Resumos
O balanço energético é um dos mais importantes mecanismos de homeostase e de sobrevivência das espécies. O sistema endocanabinóide é um novo e importante componente entre estes mecanismos. Os seus receptores e agonistas endógenos se expressam no sistema nervoso central (SNC) e perifericamente, em vários sítios, estabelecendo uma rede de comunicação periferiaSNC. Um aspecto marcante é a sua expressão no tecido adiposo, onde regula a lipogênese e aumenta a expressão de genes influentes no metabolismo dos lipídeos e dos carboidratos. Estes aspectos são importantes para o controle do peso corporal e da Síndrome Metabólica (SM). O sistema é ativado sob demanda e desativado rapidamente, atuando autócrina e paracrinamente, e as evidências sugerem que mantém-se hiperativado em estados de obesidade. Um antagonista específico do seu principal receptor (CB1), o Rimonabant, tem se mostrado importante ferramenta no controle do peso em modelos animais de obesidade e de SM. Da mesma forma, grandes estudos em humanos confirmam sua eficácia no controle do peso e das variáveis metabólicas, sugerindo um papel importante deste medicamento para o controle do risco cardiovascular associado à SM.
Endocanabinóide; Síndrome metabólica; Obesidade; Rimonabant
Energetic balance is a fundamental homeostasis mechanism, which contributes to the species' survival. The endocannabinoid system is a new and important component among such mechanisms. Its receptors and endogenous agonists are expressed in central nervous system (CNS) and at various peripheral organs, establishing a CNSperiphery net communication. A relevant aspect is its expression in the adipose tissue, where it regulates lipogenesis and increases the expression of influent genes on lipids and carbohydrate metabolism. Interestingly, it seems to be upregulated in human and animal obesity, although it is activated on demand and rapidly deactivated. Its activation increases food intake and promotes weight gain, contributing to Metabolic Syndrome (MS). Rimonabant is a specific antagonist to the main endocannabinoid receptor (CB1). In animal models of obesity and MS, as well as in humans, Rimonabant has demonstrated to be a useful tool in controlling weight and metabolic aspects. Indeed, some new human trials suggest a possible role for this substance in controlling cardiovascular risk factors related to MS.
Endocannabinoid; Metabolic syndrome; Obesity; Rimonabant
ATUALIZAÇÃO
O sistema endocanabinóide: novo paradigma no tratamento da síndrome metabólica
The endocannabinoid system: a new paradigm in the metabolic syndrome treatment
Amélio F. de Godoy-Matos; Erika Paniago Guedes; Luciana Lopes de Souza; Cynthia Melissa Valério
Serviço de Metabologia e Nutrologia do Instituto Estadual de Diabetes e Endocrinologia (IEDE/RJ), Rio de Janeiro, RJ
Endereço para correspondência Endereço para correspondência: Amélio F. de Godoy-Matos Serviço de Metabologia e Nutrologia Instituto Estadual de Diabetes e Endocrinologia (IEDE/RJ) Rua Moncorvo Filho 90 20211-340 Rio de Janeiro, RJ Fax: (21) 2579-0291 E-mail: godoymatos@openlink.com.br
RESUMO
O balanço energético é um dos mais importantes mecanismos de homeostase e de sobrevivência das espécies. O sistema endocanabinóide é um novo e importante componente entre estes mecanismos. Os seus receptores e agonistas endógenos se expressam no sistema nervoso central (SNC) e perifericamente, em vários sítios, estabelecendo uma rede de comunicação periferiaSNC. Um aspecto marcante é a sua expressão no tecido adiposo, onde regula a lipogênese e aumenta a expressão de genes influentes no metabolismo dos lipídeos e dos carboidratos. Estes aspectos são importantes para o controle do peso corporal e da Síndrome Metabólica (SM). O sistema é ativado sob demanda e desativado rapidamente, atuando autócrina e paracrinamente, e as evidências sugerem que mantém-se hiperativado em estados de obesidade. Um antagonista específico do seu principal receptor (CB1), o Rimonabant, tem se mostrado importante ferramenta no controle do peso em modelos animais de obesidade e de SM. Da mesma forma, grandes estudos em humanos confirmam sua eficácia no controle do peso e das variáveis metabólicas, sugerindo um papel importante deste medicamento para o controle do risco cardiovascular associado à SM.
Descritores: Endocanabinóide; Síndrome metabólica; Obesidade; Rimonabant
ABSTRACT
Energetic balance is a fundamental homeostasis mechanism, which contributes to the species' survival. The endocannabinoid system is a new and important component among such mechanisms. Its receptors and endogenous agonists are expressed in central nervous system (CNS) and at various peripheral organs, establishing a CNSperiphery net communication. A relevant aspect is its expression in the adipose tissue, where it regulates lipogenesis and increases the expression of influent genes on lipids and carbohydrate metabolism. Interestingly, it seems to be upregulated in human and animal obesity, although it is activated on demand and rapidly deactivated. Its activation increases food intake and promotes weight gain, contributing to Metabolic Syndrome (MS). Rimonabant is a specific antagonist to the main endocannabinoid receptor (CB1). In animal models of obesity and MS, as well as in humans, Rimonabant has demonstrated to be a useful tool in controlling weight and metabolic aspects. Indeed, some new human trials suggest a possible role for this substance in controlling cardiovascular risk factors related to MS.
Keywords: Endocannabinoid; Metabolic syndrome; Obesity; Rimonabant
É FATO QUE, NO CONTEXTO do desenvolvimento econômico mundial, o balanço de energia vem claramente pendendo para o excesso de ingestão calórica e para a diminuição do gasto de energia (1). Este balanço energético positivo tem resultado numa explosão epidêmica da obesidade e suas patologias associadas, como a Síndrome Metabólica (SM) (2,3). É compreensível, portanto, que um grande interesse científico tenha se desenvolvido visando à compreensão dos vários mecanismos endócrinos e neuroendócrinos de regulação do balanço energético.
A Cannabis sativa tem sido cultivada pelo homem há mais de 5.000 anos, seja para a obtenção de fibras para manufatura têxtil, seja para fins medicinais ou recreacionais (4). A maconha e outros derivados psicotrópicos da Cannabis sativa representam a droga ilegal mais consumida no mundo ocidental. Os relatos sobre o efeito estimulador do apetite da Cannabis sativa, sobretudo para alimentos doces e palatáveis, datam de 300 d.C. Nos anos 80, a capacidade do composto ativo da Cannabis e de seus análogos sintéticos de estimular o apetite e reduzir náuseas e vômitos estimulou o seu uso em pacientes com câncer (5,6). Apenas recentemente, porém, foram clonados os alvos celulares das substâncias canabinóides, que foram denominados receptores CB (5). Concomitantemente, devido ao seu papel no controle da ingestão alimentar e balanço energético, vem crescendo a atenção no potencial terapêutico das substâncias que interferem com o sistema endocanabinóide endógeno.
A longa história da Cannabis teve sua evolução estimulada e conduzida para a medicina após a descoberta e caracterização química do seu principal princípio ativo, o D9-Tetrahidrocanabinol (THC) (7). Seus principais derivados, utilizados na prática médica, são o Drabinol e a Nabilona. Em seqüência, em 1988, um sítio de ligação para o THC foi identificado no cérebro de ratos e, em 1990, foi clonado o primeiro receptor canabinóide (8). Após a identificação de um segundo receptor, ficou definida a nomenclatura de CB1 para o primeiro receptor e CB2 para o segundo. O CB1 é o mais abundante receptor GPCR (receptores de membrana-ligados-a-proteína G) no cérebro, enquanto o CB2 está presente nas células do sistema imunológico (5,6).
No início da década de 90, foram descobertos dois agonistas endógenos dos receptores canabinóides: a N-aracdonoil etanolamina (Anandamida) e a 2-aracdonoil glicerol (2-AG), sendo atualmente designadas como endocanabinóides (ECB) (5). As enzimas fosfolipase N-acilfosfatidiletalonamina-seletiva e lipase sn-1-diacilglicerol-seletiva são as enzimas que rapidamente hidrolisam a Anandamida e a 2-AG, respectivamente (6). Os receptores canabinóides, os endocanabinóides e as enzimas que catalisam sua síntese e degradação constituem o Sistema Endocanabinóide (SECB) (9).
O primeiro antagonista específico do receptor CB1 endocanabinóide foi descoberto em 1994, sendo denominado SR141716 ou Rimonabant (10). Esta substância vem sendo estudada como modulador do apetite e como agente para o controle do tabagismo e, principalmente, para o controle dos fatores de risco ligados à obesidade visceral (discussão adiante) (11). O antagonista específico do receptor CB2, SR144528, futuramente poderá ser usado em pesquisas com a finalidade de modular a resposta imune (10,12).
O Sistema Endocanabinóide
Os receptores canabinóides pertencem à superfamília dos receptores de membrana-ligados-a-proteína G (GPCR; G-Protein-Coupled-Receptor) (5,13). A ativação desses receptores, tipicamente, inibe a adenilato-ciclase com conseqüente fechamento dos canais de cálcio, abertura dos canais de potássio e estimulação de proteínas quinases. O CB1 é o mais abundante receptor GPCR no cérebro, expresso predominantemente nos neurônios pré-sinápticos, mas também se encontra presente no sistema nervoso periférico (6). De fundamental importância para o conhecimento médico é entender que os agonistas endógenos e os receptores CB1 se expressam em vários outros órgãos da periferia (tabela 1). Atenção especial deve ser dada à sua presença no tecido adiposo. Já os receptores CB2 estão presentes nas células do sistema imunológico (5,6). Há evidências farmacológicas e fisiológicas sugerindo a existência de outros subtipos de receptores, ainda não clonados (5).
Os principais agonistas endógenos dos receptores canabinóides, a anandamida e a 2-AG, são derivados de ácidos graxos poliinsaturados de cadeia longa, principalmente do ácido aracdônico (6). Essas substâncias não estão confinadas no SNC, mas também agem como mediadores locais em muitos tecidos, sendo produzidos por demanda após alterações agudas ou crônicas da homeostase celular (6).
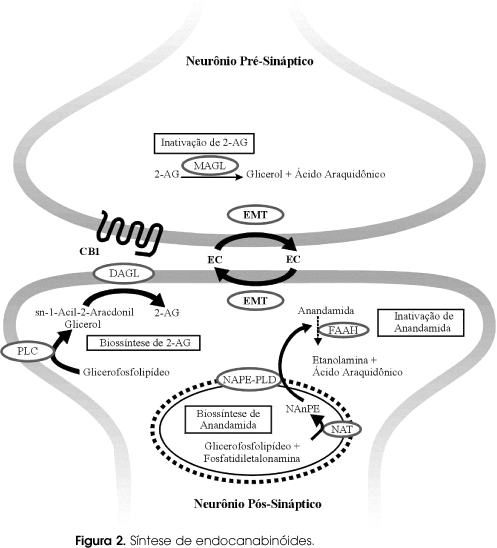
Os endocanabinóides agem como mensageiros cerebrais retrógrados (14,15). Diferentemente da seqüência usual, o estímulo começa no neurônio pós-sináptico e a excitação neuronal leva à despolarização e ao influxo de íons cálcio que estimulam várias fosfolipases, iniciando assim a síntese dos endocanabinóides (figura 1). Esses são liberados na fenda sináptica e se difundem livremente para estimular os receptores CB1 nos terminais pré-sinápticos neuronais (6). Os endocanabinóides agem como mediadores locais de forma parácrina e autócrina, sendo captados por células neuronais através de transportadores e metabolizados rapidamente. Assim, são rapidamente hidrolizados pelas enzimas fosfolipase N-acilfosfatidiletalonamina-seletiva (FAAH) e lipase sn-1-diacilglecerol seletiva (6). É importante ressaltar que os ECB não ficam armazenados nas vesículas lisossômicas, mas são prontamente sintetizados e liberados para as células, onde e quando forem necessários. Portanto, a disponibilidade dos ECB é regulada por captação-degradação (15). É interessante perceber, portanto, que a enzima de degradação da anandamida (FAAH) pode determinar maior atividade do sistema quando inativada. Com efeito, estudos muito recentes demonstraram uma associação entre um polimorfismo no gene da FAAH e obesidade em humanos (16). Ainda nesta linha de raciocínio, Engeli e cols. mostraram que os endocanabinóides anandamida e 2-AG estão aumentados no plasma de humanos obesos, e seus níveis estão inversamente relacionados com a atividade da FAAH (17). Isto corrobora dados em animais que apontam para uma hiperatividade do SECB em estados de obesidade. Jack e cols. demonstraram que a mesma mutação no gene da FAAH pode estar relacionada a um maior risco de uso de drogas ilícitas e alcoolismo, sugerindo também uma relação entre SECB e abuso e dependência de drogas (18).
Além de seus efeitos no balanço energético, o SECB tem um importante papel na regulação da secreção hormonal, através da sua ação primária no hipotálamo e direta na hipófise (5). A expressão dos receptores CB1 e a síntese de ECB nas células hipofisárias, além da habilidade dos ECB em inibir a secreção de prolactina e de GH e em aumentar a de ACTH, foram recentemente descritos (19).
A habilidade em modular o eixo hipotálamo-hipófise-adrenal e o envolvimento da resposta ao estresse são avaliados por estudos que demonstram que os ECB parecem produzir efeitos ansiolíticos dose-dependente (9). Além disso, são capazes de modular a resposta imune e inflamatória e várias funções fisiológicas, como a cardiovascular (alteração da freqüência cardíaca e vasodilatação), a respiratória (hiper ou hipoventilação e broncodilatação), a reprodutiva (inibição da secreção de testosterona e relaxamento uterino) e a ocular (diminui a pressão intra-ocular) (9).
O Sistema Endocanabinóide (SECB) e o Comportamento Alimentar
No homem, é reconhecido o efeito do uso recreacional da Cannabis, induzindo a procura por alimentos, notadamente alimentos palatáveis (este efeito é popularmente denominado de "larica") (19). Este fato e a experiência médica anterior, com o uso de canabinóides para alívio de anorexia e náuseas em pacientes com doenças consumptivas, sugeriam um papel deste sistema na modulação do apetite (19). Anatomicamente, esta relação parece se confirmar, ao se demonstrar que os endocanabinóides e o receptor CB1 estão presentes em altas concentrações em áreas hipotalâmicas que estão envolvidas no controle alimentar como a ventromedial, a dorsomedial, a lateral, os núcleos arqueados e os paraventriculares (10,20).
Uma interessante característica do sistema endocanabinóide é o fato de agir "sob demanda". Isto significa que é acionado apenas quando necessário e funciona para reparar ou modular a função de outros mediadores. Obviamente, pela abundância no SNC, os neurotransmissores são os principais candidatos à sua interação (5,6). Diversos sítios de produção de neurotransmissores relacionados ao comportamento alimentar, como CRH (Corticotropin Releasing Hormone), MCH (Melanin Concentrating Hormone), CART (Cocaine-Amphetamine Related Transcript) e a pré-pró-orexina estão localizados próximos aos receptores CB1 (20). Existem, ainda, evidências para uma relação funcional entre o SECB e o neuropeptídeo Y, a melanocortina, o GLP-1 e a grelina (20,21).
As experiências com animais de laboratório atestam a intrínseca relação entre o SECB e o controle alimentar (18-25). Por exemplo, a restrição alimentar aumenta os níveis hipotalâmicos dos ECB, que diminuem quando os animais são alimentados (22). De outra forma, a administração central e periférica de anandamida (AEA) aumenta a ingesta alimentar em roedores (20). Jamshidi e cols. injetaram anandamida no hipotálamo de ratos pré-saciados e observaram um grande e significativo aumento da ingesta alimentar após três horas do procedimento (23). Curiosamente, os níveis de anandamida estão aumentados em modelos clássicos de obesidade animal, como nos camundongos db/db, apesar de esses animais serem hiperfágicos. Já a leptina, que é secretada pelo adipócito e apresenta efeito na redução da ingestão alimentar, promoveu redução dos níveis hipotalâmicos de AEA E 2-AG, quando administrada agudamente a ratos normais ou ob/ob (24). Sugere-se, deste modo, que uma deficiência da sinalização da leptina proporcionaria o aumento dos níveis de EC e que, na obesidade, este sistema estaria hiperativo. Di Marzo e cols. demonstraram, assim, que o SECB e a leptina fazem parte do sistema homeostático que regula a ingestão alimentar e o peso corporal (24). Isto parece se confirmar em humanos. Monteleone e cols. demonstraram uma relação inversa entre a anandamida e a leptina em mulheres com peso normal. O mesmo aconteceu nas mulheres anoréticas, que exibiram baixos níveis de leptina e, provavelmente como conseqüência desta deficiência, níveis elevados de AEA (mas não de 2-AG) (25). Este fato poderia ser interpretado como uma tentativa de adaptação à restrição da ingesta causada pela drástica redução de leptina na anorexia nervosa, apesar de que fatores psicológicos parecem se sobressair aos biológicos, frustrando este mecanismo adaptativo (25).
Os receptores CB1 estão presentes no trato gastrointestinal, nas mesmas regiões onde se expressam peptídeos envolvidos no controle alimentar, sugerindo um possível papel do SECB na modulação da alimentação através de uma sinalização intestino-cérebro (21). Já foi demonstrado que o jejum aumenta os níveis de anandamida no intestino delgado, o que se relacionaria ao estímulo da ingesta alimentar (26). A grelina é um peptídeo sintetizado pela mucosa do fundo gástrico, que age como um potente orexígeno, sinalizando o início da alimentação (27). Cani e cols. demonstraram que a administração do antagonista do receptor CB1, o Rimonabant, inibiu a ingesta alimentar nos ratos em jejum, o que se associou a redução significativa dos níveis de ghrelina (18). Estes resultados sugerem um papel do SECB na regulação da secreção de peptídeos gastrointestinais orexígenos.
Uma forma de demonstrar o papel do SECB no controle do balanço energético é, naturalmente, a sua inativação (28). Isto pode ser feito inativando geneticamente o seu principal receptor (CB1) ou utilizando um antagonista CB1 específico (6). Assim, foi demonstrado que camundongos nocauteados para o gene do receptor CB1 consomem significativamente menos comida (6). Cota e cols. demonstraram também que os animais nocauteados (KO) para o gene do receptor CB1 ganhavam menos peso do que os animais primitivos (20). Interessantemente, ajustando a ingestão alimentar dos dois grupos, estes autores demonstraram que, ainda assim, os animais KO pesavam menos que os controles. Mais curioso, submetidos a estudo de composição corporal com Ressonância Nuclear Magnética, os animais KO apresentavam menor percentual de gordura corporal. Estes dados sugerem que a inativação do sistema diminui o peso corporal não só pelo controle da ingestão alimentar, mas, também perifericamente, diminuindo a lipogênese (20).
Um antagonista seletivo do receptor CB1, denominado Rimonabant, foi desenvolvido após um melhor entendimento do SECB (9). Estudos com o Rimonabant demonstram uma potente diminuição na procura por alimentos palatáveis (doces, por exemplo) em animais alimentados ad libitum ou uma diminuição da ingesta, de curta duração, em animais sob restrição alimentar (5,6). Num modelo de obesidade animal mais semelhante à obesidade humana, animais submetidos a uma dieta rica em gordura quando tratados cronicamente com Rimonabant, diminuíram transitoriamente a ingestão alimentar e perderam peso significativamente (29). Quando administrado para animais nocauteados para o receptor CB1, o medicamento não exerceu qualquer efeito, atestando que sua ação na regulação do apetite é, realmente, via o sistema endocanabinóide (29). Num estudo subseqüente, o mesmo grupo de autores demonstrou que animais KO para o receptor CB1 se mostraram resistentes em ganhar peso sob uma dieta hiperlipídica (30). Em confirmação aos experimentos de Cota e cols., discutido anteriormente, os animais nocauteados para o gene do receptor CB1 mostravam uma composição corporal de menor percentual de gordura (20,30).
Os mais importantes estudos para elucidar os mecanismos de controle do peso pelo SECB talvez venham de uma recente série de experimentos produzidos por Jbilo e cols. (31). Estes autores demonstraram, no mesmo modelo de obesidade induzida por dieta e tratados com Rimonabant ou placebo, que: 1) o tecido adiposo branco (WAT) e o tecido adiposo marrom (BAT) lombar eram 64% e 46% menor (p< 0,001) nos animais tratados com Rimonabant em comparação ao placebo, 2) a diferença na média do diâmetro do adipócito era 57% menor e a capacidade de estocagem de gordura estimada era 90% menor no grupo tratado com Rimonabant (estas diferenças não eram explicadas pela ingestão alimentar, já que apenas no início da experiência houve redução da ingesta), 3) através da análise de uma série de genes pela técnica do chip de DNA, ficou claro que o Rimonabant induziu genes em exata oposição aos efeitos da dieta hiperlipídica e em semelhança ao modelo do nocaute do receptor CB1 (31). A cascata de modulação destes genes favorecia a indução de genes que aumentam a lipólise (através da b-oxidação de ácidos graxos e do ciclo dos ácidos tricarboxílicos/TCA), aumentam o gasto energético (através dos ciclos fúteis ou dos substratos) e regulam a homeostase da glicose. Estas alterações contribuíram para uma redução do estado inflamatório induzido pela obesidade. Assim, os efeitos do SECB se dão através de mecanismos fenotípicos e genotípicos (31).
Estes e outros dados sugerem que o sistema ECB atua na ingestão alimentar de duas maneiras no SNC: 1) via sistema mesolímbico, reforçando e incentivando a procura por alimentos com capacidade de proporcionar mais prazer, e 2) via hipotálamo, agindo "sob demanda" para induzir o apetite, modulando ou regulando substâncias orexígenas ou anorexígenas quando de uma restrição alimentar (20). Além disso, as suas ações se estendem à periferia, através do eixo enteralSNC (21). Talvez mais importante, sua atividade no tecido adiposo, controlando a lipogênese, aparenta ser fundamental para o controle do peso e das alterações metabólicas conseqüentes, como será discutido adiante (31).
O fato de os receptores ECB não estarem confinados apenas ao SNC, mas disseminados em vários outros tecidos, torna mais consistente o conceito de estresse e de mecanismos para restabelecimento da homeostase (5,6,20). É interessante notar que, além do sedentarismo e do aumento da oferta de alimentos, o estresse crônico ajuda a explicar o papel do ambiente na gênese da obesidade. Neste caso, condições de estresse crônico levariam a uma hiperestimulação da síntese de ECB. Os estudos com a atividade da FAAH em humanos, discutidos anteriormente, sugerem também que a hiperatividade do SECB depende de uma menor degradação, resultando em hiperativação permanente dos receptores CB1 (16). Isto contribui para a manutenção da obesidade e conseqüente instalação da SM.
Evidências do papel do sistema ECB na SM em modelos animais
O tecido adiposo é fundamental para a fisiopatologia da SM devido à função armazenadora de energia, à função endócrina do adipócito, ao conceito da deposição ectópica de gordura e ao papel do sistema PPAR, entre outros (32-34). Só recentemente, porém, conhecimentos da relação do sistema ECB com o tecido adiposo surgiram e, naturalmente, as inferências sobre o seu papel na SM são uma questão lógica. O primeiro achado, na verdade, vem de longa data, quando ficou demonstrado que o canabinóide D9-Tetrahidrocanabinol (THC) era lipossolúvel (5,6,14). Mais importante, porém, foram os estudos de Cota e cols. (20) e de Bensaid e cols. (35), que demonstraram em camundongos e ratos, respectivamente, a presença de receptores CB1 no tecido adiposo. Ainda mais, foi demonstrado que o uso de um agonista do receptor CB1 (WIN-55,212) estimulava de maneira dose-dependente a lipase lipoprotéica e a lipogênese em cultura de células adiposas (20). Um outro achado que liga o sistema ECB com o tecido adiposo e a SM foi publicado por Bensaid e col. (35). Estes investigadores demonstraram que o Rimonabant induzia um aumento da expressão da proteína exclusiva do adipócito, a Acrp30 (adiponectina), no tecido adiposo de ratos Zucker obesos. O mesmo se observou em cultura de adipócitos, realçando um efeito direto no adipócito (35).
O fígado é um importante sítio de metabolismo da glicose e de lipídeos. Seu envolvimento na SM está bem estabelecido (19,36). De fato, a esteato-hepatite não alcoólica é um constituinte dos estados de obesidade e resistência à insulina (37,38). Como sobejamente conhecido, a resistência periférica à ação da insulina está relacionada de forma independente com a obesidade visceral (39,40). Da mesma forma, a resistência hepática à ação da insulina relaciona-se com o teor de gordura hepática, que por sua vez está relacionado com o depósito de gordura visceral (40,41). Ainda mais, a oferta de ácidos graxos para o fígado, avaliada pela sua concentração no território venoso esplâncnico, está diretamente relacionada com o teor de gordura visceral (42). Mediadores moleculares como o SREBP-1 (Steroid Regulator Element Binding Protein 1) estão envolvidos na fisiopatologia da esteatose e da injúria hepática nos estados de obesidade e resistência à insulina (43). Este é um fator de transcrição que atua na cascata da lipogênese hepática, via transcrição dos genes de enzimas-chave como a acetil-CoA-carboxilase-1 (ACC1) e a sintetase de ácidos graxos (FAS). Recentemente, foi demonstrado que o fígado também expressa receptores CB1, através dos quais há a indução do SREBP-1 e, conseqüentemente, indução da lipogênese hepática (44). Em animais com obesidade induzida por dieta, estes autores demonstraram um aumento do endocanabinóide anandamida, da expressão do receptor CB1 e de esteatose hepática (44). Mais ainda, mostraram que o tratamento destes animais com Rimonabant diminuiu a síntese hepática de ácidos graxos. Interessantemente, os animais KO para o receptor CB1 não só se mostraram resistentes ao ganho de peso, como também à esteatose (44).
Papel do antagonista do receptor CB1 na SM em humanos
Alguns estudos de fase III com o antagonista do receptor CB1, Rimonabant, já estão em andamento, e seus dados já estão sendo apresentados em congressos e simpósios mais recentes (tabela 2). O programa RIO (Rimonabant In Obesity) tem selecionado mais de 6.600 pacientes com sobrepeso/obesidade em todo o mundo para 4 estudos fase III: 1 estudo de 2 anos de duração (RIONorth America) e 3 estudos de 1 ano (RIOEurope, RIOLipids e RIODiabetes) (45-48). Dois destes estudos já foram publicados e serão discutidos adiante. O objetivo destes estudos foi a avaliação do papel de Rimonabant no manejo da obesidade, manutenção da perda de peso, prevenção da recuperação do peso após perda de peso inicial e melhora dos fatores de risco relacionados à obesidade, como diabetes e dislipidemia (47,48). Em geral, os resultados preliminares destes estudos apontam para um efeito significativo do Rimonabant em todos os parâmetros da SM (47,48).
No RIOEurope (Rimonabant in Obesity Europe), 1.507 pacientes com obesidade ou com excesso de peso associado a dislipidemia ou hipertensão foram randomizados, de forma duplo-cega, para receber placebo ou Rimonabant nas doses de 5 mg ou 20 mg ao dia (47). Em 1 ano, a perda de peso foi significativamente maior nos grupos Rimonabant quando comparados com o placebo (-3,4 ± 5,7 Kg para a dose de 5 mg e -6,6 ± 7,2 para a dose de 20 mg versus -1,8 ± 6,4 Kg para o grupo placebo). Na população que completou o tratamento, mais de 67% atingiu uma perda de 5% ou mais do peso inicial e 39% perdeu 10% ou mais, comparados com 30,5% e 12,4%, respectivamente, no grupo placebo. Ainda mais importante, a redução na circunferência abdominal foi -1,5 ± 7,3 cm para o placebo contra -8,5 ± 7,4 cm para o grupo 20 mg de Rimonabant (p< 0,001). Os grupos em uso de Rimonabant também obtiveram significativas melhoras nos níveis de triglicérides, HDL-colesterol e resistência à insulina (medida pelo HOMA), de forma que ao final do tratamento houve uma redução de 64,8% no número de pacientes que preenchiam o diagnóstico para SM no grupo de 20 mg do Rimonabant, contra 33,9% de redução no grupo placebo (p< 0,001). A glicemia de jejum diminuiu significativamente apenas no grupo de 20 mg, quando comparado ao placebo (34). Os dados do estudo RIONorth America, com dois anos de duração, mas ainda não publicados, são semelhantes ao relatados para o RIOEurope, sugerindo benefícios por um período mais prolongado.
À primeira vista, parece que os efeitos do Rimonabant nos parâmetros metabólicos estariam relacionados à perda de peso. Entretanto, nos estudos apresentados, modelos de regressão logística e/ou ANCOVA, usando a perda de peso como co-variável, mostraram que a melhora nos níveis de triglicérides e HDL-C eram parcialmente independentes da perda de peso (cerca de 40% para o HDL e 55% para os triglicérides) (47,48).
O estudo RIOLipids, recentemente publicado, envolveu 1.036 pacientes com excesso de peso (IMC entre 27 e 40 Kg/m2), hipertrigliceridemia e relação colesterol total/HDL-colesterol maior que 4,5 ou 5,0 para mulheres ou homens, respectivamente (48). Os pacientes não estavam previamente sob tratamento para a dislipidemia. Após randomização, os pacientes receberam orientação para uma dieta com 600 Kcal de déficit calórico mais placebo ou Rimonabant, 5 ou 20 mg/dia, por um período de 12 meses. Comparados com o placebo, os pacientes que receberam Rimonabant 20 mg perderam mais peso (média de 6,7 ± 0,5 kg; p< 0,001) e diminuíram mais a circunferência abdominal (média de 5,8 ± 0,5 cm; p< 0,001). Consistentes foram também os resultados no perfil lipídico: o HDL-C aumentou 10,0 ± 3,5% enquanto os triglicerídeos diminuíram em média 13,0 ± 3,5%. Interessante, os aumentos do HDL-C e da relação colesterol total/HDL-C demonstraram ser dose-dependente e houve uma redistribuição do perfil das partículas do LDL-C em favor de partículas maiores e menos densa ("flutuante" e menos aterogênica) no grupo Rimonabant 20 mg. Além disso, Rimonabant na dose de 20 mg conduziu a um aumento nos níveis de adiponectina de 57,7%. Foi estimado que cerca de 57% deste aumento foi independente da perda de peso. Isto parece confirmar, em humanos, os estudos que demonstram uma ação direta indutora da adiponectina no tecido adiposo.
O perfil de segurança do medicamento parece ser adequado (47,48). Até agora, cerca de 5.580 pacientes fizeram parte dos estudos RIO. Os dados acumulados mostram uma incidência de apenas 3,6% de efeitos colaterais acima do placebo. Os efeitos adversos mais comuns foram: náuseas (1,3%), diarréia (1,3%), tonturas (0,6%), depressão (1,4%) e ansiedade (0,7%) (47,48). No estudo RIOLipids, os efeitos adversos mais freqüentes, em ordem decrescente, foram: náuseas, tonteiras, gripe, ansiedade, diarréia e insônia (48), não diferindo muito em relação ao placebo. As taxas de depressão e ansiedade medidas pela "Escala Hospitalar de Depressão e Ansiedade" não foram diferentes entre os grupos e mantiveram-se em níveis baixos (em torno de 3,0 para depressão e 5,0 para ansiedade numa escala que vai de 0 a 21 pontos). Contudo, a taxa de descontinuação no estudo por eventos adversos foi maior no grupo Rimonabant 20 mg. Se exploradas mais detalhadamente, a depressão contribuiu com 2,9% e ansiedade 1,7% no grupo Rimonabant 20 mg, contra 0,6% no placebo (48).
Papel do antagonismo do receptor CB1 na dependência de nicotina
De acordo com a OMS, atualmente cerca de 47% dos homens e 12% das mulheres são tabagistas em todo o mundo (49). A relação entre tabagismo e mortalidade cardiovascular é bem definida, e o tabagismo permanece como a mais prevenível causa de morte cardiovascular (50,51).
A nicotina, presente no cigarro, é a substância responsável pelo desenvolvimento da dependência (52). O consumo de nicotina é motivado pela liberação de dopamina no nucleus accumbens. O uso crônico de nicotina resulta em hiperestimulação persistente do SECB no nucleus accumbens, com reforço da liberação de dopamina e abuso da nicotina (53).
O bloqueio seletivo do receptor CB1 tem demonstrado ser eficaz no tratamento da dependência à nicotina (45,49,50). O estudo STRATUS (STudies with Rimonabant And Tobacco USe), envolvendo 6.500 pacientes em todo o mundo, foi desenvolvido para avaliar o papel do antagonista seletivo de CB1 (Rimonabant) na interrupção do tabagismo, abstinência a longo prazo e prevenção de ganho de peso (após o paciente parar de fumar) (41). O projeto engloba 2 estudos fase III de 10 semanas de duração seguidas de 10 semanas de follow-up (STRATUSUS nos Estados Unidos e STRATUSEU na Europa) e um estudo de 1 ano de tratamento seguido por 1 ano de seguimento (STRATUSWW estudo mundial) (45,54,55).
Resultados preliminares do STRATUSUS demonstraram que a taxa de abandono do tabagismo foi significativamente maior com Rimonabant 20 mg ao dia quando comparado ao placebo, com também significativa redução do ganho de peso após a interrupção do tabagismo (45,54,55). O perfil de segurança do Rimonabant foi favorável neste estudo (45,54,55).
Considerando o papel do tabagismo como expressivo fator de risco cardiovascular, aliado ao ganho de peso associado à interrupção do tabagismo e ao possível desenvolvimento da SM, estes estudos orientam para o potencial terapêutico do bloqueio do SECB nesta situação.
CONCLUSÕES
Em resumo, o sistema ECB está relacionado com o controle energético e funciona induzindo a ingestão alimentar. Notadamente, é estimulado transitoriamente em situações de restrição alimentar e na presença de alimentos de alto valor de recompensa (palatável). Além disso, atua perifericamente, contribuindo para um "diálogo" entre a periferia e o SNC, e aumentando a lipogênese. O bloqueio deste sistema através de um antagonista seletivo do receptor CB1 (Rimonabant) diminui a ingestão alimentar, auxilia no controle do peso em animais e em humanos e contribui para uma melhora nos parâmetros metabólicos que compõem a SM (figura 2). Alguns desses efeitos parecem ser parcialmente independentes do controle do peso.
Recebido em 14/01/06
Aceito em 27/01/06
- 1. Webber J. Energy balance in obesity. Proc Nutr Soc 2003;62:539-43.
- 2. Flegal KM. Epidemiologic aspects of overweight and obesity in the United States. Physiol Behav 2005;86:599-602.
- 3. Berenson GS. Obesity a critical issue in preventive cardiology: the Bogalusa Heart Study. Prev Cardiol 2005;8:234-41.
- 4. Smith R, Fathi Z. Recent advances in the development and research of CB1 antagonists. Drugs 2005;8:55-63.
- 5. Di Marzo V, Bifulco M, Petrocellis L. The endocannabinoid system and its therapeutic exploitation. Nat Rev Drug Disc 2004;3:771-84.
- 6. Di Marzo V, Matias I. Endocannabinoid control of food intake and energy balance. Nat Neurosci 2005;8:585-9.
- 7. Gaoni Y, Mechoulan R. Isolation, structure, and partial synthesis of an active constituent of hashish. J Am Chem Soc 1964;86:1646-7.
- 8. Matsuda LA, Lolait SJ, Brownstein MJ, Young AC, Bonner TI. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990;346:561-4.
- 9. Cota D, Marsiano G, Lutz B, Vicennati V, Stalla GK, Pasquali R, et al. Endogenous cannabinoid system as a modulator of food intake. Int J Obes 2003;27:289-301.
- 10. Howlett AC, Breivogel CS, Childers SR, Deadwyler SA, Hampson RE, Porrino LJ. Cannabinoid physiology and pharmacology: 30 years of progress. Neuropharmacol 2004;47:345-58.
- 11. Cox SL. Rimonabant hydrochloride: an investigational agent for the management of cardiovascular risk factors. Drugs Today (Barc) 2005;41:499-508.
- 12. Cabral GA, Dove Pettit DA. Drugs and immunity: cannabinoids and their role in decreased resistance to infectious disease. J Neuroimmunol 2005;83:116-23.
- 13. Howlett AC. The cannabinoid receptors. Prostaglandins Other Lipid Mediat 2002;68-9:619-31.
- 14. De Petrocellis L, Cascio MG, Di Marzo V. The endocannabinoid system: a general view and latest additions. Br J Pharmacol 2004;141:765-74.
- 15. Christie MJ, Vaughan CW. Neurobiology: Cannabinoids act backwards. Nature 2001;410:527-30.
- 16. Sipe JC, Waalen J, Gerber A, Beutler E. Overweight and obesity associated with a missense polymorphism in fatty acid amide hydrolase (FAAH). Int J Obes 2005;29:755-9.
- 17. Engeli S, Böhnke J, Feldpausch M, Gorzelniak K, Janke J, Bátkai S, et al. Activation of the peripheral Endocannabinoid system in human obesity. Diabetes 2005;2838-43.
- 18. Sipe JC, Chiang K, Gerber AL, Beutlar E, Cravatt BF. A missense mutation in human fatty acid amide hydrolase associated with problem drug use. PNAS 2002;99:8394-9.
- 19. Godoy-Matos AF. O sistema canabinóide. In: Godoy-Matos AF, editor. Síndrome metabólica São Paulo: Atheneu, 2005
- 20. Cota D, Marsicano G, Tschöp M, Grübler Y, Flachskamm C, Schubert M, et al. The endogenous cannabinoid system affects energy balance via central orexigenic drive and peripheral lipogenesis. J Clin Invest 2003;112:423-31.
- 21. Cani PC, Montoya ML, Neyrinck AM, Delzenne NM, Lambert DM. Short communication Potential modulation of plasma ghrelin and glucagon-like peptide-1 by anorexigenic cannabinoid compounds, SR141716A (rimonabant) and oleoylethanolamide. Br J Nutr 2004;92:757-61.
- 22. Kirkham TC, Williams CM, Fezza F, Di Marzo V. Endocannabinoid levels in rat limbic forebrain and hypothalamus in relation to fasting, feeding and satiation: stimulation of eating by 2-arachidonoyl glycerol. Br J Pharmacol 2002;136:550-7.
- 23. Jamshidi N, Taylor DA. Anandamide administration into the ventromedial hypothalamus stimulates appetite in rats. Br J Pharmacol 2001;134:1151-4.
- 24. Di Marzo V, Goparaju SK, Wang L, Liu J, Batkai S, Jarai Z, et al. Leptin-regulated endocannabinoids are involved in maintaining food intake. Nature 2001;412:822-5.
- 25. Monteleone P, Matias I, Martiadis V, De Petrocellis L, Maj M, Di Marzo V. Blood levels of the endocannabinoid anandanida are increased in anorexia nervosa and binge-eating disorder, but not in Bulimia Nervosa. Neuropsycopharmacol 2005;30:1216-21.
- 26. Gomez R, Navarro M, Ferrer B, Trigo JM, Bilbao A, Del Arco I, et al. A peripheral mechanism for CB1 cannabinoid receptor-dependent modulation of feeding. J Neurosci 2002;22:9612-27.
- 27. Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H, Kangawa K. Ghrelin is a growth-hormone-releasing acylated from stomach. Nature 1999;402:656-60.
- 28. Giuffrida A, Beltramo M, Piomelli D. Mechanisms of endocannabinoid inactivation: biochemistry and pharmacology. J Pharmacol Exp Ther 2001;298:7-14.
- 29. Trillou C R, Arnone M, Delgorge C, Gonalons N, Keane P, Maffrand JP, et al. Anti-obesity effect of SR141716, a CB1 receptor antagonist, in diet-induced obese mice. Am J Physiol Regul Comp Physiol 2003;284:R345-53.
- 30. Ravinet Trillou C, Delgorge C, Menet C, Arnone M, Soubrie P. CB1 cannabinoid receptor knockout in mice leads to leanness, resistance to diet-induced obesity and enhanced leptin sensitivity. Int J Obes Relat Metab Disord 2004;28:640-8.
- 31. Jbilo O, Ravinet-Trillou C, Arnone N, Buisson I, Bribes E, et al. The CB1 receptor antagonist rimonabant reverses the diet-induced obesity phenotype through the regulation of lypolisis and energy balance. FASEB J 2005;19:1567-9.
- 32. Heilbronn L, Smith SR, Ravussin E. Failure of fats cell proliferation, mitochondrial function and fat oxidation results in ectopic fat storage, insulin resistance and type 2 diabetes mellitus. Inter J Obes 2004;28:S12-S21.
- 33. Unger RH. Mini review: Weapons of lean body mass destruction: the role of ectopic lipids in the metabolic syndrome. Endocrinology 2003;144:5159-65.
- 34. Ali AT, Crowther NJ. Body fat distribution and insulin resistance. SAMJ 2005;95:878-90.
- 35. Bensaid M, Gary-Bobo M, Esclangon A. The cannabinoid CB1 receptor antagonist SR141716 increases Acrp30 mRNA expression in adipose tissue of obese fa/fa rats and in cultured cells. Mol Pharmacol 2003;63:908-14.
- 36. Hamaguchi M, Kojima T, Takeda N, Nakagawa T, Taniguchi H, Fujii K, et al. Metabolic syndrome as a predictor of nonalcoholic fatty liver disease. Ann Intern Med 2005;143:722-8.
- 37. Medina J, García-Buey L, Fernandez-Salazar LI, Moreno-Otero R. Approach to the pathogenesis and treatment of non alcoholic steatohepatitis. Diabetes Care 2004;27:2057-66.
- 38. Diehl AM, Li ZP, Lin HZ, Iang SQ. Cytoquines and the pathogenesis of non-alcoholic steatohepatitis. Gut 2005;54:303-6.
- 39. Baneji MA, Faridi N, Atluri R, Chaiken RL, Lebovitz HE. Body composition, visceral fat, leptin, and insulin resistance in Asian Indian men. J Clin Endocrinol Metab 1999;84:137-44.
- 40. Wajchenberg BL. Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome. Endocr Rev 2000;21:697-738.
- 41. Thamer C, Machann J, Haap M, Stefan N, Heller E, Schnödt B, et al. Intrahepatic lipids are predicted by visceral adipose tissue mass in healthy subjects.Diabetes Care 2004;27:2726-9.
- 42. Nielsen S, Guo Z, Johnson CM, Hensrud DD, Jensen MD. Splanchnic lipolysis in human obesity. J Clin Invest 2004;113:1582-8.
- 43. Browning JD, Horton JD. Molecular mediators of hepatic steatosis and liver injury. J Clin Invest 2004;114:147-52.
- 44. Osei-Hyiaman D, DePetrillo M, Pacher P, Liu J, Radaeva S, Bátkai S, et al. Endocannabinoid action at hepatic CB1 receptors regulates fatty acid synthesis: role in diet-induced obesity. J Clin Invest 2005;115:1298-305.
- 45. Dale L, Anthenelli R, Despres J-P, Golay A, Sjostrom L. Effects of rimonabant in the reduction of major cardiovascular risk factors. Results from the STRATUSUS Trial (Smoking Cessation in Smokers Motivated to Quit) and the RIOLipids Trial (Weight Reducing and Metabolic Effects in Overweight/Obese Patients with Dyslipidemia). Late-Breaking Clinical Trials II. American College of Cardiology Scientific Session 2004, March 710, 2004, New Orleans, Louisiana.
- 46. Van Gaal L. RIOEurope: a randomized double-blind study of weight reducing effect and safety of rimonabant in obese patients with or without comorbidities. Hot Line I Prevention and medical treatment I. ESC Congress 2004, August 28 - September 1, 2004, Munich, Germany.
- 47. Van Gaal LF, Rissanen A, Scheen AJ, Ziegler O, Rössner S; the RIOEurope Study Group. Effects of the cannabinoid-1 receptor blocker rimonabant on weight reduction and cardiovascular risk factors in overweight patients: 1-year experience from the RIOEurope study. Lancet 2005;365:1389-97.
- 48. Després JP, Golay A, Sjöström L, the Rimonabant in ObesityLipids Study group. Effects of rimonabant on metabolic risk factors in overweight patients with dyslipidemia. NEJM 2005;353:2121-34.
- 49.Tobacco or health: A global status report Geneva: World Health Organization, 1997
- 50. Centers for Disease Control and Prevention. Annual smoking-attributable mortality, years of potential life lost, and economic costs United States, 1995-1999. Morb Mortal Wkly Rep 2002;51:300-3.
- 51. Godtfredsen NS, Holst C, Prescott E, Vestbo J, Osler M. Smoking reduction, smoking cessation, and mortality: a 16-year follow-up of 19,732 men and women from The Copenhagen Centre for Prospective Population Studies. Am J Epidemiol 2002;156:994-1001.
- 52. Balfour DJ. Neuroplasticity within the mesoaccumbens dopamine system and its role in tobacco dependence. Curr Drug Targets CNS Neurol Disord 2002;1:413-21.
- 53. De Vries TJ, De Vries W, Janssen MCW, Schoffelmeer ANM. Suppression of conditioned nicotine and sucrose seeking by the cannabinoid-1 receptor antagonist SR141716A. Behav Brain Res 2005;161:164-8.
- 54. Henningfield JE, Fant RV, Buchhalter AR, Stitzer ML. Pharmacotherapy for Nicotine Dependence. CA Cancer J Clin 2005;55:281-99.
- 55. Anthenelli R. Smoking cessation in smokers motivated to quit. Presented at: American College of Cardiology Scientific Sessions; March 710, 2004; New Orleans, LA.
Datas de Publicação
-
Publicação nesta coleção
23 Maio 2006 -
Data do Fascículo
Abr 2006
Histórico
-
Aceito
27 Jan 2006 -
Recebido
14 Jan 2006