Abstract
We investigated whether essential oil and aqueous and ethanolic extracts from M. vittoriana leaves have phytotoxic effects on the germination and initial development, and cytogenotoxic effects on the cell cycle, of model plants. The essential oil and extracts of M. vittoriana were characterized and used as treatments in phytotoxicity and cytotoxicity tests. The results indicated a reduction in germinative parameters and plant growth, with the higher concentrations of extracts and essential oil having the most evident effects. The cell cycle was also affected with a reduction of the mitotic index and the presence of chromosomal and nuclear alterations. All treatments showed clastogenic and aneugenic modes of action. The results can be associated with the synergistic effects of metabolites found in the extracts and essential oil, mainly the presence of the sesquiterpene germacrene D in the essential oil and of catechins, saponins, and tannins in the extracts. These substances inhibit plant germination and growth, confirming the phytotoxic effects of M. vittoriana in plant models, which should now be tested under field conditions.
Key words
Atlantic Forest; cytotoxicity; essential oil; plant bioassays
INTRODUCTION
Plants interact in different ways in search of abiotic resources. Such interactions may hinder or facilitate the establishment of neighboring species (Inderjit 2005INDERJIT. 2005. Soil microorganisms: An important determinant of allelopathic activity. Plant Soil 274: 227-236.). Allelopathy is a form of interference generally associated with the negative influence of one plant on the growth and development of other plants through biomolecules (allelochemicals) released into the environment (Chengxu et al. 2011CHENGXU W, MINGXING Z, XUHUI C & BO Q. 2011. Review on allelopathy of exotic invasive plants. Procedia Eng 18: 240-246., Reigosa et al. 1999REIGOSA MJ, SÁNCHEZ-MOREIRAS A & GONZÁLEZ L. 1999. Ecophysiological approach in allelopathy. Crit Rev Plant Sci 18: 577-608.). These compounds are products of specialized metabolism and are emitted through leaching of aerial parts, root exudation, volatilization or by decomposing plant residues (Rice 1984RICE EL. 1984. Allelopathy, 2nd ed., Orlando: Academic Press, 423 p., Weir et al. 2004WEIR TL, PARK SW & VIVANCO JM. 2004. Biochemical and physiological mechanisms mediated by allelochemicals. Curr Opin Plant Biol 7: 472-479.). Different physiological processes of other plants can be affected, such as photosynthesis, nutrient absorption, and cell division or elongation (Inderjit & Duke 2003INDERJIT & DUKE SO. 2003. Ecophysiological aspects of allelopathy. Planta 217: 529-539.).
Allelopathy is a natural and common phenomenon of different plant communities (Blanco 2007BLANCO JA. 2007. The representation of allelopathy in ecosystem-level forest models. Ecol Modell 209: 65-77., Smith 1989SMITH AE. 1989. The potential allelopathic characteristics of bitter sneezeweed (Helenium amarum). Weed Sci 37: 665-669.). Allelochemicals play an important role in controlling species diversity and can determine the organization of natural communities (Chou 1999CHOU CH. 1999. Roles of allelopathy in plant biodiversity and sustainable agriculture. Crit Rev Plant Sci 18: 609-636.) through changes in structural and successional patterns, and by favoring the dominance of introduced species (Callaway & Ridenour 2004CALLAWAY RM & RIDENOUR WM. 2004. Novel weapons: Invasive success and the evolution of increased competitive ability. Front Ecol Environ 2: 436-443., Fernandez et al. 2013FERNANDEZ C, SANTONJA M, GROS R, MONNIER Y, CHOMEL M, BALDY V & BOUSQUET-MÉLOU A. 2013. Allelochemicals of Pinus halepensis as drivers of biodiversity in Mediterranean open mosaic habitats during the colonization stage of secondary succession. J Chem Ecol 39: 298-311., Kato-Noguchi et al. 2017KATO-NOGUCHI H, KIMURA F, OHNO O & SUENAGA K. 2017. Involvement of allelopathy in inhibition of understory growth in red pine forests. J Plant Physiol 218: 66-73.). In this sense, allelopathy has demonstrated the relevance of plant-plant chemical interactions for modulating the structure and functioning of communities, which was previously limited to competition for resources through the removal or reduction of shared environmental factors in the habitat (Inderjit et al. 2011INDERJIT, WARDLE DA, KARBAN R & CALLAWAY RM. 2011. The ecosystem and evolutionary contexts of allelopathy. Trends Ecol Evol 26: 655-662., Tukey 1969TUKEY HB. 1969. Implications of allelopathy in agricultural plant science. Bot Rev 35: 1-16.). Some tropical plants are especially known for developing allelochemicals that allow them to successfully outcompete their neighbors (Ooka & Owens 2018OOKA JK & OWENS DK. 2018. Allelopathy in tropical and subtropical species. Phytochem Rev 17: 1225-1237.). However, our understanding about the role that allelopathy plays in shaping the structure and organization of tropical plant communities is still limited given the great diversity of these systems.
The pantropical Myrtaceae is one of the most species rich plant families in tropical ecosystems (Beech et al. 2017BEECH E, RIVERS M, OLDFIELD S & SMITH PP. 2017. GlobalTreeSearch: The first complete global database of tree species and country distributions. J Sustain For 36: 454-489., Zappi et al. 2015ZAPPI DC ET AL. 2015. Growing knowledge: An overview of Seed Plant diversity in Brazil. Rodriguésia 66: 1085-1113.) and allelopathic effects have been demonstrated for several of its species (e.g., Caputo et al. 2020CAPUTO L, SMERIGLIO A, TROMBETTA D, CORNARA L, TREVENA G, VALUSSI M, FRATIANNI F, FEO DV & NAZZARO F. 2020. Chemical composition and biological activities of the essential oils of Leptospermum petersonii and Eucalyptus gunnii. Front Microbiol 11: 1-15., Kapoor et al. 2019KAPOOR D, RINZIM, TIWARI A, SEHGAL A, LANDI M, BRESTIC M & SHARMA A. 2019. Exploiting the allelopathic potential of aqueous leaf extracts of Artemisia absinthium and Psidium guajava against parthenium hysterophorus, a widespread weed in India. Plants 8: 1-13., Verdeguer et al. 2020VERDEGUER M, CASTAÑEDA LG, TORRES-PAGAN N, LLORENS-MOLINA J & CARRUBBA A. 2020. Control of Erigeron bonariensis with Thymbra capitata, Mentha piperita, Eucalyptus camaldulensis, and Santolina chamaecyparissus essential oils. Molecules 25: 562.). Studies conducted with representatives of the genus Myrcia have demonstrated their ability to affect the growth and development of undesirable species in crop systems (Imatomi et al. 2015IMATOMI M, NOVAES P, MIRANDA MAFM & GUALTIERI SCJ. 2015. Phytotoxic effects of aqueous leaf extracts of four Myrtaceae species on three weeds. Acta Sci Agron 37: 241-248.). Phytotoxic effects have been demonstrated for species such as M. bella Cambess., M. multiflora DC., M. splendens DC (Imatomi et al. 2013aIMATOMI M, NOVAES P & GUALTIERI SCJ. 2013a. Interspecific variation in the allelopathic potential of the family Myrtaceae. Acta Bot Brasilica 27: 54-61.), M. guianensis (Aubl.) DC. (Franco et al. 2015FRANCO DM, SALDANHA LL, SILVA EM, NOGUEIRA F, DOKKEDAL AL, SANTOS C & ALMEIDA LFR. 2015. Effects of leaf extracts of Myrcia guianensis (Aubl.) DC. on growth and gene expression during root development of Sorghum bicolor (L.) Moench. Allelopath J 35: 237-248., Souza Filho et al. 2006SOUZA FILHO APS, SANTOS RA, SANTOS LS, GUILHON GMP, SANTOS AS, ARRUDA MSP, MULLER AH & ARRUDA AC. 2006. Allelophatic potential of Myrcia guianensis. Planta Daninha 24: 649-656.), and M. tomentosa DC. (Imatomi et al. 2013bIMATOMI M, NOVAES P, MATOS AP, GUALTIERI SCJ, MOLINILLO JMG, LACRET R, VARELA RM & MACÍAS FA. 2013b. Phytotoxic effect of bioactive compounds isolated from Myrcia tomentosa (Myrtaceae) leaves. Biochem Syst Ecol 46: 29-35.). However, given the great diversity and widespread distribution of this plant family in the tropics, our knowledge is still largely limited. There have been advances with well-known and non-native species, or native species with widespread distributions (Caputo et al. 2020CAPUTO L, SMERIGLIO A, TROMBETTA D, CORNARA L, TREVENA G, VALUSSI M, FRATIANNI F, FEO DV & NAZZARO F. 2020. Chemical composition and biological activities of the essential oils of Leptospermum petersonii and Eucalyptus gunnii. Front Microbiol 11: 1-15., Qin et al. 2018QIN F, LIU S & YU S. 2018. Effects of allelopathy and competition for water and nutrients on survival and growth of tree species in Eucalyptus urophylla plantations. For Ecol Manage 424: 387-395., Ruwanza et al. 2015RUWANZA S, GAERTNER M, ESLER KJ & RICHARDSON DM. 2015. Allelopathic effects of invasive Eucalyptus camaldulensis on germination and early growth of four native species in the Western Cape, South Africa. South For 77: 91-105., Vasconcelos et al. 2019VASCONCELOS LC, SANTOS ES, BERNARDES CO, FERREIRA MFS, FERREIRA A, TULER AC, CARVALHO JAM, PINHEIRO PF & PRAÇA-FONTES MM. 2019. Phytochemical analysis and effect of the essential oil of Psidium L. species on the initial development and mitotic activity of plants. Environ Sci Pollut Res 26: 26216-26228.). Nonetheless, we have little knowledge about how endemic species of Myrtaceae make use of allelochemicals to succeed in tropical plant communities. Investigating the allelopathic effects of native species of this family will shed light on the mechanisms responsible for floristic composition, vegetation structure, and spatial patterns in tropical plant communities.
Garbin et al. (2016)GARBIN ML, GUIDONI-MARTINS KG, HOLLUNDER RK, MARIOTTE P, SCARANO FR & CARRIJO TT. 2016. Spatial segregation of subordinate species is not controlled by the dominant species in a tropical coastal plant community. Perspect Plant Ecol Evol Syst 18: 23-32. revealed that, in a sandy coastal plant community in the Atlantic Forest, shrub patches with a greater abundance of Myrcia vittoriana Kiaersk. were not occupied by Erythroxylum subsessile (Mart.) O.E.Schulz (Erythroxylaceae) and Myrsine parvifolia A.DC (Primulaceae). Based on the spatial patterns observed for these species, Garbin et al. (2016)GARBIN ML, GUIDONI-MARTINS KG, HOLLUNDER RK, MARIOTTE P, SCARANO FR & CARRIJO TT. 2016. Spatial segregation of subordinate species is not controlled by the dominant species in a tropical coastal plant community. Perspect Plant Ecol Evol Syst 18: 23-32. suggested that, among other factors, M. vittoriana could be exerting allelopathic effects on E. subsessile and M. parvifolia. Thus, testing the allelopathic effects of M. vittoriana can help to understand the spatial distribution patterns reported for these species in their natural environment, as well as for other native species of Myrtaceae found in tropical ecosystems. A first step towards approaching this target is to test the phytotoxic effects of M. vittoriana on model plants.
This study, therefore, aimed to investigate the phytotoxic effects of the essential oil and the aqueous and ethanolic extracts of M. vittoriana through their activities on the initial development (phytotoxicity) and cell cycle (cytotoxicity) of the model plants Lactuca sativa L. (lettuce) and Allium cepa L. (onion). The compounds present in the essential oil and extracts of M. vittoriana were also studied and identified phytochemically. The investigation of the cytotoxicity effects of compounds of M. vittoriana on the cell cycle of L. sativa and A. cepa will help to explain morphological and physiological changes found at the cellular level and the mode of action of these substances.
MATERIALS AND METHODS
Plant material
Fresh leaves of M. vittoriana were collected from individual plants growing in a sandy coastal plain plant community in southeastern Brazil (22° 23’S, 41°45 ’W, sea level) (Figure 1). Healthy leaves were collected in the field during the morning from nine individuals distributed among three vegetation patches (three individuals per patch), previously studied by Garbin et al. (2016)GARBIN ML, GUIDONI-MARTINS KG, HOLLUNDER RK, MARIOTTE P, SCARANO FR & CARRIJO TT. 2016. Spatial segregation of subordinate species is not controlled by the dominant species in a tropical coastal plant community. Perspect Plant Ecol Evol Syst 18: 23-32.. Some of the collected leaves were dried in an oven at 40° for 72 hours and then crushed into small pieces for use in the preparation of leaf extracts. The tests were performed using seeds of the eudicotyledonous L. sativa L. (commercial cultivar Alface Grandes Lagos Americana, Isla seeds), and the monocotyledonous A. cepa L. (commercial cultivar Baia Periforme, Isla seeds).

Individual of Myrcia vittoriana Kiaersk. in a shrub patch: (a) Individual. (b) The appearance of a vegetation patch in a sandy coastal plain environment. Photo: Amélia Carlos Tuler.
Extraction and yield of essential oil
Essential oil was extracted from fresh leaves of M. vittoriana by hydrodistillation using a Clevenger apparatus. About 420g of freshly perforated plant material was placed with distilled water in a 5L round-bottom flask equipped with an essential oil extraction condenser. The contents were boiled for approximately four hours, after which the hydrolysate (a mixture of water and oil) was collected through the condenser nozzle and centrifuged at 5000 rpm for 5 minutes to separate the aqueous and oily phases. The light-yellow oil (supernatant) was removed with the aid of a Pasteur pipette and stored in an amber glass bottle in a freezer at temperatures below 0°C (Pinheiro et al. 2013PINHEIRO PF, QUEIROZ VT, RONDELLI VM, COSTA AV, MARCELINO TP & PRATISSOLI D. 2013. Insecticidal activity of citronella grass essential oil on Frankliniella schultzei and Myzus persicae. Ciênc Agrotec 37: 138-144.). The extraction procedure was repeated four times. Yield was calculated using the ratio between the mass of the extracted oil and the fresh mass of the sample (%, m / m).
Chemical characterization of essential oil
The essential oil samples obtained from the leaves of M. vittoriana were analyzed by gas chromatography with a flame ionization detector (GC/FID) (Shimadzu GC-2010 Plus apparatus) and by gas chromatography coupled to mass spectrometry (GC/MS) (Shimadzu QPMS-2010) following the methodology of Souza et al. (2017)SOUZA TS, FERREIRA MFS, MENINI L, SOUZA JRCL, PARREIRA LA, CECON PR & FERREIRA A. 2017. Essential oil of Psidium guajava: Influence of genotypes and environment. Sci Hortic 216: 38-44., with adaptations. The following chromatographic conditions were used in both analyses: fused silica capillary column (30 m x 0.25 mm) with Rtx®-5MS stationary phase (0.25 μm film thickness); N2 (in GC/FID analysis) and He (in CG/MS analysis) as carrier gases with a flow rate of 3.0 mL/min; the oven temperature followed a schedule in which it remained at an initial temperature of 40°C for 3 minutes and then gradually increased by 3°C/minute until it reached 240°C, maintaining this temperature for 5 minutes; injector temperature of 250°C; detector temperature of 280°C; split ratio of 1:30. The GC/MS analyses were performed on equipment operating by electronic impact with 70 eV; scan speed 1,000; scanning interval of 0.50 fragments / second and detected fragments from 29 to 400 (m/z).
Chemical components of oil samples were identified by comparing their mass spectra with those available in the Willey7, NIST05, NIST05s spectrotest databases, using co-injection of standards and Retention Indexes (RIs). A mixture of linear n-alkanes (C7 to C40) was used for the calculation of RIs. The calculated RI for each compound was compared with values in the literature (Adams 2007ADAMS RP. 2007. Identification of essential oil components by gaschromatography/mass spectrometry, 4th ed., Carol Stream: Allured publishing corporation, 811 p.). The relative percentage of each essential oil compound was calculated as the ratio between the integral area of its respective peaks and the total area of all constituents in the sample. These data were obtained by gas chromatography analysis with a flame ionization detector (GC/FID). The compounds with a relative area greater than 1% for all oils were used to define the chemical composition of the oil.
Obtaining leaf extracts
Aqueous extract was prepared with 30g of dry leaves placed in 300 mL of distilled water heated to 100°C. After 10 minutes of infusing, the extract was filtered. The aqueous extract was not concentrated. The ethanolic extract was obtained from a mixing of 10g of dry leaves and 100 mL of 70% alcohol in a shaker for 72 hours. The solution was then filtered and concentrated in a vacuum on a rotary evaporator until it reached half of its initial volume.
Phytochemical screening of extracts
The main classes of secondary metabolites present in the aqueous and ethanolic extracts of leaves of M. vittoriana were determined by phytochemical screening using the qualitative methodologies described by Matos (2009)MATOS FJA. 2009. Introdução à fitoquímica experimental, 3ª ed., Fortaleza: Universidade Federal do Ceará. for phenols, condensed tannins, hydrolysable tannins, anthocyanins, anthocyanins, aurones, chalcones, flavonoids, xanthones, catechins, steroids, saponins, fixed acids, resins, and alkaloids; Joshi et al. (2013)JOSHI A, BHOBE M & SATTARKAR A. 2013. Phytochemical investigation of the roots of Grewia microcosm Linn. Am J Chem Pharm Res 1: 54-65. for anthraquinone glycoside; and Ayoola et al. (2008)AYOOLA G, COKER H, ADESEGUN S, ADEPOJU-BELLO A, OBAWEYA K, EZENNIA E & ATANGBAYILA T. 2008. Phytochemical screening and antioxidant activities of some selected medicinal plants used for malaria therapy in southwestern Nigeria. Trop J Pharm Res 7: 1019-1024. for cardiac glycoside and terpenoids. The presence or absence of these secondary metabolites was determined by observing the reactions carried out that indicate a positive or negative result for each investigated metabolite (Bessa et al. 2014BESSA NGF ET AL. 2014. Prospecção fitoquímica preliminar de plantas nativas do cerrado de uso popular medicinal pela comunidade rural do assentamento vale verde - Tocantins. Rev Bras Plantas Med 15: 692-707.).
Exposure of plants to essential oil and extracts
Plant growth assays were carried out using solutions of essential oil obtained from the leaves of M. vittoriana. These solutions were prepared to concentrations of 3000, 1500, 750, 375, and 187.5 µg mL-1 using the solvent dichloromethane. The aqueous and ethanolic extracts were diluted with distilled water and tested at the concentrations of 100, 50, 25, 12.5, and 6.25 (% v v-1). Essential oil concentrations were chosen based on Dutra et al. (2020)DUTRA QP, CHRIST JA, CARRIJO TT, ALVES TA, ALVES TA, MENDES LA, & PRAÇA-FONTES MM. 2020. Phytocytotoxicity of volatile constituents of essential oils from Sparattanthelium Mart. species (Hernandiaceae). Sci Rep 10:1-11. and Vasconcelos et al. (2019)VASCONCELOS LC, SANTOS ES, BERNARDES CO, FERREIRA MFS, FERREIRA A, TULER AC, CARVALHO JAM, PINHEIRO PF & PRAÇA-FONTES MM. 2019. Phytochemical analysis and effect of the essential oil of Psidium L. species on the initial development and mitotic activity of plants. Environ Sci Pollut Res 26: 26216-26228., and extract concentrations were based on the thesis of Alves T.A. (unpublished data). Glyphosate herbicide (1 mL L-1) was used as a positive control. The solvent dichloromethane used in the preparation of the essential oil solutions was also tested but showed statistically similar results to the water control, and so distilled water was chosen as a negative control (solvent data not shown). The experimental design of the plant tests was completely randomized with five replicates for each treatment. Each replicate corresponded to a Petri dish (9 cm in diameter) containing 25 seeds. The seeds of L. sativa and A. cepa were placed on filter paper moistened with 2 mL of the appropriate solution of essential oil or extract. The plates were wrapped with plastic film and placed under Biochemical Oxygen Demand (BOD) at 24 ± 2°C for the entire experimental period.
Macroscopic analysis
For macroscopic analysis, the number of germinated seeds was evaluated at 8-h intervals from 8 to 48 h for L. sativa and at 12-h intervals from 12 to 96 h for A. cepa. The germination speed index (GSI) was then calculated by the formula (N 1) × 1 + (N 1+N 2) × 1 ⁄ 2 + (N 3 + N 2) × 1 ⁄ 3…(N y - (Ny-1)) × 1 ⁄ y, where: Ny = number of seeds germinated in a given period; and y = total number of time slots. Germination percentage and root growth of L. sativa were obtained after 48 h of exposure, while aerial growth was measured after 120 h (Pinheiro et al., 2015PINHEIRO PF, COSTA AV, ALVES TA, GALTER IN, PINHEIRO CA, PEREIRA AF, OLIVEIRA CMR & FONTES MMP. 2015. Phytotoxicity and cytotoxicity of essential oil from leaves of Plectranthus amboinicus, carvacrol, and thymol in plant bioassays. J Agric Food Chem 63: 8981-8990.). Germination percentage and root growth of A. cepa were obtained after 96 h of exposure. Since seedlings of A. cepa have slower growth compared to L. sativa, and do not show green shoots within the 120 h period (Silveira et al. 2017SILVEIRA GL, GABRIELA M, LIMA F, REIS GB, PALMIERI MJ & ANDRADE-VIERIA LF. 2017. Toxic effects of environmental pollutants: Comparative investigation using Allium cepa L. and Lactuca sativa L. Chemosphere 178: 359-367.), the aerial growth of A. cepa was not evaluated. All measurements were performed with the aid of a digital calliper.
Microscopic analysis
Seeds of L. sativa and A. cepa for microscopic analysis were submitted to the same conditions, solutions and controls tested in macroscopic analysis for 48 and 96 h, respectively. Methyl methanesulfonate (MMS) (0.004 M) was also used as a positive control. MMS is a DNA alkylating agent used as a positive control in cytogenotoxic tests (Mauro et al. 2014MAURO MO, PESARINI JR, MARIN-MORALES MA, MONREAL MTFD, MONREAL ACD, MANTOVANI MS & OLIVEIRA RJ. 2014. Evaluation of the antimutagenic activity and mode of action of the fructooligosaccharide inulin in the meristematic cells of Allium cepa culture. Genet Mol Res 13: 4808-4819.). The roots of three replicates were then fixed in ethanol:acetic acid (3:1) and stored at -18 °C in a freezer. The fixing solution was changed after 10 minutes and 24 h. The previously selected roots were then washed in distilled water and hydrolyzed in 5N HCl for 18 minutes at room temperature. Slides with meristems were prepared using the crushing technique, stained with 2% (v v-1) acetic orcein and covered with a coverslip and sealed. One thousand meristematic cells were evaluated per slide, for a total of 3000 cells per treatment for L. sativa and A. cepa.
The different stages of mitotic division were observed and recorded under an optical microscope, as were any chromosomal and nuclear alterations found. The following cytotoxicity parameters were calculated according to Aragão et al. (2015)ARAGÃO FB, PALMIERI MJ, FERREIRA A, COSTA AV, QUEIROZ VT, PINHEIRO PF & ANDRADE-VIEIRA LF. 2015. Phytotoxic and cytotoxic effects of eucalyptus essential oil on lettuce (Lactuca sativa L.). Allelopath J 35: 259-272.: mitotic index (MI), percentage of nuclear alterations (NA) (condensed nucleus and micronucleus), and chromosomal alterations (CA) (lost chromosome, adherent chromosome, c-metaphase, and bridge).
Statistical analysis
Data from the macroscopic and microscopic analyses were submitted to analysis of variance (ANOVA) and the average values to Dunnett’s test at p ≤ 0.05. This test was chosen because it allows multiple comparisons of various treatments with a control and is sensitive to small differences between groups (Dunnett 1955DUNNETT CW. 1955. A Multiple Comparison Procedure for Comparing Several Treatments with a Control. J Am Stat Assoc 50: 1096-1121., McHugh 2011MCHUGH ML. 2011. Multiple comparison analysis testing in ANOVA. Biochem Medica 21: 203-209.). All statistical analyses were performed using the program R, version 4.0.0 (R Core Team 2020R CORE TEAM. 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.).
RESULTS
Yield and chemical characterization of essential oil
The essential oil of M. vittoriana had an average yield of 0.098% (m / m) compared to plant fresh weight. Gas chromatography analysis revealed the presence of 17 compounds; however, only 15 were identified, constituting 95.82% of the total relative area of the compounds present in the oil. The composition of the oil was predominantly sesquiterpenes, with 81.76% hydrogenated sesquiterpenes and 14.06% oxygenated sesquiterpenes (Table I). The main compounds found were germacrene D (21.90%), germacrene B (17.30%), and bicyclogermacrene (11.90%), all hydrogenated sesquiterpenes.
Chemical characterization and relative area (%) of the compounds present in the essential oil of the leaves of Myrcia vittoriana.
Phytochemical screening of extracts
Phytochemical screening was performed for 19 classes of chemical compounds (Table II), of which only four were not found in any of the extracts. The aqueous extract had an exclusive positive result for saponins while the ethanolic extract had exclusive positive results for alkaloids, steroids, anthocyanins, anthocyanidins, aurones, chalcones, xanthones, and cardiac glycosides. Of the remaining classes, fixed acids, catechins, simple phenols, flavonoids, hydrolysable tannins, and terpenoids showed positive results for both extracts.
Classes of secondary metabolites found in the aqueous and ethanolic extracts of Myrcia vittoriana.
Macroscopic effects
The essential oil and aqueous and ethanolic extracts of the leaves of Myrcia vittoriana significantly affected (p <0.05) the germination and growth variables of the model plants, L. sativa and A. cepa, used as receivers (Figures 2, 3, 4 and 5). The essential oil caused a significant reduction in germination rate, compared to water, only for A. cepa at concentrations of 187.5, 1500, and 3000 μg mL-1 with a decrease of 61.6% for the highest concentration (Figure 2a). For L. sativa, the highest tested concentrations of the ethanolic extract and aqueous extract [100 (% v v-1)] showed robust phytotoxic effects with reductions of 19.2 % and 84.8% in germination percentage, respectively. The extracts did not affect the germination percentage of A. cepa seeds (Figures 2b-c).

Effects of essential oil (a), aqueous extract (b) and ethanolic extracts (c) of Myrcia vittoriana on the germination (%) of Lactuca sativa and Allium cepa. Bars (mean ± SE; n = 5) with lowercase letter ‘a’ are statistically identical to water; bars followed by the letter ‘b’ are statistically identical to glyphosate by Dunnett’s test (p <0.05).

Effects of essential oil (a), aqueous extract (b) and ethanolic extracts (c) of Myrcia vittoriana on the germination speed index (GSI) of Lactuca sativa and Allium cepa. Bars (mean ± SE; n = 5) with lowercase letter ‘a’ are statistically identical to water; bars followed by the letter ‘b’ are statistically identical to glyphosate by Dunnett’s test (p <0.05).

Effects of essential oil (a), aqueous extract (b) and ethanolic extracts (c) of Myrcia vittoriana on root growth (mm) of Lactuca sativa and Allium cepa. Bars (mean ± SE; n = 5) with lowercase letter ‘a’ with lowercase letter ‘a’ are statistically identical to water; bars followed by the letter ‘b’ are statistically identical to glyphosate by Dunnett’s test (p <0.05).

Effects of essential oil (a), aqueous extract (b) and ethanolic extracts (c) of Myrcia vittoriana on the growth of the aerial part (mm) of Lactuca sativa. Bars (mean ± SE; n = 5) with lowercase letter ‘a’ are statistically identical to water; bars followed by the letter ‘b’ are statistically identical to glyphosate by Dunnett’s test (p <0.05).
The germination speed index (GSI) for A. cepa was reduced by all essential oil concentrations except for 375µg mL-1, with the most evident reduction being for the highest concentration (Figure 3a). The GSI for L. sativa was negatively affected by the aqueous extract, with significant differences from all the controls for the concentrations of 25, 50 and 100 (% v v-1) and with GSI decreasing in accordance with increasing concentrations (Figure 3b). The concentration 100 (% v v-1) of ethanolic extract also affected the GSI of L. sativa, with significant differences from all the controls. This concentration [100 (% v v-1)] was responsible for the most significant reduction in the GSI of L. sativa seeds treated with aqueous and ethanol extracts (Figure 3c). None of the tested extracts influenced the GSI of A. cepa seeds (Figures 3b-c).
The essential oil of M. vittoriana also affected root growth of the model plants (Figure 4). The concentrations of 750 to 3000 µg mL-1 inhibited root growth of A. cepa, with the inhibitory effects of these concentrations not differing significantly from the positive control glyphosate, a commercial herbicide. On the other hand, the concentrations of 375 to 1500 µg mL-1 caused an increase in the elongation of the roots of L. sativa, with a stimulatory effect comparable to that of the negative control (Figure 4a). Almost all concentrations of aqueous extract of M. vittoriana suppressed root elongation of L. sativa and A. cepa, with the exceptions being 6.25 and 12.5 (% v v-1) for A. cepa (Figure 4b). The establishment of a “a dose-dependent response” was observed for these seedlings since root growth rate declined according to increases in concentrations. The ethanolic extract, at the highest concentration (100% v v-1), also affected the growth of L. sativa roots (Figure 4c).
The growth of the aerial part of L. sativa was strongly inhibited by the essential oil of M. vittoriana, with all concentrations differing significantly from water (Figure 5a). The concentrations of 750 to 3000 µg mL-1 did not differ significantly from glyphosate. A dose-response relationship was also observed with increasing concentrations of essential oil. The highest concentration (3000 µg mL-1) had a 54.8% inhibition of the aerial growth of seedlings compared to water. With this concentration, a yellowish coloration was observed in the seedlings, along with a darkening of the roots and the presence of dark spots on the leaves (necrosis) (Figure 6).

Effects of the essential oil of Myrcia vittoriana (3000 µg mL-1) on the growth of the aerial part (mm) of Lactuca sativa. (a) Seedlings treated with distilled water with normal appearance and color. (b) Seedlings treated with 3000 µg mL-1 of essential oil showing yellowish color, dark roots and dark spots on the aerial part. Photo: Loren Cristina Vasconcelos.
Microscopic effects
The essential oil and aqueous and ethanolic extracts of M. vittoriana were cytotoxic to meristematic cells of both L. sativa and A. cepa (Figures 7a-c and 8a-c). A mitodepressive effect was observed only for cells of A. cepa treated with aqueous and ethanolic extracts (Figure 8b-c). Aqueous extract at concentrations of 6.25 and 12.5 (% v v-1) caused reductions in MI of 36.45% and 27.84%, respectively (Figure 8b). In comparison, ethanolic extract at the concentration of 100 (%v v-1) caused a 27.5% decrease compared to water (Figure 8c). Exposure to essential oil and aqueous and ethanolic extracts of M. vittoriana caused several types of chromosomal (CA) and nuclear (NA) alterations, however, the percentages of these alterations observed in A. cepa did not differ significantly from water for any of the treatments evaluated (p <0.05). The frequencies of CA for L. sativa seedlings treated with aqueous extract and ethanolic extract were significantly greater than that for water, and not significantly different from MMS (positive control) for all concentrations, except for 6.25 (% v v-1) aqueous extract (Figures 7b-c).

Meristematic cell analysis of L. sativa roots exposed to essential oil (a), aqueous extract (b) and ethanolic extract (c) of Myrcia vittoriana. Bars (mean ± SE; n = 3) with lowercase letter ‘a’ are statistically identical to water, bars followed by the letter ‘b’ are statistically identical to glyphosate, bars followed by the letter ‘c’ are statistically identical to MMS (methyl methanesulfonate) by Dunnett’s test (p <0.05). The three variables presented are MI: mitotic index; CA: chromosomal alterations; and NA: nuclear alterations.

Meristematic cell analysis of A. cepa roots exposed to essential oil (a), aqueous extract (b) and ethanolic extract (c) of Myrcia vittoriana. Bars (mean ± SE; n = 3) with lowercase letter ‘a’ are statistically identical to water, bars followed by the letter ‘b’ are statistically identical to glyphosate, bars followed by the letter ‘c’ are statistically identical to MMS (methyl methanesulfonate) by Dunnett’s test (p <0.05). The three variables presented are MI: mitotic index; CA: chromosomal alterations; and NA: nuclear alterations.
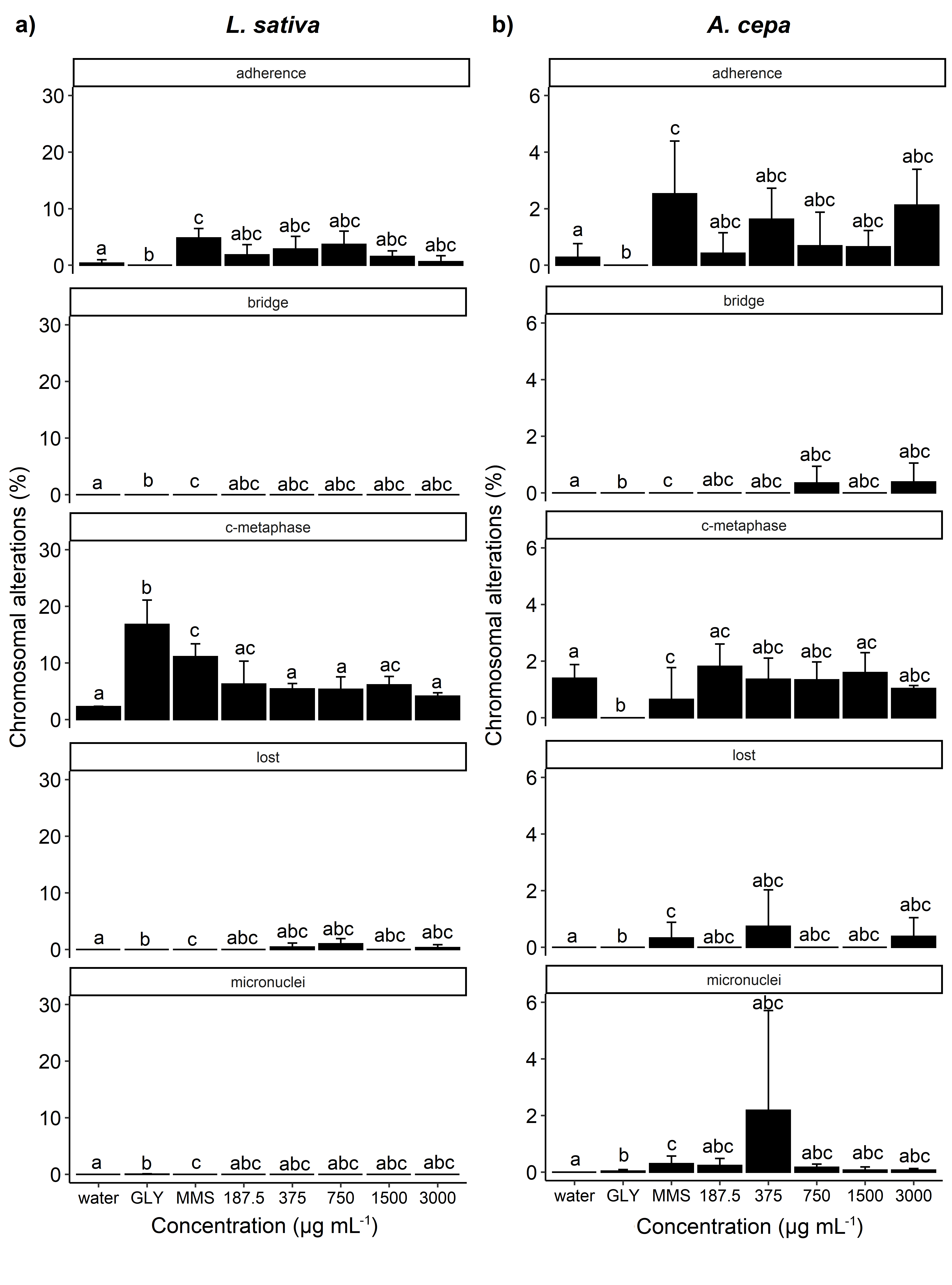
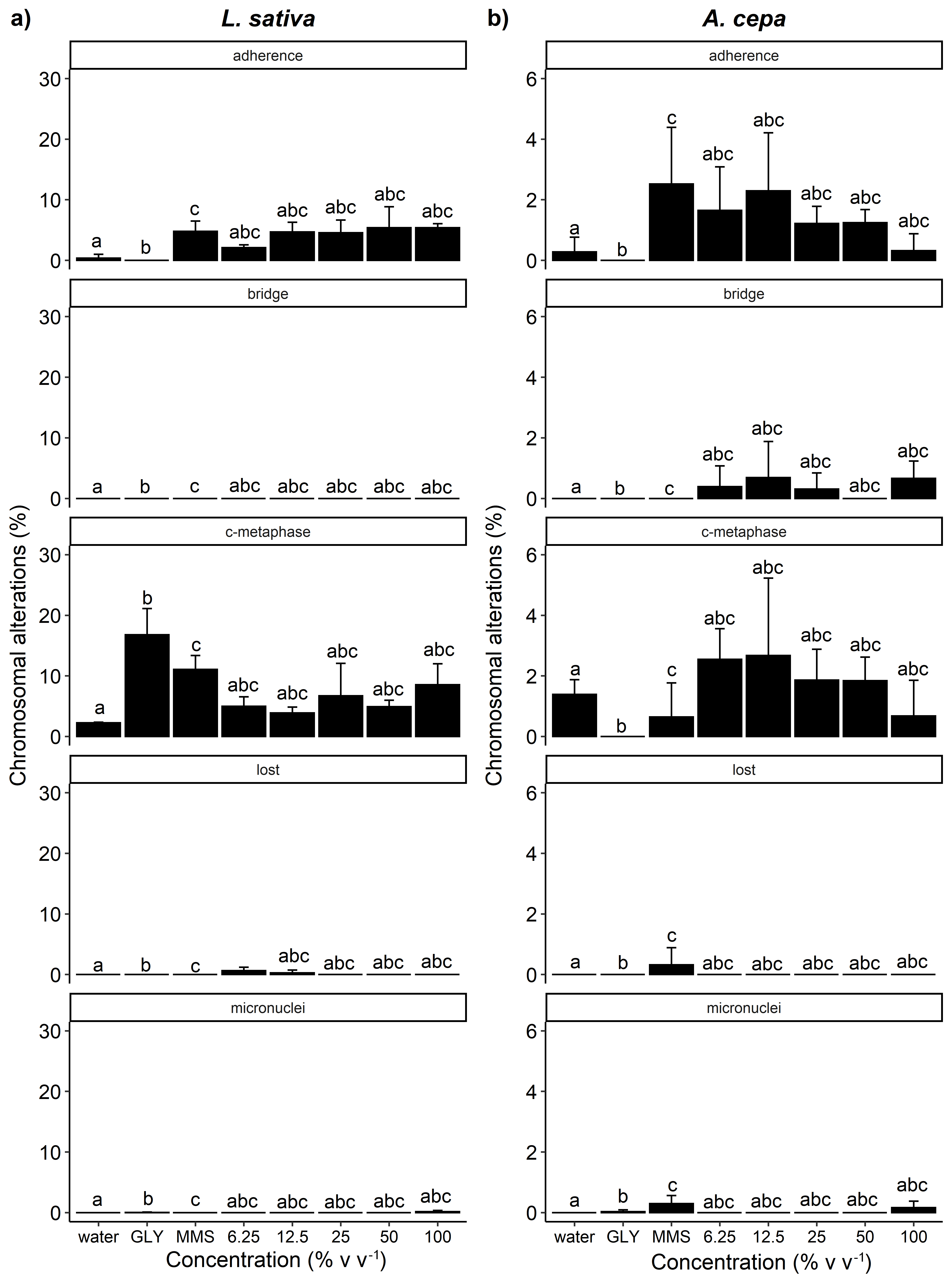
The alterations induced by the treatments were lost chromosome, adherent chromosome, c-metaphase, bridge, and micronucleus in interphase (Figure 9a-d, 10a-b, 11a-b and 12a-b). Considering the frequency of these alterations, a significant increase was observed only for L. sativa cells exposed to the ethanolic extract (Figure 12a). The adherent chromosome percentage for L. sativa increased in concentrations 12.5 and 50 (% v v-1) and did not differ significantly from MMS. The ethanolic extract concentration of 25 (% v v-1) promoted an increase in the frequency of c-metaphase, which did not differ significantly from glyphosate and MMS. The frequency of chromosomal bridge was also positively influenced by ethanolic extract at a concentration of 100 (% v v-1), when compared to positive and negative controls.

Percentage of chromosomal alterations observed in the root meristematic cells of Lactuca sativa (a) and Allium cepa (b) exposed to ethanolic extract of Myrcia vittoriana. Bars (mean ± SE; n = 3) with lowercase letter ‘a’ are statistically identical to water, bars followed by the letter ‘b’ are statistically identical to GLY (glyphosate), bars followed by the letter ‘c’ are statistically identical to MMS (methyl methanesulfonate) by Dunnett’s test (p <0.05). The five variables shown are adherent chromosome, bridge in anaphase and telophase, c-metaphase, lost chromosome, and micronucleus in interphase.

Main alterations found in Lactuca sativa root meristematic cells exposed to essential oil, aqueous extract and ethanolic extract of Myrcia vittoriana. (a) Metaphase with adherent chromosome. (b) Interphase with micronucleus. (c) C-metaphase. (d) Chromosomal bridge. Bar = 10 µm.
Other alterations also showed considerable increases in relation to the controls, although these increases were not significant. Meristematic cells of L. sativa and A. cepa with adherent chromosome were observed in all treatments, except for the negative control. Likewise, c-metaphase was also observed in all treatments (Figures 10, 11 and 12), except for A. cepa cells exposed to essential oil. Lost chromosome was induced in L. sativa cells by all treatments evaluated (Figures 10a, 11a and 12a), while this same alteration was not observed in any of the tested controls. The essential oil of M. vittoriana also induced lost chromosome in A. cepa at concentrations of 350 and 3000 µg mL-1, while this alteration was only observed in the MMS control (Figure 10b). Chromosomal bridge and micronucleus in interphase were not observed in the negative control of any of the plant models, however, they were observed in the evaluated treatments. Chromosomal bridge was observed for A. cepa in all treatments tested, but most frequently for aqueous extract (Figure 11b). Micronucleus in interphase was also identified in meristematic cells of A. cepa exposed to treatments, with essential oil being the main cause of this alteration (Figure 10b). Aqueous extract also produced micronucleus in interphase in all plant models, but only at the concentration of 100 (% v v-1) (Figure 11a-b). Ethanolic extract at the concentration of 100 (% v v-1) induced chromosomal bridge in anaphase cells of L. sativa, while the same concentration of aqueous extract also caused the appearance of micronucleus in interphase (Figure 12a). Micronucleus in interphase was also observed in L. sativa cells treated with ethanolic extract at concentrations of 12.5 and 100 (% v v-1) (Figure 12b).

Percentage of chromosomal alterations observed in the root meristematic cells of Lactuca sativa (a) and Allium cepa (b) exposed to essential oil of Myrcia vittoriana. Bars (mean ± SE; n = 3) with lowercase letter ‘a’ are statistically identical to water, bars followed by the letter ‘b’ are statistically identical to GLY (glyphosate), bars followed by the letter ‘c’ are statistically identical to MMS (methyl methanesulfonate) by Dunnett’s test (p <0.05). The five variables shown are adherent chromosome, bridge in anaphase and telophase, c-metaphase, lost chromosome, and micronucleus in interphase.

Percentage of chromosomal alterations observed in the root meristematic cells of Lactuca sativa (a) and Allium cepa (b) exposed to aqueous extract of Myrcia vittoriana. Bars (mean ± SE; n = 3) with lowercase letter ‘a’ are statistically identical to water, bars followed by the letter ‘b’ are statistically identical to GLY (glyphosate), bars followed by the letter ‘c’ are statistically identical to MMS (methyl methanesulfonate) by Dunnett’s test (p <0.05). The five variables shown are adherent chromosome, bridge in anaphase and telophase, c-metaphase, lost chromosome, and micronucleus in interphase.
DISCUSSION
The essential oil of M. vittoriana was composed entirely of sesquiterpenes, with the hydrocarbon sesquiterpenes germacrene D, germacrene B and bicyclogermacrene being the major compounds, as has been described for other species of Myrcia [e.g., M. tomentosa (Aubl.) DC., M. sylvatica (G. Mey.) DC., M. cuprea (O. Berg) Kiaersk., M. pubipetala Miq., M. lajeana D. Legrand and M. alagoensis O. Berg; Franco et al. 2021FRANCO CDJP, FERREIRA OO, MORAES AAB, VARELA ELP, NASCIMENTO LDD, PERCÁRIO S, OLIVEIRA MS & ANDRADE EHDA. 2021. Chemical composition and antioxidant activity of essential oils from Eugenia patrisii Vahl, E. punicifolia (Kunth) DC., and Myrcia tomentosa (Aubl.) DC., leaf of family Myrtaceae. Molecules 26: 3292., Jerônimo et al. 2021JERÔNIMO LB, COSTA JS, PINTO LC, MONTENEGRO RC, SETZER WN, MOURÃO RHV, SILVA JKR, MAIA JGS & FIGUEIREDO PLB. 2021. Antioxidant and cytotoxic activities of Myrtaceae essential oils rich in terpenoids from Brazil. Nat Prod Commun 16: 1934578X21996156., Zoghbi et al. 2003ZOGHBI MDGB, ANDRADE EHA, SILVA MHL, CARREIRA LMM & MAIA JGS. 2003. Essential oils from three Myrcia species. Flavour Fragr J 18: 421-424., Limberger et al. 2004LIMBERGER RP, SOBRAL M, HENRIQUES AT, MENUT C & BESSIÈRE JM. 2004. Óleos voláteis de espécies de Myrcia nativas do Rio Grande do Sul. Quim Nova 27: 916-919., Silva et al. 2013SILVA ADN, UETANABARO APT & LUCCHESE AM. 2013. Chemical composition and antibacterial activity of essential oils from Myrcia alagoensis (Myrtaceae). Nat Prod Commun 8: 1934578X1300800235.]. Germacrene D is considered a bioenergetic precursor to many sesquiterpenes (Bülow & König 2000BÜLOW N & KÖNIG WA. 2000. The role of germacrene D as a precursor in sesquiterpene biosynthesis: Investigations of acid catalyzed, photochemically and thermally induced rearrangements. Phytochemistry 55: 141-168.) and has been reported for its numerous biological activities. Studies have associated the presence of germacrene D with the phytotoxic (El Ayeb-Zakhama et al. 2016EL AYEB-ZAKHAMA A, SAKKA-ROUIS L, BERGAOUI A, FLAMINI G, JANNET HB & HARZALLAH-SKHIRI F. 2016. Chemical composition and allelopathic potential of essential oils from Tipuana tipu (Benth.) Kuntze cultivated in Tunisia. Chem Biodivers 13: 309-318., Scrivanti & Anton 2019SCRIVANTI LR & ANTON AM. 2019. Allelopathic effect of endemic South American Bothriochloa species (Poaceae: Andropogoneae). J Essent Oil Res 31: 247-254.), antifungal (Lawson et al. 2020LAWSON SK, SHARP LG, POWERS CN, MCFEETERS RL, SATYAL P & SETZER WN. 2020. Volatile compositions and antifungal activities of native american medicinal plants: focus on the Asteraceae. Plants 9: 126.), insecticidal (Birkett et al. 2008BIRKETT MA ET AL. 2008. Antiectoparasitic activity of the gum resin, gum haggar, from the East African plant, Commiphora holtziana. Phytochemistry 69: 1710-1715.) and antibacterial (Salazar et al. 2018SALAZAR GJT ET AL. 2018. Phytochemical characterization of the Baccharis dracunculifolia DC (Asteraceae) essential oil and antibacterial activity evaluation. In Crops Prod 122: 591-595.) actions of essential oils. Eom et al. (2006)EOM SH, YANG HS & WESTON LA. 2006. An evaluation of the allelopathic potential of selected perennial groundcovers: foliar volatiles of catmint (Nepeta× faassenii) inhibit seedling growth. J Chem Ecol 32: 1835-1848. associated weed growth suppression with volatile from Catmint (Nepeta x faassenii), which includes germacrene D and B among other compounds.
Phytochemical screening of the studied species confirmed the presence of fixed acid metabolites, simple phenols, flavonoids, catechins, hydrolysable tannins and terpenoids in both aqueous and ethanolic extracts, as reported by previous studies within Myrtaceae (e.g., Kich et al. 2017KICH DM ET AL. 2017. Lymphocyte genotoxicity and protective effect of Calyptranthes tricona (Myrtaceae) against H2O2-induced cell death in MCF-7 cells. Mol Cell Biochem 424: 35-43., Raj et al. 2020RAJ A, MENON V & SHARMA N. 2020. Phytochemical screening, antimicrobial, antioxidant and cytotoxic potential of different extracts of Psidium guajava leaves. Vegetos 33: 750-758., Rocha et al. 2021ROCHA T, MARQUES EJ, NASCIMENTO CM, SOUZA RRM, SILVA MC, NETA LCS & VALE VLC. 2021. Chemical and biological profile of Psidium bahianum landrum & funch (Myrtaceae). Rev Bras Bot: 1-11.). These compounds have been reported for their ability to reduce germination and growth of different plant species (John & Sarada 2012JOHN J & SARADA S. 2012. Role of phenolics in allelopathic interactions. Allelopath J 29: 215-230.). In addition, phenolic compounds are recognized for altering various physiological and biochemical processes, such as cell membrane permeability, photosynthesis, cell respiration, and lipid peroxidation, in addition to interfering in cell division and, consequently, in plant growth and development (John & Sarada 2012JOHN J & SARADA S. 2012. Role of phenolics in allelopathic interactions. Allelopath J 29: 215-230.).
The essential oil of M. vittoriana reduced all germination parameters evaluated (germination percentage and GSI) for A. cepa, as well as root growth, whereas only aerial growth of L. sativa was affected, with no effects on germination. Inhibitory effects were confirmed for germination percentage, GSI and root growth for L. sativa, whereas only root growth was negatively affected by aqueous extract for A. cepa. According to Blum (1999BLUM U. 1999. Designing laboratory plant debris-soil bioassays: some reflections. In: Inderjit et al. (Eds), Principles and practices in plant ecology: allelochemical interactions, Boca Raton: CRC press, Flórida, USA, p. 17-24.), seedling emergence and growth are the most sensitive phases in the ontogenesis of an individual plant. Growth reduction indicates inhibition of nutrient absorption, cell elongation and cell division, and causes cell death (Freitas et al. 2016FREITAS AS, CUNHA IMF, ANDRADE-VIEIRA LF & TECHIO VH. 2016. Effect of SPL (Spent Pot Liner) and its main components on root growth, mitotic activity and phosphorylation of Histone H3 in Lactuca sativa L. Ecotoxicol Environ Saf 124: 426-434.).
The growth of the aerial part of L. sativa was significantly affected by all essential oil concentrations in a dose-dependent manner, with the inhibition caused by most concentrations not differing significantly from glyphosate herbicide. The highest essential oil concentration caused depigmentation of leaves, darkening of roots, and necrosis of seedlings. Loss of pigmentation and necrosis towards the tip may occur in leaves due to the degradation of pigment molecules by oxidative damage, with serious effects on photosynthesis (Hussain et al. 2017HUSSAIN MI, BORAH P, HUSSAIN I, SHARMA RK & KALITA MC. 2017. Necrotic enteritis by beta2toxin-producing Clostridium perfringens in doom pigs of assam, India. Int J Curr Microbiol Appl Sci 6: 1872-1876., Niu et al. 2013NIU Y, WANG Y, LI P, ZHANG F, LIU H & ZHENG G. 2013. Drought stress induces oxidative stress and the antioxidant defense system in ascorbate-deficient vtc1 mutants of Arabidopsis thaliana. Acta Physiol Plant 35: 1189-1200.). In this sense, oxidative stress may have caused the suppression of shoot growth by targeting and allocating the few photoassimilates produced to the roots as an adaptive strategy for seedling survival by increasing the water and nutrient absorption capacity of the root. This would explain the induction of root growth to the detriment of aerial growth for L. sativa seedlings treated with essential oil. Therefore, it is possible that sesquiterpenes and the compounds found in the extracts are mostly responsible for the phytotoxic activity of M. vittoriana.
The present study demonstrated the cytotoxicity of the essential oil and aqueous and ethanolic extracts of M. vittoriana by documenting a decrease in the mitotic index and detecting chromosomal and nuclear alterations in the cell cycle of L. sativa and A. cepa. The mitodepressive effect caused by the aqueous and ethanolic extracts on the meristematic cells of A. cepa may have been one of the causes of root growth inhibition since root elongation depends on normal cell division. Soltys et al. (2011)SOLTYS D, RUDZIŃSKA-LANGWALD A, KUREK W, GNIAZDOWSKA A, SLIWINSKA E & BOGATEK R. 2011. Cyanamide mode of action during inhibition of onion (Allium cepa L.) root growth involves disturbances in cell division and cytoskeleton formation. Planta 234: 609-621. explained that reduced root growth caused by the allelochemical cyanamide is due to inhibition of the mitotic cycle and cell proliferation, as well as to the presence of alterations in the arrangement of the cytoskeleton in A. cepa root tip cells. According to this study, newly formed cells are the only way to elongate organs by increasing the number of cells in dimensions.
Aqueous and ethanolic extracts of M. vittoriana also reduced root growth of L. sativa. However, cytological observations revealed they did not cause a reduction in MI. The non-reduction of MI observed here can be explained by the presence of saponins in the aqueous extract and hydrolysable tannins in both the aqueous and ethanolic extracts of M. vittoriana. Luber et al. (2015)LUBER J, PALMIERI MJ, BOTELHO CM, RINALDO D & ANDRADE-VIEIRA LF. 2015. Investigation on the effects of guava (Psidium guajava L.) infusions on germination, root tips and meristematic cells of Latuca Sativa. An Acad Bras Cienc 87: 903-913. studied the phytotoxic effects of Psidium guajava L. infusion and observed reduced root growth without a decrease in MI for L. sativa, which they associated with the presence of tannins in infusions of guava cultivars. According to the authors, the roots started to grow after germination, without cell proliferation being affected by the infusions. Tannins and saponins act in defense against herbivores and pathogens (Ferreira & Aquila 2000FERREIRA AG & AQUILA EMEA. 2000. Alelopatia: Uma área emergente da ecofisiologia. Rev Bras Fisiol Veg 12: 175-204., Rice 1984RICE EL. 1984. Allelopathy, 2nd ed., Orlando: Academic Press, 423 p.). According to Marchaim et al. (1974)MARCHAIM U, WERKER E & THOMAS WDE. 1974. Changes in the anatomy of cotton seed coats caused by lucerne saponins. Bot Gaz 135: 139-146., saponine can affect germination and the initial growth of seedlings by reducing the respiration rate of seeds by making it difficult for oxygen to diffuse through the seed coat. Castro & Ferreira (2001)CASTRO H & FERREIRA F. 2001. Contribuição ao estudo das plantas medicinais (Baccharis genistelloides), Viçosa: Ed. UFV. reported the ability of hydrolysable tannins to act as inhibitors of plant germination and growth, as well as nitrifying and nitrogen-fixing bacteria. From this perspective, the reduced growth of L. sativa roots exposed to aqueous and ethanolic extracts of M. vittoriana may have been due to the action of saponins and tannins on germination, without cell proliferation being affected.
The increased frequency of chromosomal alterations in relation to the water control indicates genotoxic action by the compounds present in the essential oil and aqueous and ethanolic extracts of M. vittoriana. These alterations occur as the result of changes in the structure and/or the number of chromosomes (Leme & Marin-Morales 2009LEME DM & MARIN-MORALES MA. 2009. Allium cepa test in environmental monitoring: A review on its application. Mutat Res 682: 71-81.). Clastogenic compounds induce breakages in DNA while aneugenic compounds interfere with the mitotic spindle, leading to errors during chromosomal segregation, resulting in the elimination of genetic material and the formation of polyploid cells (Fernandes et al. 2007FERNANDES TCC, MAZZEO DEC & MARIN-MORALES MA. 2007. Mechanism of micronuclei formation in polyploidizated cells of Allium cepa exposed to trifluralin herbicide. Pestic Biochem Physiol 88: 252-259.). Microscopic analysis indicated that both essential oil and aqueous and ethanol extracts have aneugenic and clastogenic modes of action.
Adherent chromosome and c-metaphase were the most frequent chromosomal alterations in both L. sativa and A. cepa. These abnormalities are caused by aneugenic agents. According to El-Ghamery et al. (2003)EL-GHAMERY AA, EL-KHOLY MA & EL-YOUSSER MAA. 2003. Evaluation of cytological effects of Zn2+ in relation to germination and root growth of Nigella sativa L. and Triticum aestivum L. Mutat Res - Genet Toxicol Environ Mutagen 537: 29-41., adherence can be related to the later appearance of chromosomal breaks and the formation of bridges between chromatids, corroborating the results found in the present study. All treatments of A. cepa induced chromosomal bridge formation in anaphase. The ethanolic extract also caused chromosomal bridge in L. sativa. The establishment of bridges is a clastogenic alteration that occurs due to the loss of telomeres through breaks in the ends of chromosomes followed by the union of chromatids (Leme & Marin-Morales 2009LEME DM & MARIN-MORALES MA. 2009. Allium cepa test in environmental monitoring: A review on its application. Mutat Res 682: 71-81., Matsumoto et al. 2006MATSUMOTO ST, MANTOVANI MS, MALAGUTTII MIA, DIAS AL, FONSECA IC & MARIN-MORALES MA. 2006. Genotoxicity and mutagenicity of water contaminated with tannery effluents as evaluated by the micronucleus test and comet assay using the fish Oreochromis niloticus and chromosome aberrations in onion root-tips. Genet Mol Biol 29: 148-158.).
C-metaphase and lost chromosome are aneugenic changes related to the action of agents on the fibers of the mitotic spindle during metaphase (Fernandes et al. 2007FERNANDES TCC, MAZZEO DEC & MARIN-MORALES MA. 2007. Mechanism of micronuclei formation in polyploidizated cells of Allium cepa exposed to trifluralin herbicide. Pestic Biochem Physiol 88: 252-259., Leme & Marin-Morales 2009LEME DM & MARIN-MORALES MA. 2009. Allium cepa test in environmental monitoring: A review on its application. Mutat Res 682: 71-81.). Lost chromosome results from the lack of orientation of the spindle fibers during metaphase (Freitas et al. 2016FREITAS AS, CUNHA IMF, ANDRADE-VIEIRA LF & TECHIO VH. 2016. Effect of SPL (Spent Pot Liner) and its main components on root growth, mitotic activity and phosphorylation of Histone H3 in Lactuca sativa L. Ecotoxicol Environ Saf 124: 426-434.). Thus, the induction of c-metaphase and lost chromosome in the model plants may have occurred due to the action of aneugenic substances present in the essential oil and extracts of M. vittoriana leaves on the proteins that form or regulate the mitotic spindle. Micronucleus was also induced by treatments, with the exception being for essential oil in the meristematic cells of L. sativa. In the present study, micronucleus may have arisen from entire chromosomes being lost during anaphase, which may be indicative of a mutagenic effect.
CONCLUSIONS
The phytotoxic and cytotoxic effects of Myrcia vittoriana were confirmed for the plant models L. sativa and A. cepa. The sesquiterpenes, with an emphasis on germacrene D (identified in the essential oil), and the phenolic compounds (tannins, catechins, and saponins) identified in the aqueous and ethanolic extracts contributed to the phytotoxic, cytotoxic, genotoxic, and mutagenic activity observed in the treatments. Future studies should focus on the identification of polar or moderately polar constituents in fresh and dried leaf tissues of M. vittoriana and test them on native species and under field conditions. In this way, it will be possible to elucidate how these compounds of M. vittoriana are released into the environment where they affect neighboring plants in the communities. Additional evaluation of the ethanolic and aqueous extracts using mass spectrometry is also needed.
ACKNOWLEDGMENTS
TT Carrijo, ML Garbin and M.M.Praça-Fontes thank Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the research grant “Bolsa de Produtividade em Pesquisa”. The authors are also greatful to Fundação de Amparo à Pesquisa e Inovação do Espírito Santo (FAPES) for the research grant “Taxa de Pesquisa” (T.T. Carrijo #297/2021 and M.L. Garbin #299/2021), “Universal” grant (M.M. Praça-Fontes #484/2021, 2021-ZK8PJ), and “Cooperação CAPES/FAPES – PDPG” grant. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior -Brasil (CAPES) - Finance Code 001. We also thank to the Universidade Federal do Espírito Santo (UFES) for logistical support. The authors declare that they have no knowledge of competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
REFERENCES
- ADAMS RP. 2007. Identification of essential oil components by gaschromatography/mass spectrometry, 4th ed., Carol Stream: Allured publishing corporation, 811 p.
- ARAGÃO FB, PALMIERI MJ, FERREIRA A, COSTA AV, QUEIROZ VT, PINHEIRO PF & ANDRADE-VIEIRA LF. 2015. Phytotoxic and cytotoxic effects of eucalyptus essential oil on lettuce (Lactuca sativa L.). Allelopath J 35: 259-272.
- AYOOLA G, COKER H, ADESEGUN S, ADEPOJU-BELLO A, OBAWEYA K, EZENNIA E & ATANGBAYILA T. 2008. Phytochemical screening and antioxidant activities of some selected medicinal plants used for malaria therapy in southwestern Nigeria. Trop J Pharm Res 7: 1019-1024.
- BEECH E, RIVERS M, OLDFIELD S & SMITH PP. 2017. GlobalTreeSearch: The first complete global database of tree species and country distributions. J Sustain For 36: 454-489.
- BESSA NGF ET AL. 2014. Prospecção fitoquímica preliminar de plantas nativas do cerrado de uso popular medicinal pela comunidade rural do assentamento vale verde - Tocantins. Rev Bras Plantas Med 15: 692-707.
- BIRKETT MA ET AL. 2008. Antiectoparasitic activity of the gum resin, gum haggar, from the East African plant, Commiphora holtziana. Phytochemistry 69: 1710-1715.
- BLANCO JA. 2007. The representation of allelopathy in ecosystem-level forest models. Ecol Modell 209: 65-77.
- BLUM U. 1999. Designing laboratory plant debris-soil bioassays: some reflections. In: Inderjit et al. (Eds), Principles and practices in plant ecology: allelochemical interactions, Boca Raton: CRC press, Flórida, USA, p. 17-24.
- BÜLOW N & KÖNIG WA. 2000. The role of germacrene D as a precursor in sesquiterpene biosynthesis: Investigations of acid catalyzed, photochemically and thermally induced rearrangements. Phytochemistry 55: 141-168.
- CALLAWAY RM & RIDENOUR WM. 2004. Novel weapons: Invasive success and the evolution of increased competitive ability. Front Ecol Environ 2: 436-443.
- CAPUTO L, SMERIGLIO A, TROMBETTA D, CORNARA L, TREVENA G, VALUSSI M, FRATIANNI F, FEO DV & NAZZARO F. 2020. Chemical composition and biological activities of the essential oils of Leptospermum petersonii and Eucalyptus gunnii. Front Microbiol 11: 1-15.
- CARVALHO MSS, ANDRADE-VIEIRA LF, SANTOS FED, CORREA FF, GRAÇAS MCD & VILELA LR. 2019. Allelopathic potential and phytochemical screening of ethanolic extracts from five species of Amaranthus spp. in the plant model Lactuca sativa. Sci Hortic 245: 90-98.
- CASTRO H & FERREIRA F. 2001. Contribuição ao estudo das plantas medicinais (Baccharis genistelloides), Viçosa: Ed. UFV.
- CHENGXU W, MINGXING Z, XUHUI C & BO Q. 2011. Review on allelopathy of exotic invasive plants. Procedia Eng 18: 240-246.
- CHOU CH. 1999. Roles of allelopathy in plant biodiversity and sustainable agriculture. Crit Rev Plant Sci 18: 609-636.
- DUNNETT CW. 1955. A Multiple Comparison Procedure for Comparing Several Treatments with a Control. J Am Stat Assoc 50: 1096-1121.
- DUTRA QP, CHRIST JA, CARRIJO TT, ALVES TA, ALVES TA, MENDES LA, & PRAÇA-FONTES MM. 2020. Phytocytotoxicity of volatile constituents of essential oils from Sparattanthelium Mart. species (Hernandiaceae). Sci Rep 10:1-11.
- EL AYEB-ZAKHAMA A, SAKKA-ROUIS L, BERGAOUI A, FLAMINI G, JANNET HB & HARZALLAH-SKHIRI F. 2016. Chemical composition and allelopathic potential of essential oils from Tipuana tipu (Benth.) Kuntze cultivated in Tunisia. Chem Biodivers 13: 309-318.
- EL-GHAMERY AA, EL-KHOLY MA & EL-YOUSSER MAA. 2003. Evaluation of cytological effects of Zn2+ in relation to germination and root growth of Nigella sativa L. and Triticum aestivum L. Mutat Res - Genet Toxicol Environ Mutagen 537: 29-41.
- EOM SH, YANG HS & WESTON LA. 2006. An evaluation of the allelopathic potential of selected perennial groundcovers: foliar volatiles of catmint (Nepeta× faassenii) inhibit seedling growth. J Chem Ecol 32: 1835-1848.
- FERNANDES TCC, MAZZEO DEC & MARIN-MORALES MA. 2007. Mechanism of micronuclei formation in polyploidizated cells of Allium cepa exposed to trifluralin herbicide. Pestic Biochem Physiol 88: 252-259.
- FERNANDEZ C, SANTONJA M, GROS R, MONNIER Y, CHOMEL M, BALDY V & BOUSQUET-MÉLOU A. 2013. Allelochemicals of Pinus halepensis as drivers of biodiversity in Mediterranean open mosaic habitats during the colonization stage of secondary succession. J Chem Ecol 39: 298-311.
- FERREIRA AG & AQUILA EMEA. 2000. Alelopatia: Uma área emergente da ecofisiologia. Rev Bras Fisiol Veg 12: 175-204.
- FRANCO CDJP, FERREIRA OO, MORAES AAB, VARELA ELP, NASCIMENTO LDD, PERCÁRIO S, OLIVEIRA MS & ANDRADE EHDA. 2021. Chemical composition and antioxidant activity of essential oils from Eugenia patrisii Vahl, E. punicifolia (Kunth) DC., and Myrcia tomentosa (Aubl.) DC., leaf of family Myrtaceae. Molecules 26: 3292.
- FRANCO DM, SALDANHA LL, SILVA EM, NOGUEIRA F, DOKKEDAL AL, SANTOS C & ALMEIDA LFR. 2015. Effects of leaf extracts of Myrcia guianensis (Aubl.) DC. on growth and gene expression during root development of Sorghum bicolor (L.) Moench. Allelopath J 35: 237-248.
- FREITAS AS, CUNHA IMF, ANDRADE-VIEIRA LF & TECHIO VH. 2016. Effect of SPL (Spent Pot Liner) and its main components on root growth, mitotic activity and phosphorylation of Histone H3 in Lactuca sativa L. Ecotoxicol Environ Saf 124: 426-434.
- GARBIN ML, GUIDONI-MARTINS KG, HOLLUNDER RK, MARIOTTE P, SCARANO FR & CARRIJO TT. 2016. Spatial segregation of subordinate species is not controlled by the dominant species in a tropical coastal plant community. Perspect Plant Ecol Evol Syst 18: 23-32.
- HUSSAIN MI, BORAH P, HUSSAIN I, SHARMA RK & KALITA MC. 2017. Necrotic enteritis by beta2toxin-producing Clostridium perfringens in doom pigs of assam, India. Int J Curr Microbiol Appl Sci 6: 1872-1876.
- IMATOMI M, NOVAES P & GUALTIERI SCJ. 2013a. Interspecific variation in the allelopathic potential of the family Myrtaceae. Acta Bot Brasilica 27: 54-61.
- IMATOMI M, NOVAES P, MATOS AP, GUALTIERI SCJ, MOLINILLO JMG, LACRET R, VARELA RM & MACÍAS FA. 2013b. Phytotoxic effect of bioactive compounds isolated from Myrcia tomentosa (Myrtaceae) leaves. Biochem Syst Ecol 46: 29-35.
- IMATOMI M, NOVAES P, MIRANDA MAFM & GUALTIERI SCJ. 2015. Phytotoxic effects of aqueous leaf extracts of four Myrtaceae species on three weeds. Acta Sci Agron 37: 241-248.
- INDERJIT. 2005. Soil microorganisms: An important determinant of allelopathic activity. Plant Soil 274: 227-236.
- INDERJIT & DUKE SO. 2003. Ecophysiological aspects of allelopathy. Planta 217: 529-539.
- INDERJIT, WARDLE DA, KARBAN R & CALLAWAY RM. 2011. The ecosystem and evolutionary contexts of allelopathy. Trends Ecol Evol 26: 655-662.
- JERÔNIMO LB, COSTA JS, PINTO LC, MONTENEGRO RC, SETZER WN, MOURÃO RHV, SILVA JKR, MAIA JGS & FIGUEIREDO PLB. 2021. Antioxidant and cytotoxic activities of Myrtaceae essential oils rich in terpenoids from Brazil. Nat Prod Commun 16: 1934578X21996156.
- JOHN J & SARADA S. 2012. Role of phenolics in allelopathic interactions. Allelopath J 29: 215-230.
- JOSHI A, BHOBE M & SATTARKAR A. 2013. Phytochemical investigation of the roots of Grewia microcosm Linn. Am J Chem Pharm Res 1: 54-65.
- KAPOOR D, RINZIM, TIWARI A, SEHGAL A, LANDI M, BRESTIC M & SHARMA A. 2019. Exploiting the allelopathic potential of aqueous leaf extracts of Artemisia absinthium and Psidium guajava against parthenium hysterophorus, a widespread weed in India. Plants 8: 1-13.
- KATO-NOGUCHI H, KIMURA F, OHNO O & SUENAGA K. 2017. Involvement of allelopathy in inhibition of understory growth in red pine forests. J Plant Physiol 218: 66-73.
- KICH DM ET AL. 2017. Lymphocyte genotoxicity and protective effect of Calyptranthes tricona (Myrtaceae) against H2O2-induced cell death in MCF-7 cells. Mol Cell Biochem 424: 35-43.
- LAWSON SK, SHARP LG, POWERS CN, MCFEETERS RL, SATYAL P & SETZER WN. 2020. Volatile compositions and antifungal activities of native american medicinal plants: focus on the Asteraceae. Plants 9: 126.
- LEME DM & MARIN-MORALES MA. 2009. Allium cepa test in environmental monitoring: A review on its application. Mutat Res 682: 71-81.
- LIMBERGER RP, SOBRAL M, HENRIQUES AT, MENUT C & BESSIÈRE JM. 2004. Óleos voláteis de espécies de Myrcia nativas do Rio Grande do Sul. Quim Nova 27: 916-919.
- LUBER J, PALMIERI MJ, BOTELHO CM, RINALDO D & ANDRADE-VIEIRA LF. 2015. Investigation on the effects of guava (Psidium guajava L.) infusions on germination, root tips and meristematic cells of Latuca Sativa. An Acad Bras Cienc 87: 903-913.
- MARCHAIM U, WERKER E & THOMAS WDE. 1974. Changes in the anatomy of cotton seed coats caused by lucerne saponins. Bot Gaz 135: 139-146.
- MATOS FJA. 2009. Introdução à fitoquímica experimental, 3ª ed., Fortaleza: Universidade Federal do Ceará.
- MATSUMOTO ST, MANTOVANI MS, MALAGUTTII MIA, DIAS AL, FONSECA IC & MARIN-MORALES MA. 2006. Genotoxicity and mutagenicity of water contaminated with tannery effluents as evaluated by the micronucleus test and comet assay using the fish Oreochromis niloticus and chromosome aberrations in onion root-tips. Genet Mol Biol 29: 148-158.
- MAURO MO, PESARINI JR, MARIN-MORALES MA, MONREAL MTFD, MONREAL ACD, MANTOVANI MS & OLIVEIRA RJ. 2014. Evaluation of the antimutagenic activity and mode of action of the fructooligosaccharide inulin in the meristematic cells of Allium cepa culture. Genet Mol Res 13: 4808-4819.
- MCHUGH ML. 2011. Multiple comparison analysis testing in ANOVA. Biochem Medica 21: 203-209.
- NIU Y, WANG Y, LI P, ZHANG F, LIU H & ZHENG G. 2013. Drought stress induces oxidative stress and the antioxidant defense system in ascorbate-deficient vtc1 mutants of Arabidopsis thaliana. Acta Physiol Plant 35: 1189-1200.
- OOKA JK & OWENS DK. 2018. Allelopathy in tropical and subtropical species. Phytochem Rev 17: 1225-1237.
- PINHEIRO PF, COSTA AV, ALVES TA, GALTER IN, PINHEIRO CA, PEREIRA AF, OLIVEIRA CMR & FONTES MMP. 2015. Phytotoxicity and cytotoxicity of essential oil from leaves of Plectranthus amboinicus, carvacrol, and thymol in plant bioassays. J Agric Food Chem 63: 8981-8990.
- PINHEIRO PF, QUEIROZ VT, RONDELLI VM, COSTA AV, MARCELINO TP & PRATISSOLI D. 2013. Insecticidal activity of citronella grass essential oil on Frankliniella schultzei and Myzus persicae. Ciênc Agrotec 37: 138-144.
- QIN F, LIU S & YU S. 2018. Effects of allelopathy and competition for water and nutrients on survival and growth of tree species in Eucalyptus urophylla plantations. For Ecol Manage 424: 387-395.
- R CORE TEAM. 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
- RAJ A, MENON V & SHARMA N. 2020. Phytochemical screening, antimicrobial, antioxidant and cytotoxic potential of different extracts of Psidium guajava leaves. Vegetos 33: 750-758.
- REIGOSA MJ, SÁNCHEZ-MOREIRAS A & GONZÁLEZ L. 1999. Ecophysiological approach in allelopathy. Crit Rev Plant Sci 18: 577-608.
- RICE EL. 1984. Allelopathy, 2nd ed., Orlando: Academic Press, 423 p.
- ROCHA T, MARQUES EJ, NASCIMENTO CM, SOUZA RRM, SILVA MC, NETA LCS & VALE VLC. 2021. Chemical and biological profile of Psidium bahianum landrum & funch (Myrtaceae). Rev Bras Bot: 1-11.
- RUWANZA S, GAERTNER M, ESLER KJ & RICHARDSON DM. 2015. Allelopathic effects of invasive Eucalyptus camaldulensis on germination and early growth of four native species in the Western Cape, South Africa. South For 77: 91-105.
- SALAZAR GJT ET AL. 2018. Phytochemical characterization of the Baccharis dracunculifolia DC (Asteraceae) essential oil and antibacterial activity evaluation. In Crops Prod 122: 591-595.
- SCRIVANTI LR & ANTON AM. 2019. Allelopathic effect of endemic South American Bothriochloa species (Poaceae: Andropogoneae). J Essent Oil Res 31: 247-254.
- SILVA ADN, UETANABARO APT & LUCCHESE AM. 2013. Chemical composition and antibacterial activity of essential oils from Myrcia alagoensis (Myrtaceae). Nat Prod Commun 8: 1934578X1300800235.
- SILVEIRA GL, GABRIELA M, LIMA F, REIS GB, PALMIERI MJ & ANDRADE-VIERIA LF. 2017. Toxic effects of environmental pollutants: Comparative investigation using Allium cepa L. and Lactuca sativa L. Chemosphere 178: 359-367.
- SMITH AE. 1989. The potential allelopathic characteristics of bitter sneezeweed (Helenium amarum). Weed Sci 37: 665-669.
- SOLTYS D, RUDZIŃSKA-LANGWALD A, KUREK W, GNIAZDOWSKA A, SLIWINSKA E & BOGATEK R. 2011. Cyanamide mode of action during inhibition of onion (Allium cepa L.) root growth involves disturbances in cell division and cytoskeleton formation. Planta 234: 609-621.
- SOUZA FILHO APS, SANTOS RA, SANTOS LS, GUILHON GMP, SANTOS AS, ARRUDA MSP, MULLER AH & ARRUDA AC. 2006. Allelophatic potential of Myrcia guianensis. Planta Daninha 24: 649-656.
- SOUZA TS, FERREIRA MFS, MENINI L, SOUZA JRCL, PARREIRA LA, CECON PR & FERREIRA A. 2017. Essential oil of Psidium guajava: Influence of genotypes and environment. Sci Hortic 216: 38-44.
- TUKEY HB. 1969. Implications of allelopathy in agricultural plant science. Bot Rev 35: 1-16.
- VASCONCELOS LC, SANTOS ES, BERNARDES CO, FERREIRA MFS, FERREIRA A, TULER AC, CARVALHO JAM, PINHEIRO PF & PRAÇA-FONTES MM. 2019. Phytochemical analysis and effect of the essential oil of Psidium L. species on the initial development and mitotic activity of plants. Environ Sci Pollut Res 26: 26216-26228.
- VERDEGUER M, CASTAÑEDA LG, TORRES-PAGAN N, LLORENS-MOLINA J & CARRUBBA A. 2020. Control of Erigeron bonariensis with Thymbra capitata, Mentha piperita, Eucalyptus camaldulensis, and Santolina chamaecyparissus essential oils. Molecules 25: 562.
- WEIR TL, PARK SW & VIVANCO JM. 2004. Biochemical and physiological mechanisms mediated by allelochemicals. Curr Opin Plant Biol 7: 472-479.
- ZAPPI DC ET AL. 2015. Growing knowledge: An overview of Seed Plant diversity in Brazil. Rodriguésia 66: 1085-1113.
- ZOGHBI MDGB, ANDRADE EHA, SILVA MHL, CARREIRA LMM & MAIA JGS. 2003. Essential oils from three Myrcia species. Flavour Fragr J 18: 421-424.
Publication Dates
-
Publication in this collection
18 July 2022 -
Date of issue
2022
History
-
Received
6 June 2021 -
Accepted
4 Nov 2021