365; p=0.251). The number of annual increments were recorded along the dorsal axis (Fig. 1), where the increments showed mode differences compared with other regions of the otolith.

Transversal section (0.3 mm width) of otolith from a pink cusk-eel Genypterus brasiliensis (female, 946 mm, 8 annuli, caught in October 2002 south of Rio de Janeiro, Brazil, around 300 m depth) observed through reflected light and 16 x.
From a total of 263 otoliths that were sectioned, 153 (102 females and 51 males) were considered legible and/or had similar ring readings. The reading comparison was consistence for both sexes with high precision (CV of 4.2% and APE of 2.0%), except for rings 1, 8 and 10 (few specimens sampled).
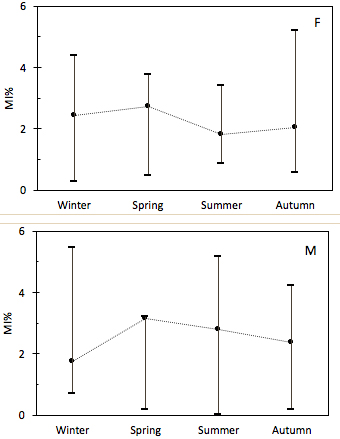
The relationship between Lt and Rt was significantly different between sexes (F=5,573.733; df:610; p=0.105) and were expressed as Lt♂=13.792+154.280Rt (r2=0.827; F=1,147.016; df: 202; p<0.001) and Lt♀=17.617+158.330Rt (r2=0.913; F=4,784.067; df=407; p<0.001). Assuming those results, 72.5% of the females and 58.8% of the males among all otolith sections analyzed corresponded to the 2 to 4 age class interval. The seasonal percentage opaque/hyaline border analysis reported peaks of deposition (austral Autumn and Winter) with no conflicts to marginal increment analysis (MAI, higher median of MI% for hyaline border in Winter to Spring), which suggest an annual deposition period during late Summer, Autumn and Winter (Fig. 2).

Marginal increment analysis (MI%, median and range) for females and males of the pink cusk-eel Genypterus brasiliensis.
The annuli readings showed 13 age classes for females and 14 for males. No difference of statistical significance was observed for the entire age period (1 to 13 years, Kruskal-Wallis: p=0.430) or for the most representative age period in the catches (5 to 8 years, Kruskal Wallis: p=0.217). However, this may indicate an approximate period for annuli formation and may assume that the number of annual increments increased consistently across the age range of fish.
The plots of average back calculated length (BPH, Fig. 3) corresponding to each age class and observed L models allowed the fitting of the von Bertalanffy growth curve for males L=983.07(1-e-0.155(t+0.236)) and females: L=1.452.38(1-e-0.0996(t+0.2757)), with ranges and confidence intervals, and age-at-length keys for each sex shown in Tables II and III, respectively. Because the largest fish measured was a female (1,100 mm), the highest estimated age was almost 14 years, which is essentially the same age for a male of 890 mm. The performances indexes (Φ’) presented values of 3.320 for females and of 3.175 for males, lower than other species of the same genus (JappJAPP AP. 1990. A new study on age and growth of kingklip Genypterus capensis off the south and west coasts of South Africa, with comments on its use for stock identification. S Afr J Mar Sci 9: 223-237. 1990, HornHORN PL. 1993. Growth, age structure, and productivity of ling Genypterus blacodes (Ophidiidae), in New Zealand waters. New Zeal J Mar Fresh 27: 385-397. 1993, WithellWITHELL AF and WANKOWSKI JWJ. 1989. Age and growth estimates for pink ling, Genypterus blacodes (Schneider), and gemfish, Rexea solandri (Curvier), from Eastern Bass Strait, Australia. Aust J Mar Freshw Res 40(2): 215-226. and Wankowski 1989, WiffWIFF R, OJEDA V and QUIROZ JC. 2007. Age and growth in pink cusk-eel (Genypterus blacodes) off the Chilean austral zone: evaluating differences between management fishing zones. J Appl Ichthyol 23: 270-272. et al. 2007, ChongCHONG J and AGUAYO M. 1990. Determinación de edad y estimación de los parâmetros de crecimiento del congrio dorado, Genypterus blacodes (Schneider, 1801) (Osteichthyes, Ophidiidae) en el Pacifico Sur-oriental. Biol Pesq 19: 55-67. and Aguayo 1990).
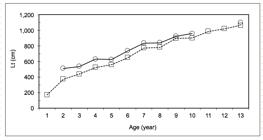
Average back calculated (BPH) and observed lengths (Lt) by age class of the pink cusk-eel Genypterus brasiliensis.
Back calculation growth (BPH) parameters fitted to von Bertalanffy function for the pink cusk-eel Genypterus brasiliensis (CI: 95% confidence interval, ASE: asymptotic standard error).
The ratio M/K (1.05 and 1.01, for females and males, respectively) invoke optimum catch sizes (Lopt) of 800.2 mm (males) and 955.0 mm (females), which corresponded to ages 7 to 9.5, respectively (see Table III). The optimum catch length-asymptotic length ratio (Lopt/L∞) was 0.658 for females and 0.814 for males, inside the interval (0.323 to 0.938, and slightly higher than the average of 0.620) assumed by CubillosCUBILLOS LA. 2003. An approach to estimate the natural mortality rate in fish stocks. Naga 26(1): 17-19. (2003) for all fish species.
DISCUSSION
The average size among specimens of Genypterus brasiliensis caught in the otter trawl (500-550 mm) and longline fisheries (720-737 mm), all from the same region, revealed a gear selectivity effect. The first interval included the mean length at first maturity of the population (L50 of 527.3 mm, authors’ unpublished data), and the latter was smaller than the length of entire populations that were mature (L100 approximately 950 mm) and close to the optimum catch size found for females (~955 mm).
Juveniles (less than 500 mm) were observed in the coastal pair trawl (in areas shallower than 40 m deep), and also in the offshore longline (deeper than 100 m) fisheries of southern Brazil (Haimovici et al. 2004, PerezPEREZ JAA. 2006. Potenciais de rendimento dos alvos da pesca de arrasto de talude do Sudeste e Sul do Brasil estimados a partir de parâmetros do ciclo de vida. BJAST 10(2): 1-11. 2006). In addition to the larger ones, fish sized in the 300-400 mm interval were common in regional fish markets (authors’ unpublished data) but with unknown fisheries areas.
Differing from the results by VillarinoVILLARINO MF and MORIOKA S. 1999. Problems in identification of 1st annual ring in sagittae of kingklip (Genypterus blacodes) and an attempt of utilization of lapilli for daily increment analysis. Seminario Final del Proyecto INIDEP – JICA sobre Evaluacion y monitoreo de recursos pesqueros 1994-1999, Mar del Plata: 6-9 Septiembre 1999, Argentina. p. 121-122. and Morioka (1999) for Genypterus blacodes (a close species with a wider geographical distribution along the south Pacific and also in the Argentine waters – south Atlantic), it was possible the recognize of the first age ring in G. brasiliensis. Although criticisms to marginal increment analysis (MAI) as an indicator of annulus formation, the requirements pointed out by CampanaCAMPANA SE. 2001. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol 59: 197-242. (2001), as the well-defined reproductive period (unpublished data), the samples obtained from throughout year (and higher than the minimum sample reported) and the APE and CV low values, may be strong reasons to be assumed the MAI results as realistic for Genypterus brasiliensis.
The estimated longevity, higher than 13 yr, did not differ from those of Chong and Aguayo (1990) (10 to 14 yr) but was smaller than Withell and Wankowski (1989) (21 yr), Horn (1993) (26-27 yr), and WrzesinskiWRZESINSKI O. 1984. Some features of the Genypterus sp. Populations off the coasts of Southwest Africa and Argentina. Rep Sea Fish Inst 19: 43-60. (1984) (30-31 yr) for G. blacodes.
The VBGF fit the back calculated lengths extremely well; however, the values of L∞ seem to be overestimated. The maximum length reported for the species was 1,100 mm, 24.3% smaller than L∞. The overestimated value of L∞ can be due to the lack of older age-classes in the samples. Even though the L∞ is only an indicative of the population, estimated from the existing data, the low representativeness of larger specimens in the samples can be an indication of overfishing.
Females and males of G. brasiliensis have about the same weight-at-length before the estimated mean length at first maturity. After maturity, females are slightly heavier, which is consistent with growth studies of other species of the genus (PaynePAYNE AIL. 1985. Growth and stock differentiation of kingklip (Genypterus capensis) on the southeast coast of South Africa. S Afr J Mar Sci 20: 49-56. 1985, Japp 1990, Horn 1993, BakerBAKER LL, WIFF R, QUIROZ JC, FLORES A, CÉSPEDES R, BARRIENTOS MA, OJEDA V and GATICA C. 2014. Reproductive ecology of the female pink cusk-eel (Genypterus blacodes): evaluating differences between fishery management zones in the Chilean austral zone. Environ Biol Fish 97(10): 1083-1093. et al. 2014, VegaVEGA R, SEPÚLVEDA C, BARNERT M, MARDONES A, ENCINA-MONTOYA F, OBERTI C, RAMIREZ D and ESTRADA JM. 2018. Reproductive capacity of the red cusk-eel Genypterus chilensis (Guichenot, 1848) in captivity. Lat Am J Aquat Res 46(2): 489-494. et al. 2018), except for Withell and Wankowski (1989). Age at length observations showed that the fastest growth prevails in the earliest five to six years of the life span, thus approximately 50% of growth of asymptotic length is reached before age five for both sexes (491.59 mm in age 4 and 726.19 mm in age 6, respectively). The age-length keys for each sex suggest that at least 65% of the sampled specimens were caught under age of 4 yr, close to the age of first maturity (equivalent to 4.3-4.5 yr). Meanwhile, the age of the entire population maturity was supposed to be reached at 8 to 9 yr. Older specimens (ages higher than 9 yr) were scarce in the population, should be viewed as a risk for the sustainability of the fisheries of the species.
Values of K and mean age at first maturity (tmat) are related to M (see Pauly 1980, VelascoVELASCO G, REIS EG and MIRANDA LV. 2003. Cálculo da taxa instantânea de mortalidade natural para Netuma barba (Lacépède, 1803) (Actinopterygii, Siluriformes, Ariidae). Acta Biologica Lepoldensia 25(2): 233-242. et al. 2003). High values of M reduce the expected lifespan and consequently tend to be associated with higher K and lower tmat. Although natural mortality rates estimated with Pauly’s method have been considered inaccurate by some authors (as GullandGULLAND JA and ROSENBERG AA. 1992. A review of length-based approaches to assessing fish stocks. FAO Fish Tec Pap 323: 1-100. and Rosenberg 1992, alternatively see KenchingtonKENCHINGTON TJ. 2014. Natural mortality estimators for information-limited fisheries. Fish Fish 15: 533-562. 2014 for some others remarks of estimation of M), other methods (such as Taylor, Alagaraja, Hoenig and Hewitt and Koenig) offered close values, particularly for females (0.101/yr, 0.155/yr, 0.152/yr and 0.101/yr). Alagaraja’s and Hoenig’s suggest much higher estimations (0.241/yr and 0.222/yr, respectively), similar to those estimated for G. blacodes (0.2/yr, at PuntPUNT AE and JAPP DW. 1994. Stock assessment of the kingklip Genypterus capensis off South Africa. S Afr J Mar Sci 14: 133-149. and Japp 1994). Conversely, the method of RiktherRIKTHER VA and EFANOV VN. 1976. On one of the approaches to estimation of natural mortality of fish populations. ICNAF Res Doc 76: 1-12. and Efanov (1976), using the mean age of maturity, showed values of 0.264/yr and 0.406/yr. The M/K ratio (a measure of the validity of mortality estimates) from M values obtained from the Pauly method (1.01 for females and 1.048 for males) were under the range of 1.5 to 2.5 reported by BevertonBEVERTON RJH and HOLT SJ. 1959. On the dynamics of exploited fish populations. Fisheries Investigation of the Ministry of Agriculture, Fisheries and Food. UK, 2 Sea Fisheries no. 19, 533 p. and Holt (1959). Even using M values from the other estimations cited above, most of values for males were outside this interval (except when using M from Rikther and Efanov, generating M/K=1.72), with extreme values for females, 1.048 to 4.072 (the later estimate using Rikther and Efanov). Then, from the inappropriate use of distinct methods for different sexes, and as the M values estimated through Pauly’s (1980) method are not equal to 1.5K, nor the length of the first maturity is less than 2/3 of L∞, the analysis of KirkwoodKIRKWOOD GP, BEDDINGTON JR and ROSSOUW JR. 1994. Harvesting species of different lifespans. In: Edwards PJ et al. (Eds), Large Scale Ecology and Conservation Biology. Oxford: UK. Blackwell Scientific, p. 199-227. et al. (1994) was unsuccessful. This situation leads us to face a distinct perspective to avoid the use of a fixed value for M. Another reason to strengthen not considering the recruitment constant is due to the annual variability of the oceanographic processes along the area, including the effective influences of the El Niño Southern Oscillation phenomena (ENSO), as discussed by PaesPAES ET and MORAES LES. 2007. A new hypothesis on the influence of the El Niño/La Niña upon the biological productivity, ecology and fisheries of the Southern Brazilian Bight. Panamjas 2(2): 94-102. and Moraes (2007), to constraint the usefulness of the invariants presented on Beverton and Holt (1959) and BeddingtonBEDDINGTON JR and KIRKWOOD GP. 2005. The estimation of potential yield and stock status using life-history parameters. Phil Trans Royal Soc Bull 360: 163-170. and Kirkwood (2005).
As exposed, it can be inferred that Genypterus brasiliensis was experiencing an excessive fishing pressure, mainly over the juvenile stratum submitted to the coastal trawl fisheries, which suggests a troublesome scenario. Haimovici et al. (2004) reported the highest densities for the species to the upper slope of 32o to 34oS followed by 22o to 23oS (the later an area influenced by upwelling waters). Both areas are also intensive fishing grounds, especially after the beginning of the decade of 2000, when chartered fishery fleets had operated in southern Brazil (Perez et al. 2002). The most intense fishery efforts continued down the outer shelf and upper slope (100 to 250 m), in the zone whereas this species was one of the most fishable resources (Perez 2006). Moreover, as noted by PattersonPATTERSON K. 1992. Fisheries for small pelagic species: an empirical approach to management targets. Rev Fish Biol Fish 2(4): 321-338. (1992), stocks can sustain exploitation rates of only 0.5 to 1.0 times the natural mortality rate and allied to the underestimation of the optimal exploitation rate for long-lived resources, the species should be at risk.
Although no fecundity studies of Genypterus brasiliensis were known to date, and by acceptance of a similar pattern to G. blacodes, which includes the elder females of the genus as BOFFFF (Big Old Fat Fecund Female Fish), and their removal by fisheries should result in potential risk to the structure of the population (HixonHIXON MA, JOHNSON DW and SOGARD SM. 2013. BOFFFFs: on the importance of conserving old-growth age structure in fishery populations. ICES J Mar Sci 71(8): 2171-2185. et al. 2013). It must be highlighted that individuals younger than 5 years old composed approximately 66% of male and 71.5% of the female catches of G. brasiliensis. In addition, the smallest fishes of valuable species (such as G. brasiliensis) often have minor interest from fisheries. Thus, the small fishes may not be reported in the landings and may be compounded in the discards (unreported catches), particularly in the trawl fisheries, generating an additional source of fisheries mortality that is not always recognized. Despite this, specimens smaller than 300 mm were reported in the double-rig trawl fisheries (Graça-Lopes et al. 2002), also in the discard of the species from the chartered pot and trawl fisheries fleets operating along upper slope off southern Brazil (Perez et al. 2002). The slow growth and a late complete maturation (7-9 yr), which suggests low biological production, allied to non-target fisheries interest, the low resilience and endemism, classify Genypterus brasiliensis as a potential vulnerable resource, jeopardizing a sustainable fishery. The catchable stock of the species is herein seen as another fisheries resource vulnerable to overexploitation. Until reliable quantitative biological data are available, precautionary management must be considered. Moreover, Genypterus brasiliensis is not cited on the worldwide IUCN and CITES red lists of endangered species. However, the southernmost Brazilian province (State of Rio Grande do Sul) includes this species on a regional list (State Law 51797 of September 8th, 2014) as near threatened.
ACKNOWLEGMENTS
This text is part of an effort to recognize new fisheries resources from the Brazilian EEZ. The authors also thank Manuel Haimovici (Universidade de Rio Grande/UFRG) and Carmen L.D.B. Rossi-Wongtschowski (Instituto Oceanografico, Universidade de São Paulo/IOUSP) for encouraging this study. Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) provided an undergraduate (ITI ‘Revizee’) scholarship to CLTE, and Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) provided a Post-doctoral scholarship to GV (Grant number 05/59715-2).
REFERENCES
- BAKER LL, WIFF R, QUIROZ JC, FLORES A, CÉSPEDES R, BARRIENTOS MA, OJEDA V and GATICA C. 2014. Reproductive ecology of the female pink cusk-eel (Genypterus blacodes): evaluating differences between fishery management zones in the Chilean austral zone. Environ Biol Fish 97(10): 1083-1093.
- BEAMISH RJ and FOURNIER DA. 1981. A Method for Comparing the Precision of a Set of Age Determinations. Can J Fish Aquat Sc 38: 982-983.
- BECKMAN DW and WILSON CA. 1995. Seasonal timing of opaque zone formation in fish otoliths. In: Secor DH et al. (Eds), Recent Developments in Fish Otolith Research. Columbia, University of South Carolina Press, p. 27-43.
- BEDDINGTON JR and KIRKWOOD GP. 2005. The estimation of potential yield and stock status using life-history parameters. Phil Trans Royal Soc Bull 360: 163-170.
- BELEM AL, CASTELAO RM and ALBUQUERQUE AL. 2013. Controls of subsurface temperature variability in a western boundary upwelling system. Geophys Res Let 40(7): 1362-1366.
- BEVERTON RJH and HOLT SJ. 1959. On the dynamics of exploited fish populations. Fisheries Investigation of the Ministry of Agriculture, Fisheries and Food. UK, 2 Sea Fisheries no. 19, 533 p.
- CAMPANA SE. 2001. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol 59: 197-242.
- CHONG J and AGUAYO M. 1990. Determinación de edad y estimación de los parâmetros de crecimiento del congrio dorado, Genypterus blacodes (Schneider, 1801) (Osteichthyes, Ophidiidae) en el Pacifico Sur-oriental. Biol Pesq 19: 55-67.
- CUBILLOS LA. 2003. An approach to estimate the natural mortality rate in fish stocks. Naga 26(1): 17-19.
- FIGUEIREDO JL and MENEZES NA. 1978. Manual de Peixes Marinhos do Sudeste do Brasil. II. Teleostei (1). Museu de Zoologia, Universidade de São Paulo. São Paulo: Brasil, 110 p.
- FROTA LO, COSTA PAS and BRAGA AC. 2004. Length-weight relationships of marine fishes from the central Brazilian coast. Naga 27(1-2): 20-26.
- GRAÇA-LOPES R, TOMÁS ARG, TUTUI SLS, SEVERINO-RODRIGUES E and PUZZI A. 2002. Fauna Acompanhante de Pesca Camaroeira no Litoral do Estado de São Paulo, Brasil. Bol Inst Pesca 28(2): 173-188.
- GULLAND JA and ROSENBERG AA. 1992. A review of length-based approaches to assessing fish stocks. FAO Fish Tec Pap 323: 1-100.
- HAIMOVICI M, AVILA-DA-SILVA AO and ROSSI-WONGTSCHOWSKI CLDB. 2004. Prospecção Pesqueira de Espécies Demersais com Espinhel-de-fundo na Zona Econômica Exclusiva da Região Sudeste-Sul. Instituto Oceanográfico. Universidade de São Paulo. Série Documentos Revizee, 112 p.
- HAIMOVICI M, MARTINS AS, FIGUEIREDO JL and VIEIRA PC. 1994. Demersal bony fish of the outer shelf and upper slope of the southern Brazil Subtropical Convergence Ecosystem. Mar Ecol Prog Ser 108: 59-77.
- HIXON MA, JOHNSON DW and SOGARD SM. 2013. BOFFFFs: on the importance of conserving old-growth age structure in fishery populations. ICES J Mar Sci 71(8): 2171-2185.
- HORN PL. 1993. Growth, age structure, and productivity of ling Genypterus blacodes (Ophidiidae), in New Zealand waters. New Zeal J Mar Fresh 27: 385-397.
- JAPP AP. 1990. A new study on age and growth of kingklip Genypterus capensis off the south and west coasts of South Africa, with comments on its use for stock identification. S Afr J Mar Sci 9: 223-237.
- KENCHINGTON TJ. 2014. Natural mortality estimators for information-limited fisheries. Fish Fish 15: 533-562.
- KIRKWOOD GP, BEDDINGTON JR and ROSSOUW JR. 1994. Harvesting species of different lifespans. In: Edwards PJ et al. (Eds), Large Scale Ecology and Conservation Biology. Oxford: UK. Blackwell Scientific, p. 199-227.
- LAI HL, GALLUCCI VF, GUNDERSON DR and DONNELLY RF. 1996. Age determination in fisheries: methods and applications to stock assessment. In: Gallucci VF et al. (Eds), Stock Assessment Quantitative Methods and Applications for Small–scale Fisheries. CRC Press, Boca Raton: Florida, USA, p. 82-178.
- MENEZES NA, BUCKUP PA, FIGUEIREDO JL and MOURA RL. 2003. Catálogo das Espécies de Peixes Marinhos do Brasil. Museu de Zoologia. Universidade de São Paulo. São Paulo: Brasil, 160 p.
- NAKAMURA I. 1986. Family Ophidiidae. In: Nakamura I et al. (Eds), Important Fishes Trawled off Patagonia. Japan Marine Fisheries Resources Research Center, 369 p.
- NIELSEN JG, COHEN DM, MARKIE DF and ROBINS CR. 1999. FAO Species Catalogue. Ophidiiform Fishes of the World. (Order Ophidiiformes). FAO Fish Synop 18: 1-178.
- PAES ET and MORAES LES. 2007. A new hypothesis on the influence of the El Niño/La Niña upon the biological productivity, ecology and fisheries of the Southern Brazilian Bight. Panamjas 2(2): 94-102.
- PATTERSON K. 1992. Fisheries for small pelagic species: an empirical approach to management targets. Rev Fish Biol Fish 2(4): 321-338.
- PAULY D. 1980. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. ICES J Mar Sci 39(2): 175-192.
- PAULY D and MUNRO JL. 1984. Once more on the comparison of growth in fish and invertebrates. Fishbyte 2(1): 21.
- PAYNE AIL. 1977. Stock differentiation and growth of the southern African kingklip Genypterus capensis. Invest Rep Sea Fish B S Afr 133: 1-32.
- PAYNE AIL. 1985. Growth and stock differentiation of kingklip (Genypterus capensis) on the southeast coast of South Africa. S Afr J Mar Sci 20: 49-56.
- PEREZ JAA. 2006. Potenciais de rendimento dos alvos da pesca de arrasto de talude do Sudeste e Sul do Brasil estimados a partir de parâmetros do ciclo de vida. BJAST 10(2): 1-11.
- PEREZ JAA, WAHRLICH R, PEZZUTO PR, SCHWINGEL PR, LOPES FRA and RODRIGUES-RIBEIRO M. 2002. Deep-sea fishery off southern Brazil: recent trends of the Brazil fishing industry. J North Atlant Fish Sci 31: 1-18.
- PIOLA AR, CAMPOS EJD, MOLLER JUNIOR O, CHARO M and MARTINEZ C. 2000. Subtropical shelf front off eastern South America. J Geophys Res 105(3): 6565-6578.
- PUNT AE and JAPP DW. 1994. Stock assessment of the kingklip Genypterus capensis off South Africa. S Afr J Mar Sci 14: 133-149.
- RIKTHER VA and EFANOV VN. 1976. On one of the approaches to estimation of natural mortality of fish populations. ICNAF Res Doc 76: 1-12.
- TURNER SC. 1986. Population dynamics of and impact of fishing on tilefish, Lopholatilus chamaeleonticeps, in the middle Atlantic-southern New England region during the 1970s and early 1980s. PhD thesis, New Brunswick Rutgers. University of New Jersey: NJ, USA, 289 p.
- TUTUI SLS, BASTOS GCC, TOMÁS ARG, TIAGO GG and ZAVALA-CAMIN LA. 2000. Species composition of the exploratory fisheries with bottom longline off southeastern Brazil. Cienc Cult 52(1): 55-58.
- UTHSCSA - UNIVERSITY OF TEXAS HEALTH SCIENCE CENTER AT SAN ANTONIO. 2008. Image Tool. Texas.
- VEGA R, SEPÚLVEDA C, BARNERT M, MARDONES A, ENCINA-MONTOYA F, OBERTI C, RAMIREZ D and ESTRADA JM. 2018. Reproductive capacity of the red cusk-eel Genypterus chilensis (Guichenot, 1848) in captivity. Lat Am J Aquat Res 46(2): 489-494.
- VELASCO G, REIS EG and MIRANDA LV. 2003. Cálculo da taxa instantânea de mortalidade natural para Netuma barba (Lacépède, 1803) (Actinopterygii, Siluriformes, Ariidae). Acta Biologica Lepoldensia 25(2): 233-242.
- VILLARINO MF and MORIOKA S. 1999. Problems in identification of 1st annual ring in sagittae of kingklip (Genypterus blacodes) and an attempt of utilization of lapilli for daily increment analysis. Seminario Final del Proyecto INIDEP – JICA sobre Evaluacion y monitoreo de recursos pesqueros 1994-1999, Mar del Plata: 6-9 Septiembre 1999, Argentina. p. 121-122.
- WIFF R, OJEDA V and QUIROZ JC. 2007. Age and growth in pink cusk-eel (Genypterus blacodes) off the Chilean austral zone: evaluating differences between management fishing zones. J Appl Ichthyol 23: 270-272.
- WILKINSON P. 2000. Systat for Windows. Statistics ver. 10. SPSS, Evanston: IL, USA.
- WITHELL AF and WANKOWSKI JWJ. 1989. Age and growth estimates for pink ling, Genypterus blacodes (Schneider), and gemfish, Rexea solandri (Curvier), from Eastern Bass Strait, Australia. Aust J Mar Freshw Res 40(2): 215-226.
- WRZESINSKI O. 1984. Some features of the Genypterus sp. Populations off the coasts of Southwest Africa and Argentina. Rep Sea Fish Inst 19: 43-60.
- ZAR JH. 1999. Biostatistical Analysis. 4th ed., New Jersey: Prentice Hall, 663 p.
Publication Dates
-
Publication in this collection
12 Aug 2019 -
Date of issue
2019
History
-
Received
8 Oct 2017 -
Accepted
17 Oct 2018