
Resumos
Este trabalho revisa as alterações cerebrais de serotonina quando da oferta de nutrientes (carboidratos, proteínas e aminoácidos) durante atividade física. Utilizando a estratégia nutricional, o foco é o aminoácido precursor da serotonina cerebral: o triptofano; sendo um aminoácido essencial, é possível sua modulação via dieta. Uma abordagem emergente e polêmica está relacionada à fadiga durante atividade de curta e longa duração e sua relação com a função serotoninérgica cerebral. Os mecanismos propostos para o desenvolvimento de fadiga precoce durante o exercício se apresentam amplamente inexplorados. Assim serão discutidos os prováveis mecanismos envolvidos na "hipótese da fadiga central" e a oferta de carboidratos e aminoácidos como estratégia para retardar este fato durante atividade física e alcançar melhora no rendimento esportivo.
Proteínas; Exercício; Suplementação; Aminoácidos de cadeia ramificada; Triptofano; Serotonina
There are two major aspects focused in the present work. The first one concerns the tryptophan: a cerebral serotonin precursor aminoacid that alters serotonin and its relation to the offer of nutrients (carbohydrates, proteins and aminoacids). As an essential aminoacid it can be modulated through diet, having its physiological and behavioral effects changed, as it can be learned in the present study. The second aspect is related to the precocious fatigue during sports and activities of short and long duration and its relation with the serotoninergic cerebral function. What causes this precocious fatigue and how it is developed is yet largely unknown. The present work reviews the mechanism involved in the "central fatigue hypothesis" and the offer of carbohydrates and aminoacids as a strategy to retard this effect during physical activity.
Proteins; Exercise; Supplementation; Branched-chain aminoacids; Tryptophan; Serotonin
REVISÃO
Implicações do sistema serotoninérgico no exercício físico
Serotoninergic system and its implications on physical exercise
Luciana Rossi; Julio Tirapegui
Departamento de Alimentos e Nutrição Experimental, Faculdade de Ciências Farmacêuticas, Universidade de São Paulo, São Paulo, SP
Endereço para correspondência Endereço para correspondência Julio Tirapegui Av. Prof. Lineu Prestes 580, bloco 14 Conjunto das Químicas Cidade Universitária, USP 05508-900 São Paulo, SP
RESUMO
Este trabalho revisa as alterações cerebrais de serotonina quando da oferta de nutrientes (carboidratos, proteínas e aminoácidos) durante atividade física. Utilizando a estratégia nutricional, o foco é o aminoácido precursor da serotonina cerebral: o triptofano; sendo um aminoácido essencial, é possível sua modulação via dieta. Uma abordagem emergente e polêmica está relacionada à fadiga durante atividade de curta e longa duração e sua relação com a função serotoninérgica cerebral. Os mecanismos propostos para o desenvolvimento de fadiga precoce durante o exercício se apresentam amplamente inexplorados. Assim serão discutidos os prováveis mecanismos envolvidos na "hipótese da fadiga central" e a oferta de carboidratos e aminoácidos como estratégia para retardar este fato durante atividade física e alcançar melhora no rendimento esportivo.
Descritores: Proteínas; Exercício; Suplementação; Aminoácidos de cadeia ramificada; Triptofano; Serotonina
ABSTRACT
There are two major aspects focused in the present work. The first one concerns the tryptophan: a cerebral serotonin precursor aminoacid that alters serotonin and its relation to the offer of nutrients (carbohydrates, proteins and aminoacids). As an essential aminoacid it can be modulated through diet, having its physiological and behavioral effects changed, as it can be learned in the present study. The second aspect is related to the precocious fatigue during sports and activities of short and long duration and its relation with the serotoninergic cerebral function. What causes this precocious fatigue and how it is developed is yet largely unknown. The present work reviews the mechanism involved in the "central fatigue hypothesis" and the offer of carbohydrates and aminoacids as a strategy to retard this effect during physical activity.
Keywords: Proteins; Exercise; Supplementation; Branched-chain aminoacids; Tryptophan; Serotonin
A SETOTONINA OU 5-HIDROXITRIPTAMINA (5-HT) compõe o grupo das aminas biogênicas (neurotransmissores), que incluem também as catecolaminas (adrenalina, noradrenalina e dopamina). Estes compostos possuem grupos funcionais amina e regulam importantes vias do metabolismo dos mamíferos. São sintetizados na sua grande maioria a partir da decarboxilação de aminoácidos aromáticos (fenilalanina, tirosina e triptofano). A tirosina pode ser obtida da fenilalanina e é o aminoácido precursor das catecolaminas; já a serotonina é obtida do aminoácido essencial triptofano (1).
Os níveis de serotonina cerebrais estão relacionados a alterações de comportamento e humor, ansiedade, agressividade, depressão, sono, fadiga, supressão de apetite, etc. Os mecanismos bioquímicos precisos pelos quais os neurônios serotoninérgicos controlam estas funções ainda não estão totalmente esclarecidos (2). Existem, tanto em humanos como em animais (ratos e macacos), evidências de que a síntese de serotonina cerebral possa ser modulada dietéticamente através da oferta de macronutrientes, destacando-se principalmente a ingestão de carboidratos, proteínas e aminoácidos isoladamente (3-6).
Este trabalho revisa os mecanismos, durante exercício de curta e longa duração, relacionados à modulação da serotonina cerebral através dos quais uma manipulação nutricional pode afetar o rendimento, pela redução dos sintomas de fadiga.
Triptofano e Serotonina
O triptofano é um aminoácido aromático essencial, tanto para homens como animais. Sua essencialidade não se restringe apenas à sua contribuição no crescimento normal e síntese protéica, mas também na regulação de importantes mecanismos fisiológicos. Entre suas diversas funções temos: precursor do neurotransmissor serotonina (5-hidroxitriptamina: 5-HT), e sua influência no sono, comportamento, fadiga, ingestão alimentar entre outras. Além disto, o triptofano é precursor da vitamina B3 (niacina) e é um dos aminoácidos que estimula a secreção de insulina e hormônio do crescimento (7).
O triptofano, não podendo ser produzido pelo organismo, é obtido da degradação das proteínas cerebrais ou da circulação plasmática (pool aminoacídico), sendo este fornecido através da ingestão dietética normal ou degradação protéica corporal (8). No plasma, o triptofano pode circular livre (10%), ou, principalmente, ligado a uma proteína de transporte: a albumina (90%). Na barreira hematoencefálica, o triptofano livre (TRPL) compete com outros cinco aminoácidos (aminoácidos neutros: AN) para seu transporte e conseqüente síntese de serotonina cerebral (9). Os cinco aminoácidos competidores pela passagem através da barreira hematoencefálica são: leucina, isoleucina e valina (aminoácidos de cadeia ramificada: ACR), além da tirosina e fenilalanina (aminoácidos aromáticos) (10) (figura 1a). Entre os cinco aminoácidos competidores, o que possui a menor concentração plasmática é o triptofano (50µM). É estimado que a razão entre TRP:AN seja de 1:100 (11).
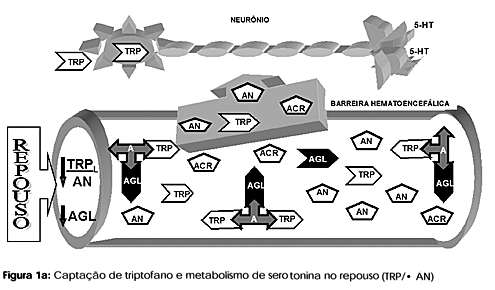
Uma abordagem utilizada na investigação da captação cerebral de triptofano é a proporção plasmática entre TRP e a soma dos aminoácidos neutros (AN): (TRP/< AN). Os mecanismos que aumentam a concentração de TRP, sobre a soma de seus competidores, são prováveis de aumentar a síntese de serotonina cerebral (12). Outro aspecto da competição pela barreira hematoencefálica é o aumento da concentração da parcela livre do triptofano. A albumina também transporta ácidos graxos livres, quanto maior sua concentração plasmática mais triptofano livre é deslocado, melhorando sua captação cerebral (13). É aceito que a taxa de síntese de serotonina é dependente da disponibilidade do substrato (triptofano livre) no plasma, uma vez que a enzima limitante do processo no sistema nervoso central (triptofano hidroxilase) (14) está cerca de 50% saturada (8). Abordagens indiretas, para a medida da concentração de serotonina cerebral, são utilizadas principalmente em humanos, devido à impossibilidade de determinações diretas da mesma.
Modulação Dietética da Serotonina e Fadiga
A síntese de serotonina cerebral pode ser modulada por três fatores: quantidade de triptofano total no plasma (proporção entre a parcela livre e a ligada à albumina); transporte de triptofano livre pela barreira hematoencefálica contra seus competidores (TRP/<AN) e a atividade da enzima triptofano hidroxilase. Os dois primeiros mecanismos possuem possibilidade de manipulação dietética.
Cunliffe e cols., em diversos experimentos (3,5), propuseram testes para determinar fadiga subjetiva mediada pela alteração da concentração da serotonina cerebral: a denominada fadiga central. Em um primeiro experimento para avaliar a relação entre alteração da composição de refeições e da proporção TRP/< AN em humanos, foram feitos diversos testes para acessar a fadiga central e periférica. A fadiga periférica seria aquela relacionada ao cansaço ou incapacidade muscular para realização da tarefa, e a fadiga central aquela relacionada ao sistema nervoso central (SNC). Os indivíduos que consumiram dietas hiperglicídicas apresentaram mais sintomas subjetivos de fadiga e menor tempo de reação, ocasionando uma piora no estado de alerta e atenção para este grupo. Em um segundo estudo, a oferta direta de triptofano (30mg/kg) para indivíduos saudáveis também apresentou resultados semelhantes aos anteriores, porém com redução de sensação de desconforto em teste ergométrico. O efeito opióide e analgésico da administração de triptofano seria a razão do maior rendimento. Há uma aplicação direta destes estudos sobre fadiga central ou cerebral na atividade física como veremos a seguir.
Serotonina e Atividade Física
Tanto a fadiga central como a periférica podem ocorrer em indivíduos que se exercitam em alta intensidade (15). As modificações nutricionais envolvendo principalmente carboidratos, proteínas e aminoácidos, podem, junto com outros fatores, alterar a neuroquímica cerebral (16). Assim, a fadiga central pode ser tanto relacionada à nutrição como a certas desordens como depressão, síndrome pré-menstrual, insônia (17), entre outras. Com relação à sua associação com atividade física, a fadiga central possui mecanismos ainda mais promissores.
Fadiga Periférica x Central
Para efeito de discussão, definiremos fadiga como um conjunto de manifestações produzidas por trabalho ou exercício prolongado, que tem como conseqüência redução ou prejuízo na capacidade funcional de manter ou continuar o rendimento esperado (18). A fadiga é um sinalizador de que a intensidade de treinamento previamente tolerada deve ser reduzida. No nível bioquímico podemos considerar a fadiga periférica, como aquela onde a perda da força e potência que ocorre é independente da ação neuronal; e a central, que é uma redução progressiva do direcionamento voluntário para os neurônios motores durante o exercício. Na periférica, ocorreriam nos músculos em atividade diversas alterações, tais como: depleção de fosfocreatina; acúmulo de prótons e fosfato. Já na central, os mecanismos relacionados à fadiga seriam a hipoglicemia e alteração plasmática na concentração aminoacídica, principalmente em relação à razão TRP/< AN (19). A vantagem da abordagem bioquímica da fadiga central é que há a possibilidade de, através do binômio duração do exercício-e-nutrição, influenciar o metabolismo serotoninérgico central pela manipulação do seu precursor, o triptofano.
Atividade Física de Alta Intensidade e Curta Duração
O uso de triptofano era muito comum na década de 80, principalmente associado ao tratamento de insônia, depressão e síndrome pré-menstrual, entre outros. Entre atletas, seu uso era direcionado na tentativa de aumentar a secreção do hormônio do crescimento (GH) (20). Ainda diversos estudos apontavam que a serotonina (5-HT) poderia estar relacionada à determinação do limiar de dor e respostas comportamentais ao estímulo doloroso (3,21). Baseados nesta última visão, Segura & Ventura (22) conduziram um experimento com a suplementação de 1,2g de triptofano em 20 atletas durante corrida de esteira a 80% do VO2máx. Os indivíduos suplementados foram capazes de, em um segundo teste, aumentar em 49,4% seu tempo total de exercício em relação ao grupo placebo. Os autores atribuíram este aumento acentuado no rendimento aos efeitos do triptofano e da serotonina no sistema opióide cerebral, fazendo com que houvesse uma menor percepção do esforço. Nos anos subseqüentes, diversas críticas foram feitas em relação a este estudo, como o fato de dois atletas terem um aumento de rendimento de 160 e 260% (11,23), e também ao fato dos autores não discutirem os potenciais efeitos tóxicos do triptofano (24). No início da década de 90, o Food and Drug Administration (FDA) ordenou a retirada do mercado de suplementos e fórmulas com altas doses de triptofano. O banimento do aminoácido se deu devido ao aumento no número de casos da síndrome da Eosinofilia-Mialgia (SEM) como conseqüência do consumo de suplementos com triptofano (12). A SEM é caracterizada pela eosinofilia e severas mialgias generalizadas. Houve, segundo registro da FDA, 21 casos de mortes confirmadas. Contaminantes presentes nos suplementos de triptofano, formados durante o seu processo de purificação, foram identificados como a causa da doença e mortes (20).
Atividade Física de Longa Duração
Desde que o grupo de Eric Newsholme, em 1987, propôs a chamada "hipótese da fadiga central" durante atividades de longa duração, o triptofano como precursor da serotonina tem recebido especial atenção por pesquisadores da área. Segundo esta hipótese, o aumento na atividade serotoninérgica cerebral teria conseqüência no desenvolvimento de fadiga precoce durante a realização do exercício de longa duração em atletas.
Mecanismo da Fadiga Central Durante o Exercício
Durante a atividade de longa duração, dois eventos contribuem para o maior influxo de triptofano pela barreira hematoencefálica. Primeiro, os ACR seriam captados pelo músculo esquelético para serem oxidados para fins energéticos, concomitantemente à queda da glicemia, efeito comum durante tais eventos. Com a redução da competição na barreira hematoencefálica, aumentaria a captação de triptofano e seu influxo cerebral (13). Em segundo lugar, com o prosseguimento do exercício, o organismo também começa a mobilizar para fins energéticos ácidos graxos (AGL) do tecido adiposo. Os AGL competem pelo transporte com o triptofano, sendo que esta competição pelos locais de ligação da albumina faz com que ocorra maior disponibilidade de triptofano livre (25). A albumina humana possui apenas um local de ligação para o triptofano, já a bovina possuí dois (26). O sinergismo entre deslocamento da parcela livre, mediada por AGL, e menor competição, mediada pela redução plasmática dos ACR, tem como conseqüência maior produção de serotonina cerebral e desencadeamento de fadiga central precoce (figuras 1a e 1b).
Modulação Dietética da Serotonina Cerebral
Com respeito à fadiga central, algumas estratégias nutricionais têm sido experimentalmente empregadas nesta área. Elas basicamente envolvem a suplementação com ACR e/ou carboidratos durante o exercício (27,28). Na suplementação com carboidratos, o foco é atenuar o aumento na concentração plasmática de AGL induzido pelo exercício e elevação dos hormônios contra-regulatórios (catecolaminas) (29). Os AGL possuem maior afinidade pela albumina do que o triptofano, assim o seu aumento disponibilizaria mais triptofano livre para a passagem pela barreira hematoencefálica. Durante a atividade física, há diminuição na concentração de insulina, para não haver aumento na sua secreção pancreática, soluções com até 12% de carboidrato são oferecidas (12,30). Uma concentração maior teria como efeito hipoglicemia e interrupção do exercício. A maioria dos estudos na área utilizam soluções de 6% de glicose (28,31).
Na suplementação com ACR, o objetivo é manter a concentração aminoacídica dos competidores do triptofano. Estudos realizados tanto em ratos como em seres humanos, têm demonstrado boas evidências para reduzir os sintomas de fadiga central (27,31). Em nosso laboratório, ratos Wistar treinados por 6 semanas em natação e suplementados na ração com 50% a mais de ACR foram submetidos a teste de exaustão. O grupo suplementado teve tempo médio até a exaustão de 345min, que foi 29% maior em relação ao do controle. Este mesmo grupo apresentou, no momento da exaustão, concentração de serotonina cerebral 33,3% menor, confirmando o efeito positivo da suplementação no rendimento e retardo da fadiga central (32). Estes resultados estão de acordo com Calders e cols. (33), que utilizaram 30mg de ACR e obtiveram melhora de 30,8% no rendimento dos ratos suplementados em teste exaustivo na esteira. Embora os experimentos utilizando animais indiquem o grande potencial da "hipótese da fadiga central", estudos laboratorialmente controlados com humanos não têm confirmado a hipótese (11,28) (tabela 1). Várias questões metodológicas são apontadas para resultados negativos a respeito, mas, recentemente, um estudo conduzido por Welsh e cols. (35) levantou uma questão importante a respeito desta problemática. Os pesquisadores infundiram soluções contendo ACR, carboidratos ou água no estômago de ratos durante teste em esteira. Não foi observada alteração de serotonina cerebral nos animais que foram sacrificados após 60 e 90min de exercício. No experimento, os animais não foram submetidos à fadiga. Porém, após 120 minutos, o grupo que recebeu tanto carboidrato como ACR apresentou menores concentrações cerebrais de serotonina. Estes resultados mostram a importância da duração do exercício para que ocorram alterações significativas do metabolismo serotoninérgico.
CONSIDERAÇÕES FINAIS
O triptofano é um aminoácido essencial tanto para humanos como animais, porém sua importância não se restringe apenas à contribuição no crescimento e síntese protéica. Como precursor da serotonina cerebral, o triptofano exerce papel fundamental em diversos mecanismos fisiológicos e comportamentais como sono, depressão, ingestão alimentar, fadiga, entre outros. Em relação à atividade física, há envolvimento do triptofano tanto em exercícios de longa como de curta duração. Nos exercícios de longa duração, foi proposto o mecanismo da "hipótese da fadiga central" que se encontra até agora inexplorado. As dificuldades no estudo da determinação da serotonina residem na obtenção da sua concentração cerebral e de seus metabólitos. Ainda assim, há evidências indiretas de seu papel dentro do desenvolvimento de fadiga associada a esportes de longa duração.
AGRADECIMENTOS
Os autores agradecem à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Capes), CNPq, FAPESP e à Prof. Dra. Inar Alves de Castro pela revisão crítica do manuscrito.
Recebido em 25/04/03
Revisado em 05/09/03
Aceito em 04/11/03
- 1. Meeusen R, de Meirleir K. Exercise and brain neurotransmission. Sports Med1995;20:160-88.
- 2. Bell C, Abrams J, Nutt D. Tryptophan depletion and its implications for psychiatry. Br J Psyc 2001;178:399-405.
- 3. Cunliffe A, Obeid OA, Powell-Tuck J. A placebo controlled investigation of the effects of tryptophan or placebo on subjective and objective measures of fatigue. Eur J Clin Nutr 1998;52:425-30.
- 4. Lyons PM, Truswell AS. Serotonin precursor influenced by type of carbohydrate meal in healthy adults. Am J Clin Nutr 1988;47:433-9.
- 5. Cunliffe A, Obeid OA, Powell-Tuck J. Post-prandial changes in measures of fatigue: effect of a mixed or a pure carbohydrate or pure fat meal. Eur J Clin Nutr 1997;51:831-8.
- 6. Blundell JE. Serotonin and the biology of feeding. Am J Clin Nutr 1992;55:S155-9.
- 7. Cuq JA, Friedman M. Effect of heat on tryptophan in food: chemistry, toxicology and nutritional consequences. In: Friedman, M. Absorption and utilization of amino acids III 1st ed. Florida:CRC Press; 1989 p.103-28.
- 8. Wurtman RJ, Fernstrom JF. Control of brain monoamine synthesis by diet and plasma amino acids. Am J Clin Nutr 1975;28:638-47.
- 9. Pardridge WM, Choi TB. Neutral amino acid transport at the human blood-brain barrier. Federation Proc 1986;45:2073-8.
- 10. Rossi L, Tirapegui J. Aspectos atuais sobre exercício físico, fadiga e nutriçăo. Rev Paul Educ Fís 1999;13:67-82.
- 11. Jakeman PM. Amino acid metabolism, branched-chain amino acid feeding and brain monoamine function. Proc Nutr Soc 1998;57:35-41.
- 12. Fernstrom JD. Dietary amino acids and brain function. J Am Diet Assoc 1994;94:71-7.
- 13. Davis JM. Carbohydrates, branched-chain amino acids, and endurance: the central fatigue hypothesis. Int J Sports Nutr 1995;5:S29-38.
- 14. Chaouloff F. Physiopharmacological interactions between stress hormones and central serotoninerg system. Brain Res Rev 1993;18:1-31.
- 15. Davis JM, Bailey SP. Possible mechanisms of central nervous system fatigue during exercise. Med Sci Sports Exer 1997;29:45-57.
- 16. Davis JM, Alderson NL, Welsh RS. Serotonin and central nervous system fatigue: nutritional considerations. Am J Clin Nutr 2000;72:573S-8.
- 17. Lieberman HR, Corkin S, Spring BJ, Wurtman RJ, Growdon JH. The effects of dietary neurotransmitter precursors on human behavior. Am J Clin Nutr 1985;42:366-70.
- 18. Rossi L, Tirapegui J. Aminoácidos: bases atuais para sua suplementaçăo na atividade física. Rev Bras Cięncias Farm 2000;36:37-51.
- 19. Hoomann W, Strüder HK. Exercise, physical activity, nutrition, and the brain. Nutr Rev 1996;54:S37-43.
- 20. Pasquale M. Essential aminoacids. In: Pasquale M, editor. Aminoacids and proteins for the athlete: the anabolic edge. 1st ed. New York:CRC Press; 1997 p.105-26.
- 21. Prasad C. Food, mood and health: a neurobiologic outlook. Braz J Med Biol Res 1998;31:1517-27.
- 22. Segura R, Ventura JL. Effect of L-tryptophan supplementation on exercise performance. Int J Sports Med 1988;9:301-5.
- 23. Stensrud T, Ingjer F, Holm H, Stromme SB. L-tryptophan supplementation does not improve running performance. Int J Sports Med 1992;13:481-6.
- 24. Chaouloff F. About the effect of L-tryptophan on exercise performance: lacunae e pitfalls. Int J Sports Med 1989;10:383.
- 25. Curzon G, Knott PJ. Effects on plasma and brain tryptophan in the rat of drugs and hormones that influence the concentration of unesterified fatty acid in the plasma. Br J Pharmac 1974;50:197-204.
- 26. Strüder HK, Weicker H. Physiology and pathophysiology of the serotonergic system and its implications on mental and physical performance. Part I. Int J Sports Med 2001;22:467-81.
- 27. Blomstrand E, Anderson S, Hassmen P, Ekblom B, Newsholme EA. Effect of branched-chain aminoacid and carbohydrate supplementation on the exercise-induced change in plasma and muscle concentration of amino acids in human subjects. Acta Physiol Scand 1995;153:87-96.
- 28. Van Hall G, Raaymakers JSH, Saris WHM, Wagenmakers AJM. Ingestion of branched-chain aminoacids and tryptophan during sustained exercise: failure to affect performance. J Appl Physiol 1995;486:789-94.
- 29. Davis JM, Bailey SP, Woods JA, Galiano FJ, Hamilton M, Bartoli WP. Effects of carbohydrate feedings on plasma free-tryptophan and branched-chain amino acids during prolonged cycling. Eur J Appl Physiol 1992;65:513-9.
- 30. Galiano FJ, Davis JM, Bailey SP, Woods JA, Hamilton M. Physiologic, endocrine and performance effects of adding branched-chain amino acids to a 6% carbohydrate-electrolyte beverage during prolonged cycling. Med Sci Sports Exerc 1991;23:14S.
- 31. Blomstrand E, Hassmen P, Newsholme EA. Effect of branched-chain aminoacid supplementation on mental performance. Acta Physiol Scand 1991;136:473-81.
- 32. Rossi L. Efeitos da suplementaçăo com aminoácidos de cadeia ramificada e sua relaçăo com a fadiga periférica e central. Săo Paulo, 2001 [Dissertaçăo de Mestrado Universidade da Săo Paulo Faculdade de Cięncias Farmacęuticas].
- 33. Calders P, Pannier JP, Matthys DM, Lacroix EM. Pre-exercise branched-chain aminoacid administration increases endurance performance in rats. Med Sci Sports Exerc 1997;29:1182-6.
- 34. Strüder HK, Weicker H. Physiology and pathophysiology of the serotonergic system and its implications on mental and physical performance. Part II. Int J Sports Med 2001;22:482-97.
- 35. Weslh RS, Waskovich M, Alderson NL, Davis JM. Carbohydrate and branched-chain aminoacid feedings suppress brain 5-HT during prolonged exercise. Med Sci Sports Exerc 1997;29:S19.
Datas de Publicação
-
Publicação nesta coleção
07 Jul 2004 -
Data do Fascículo
Abr 2004
Histórico
-
Aceito
04 Nov 2003 -
Revisado
05 Set 2003 -
Recebido
25 Abr 2003


