Resumos
A microbiota intestinal, adquirida no período pós-natal, é composta por grande diversidade de bactérias que desempenham diferentes funções no hospedeiro humano, entre elas a absorção de nutrientes, proteção contra patógenos e modulação do sistema imune. O conteúdo bacteriano intestinal ainda não é totalmente conhecido, mas sabe-se que é influenciado por fatores internos e principalmente externos que modulam sua composição e função. Estudos indicam que a microbiota intestinal difere em indivíduos magros e obesos e ainda naqueles que mantêm hábitos alimentares diferentes. Há evidências de que as relações entre dieta, inflamação, resistência à insulina e risco cardiometabólico são em parte mediadas pela composição de bactérias intestinais. Conhecimentos sobre a microbiota poderão reverter em diferentes estratégias para manipular as populações bacterianas e promover saúde. Esta revisão aborda a relevância do conhecimento sobre o papel de fatores ou padrões alimentares na composição da microbiota, assim como mecanismos fisiopatológicos de doenças metabólicas crônicas e as potencialidades de prebióticos e probióticos sobre o perfil de risco cardiometabólico.
Microbiota intestinal; hábitos alimentares; inflamação; resistência à insulina; adiposidade corporal
The gut microbiota obtained after birth is composed of a large range of bacteria that play different roles in the human host, such as nutrient uptake, protection against pathogens and immune modulation. The intestinal bacterial content is not completely known, but it is influenced by internal, and mainly by external factors, which modulate its composition and function. Studies indicate that the gut microbiota differs in lean and obese individuals, and in individuals with different food habits. There is evidence that the relationship between diet, inflammation, insulin resistance, and cardiometabolic risk are, in part, mediated by the composition of intestinal bacteria. Knowledge about the gut microbiota may result in different strategies to manipulate bacterial populations and promote health. This review discusses the relevance of understanding the role of dietary factors or patterns in the composition of the microbiota, as well as pathophysiological mechanisms of chronic metabolic diseases, and the potential of prebiotics and probiotics on the cardiometabolic risk profile.
Gut microbiota; food habits; inflammation; insulin resistance; body adiposity
INTRODUÇÃO
A MICROBIOTA INTESTINAL DOS SERES HUMANOS
Doenças crônicas não transmissíveis (DCNTs), como diabetes melito tipo 2 (DM2),
hipertensão arterial, dislipidemia e doença cardiovascular aterosclerótica,
assumem importância crescente na saúde pública mundial em decorrência de suas
incapacitações e mortalidade precoce. A adiposidade corporal excessiva tem papel
central na gênese dessas doenças, sendo alarmante o aumento das cifras de
obesidade no Brasil no mundo (11 Instituto Brasileiro de Geografia e Estatística (IBGE). Diretoria
de Pesquisas, Coordenação de Trabalho e Rendimento. Pesquisa de Orçamentos
Familiares 2008-2009: Despesas, rendimentos e condições de vida. Rio de Janeiro:
IBGE; 2010.). Os
determinantes de obesidade são múltiplos, envolvendo fatores genéticos e
ambientais, dentre os quais se destacam as dietas com alta densidade energética
e a inatividade física. O papel das bactérias que colonizam o intestino humano
como agentes etiopatogênicos desta e de outras DCNTs ganha destaque diante por
representarem alvo potencial de intervenção (22 Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, et al. The
gut microbiota as an environmental factor that regulates fat storage. Proc Natl
Acad Sci USA. 2004;101(44):15718-23.
3 Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI.
Obesity alters gut microbial ecology. Proc Natl Acad Sci USA.
2005;102(31):11070-5.
4 Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human
gut microbes associated with obesity. Nature.
2006;444(7122):1022-3.
5 Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et
al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes.
2007;56:1761-72.-66 Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM, et
al. Selective increases of bifidobacteria in gut microflora improve
high-fat-diet-induced diabetes in mice through a mechanism associated with
endotoxaemia. Diabetologia. 2007;50(11):2374-83.).
Estima-se que em 2050 ocorrerão 15,4 milhões de mortes no mundo em decorrência de DCNTs (77 Alwan A, Maclean DR, Riley LM, d’Espaignet ET, Mathers CD, Stevens GA, et al. Monitoring and surveillance of chronic non-communicable diseases: progress and capacity in high-burden countries. Lancet. 2010;376(9755):1861-8.) e a população brasileira contribuirá de forma marcante para esse cenário. Dessa forma, justifica-se aprofundar o conhecimento sobre fatores envolvidos na sua gênese, dentre estes o papel da microbiota intestinal, cuja composição pode determinar predisposição ou proteção contra doenças.
O trato gastrointestinal (TGI) humano é o sítio orgânico mais densamente povoado por micro-organismos comensais e simbióticos, na maioria bactérias, mas também fungos, archaea e vírus (88 Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human gut microbiome. Nature. 2011;473(7346):174-80.), abrigando dez vezes mais bactérias que o número de células que formam nosso organismo (99 Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010;464(7285):59-65.).
Indivíduos apresentam composições bacterianas distintas, sendo em parte definidas
geneticamente e em outra determinadas por características individuais e
ambientais, como modo de nascimento (parto normal ou cesariana), idade e hábitos
alimentares, o que resulta numa grande variabilidade intra e interindividual
(1010 Penders J, Thijs C, Vink C, Stelma FF, Snijders B, Kummeling I, et
al. Factors influencing the composition of the intestinal microbiota in early
infancy. Pediatrics. 2006;118(2):511-21.
11 Kalliomäki M, Collado MC, Salminen S, Isolauri E. Early
differences in fecal microbiota composition in children may predict overweight.
Am J Clin Nutr. 2008;87(3):534-8.-1212 Huttenhower C, Gevers D, Knight R, Abubucker S, Badger JH,
Chinwalla AT, et al. Structure, function and diversity of the healthy human
microbiome. Nature. 2012;486(7402):207-14.). Uma das formas de avaliar as comunidades que habitam o TGI é
pela classificação taxonômica que distribui as bactérias em filos, classes,
ordem, família, gênero e espécie (88 Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR,
et al. Enterotypes of the human gut microbiome. Nature.
2011;473(7346):174-80.,1212 Huttenhower C, Gevers D, Knight R, Abubucker S, Badger JH,
Chinwalla AT, et al. Structure, function and diversity of the healthy human
microbiome. Nature. 2012;486(7402):207-14.). Calcula-se que, na microbiota
intestinal, existam cerca de mil espécies, distribuídas em mais de 50 diferentes
filos. Estudos de metagenômica indicam que, na microbiota humana, haja cerca de
3,3 milhões de diferentes genes, 150 vezes mais que o genoma humano (99 Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, et al.
A human gut microbial gene catalogue established by metagenomic sequencing.
Nature. 2010;464(7285):59-65.).
Na maioria dos indivíduos, cerca de 90% dos filos são Firmicutes e Bacteroidetes, sendo o restante composto por Actinobacterias (família Bifidobacteriaceae) e Proteobacterias (família Enterobacteriaceae). Em seguida, na ordem de frequência, aparecem os filos Synergistetes, Verrucomicrobia, Fusobacteria e Euryarchaeota, representando percentual pequeno da nossa microbiota. As principais integrantes dos Firmicutes são as classes Bacilli, Clostridia e Molicutes e dos Bacteroidetes são as Bacteroides, Flavobacteria e Sphingobacterias (88 Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human gut microbiome. Nature. 2011;473(7346):174-80.,99 Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010;464(7285):59-65.,1212 Huttenhower C, Gevers D, Knight R, Abubucker S, Badger JH, Chinwalla AT, et al. Structure, function and diversity of the healthy human microbiome. Nature. 2012;486(7402):207-14.,1313 de Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA. 2010;107(33):14691-96.).
A passagem pelo canal vaginal no momento do parto já começa a influenciar a colonização do TGI do recém-nascido. No período pós-natal, o tipo e a duração da amamentação também são impactantes. Em comparação à microbiota intestinal adulta, os lactentes apresentam maior variabilidade da composição microbiana, abrigando menos espécies com menor estabilidade. Entre 2 e 3 anos de idade, o ecossistema passa a ser estável e comparável a de um adulto, dominado pelos filos Bacteroidetes e Firmicutes (1414 Angelakis E, Armougom F, Million M, Raoult D. The relationship between gut microbiota and weight gain in humans. Future Microbiol. 2012;7(1):91-109.). Há estabilização da microbiota após a primeira infância, mas ocorrerão modificações em situações específicas ao longo da vida (1010 Penders J, Thijs C, Vink C, Stelma FF, Snijders B, Kummeling I, et al. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics. 2006;118(2):511-21.,1111 Kalliomäki M, Collado MC, Salminen S, Isolauri E. Early differences in fecal microbiota composition in children may predict overweight. Am J Clin Nutr. 2008;87(3):534-8.). Indivíduos adultos podem ter variações na proporção das bactérias em consequência de alterações ambientais ou de estados patológicos (33 Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI. Obesity alters gut microbial ecology. Proc Natl Acad Sci USA. 2005;102(31):11070-5.,88 Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human gut microbiome. Nature. 2011;473(7346):174-80.,1515 Wu GD, Chen J, Hoffmann C, Bittinger K, Chen Y, Sue A, et al. Linking long-term dietary patterns with gut microbial enterotypes. 2011;334(6052):105-8.). Com o envelhecimento, observam-se redução na população de Bacteroides, Bifidobacteria e menor produção de ácidos graxos de cadeia curta, assim como crescimento de anaeróbios facultativos, tais como Fusobacteria, Clostridia, Eubacteria, e maior atividade proteolítica. Essas variações parecem estar relacionadas a perda de paladar, olfato e menor ingestão alimentar (1616 Woodmansey EJ. Intestinal bacteria and ageing. J Appl Microbiol. 2007;102(5):1178-86.).
Apesar do crescente interesse no estudo da microbiota intestinal, há importantes lacunas no conhecimento sobre como esse ecossistema pode afetar a saúde, particularmente no que se refere às DCNTs. Esta revisão fornece uma visão geral dos avanços nesta área, informando sobre a participação da microbiota intestinal no contexto de frequentes anormalidades metabólicas que acometem o homem moderno e as possibilidades de modulação da alimentação.
MICROBIOTA, OBESIDADE E RISCO CARDIOMETABÓLICO
A obesidade participa na gênese da resistência à insulina bem como de morbidades,
tais como a intolerância à glicose, dislipidemia e hipertensão arterial. A
cascata de mecanismos intracelulares e eventos sistêmicos envolvidos na
agregação desses fatores de risco continua sendo alvo de investigações. A esse
contexto, acrescenta-se o interesse pelo papel da microbiota intestinal como
fator intermediário entre componentes ambientais e comportamentais e a
ocorrência de obesidade e os distúrbios metabólicos. O conhecimento sobre a
participação de bactérias intestinais em mecanismos fisiopatológicos evoluiu em
grande parte com resultados obtidos em modelos animais (33 Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI.
Obesity alters gut microbial ecology. Proc Natl Acad Sci USA.
2005;102(31):11070-5.,55 Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et
al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes.
2007;56:1761-72.,1717 Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon
JI. An obesity-associated gut microbiome with increased capacity for energy
harvest. Nature. 2006;444(7122):1027-31.
18 Cani PD, Possemiers S, Van de Wiele T, Guiot Y, Everard A, Rottier
O, et al. Changes in gut microbiota control inflammation in obese mice through a
mechanism involving GLP-2-driven improvement of gut permeability. Gut.
2009;58(8):1091-103.
19 Vijay-Kumar M, Aitken JD, Carvalho FA, Cullender TC, Mwangi S,
Srinivasan S, et al. Metabolic syndrome and altered gut microbiota in mice
lacking Toll-like receptor 5. Science. 2010;328(5975):228-31.-2020 Caricilli AM, Picardi PK, de Abreu LL, Ueno M, Prada PO, Ropelle
ER, et al. Gut microbiota is a key modulator of insulin resistance in TLR 2
knockout mice. PLoS Biol. 2011;9(12):e1001212.).
A hipertrofia do tecido adiposo acarreta distúrbios metabólicos e hemodinâmicos pela produção de diversas adipocinas que têm papel na gênese da resistência à insulina e aterosclerose (2121 Fantuzzi G, Mazzone T. Adipose tissue and atherosclerosis. Arterioscler Thromb Vasc Biol. 2007;27:996-1003.). Ambos os processos são mediados por citocinas inflamatórias, tais como TNF-a, IL-6, IL-2 e INF-g, secretadas tanto pelos adipócitos como por monócitos que infiltram esse tecido. Reconhece-se que indivíduo obeso apresenta um estado crônico de inflamação subclínica que favorece a resistência à insulina, evento central na geração de risco cardiometabólico (2222 Tchernof A, Després JP. Pathophysiology of human visceral obesity: an update. Physiol Rev. 2013;93(1):359-404.).
No início deste século, consolidaram-se as evidências de associação da microbiota com excesso de peso corporal, a partir de pesquisas em modelos animais e também em humanos (33 Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI. Obesity alters gut microbial ecology. Proc Natl Acad Sci USA. 2005;102(31):11070-5.,44 Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444(7122):1022-3.,55 Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761-72.). Constatou-se que indivíduos magros e obesos apresentam diferente composição de microbiota. Diferentes composições da microbiota, especialmente relacionadas à alimentação, podem aumentar a produção de citocinas pró-inflamatórias, alterando a expressão de genes do hospedeiro e induzindo estado patogênico capaz de facilitar o desenvolvimento de DCNTs (55 Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761-72.,1919 Vijay-Kumar M, Aitken JD, Carvalho FA, Cullender TC, Mwangi S, Srinivasan S, et al. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science. 2010;328(5975):228-31.). Considerando que a obesidade é o principal fator de risco para o DM2, o papel da microbiota também na sua gênese passou a ser alvo de investigação.
Os estudos pioneiros que observaram a relação da microbiota com obesidade foram realizados em animais livres de bactérias (germ-free) (22 Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, et al. The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci USA. 2004;101(44):15718-23.,33 Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI. Obesity alters gut microbial ecology. Proc Natl Acad Sci USA. 2005;102(31):11070-5.). Inicialmente, verificou-se que camundongos com microbiota intestinal preservada apresentaram 42% mais gordura corporal que os livres de bactérias (22 Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, et al. The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci USA. 2004;101(44):15718-23.). Na sequência, Ley e cols. observaram que camundongos ob/ob, geneticamente obesos, apresentavam maior proporção de Firmicutes em relação a Bacteroidetes quando comparados a controles eutróficos (33 Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI. Obesity alters gut microbial ecology. Proc Natl Acad Sci USA. 2005;102(31):11070-5.). Outros confirmaram essa observação, reforçando a hipótese da participação de certos filos de bactérias na regulação do balanço energético (1717 Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027-31.,2323 Turnbaugh PJ, Bäckhed F, Fulton L, Gordon JI. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe. 2008; 3(4):213-23.).
Em 2006, estudos em humanos obesos também verificaram menor proporção de Bacteroidetes em comparação com eutróficos. Ademais, quando perdem peso, a proporção de Firmicutes cai e torna-se mais parecida com a de indivíduos magros (44 Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444(7122):1022-3.). Da mesma forma, Turnbaugh e cols., avaliando a microbiota intestinal de gêmeos, encontraram menor diversidade bacteriana em obesos, que apresentaram 75% de Actinobacteria, 25% de Firmicutes e 0% de Bacteroidetes, enquanto magros apresentaram 0% de Actinobacteria, 58% de Firmicutes e 42% de Bacteroidetes (2424 Turnbaugh PJ, Hamady M, Yatsunenko T, Cantarel BL, Duncan A, Ley RE, et al. A core gut microbiome in obese and lean twins. Nature. 2009;457(7228):480-4.).
Estudos envolvendo transplante da microbiota de roedores magros e obesos para animais germ free mostraram que, em poucas semanas, houve pequeno ganho de gordura corporal e maior extração de calorias provenientes da ração nos animais que receberam a microbiota de doadores obesos comparados àqueles que receberam a microbiota de doadores magros, mesmo sem acréscimo no consumo de ração. Esses dados sugerem que características da microbiota dos animais obesos per se contribuem para ganho ponderal (22 Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, et al. The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci USA. 2004;101(44):15718-23.,1717 Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027-31.,2323 Turnbaugh PJ, Bäckhed F, Fulton L, Gordon JI. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe. 2008; 3(4):213-23.).
Apesar de os estudos pioneiros terem encontrado associação desfavorável dos Firmicutes e benéfica dos Bacteroidetes em relação à obesidade e ao risco cardiometabólico, vale destacar que outros pesquisadores não obtiveram os mesmos resultados. Duncan e cols. observaram a mesma proporção dos dois filos em obesos e eutróficos, enquanto Schwiertz e cols., maior proporção de Bacteroidetes em relação aos Firmicutes em obesos (2525 Duncan SH, Lobley GE, Holtrop G, Ince J, Johnstone AM, Louis P, Flint HJ. Human colonic microbiota associated with diet, obesity and weight loss. Int J Obes. 2008;32(11):1720-4.,2626 Schwiertz A, Taras D, Schäfer K, Beijer S, Bos NA, Donus C, et al. Microbiota and SCFA in lean and overweight healthy subjects. Obesity. 2010;18(1):190-5.). Portanto, não é universalmente constante o padrão de distribuição de filos da microbiota intestinal entre indivíduos obesos e magros.
Tem-se buscado o entendimento da participação dos gêneros e espécies em mecanismos fisiopatogênicos do ganho de peso, admitindo-se que não somente a proporção de filos influenciaria a adiposidade corporal, mas também a diversidade dos micro-organismos da microbiota (88 Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human gut microbiome. Nature. 2011;473(7346):174-80.,2424 Turnbaugh PJ, Hamady M, Yatsunenko T, Cantarel BL, Duncan A, Ley RE, et al. A core gut microbiome in obese and lean twins. Nature. 2009;457(7228):480-4.). Vale destacar que, no filo Firmicutes, há mais de 250 gêneros, enquanto no Bacteroidetes, cerca de 20 (2727 Bajzer M, Seeley RJ. Physiology: obesity and gut flora. Nature. 2006;444(7122):1009-10.). Em estudo envolvendo crianças, as eutróficas apresentavam maior quantidade de Bifidobacterium spp. e menor de Staphylococcus aureus que aquelas com sobrepeso (1111 Kalliomäki M, Collado MC, Salminen S, Isolauri E. Early differences in fecal microbiota composition in children may predict overweight. Am J Clin Nutr. 2008;87(3):534-8.). Outros pesquisadores passaram a analisar a distribuição das bactérias segundo os grupos de gêneros e espécies da microbiota (88 Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human gut microbiome. Nature. 2011;473(7346):174-80.,1515 Wu GD, Chen J, Hoffmann C, Bittinger K, Chen Y, Sue A, et al. Linking long-term dietary patterns with gut microbial enterotypes. 2011;334(6052):105-8.). Essa abordagem por agrupamentos mais específicos foi chamada de enterótipos. Em um estudo pioneiro, houve a identificação de três enterótipos, Bacteroides, Prevotella e Ruminococcus, com base em uma amostra de 39 indivíduos de seis diferentes nacionalidades (88 Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human gut microbiome. Nature. 2011;473(7346):174-80.).
Apesar de fortes evidências da participação da microbiota para o estado nutricional, oriundas tanto de modelos animais (33 Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI. Obesity alters gut microbial ecology. Proc Natl Acad Sci USA. 2005;102(31):11070-5.,44 Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444(7122):1022-3.) como humanos (44 Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444(7122):1022-3.,2525 Duncan SH, Lobley GE, Holtrop G, Ince J, Johnstone AM, Louis P, Flint HJ. Human colonic microbiota associated with diet, obesity and weight loss. Int J Obes. 2008;32(11):1720-4.), ainda não estão esclarecidas as implicações dessa compreensão no controle da obesidade. Se alterações da composição da microbiota são fatores causais de obesidade ou resultado dela, tem sido motivo de discussões na literatura. Ainda não está estabelecido o melhor método (“padrão-ouro”) para pesquisa de simbiontes bacterianos, e o emprego de diferentes metodologias e tipos de amostras e análises dificulta a comparabilidade dos estudos e generalização dos achados (1414 Angelakis E, Armougom F, Million M, Raoult D. The relationship between gut microbiota and weight gain in humans. Future Microbiol. 2012;7(1):91-109.). Além de diferenças de ordem metodológica, há problemas com o tamanho pequeno das amostras e uso inadequado de grupos controles.
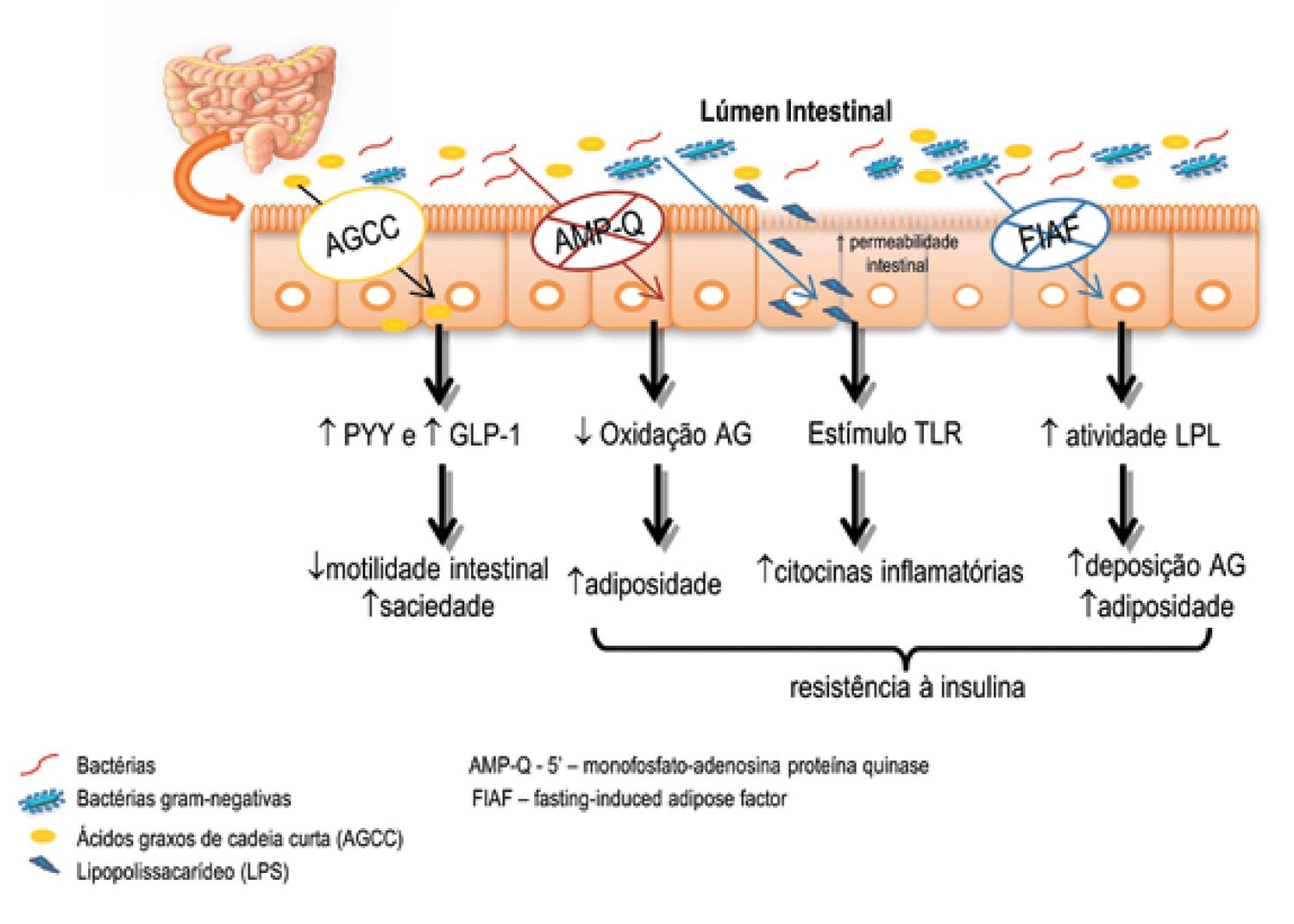
Os mecanismos envolvidos na relação entre microbiota (e sua composição) e doenças metabólicas que elevam o risco cardiovascular não estão totalmente claros, mas alguns dos mais consistentes estão descritos a seguir (Figura 1).
MECANISMOS ENVOLVIDOS NA RELAÇÃO MICROBIOTA – DOENÇAS METABÓLICAS
Fasting Induced Adipose Factor – FIAF
O Fasting Induced Adipose Factor (FIAF) é um inibidor da lipase de lipoproteína (LPL), produzido pelo intestino, fígado e tecido adiposo. Quando suprimido pela ação da microbiota intestinal, há aumento da atividade da LPL que determina a maior absorção de ácidos graxos e acúmulo de triglicerídeos nos adipócitos (22 Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, et al. The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci USA. 2004;101(44):15718-23.). Investigações em animais FIAF-deficientes mostraram que, quando alimentados com dieta ocidental, ganham mais peso corporal que animais FIAF+/+ wild-type; coerente com sua maior adiposidade, apresentam também maiores concentrações de leptina e insulina (2828 Bäckhed F, Manchester JK, Semenkovich CF, Gordon JI. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc Natl Acad Sci USA. 2007;104(3):979-84.).
Não está clara a real importância do FIAF intestinal em comparação ao produzido pelo tecido adiposo na regulação da deposição de triglicerídeos. Apesar de indícios de que possa favorecer o aumento da adiposidade corporal, a real contribuição do FIAF intestinal e sua regulação por meio de interações com populações bacterianas requerem mais investigações.
MONOFOSFATO-ADENOSINA PROTEÍNA QUINASE ATIVADA (AMP-Q)
O segundo mecanismo proposto envolve a inibição da via da 5’-monofosfato-adenosina proteína quinase (AMP-Q), enzima ativada pela adenosina monofosfato (AMP), que regula o metabolismo energético celular. Quando inibida, essa enzima ativa processos anabólicos e bloqueia catabólicos. Há evidências de que a AMP-Q desempenhe importante papel na regulação do metabolismo de ácidos graxos e da glicose, assim como na regulação do apetite. Observou-se que camundongos germ-free, mesmo com uma dieta hipercalórica, mantiveram peso, fato atribuído à elevação da atividade da AMP-Q no fígado e músculo esquelético e à maior oxidação de ácidos graxos, melhorando a sensibilidade à insulina. Tais achados sugerem que a presença da microbiota suprime a oxidação de ácidos graxos muscular via mecanismos que envolvem a inibição da AMP-Q e, portanto, favorecem a adiposidade corporal e a geração de resistência à insulina (2828 Bäckhed F, Manchester JK, Semenkovich CF, Gordon JI. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc Natl Acad Sci USA. 2007;104(3):979-84.,2929 Kola B. Role of AMP-activated protein kinase in the control of appetite. J Neuroendocrinol. 2008;20:942-51.).
EIXO CÉREBRO-INTESTINAL
O terceiro mecanismo diz respeito à sensibilidade do epitélio intestinal a produtos bacterianos. Uma linha de investigação recente refere-se ao impacto que a microbiota intestinal pode exercer no comportamento alimentar e no sistema nervoso central (SNC), de modo a influenciar a regulação central do apetite e saciedade (3030 Muccioli GG, Naslain D, Bäckhed F, Reigstad CS, Lambert DM, Delzenne NM, et al. The endocannabinoid system links gut microbiota to adipogenesis. Mol Syst Biol. 2010;6(392):1-15.). O intestino humano é capaz de digerir fibras dietéticas em grande parte devido à síntese de enzimas pela microbiota. Tais enzimas permitem a metabolização de polissacarídeos não digeríveis a monossacarídeos e a ácidos graxos de cadeia curta (AGCC), principalmente acetato, propionato e butirato (3131 Roberfroid M, Gibson GR, Hoyles L, McCartney AL, Rastall R, Rowland I, et al. Prebiotic effects: metabolic and health benefits. Br J Nutr. 2010;104(Suppl 2):S1-63.). Esses AGCC representam importante fonte de energia, de modo a favorecer a adiposidade corporal. Além disso, difundem-se nas células de forma passiva ou por transportadores da via do ácido monocarboxílico e podem atuar como sinalizadores celulares (3232 Le Poul E, Loison C, Struyf S, Springael JY, Lannoy V, Decobecq ME, et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J Biol Chem. 2003;278(28):25481-9.).
Por outro lado, há outros efeitos indiretos que podem influenciar a motilidade
intestinal e a produção de hormônios intestinais, apresentando papel na regulação da
saciedade. Os ácidos graxos de cadeia curta possuem a capacidade de se ligar aos
receptores acoplados a proteína G (GPCRs): Gpr41 e Gpr43. Atualmente, esses têm sido
denominados de receptores de ácidos graxos livres (free fatty acids
receptors – FFAR), FFAR2 e FFAR3, respectivamente (3333 Tazoe H, Otomo Y, Kaji I, Tanaka R, Karaki SI, Kuwahara A. Roles
of short-chain fatty acids receptors, GPR41 and GPR43 on colonic functions. J
Physiol Pharmacol. 2008;59(Suppl 2):251-62.
34 Holzer P, Reichmann F, Farzi A. Neuropeptide Y, peptide YY and
pancreatic polypeptide in the gut-brain axis. Neuropeptides.
2012;46(6):261-74.-3535 Ulven T. Short-chain free fatty acid receptors FFA2/GPR43 and
FFA3/GPR41 as new potential therapeutic targets. Front Endocrinol.
2012;3:111.).
Tais receptores são expressos pelas células do epitélio intestinal, em células
enteroendócrinas L, produtoras do peptídeo YY (PYY), hormônio que age inibindo a
secreção gástrica, esvaziamento gástrico, contração da vesícula biliar e reduz o
tempo de trânsito gastrointestinal. Dessa forma, ao serem ativados, esses receptores
aumentam a produção de PYY, fato que favorece a redução da motilidade intestinal e
propicia maior absorção de nutrientes do lúmen intestinal, em especial dos ácidos
graxos de cadeia curta, que são substratos para a lipogênese no fígado. Da mesma
forma, o PYY realiza suas funções hormonais no SNC, inibindo neurônios orexígenos do
núcleo arqueado, induzindo saciedade. Nesse contexto, camundongos Gpr41-/-, com ou
sem flora intestinal, apresentaram menor peso que camundongos Gpr41+/+, apesar do
mesmo consumo alimentar. Sugere-se que a Gpr41-/- esteja associada com menor
produção de PYY e com maior velocidade de trânsito intestinal, que reduz a absorção
de nutrientes (AGCC) (3333 Tazoe H, Otomo Y, Kaji I, Tanaka R, Karaki SI, Kuwahara A. Roles
of short-chain fatty acids receptors, GPR41 and GPR43 on colonic functions. J
Physiol Pharmacol. 2008;59(Suppl 2):251-62.
34 Holzer P, Reichmann F, Farzi A. Neuropeptide Y, peptide YY and
pancreatic polypeptide in the gut-brain axis. Neuropeptides.
2012;46(6):261-74.-3535 Ulven T. Short-chain free fatty acid receptors FFA2/GPR43 and
FFA3/GPR41 as new potential therapeutic targets. Front Endocrinol.
2012;3:111.). A expressão de Gpr43 parece não estar
envolvida na diferenciação do tecido adiposo, mas está possivelmente relacionada a
processos inflamatórios associados ao TNF-α (3636 Dewulf EM, Ge Q, Bindels LB, Sohet FM, Cani PD, Brichard SM, et
al. Evaluation of the relationship between GPR43 and adiposity in human. Nutr
Metab. 2013;10(1):11.).
Estudos realizados com prebióticos indicaram que uma maior produção intestinal de ácidos graxos de cadeia curta está associada ao aumento da saciedade e consequente redução da ingestão alimentar. Tais efeitos em parte relacionam-se ao aumento dos glucagon-like peptide (GLP-1 e GLP-2) e PYY, que, em conjunto com redução da grelina, ocasionam efeitos hipotalâmicos relacionados ao mecanismo de recompensa (1818 Cani PD, Possemiers S, Van de Wiele T, Guiot Y, Everard A, Rottier O, et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut. 2009;58(8):1091-103.,3737 Delzenne NM, Cani PD, Neyrinck AM. Modulation of glucagon-like peptide 1 and energy metabolism by inulin and oligofructose: experimental data. J Nutr. 2007;137(11 Suppl):2547S-51S.,3838 Cani PD, Lecourt E, Dewulf EM, Sohet FM, Pachikian BD, Naslain D, et al. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am J Clin Nutr. 2009;90:1236-43.). A estimulação do GLP-1 melhora a resposta glicêmica e insulinêmica, enquanto a secreção do GLP-2 em roedores obesos associou-se à redução da inflamação sistêmica (3939 Delzenne NM, Neyrinck AM, Cani PD. Modulation of the gut microbiota by nutrients with prebiotic properties: consequences for host health in the context of obesity and metabolic syndrome. Microb Cell Fact. 2011;10(Suppl 1):S10.).
O sistema endocanabinoide está envolvido em vários processos fisiológicos, entre eles apetite, motilidade intestinal, homeostase da glicose, função barreira do intestino e modulação da resposta inflamatória. A interação entre peptídeos estimulados pela microbiota intestinal e o sistema endocanabinoide parece estar envolvida no controle da permeabilidade intestinal e endotoxemia metabólica presente na obesidade (3030 Muccioli GG, Naslain D, Bäckhed F, Reigstad CS, Lambert DM, Delzenne NM, et al. The endocannabinoid system links gut microbiota to adipogenesis. Mol Syst Biol. 2010;6(392):1-15.,4040 Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci USA. 2013;110(22):9066-71.).
No entanto, mais estudos são necessários para compreender com precisão a função desses receptores na ação parácrina e autócrina dos ácidos graxos de cadeia curta produzidos pela microbiota intestinal.
LIPOPOLISSACARÍDEOS – LPS
As concentrações sanguíneas de lipopolissacarídeos (LPS) sofrem significativa influência da microbiota intestinal. Principalmente as bactérias Gram-negativas apresentam em sua superfície celular os LPS que funcionam como antígenos, estimulando a resposta imune do hospedeiro. Assim, a microbiota intestinal representa um grande reservatório dessa endotoxina, especialmente naqueles indivíduos que habitualmente consomem dieta com alto teor de gorduras. Esse padrão dietético pode afetar a permeabilidade intestinal por meio da secreção de mediadores, tais como o fator de necrose tumoral alfa (TNF-a), IL-1β, IL-4 e IL-13, bem como via PAR-2 (receptor ativado por protease-2), que favorecem a translocação de LPS para circulação (55 Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761-72.,2020 Caricilli AM, Picardi PK, de Abreu LL, Ueno M, Prada PO, Ropelle ER, et al. Gut microbiota is a key modulator of insulin resistance in TLR 2 knockout mice. PLoS Biol. 2011;9(12):e1001212.).
A identificação dos Toll-Like Receptors (TLR) permitiu entender como nosso organismo reconhece antígenos (como o LPS) e como ocorrem as reações pró-inflamatórias e distúrbios metabólicos. Essas anormalidades não estão presentes em animais knockout para esse tipo de receptor (1919 Vijay-Kumar M, Aitken JD, Carvalho FA, Cullender TC, Mwangi S, Srinivasan S, et al. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science. 2010;328(5975):228-31.,2020 Caricilli AM, Picardi PK, de Abreu LL, Ueno M, Prada PO, Ropelle ER, et al. Gut microbiota is a key modulator of insulin resistance in TLR 2 knockout mice. PLoS Biol. 2011;9(12):e1001212.). Vários TRLs (TRL 1, 3, 5, 6, 7, 9 e 10) têm sido descritos como mecanismos para gerar inflamação e resistência à insulina, destacando-se especialmente as descobertas relativas ao TRL2, TRL4 e TLR5.
São inúmeras as evidências de que LPS induzem inflamação e resistência à insulina (55 Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761-72.,4141 Cani PD, Bibiloni R, Knauf C, Waget A, Neyrinck AM, Delzenne NM, et al. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes. 2008;57(6):1470-81.). Essas anormalidades são desencadeadas, em parte, pela ligação dos LPS ao complexo CD14 e ao TLR4 das células imunes inatas, funcionando como gatilho para síntese de citocinas pró-inflamatórias não apenas pelas células do sistema imune, como também pelo tecido adiposo, promovendo uma endotoxemia metabólica. Assim, altas concentrações circulantes de LPS, associadas à ingestão de dietas ricas em gorduras, podem desencadear inflamação subclínica crônica, que participa na gênese da obesidade, do DM2 e outras (55 Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761-72.,4242 Tsukumo DML, Carvalho-filho MA, Carvalheira BC, Prada O, Hirabara SM, Schenka A, et al. Loss-of-function mutation in toll-like receptor 4 prevents diet-induced obesity and insulin resistance. Diabetes. 2007;56:1986-98.).
Estudo recente verificou que camundongos TLR2-/- alimentados com dieta balanceada apresentavam maior proporção de Firmicutes, aumento da inflamação pela maior captação de LPS e da resistência à insulina do que controles, e o tratamento com antibióticos foi capaz de reverter esse quadro. No mesmo estudo, camundongos germ free TLR2-/- apresentaram aumento da sensibilidade à insulina em tecido adiposo e muscular. Tais achados sugeriram fortemente que a microbiota participava de alterações inflamatórias e metabólicas (2020 Caricilli AM, Picardi PK, de Abreu LL, Ueno M, Prada PO, Ropelle ER, et al. Gut microbiota is a key modulator of insulin resistance in TLR 2 knockout mice. PLoS Biol. 2011;9(12):e1001212.).
Outros investigadores observaram que camundongos TLR5-/- apresentavam alterações na composição da microbiota intestinal e características da síndrome metabólica. Esses animais exibiam hiperfagia, dislipidemia, hipertensão arterial, resistência à insulina e aumento da adiposidade corporal. Possivelmente, essas ações eram mediadas pelo LPS, atuando em TLRs (1919 Vijay-Kumar M, Aitken JD, Carvalho FA, Cullender TC, Mwangi S, Srinivasan S, et al. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science. 2010;328(5975):228-31.,2020 Caricilli AM, Picardi PK, de Abreu LL, Ueno M, Prada PO, Ropelle ER, et al. Gut microbiota is a key modulator of insulin resistance in TLR 2 knockout mice. PLoS Biol. 2011;9(12):e1001212.).
Vários TLRs respondem às estruturas bacterianas e, uma vez ativados, podem induzir ou atenuar a resistência à insulina. Tomadas em conjunto, as descobertas sobre os mecanismos de ação da microbiota sugerem que esse ecossistema pode contribuir diretamente para o metabolismo do hospedeiro, afetando a homeostase energética, modificando as atividades enteroendócrinas e, ainda, contribuindo para a inflamação crônica subclínica, via sinalizações celulares pró-inflamatórias. No entanto, outros mecanismos moleculares devem estar envolvidos na modulação da microbiota (1919 Vijay-Kumar M, Aitken JD, Carvalho FA, Cullender TC, Mwangi S, Srinivasan S, et al. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science. 2010;328(5975):228-31.,2020 Caricilli AM, Picardi PK, de Abreu LL, Ueno M, Prada PO, Ropelle ER, et al. Gut microbiota is a key modulator of insulin resistance in TLR 2 knockout mice. PLoS Biol. 2011;9(12):e1001212.). De todos os fatores exógenos capazes de alterar a composição da microbiota intestinal, nenhum deles é mais severo do que os causados pelos antibióticos. Inclusive, esses fármacos têm sido empregados em investigações experimentais sobre o papel da microbiota na geração de inflamação e resistência à insulina, defeitos básicos em doenças como a obesidade e o DM2 (2020 Caricilli AM, Picardi PK, de Abreu LL, Ueno M, Prada PO, Ropelle ER, et al. Gut microbiota is a key modulator of insulin resistance in TLR 2 knockout mice. PLoS Biol. 2011;9(12):e1001212.,4343 Carvalho BM, Guadagnini D, Tsukumo DM, Schenka AA, Latuf-Filho P, Vassallo J, et al. Modulation of gut microbiota by antibiotics improves insulin signalling in high-fat fed mice. Diabetologia. 2012;55(10):2823-34.). Porém, a modulação da microbiota por meio da dieta talvez seja a forma mais simples, fisiológica, eficaz e de maior adesão para obter modificações no perfil de risco cardiometabólico.
MODULAÇÃO DA MICROBIOTA INTESTINAL
Micro-organismos que colonizam o intestino podem alterar a expressão gênica em células da mucosa intestinal e, em última instância, alterar a função do TGI. Em geral, a microbiota intestinal é composta em sua maioria por bactérias não patogênicas e promotoras de saúde, mas, em pequena parte, por bactérias potencialmente patogênicas.
A dieta se constitui em fator determinante das características da colonização intestinal. Esta é altamente influenciada pelos hábitos alimentares de longo prazo e por fenótipos do hospedeiro, não sendo abruptamente alterada por intervenções de curto prazo (1515 Wu GD, Chen J, Hoffmann C, Bittinger K, Chen Y, Sue A, et al. Linking long-term dietary patterns with gut microbial enterotypes. 2011;334(6052):105-8.,4444 Zhang C, Zhang M, Wang S, Han R, Cao Y, Hua W, et al. Interactions between gut microbiota, host genetics and diet relevant to development of metabolic syndromes in mice. ISME J. 2010;4(2):232-41.). Há importantes lacunas no conhecimento sobre como a alimentação e outros hábitos de vida poderiam alterar a composição da microbiota e como esta última modularia positiva ou negativamente o balanço energético (1414 Angelakis E, Armougom F, Million M, Raoult D. The relationship between gut microbiota and weight gain in humans. Future Microbiol. 2012;7(1):91-109.). Algumas dietas podem modificar o padrão de colonização intestinal desde o início da vida (1010 Penders J, Thijs C, Vink C, Stelma FF, Snijders B, Kummeling I, et al. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics. 2006;118(2):511-21.,1111 Kalliomäki M, Collado MC, Salminen S, Isolauri E. Early differences in fecal microbiota composition in children may predict overweight. Am J Clin Nutr. 2008;87(3):534-8.). Sugere-se que a alimentação tenha efeito direto sobre a microbiota, que, em última análise, resultaria em mudanças em reações bioquímicas no lúmen intestinal (4545 Hemarajata P, Versalovic J. Effects of probiotics on gut microbiota: mechanisms of intestinal immunomodulation and neuromodulation. Ther Adv Gastroenterol. 2013;6(1):39-51.). No entanto, a definição desse efeito ainda é algo a ser explorado, visto que a maioria dos estudos existentes apresenta fatores de confusão e/ou baixa comparabilidade por lançarem mão de diferentes modelos experimentais, tais como diferentes métodos de análise, espécie estudada (maioria dos estudos feitos em animais), falta de padronização dos graus de adiposidade corporal, da idade dos participantes e da dieta empregada. Os achados diante de diferentes padrões dietéticos ou nutrientes específicos serão discutidos em tópicos a seguir (Tabela 1).
DIETA HIPERLIPÍDICA (“OCIDENTAL”)
Numerosas evidências apontam que a qualidade da dieta tem importante potencial modulador da composição da microbiota intestinal, principalmente no que se refere ao teor de gorduras, que, quando excessivo, pode afetar a integridade da mucosa e prejudicar sua permeabilidade (55 Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761-72.,6, 18,38,44,46). Estudos em roedores mostraram que a composição dietética tem papel determinante na modulação da microbiota intestinal, influenciando em 57% a variação da microbiota, enquanto apenas 12% estariam relacionados a fatores genéticos (4444 Zhang C, Zhang M, Wang S, Han R, Cao Y, Hua W, et al. Interactions between gut microbiota, host genetics and diet relevant to development of metabolic syndromes in mice. ISME J. 2010;4(2):232-41.).
Em experimentos utilizando camundongos germ-free transplantados com microbiota intestinal humana, alimentados com ração com alto teor de gordura e açúcar, foram detectadas alterações rápidas na composição da microbiota intestinal com aumento no número de Firmicutes e redução de Bacteroidetes (4747 Goodman AL, Kallstrom G, Faith JJ, Reyes A, Moore A, Dantas G, et al. Extensive personal human gut microbiota culture collections characterized and manipulated in gnotobiotic mice. Proc Natl Acad Sci USA. 2011;108(15):6252-7.). Estudo recente em humanos detectou a presença mais frequente de dois enterótipos na microbiota intestinal: o tipo 1, rico em Bacteroides, fortemente associado ao consumo de proteína animal e gordura saturada, e o tipo 2, rico em Prevotella, associado à dieta baseada em carboidratos, composta por açúcares simples e fibras (1515 Wu GD, Chen J, Hoffmann C, Bittinger K, Chen Y, Sue A, et al. Linking long-term dietary patterns with gut microbial enterotypes. 2011;334(6052):105-8.).
Há evidências consistentes de que o LPS induz inflamação subclínica e resistência à insulina. Em condições normais, a presença de LPS no lúmen intestinal não é nociva à saúde. Porém, certos padrões dietéticos podem exacerbar a transferência de LPS para circulação (55 Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761-72.,66 Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM, et al. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia. 2007;50(11):2374-83.,1818 Cani PD, Possemiers S, Van de Wiele T, Guiot Y, Everard A, Rottier O, et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut. 2009;58(8):1091-103.,3838 Cani PD, Lecourt E, Dewulf EM, Sohet FM, Pachikian BD, Naslain D, et al. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am J Clin Nutr. 2009;90:1236-43.,4646 Mani V, Hollis JH, Gabler NK. Dietary oil composition differentially modulates intestinal endotoxin transport and postprandial endotoxemia. Nutr Metab. 2013;10(1):6.). A dieta hiperlipídica pode afetar indiretamente a permeabilidade intestinal por meio da ativação de mastócitos na mucosa intestinal, os quais a regulam diretamente por meio da secreção de mediadores – tais como TNF-a, IL-1B, IL-4 e IL-13 – que favorecem a translocação do LPS pela parede intestinal (55 Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761-72.,1818 Cani PD, Possemiers S, Van de Wiele T, Guiot Y, Everard A, Rottier O, et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut. 2009;58(8):1091-103.,3838 Cani PD, Lecourt E, Dewulf EM, Sohet FM, Pachikian BD, Naslain D, et al. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am J Clin Nutr. 2009;90:1236-43.). Estudo envolvendo 201 homens sugeriu que alterações na dieta, com aumento do valor calórico total independente do teor de gordura, eram capazes de induzir endotoxemia metabólica (4848 Amar J, Burcelin R, Ruidavets JB, Cani PD, Fauvel J, Alessi MC, et al. Energy intake is associated with endotoxemia in apparently healthy men. Am J Clin Nutr. 2008;87(5):1219-23.). Esses resultados contrastam com os obtidos em modelo animal nos quais foi a dieta hiperlipídica que provocou aumento significativo nas concentrações circulantes de LPS (66 Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM, et al. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia. 2007;50(11):2374-83.,4949 de La Serre CB, Ellis CL, Lee J, Hartman AL, Rutledge JC, Raybould HE. Propensity to high-fat diet-induced obesity in rats is associated with changes in the gut microbiota and gut inflammation. Am J Physiol Gastrointest Liver Physiol. 2010;299(2):G440-8.).
Vale destacar que dietas com alto teor de gorduras podem induzir alterações na microbiota intestinal sem necessariamente associarem-se à obesidade. Estudo de ratos alimentados com ração hiperlipídica concluiu que essa alimentação causou redução significativa no número total de bactérias com aumento da proporção relativa das famílias Clostridiales e Bacteroidales, alterações estas encontradas independentemente do peso dos animais (4949 de La Serre CB, Ellis CL, Lee J, Hartman AL, Rutledge JC, Raybould HE. Propensity to high-fat diet-induced obesity in rats is associated with changes in the gut microbiota and gut inflammation. Am J Physiol Gastrointest Liver Physiol. 2010;299(2):G440-8.).
O efeito desse padrão dietético pode ser observado comparando-se a microbiota intestinal de 15 crianças com dieta tipicamente urbana (Florença, Itália) com 15 crianças que viviam em zona rural (Burkina Faso, África). Nas crianças africanas, observou-se menor proporção de bactérias da família Enterobacteriaceae (Shigella e Escherichia coli), maior quantidade de Bacteroidetes e menor quantidade de Firmicutes, comparadas às italianas. Vale destacar que a dieta das crianças africanas era basicamente vegetariana com baixo consumo de gordura e proteína animal, enquanto a das italianas era tipicamente ocidental com pouca fibra e muita gordura. Esse estudo não forneceu detalhes sobre as proporções entre os tipos de gorduras consumidas (1313 de Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA. 2010;107(33):14691-96.).
A dieta tipicamente ocidental, além de hiperlipídica, provê um baixo consumo de fibras, que leva a menor produção pela microbiota de produtos imunomodulatórios considerados essenciais, tais como os ácidos graxos de cadeia curta (1313 de Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA. 2010;107(33):14691-96.).
Recentemente, além da relação entre gordura presente na carne vermelha e microbiota, verificou-se que a L-carnitina encontrada em sua composição é metabolizada pela microbiota intestinal. As bactérias transformam a L-carnitina em trimetilamina (TMA) que, por sua vez, é absorvida e metabolizada pelo fígado em N-óxido de trimetilamina, que acelera a aterosclerose em camundongos (5050 Koeth RA, Wang Z, Levison BS, Buffa JA, Org E, Sheehy BT, et al. Intestinal microbiota metabolism of l-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med. 2013;19(5):576-85.).
PREBIÓTICOS
Muito se tem investido na busca de nutrientes com capacidade seletiva de modificação favorável na microbiota intestinal, especialmente aumentando a quantidade de bifidobactérias e lactobacilos (3131 Roberfroid M, Gibson GR, Hoyles L, McCartney AL, Rastall R, Rowland I, et al. Prebiotic effects: metabolic and health benefits. Br J Nutr. 2010;104(Suppl 2):S1-63.). Denomina-se prebiótico o ingrediente alimentar (galacto-oligossacarídeos, xilo-oligossacarídeos, fruto-oligossacarídeo, inulina, fosfo-oligossacarídeos, isomalto-oligossacarídeos, lactulose, pectina) que, após fermentação, promove mudanças na composição e/ou atividade de bactérias gastrointestinais, conferindo benefícios à saúde do hospedeiro (5151 Gibson GR, Probert HM, Loo JV, Rastall RA, Roberfroid MB. Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutr Res Rev. 2004;17:259-75.).
Camundongos obesos tratados com prebióticos (oligofrutose) apresentaram redução da concentração plasmática de LPS e citocinas, além de menor expressão hepática de marcadores de inflamação e de oxidação. A redução de inflamação associou-se à maior integridade do epitélio intestinal e melhor permeabilidade, possivelmente via GLP-1 e GLP-2 e modulação do sistema endocanabinoide (3838 Cani PD, Lecourt E, Dewulf EM, Sohet FM, Pachikian BD, Naslain D, et al. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am J Clin Nutr. 2009;90:1236-43.,5252 Everard A, Lazarevic V, Derrien M, Girard M, Muccioli GG, Muccioli GM, et al. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes. 2011;60(11):2775-86.). Recentemente, verificou-se que a administração desse mesmo prebiótico foi capaz de aumentar a quantidade de Akkermansia muciniphila, reduzida nos camundongos obesos (4040 Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci USA. 2013;110(22):9066-71.). Essa bactéria mostra-se capaz de controlar a função barreira do intestino, o estoque de massa adiposa e a homeostase da glicose em animais obesos e diabéticos tipo 2. Apesar de ser Gram-negativa e conter LPS em sua superfície, a A. muciniphila é dotada de capacidade de aumentar a espessura do muco que reveste o epitélio intestinal, diminuindo, dessa forma, a endotoxemia. No entanto, ainda é precoce afirmar sobre seus benefícios de longo prazo no organismo humano (4040 Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci USA. 2013;110(22):9066-71.,5252 Everard A, Lazarevic V, Derrien M, Girard M, Muccioli GG, Muccioli GM, et al. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes. 2011;60(11):2775-86.).
Estudos em animais e humanos obesos revelam que o uso de prebióticos traz benefícios cardiometabólicos. Descrevem-se efeitos indutores de saciedade, perda de peso e redução de circunferência da cintura, e observam-se aumentos nos níveis plasmáticos de PYY, GLP-1 e GLP-2 (3939 Delzenne NM, Neyrinck AM, Cani PD. Modulation of the gut microbiota by nutrients with prebiotic properties: consequences for host health in the context of obesity and metabolic syndrome. Microb Cell Fact. 2011;10(Suppl 1):S10.). Os benefícios dos prebióticos têm sido atribuídos ao fato de serem fermentados por bactérias como Bifidobacterium spp. (66 Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM, et al. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia. 2007;50(11):2374-83.).
O vinho tinto tem sido apontado como possível modulador da microbiota; parte dos polifenóis contidos nessa bebida não é absorvida no intestino delgado, chegando intacto no cólon, onde agem como prebióticos por interagirem com a microbiota. Polifenóis do vinho tinto determinam aumento nas populações de Proteobacteria, Fusobacteria, Firmicutes e Bacteroidetes e nos gêneros Enterococcus, Bacteroides e Prevotella e redução dos patógenos do gênero Clostridium (Clostridium histolyticum). Porém, a elevação do Bifidobacterium spp. parece ser dos principais mecanismos protetores do vinho tinto, capazes de reduzir a colesterolemia e as concentrações de proteína C reativa (5353 Queipo-Ortuño MI, Boto-Ordóñez M, Murri M, Gomez-Zumaquero JM, Clemente-Postigo M, Estruch R, et al. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am J Clin Nutr. 2012;95(6):1323-34.).
PROBIÓTICOS
Probióticos são micro-organismos viáveis que conferem benefícios à saúde quando administrados em doses suficientes (5454 Reid G, Jass J, Sebulsky MT, McCormick JK. Potential uses of probiotics in clinical practice. Clin Microbiol Rev. 2003;16:658-66.). As bactérias que têm sido frequentemente utilizadas como probióticos são Lactobacilos e Bifidobactéria, mas alguns streptococos e enterococos e até mesmo a Escherichia coli também se associam a efeitos benéficos (5555 Devine DA, Marsh PD. Prospects for the development of probiotics and prebiotics for oral applications. J Oral Microbiol. 2009; 1:1-11.).
Estudo realizado em mulheres obesas, por meio da suplementação de inulina durante três meses, verificou aumento das Bifidobactérias e Faecaliumbactéria com diminuição da concentração sérica de LPS e ligeira redução de tecido adiposo (5656 Dewulf EM, Cani PD, Claus SP, Fuentes S, Puylaert PG, Neyrinck AM, et al. Insight into the prebiotic concept: lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut. 2012; 62(8):1112-21.). Outros verificaram que baixas quantidades dessas bactérias associavam-se fortemente com obesidade e diabetes. Sugeriram que tal condição favoreceria translocação do LPS, portanto, endotoxemia e inflamação, uma vez que as bifidobactérias são essenciais para adequado funcionamento da barreira da mucosa intestinal (55 Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761-72.,1111 Kalliomäki M, Collado MC, Salminen S, Isolauri E. Early differences in fecal microbiota composition in children may predict overweight. Am J Clin Nutr. 2008;87(3):534-8.,1818 Cani PD, Possemiers S, Van de Wiele T, Guiot Y, Everard A, Rottier O, et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut. 2009;58(8):1091-103.).
Recentemente, os efeitos da suplementação com essas bactérias têm sido descritos em estudos clínicos que indicam sua capacidade em induzir alterações benéficas no TGI ou melhorar a funcionalidade das comunidades microbianas preexistentes por meio de mecanismos como a competição por nutrientes, produção de substratos de crescimento ou inibição e, ainda, modulação da imunidade intestinal. Cepas probióticas de Lactobacilos melhoram a integridade da barreira intestinal, resultando na manutenção da tolerância imunológica e diminuição da translocação de bactérias pela mucosa intestinal (4545 Hemarajata P, Versalovic J. Effects of probiotics on gut microbiota: mechanisms of intestinal immunomodulation and neuromodulation. Ther Adv Gastroenterol. 2013;6(1):39-51.). Voluntários saudáveis submetidos a experimento de seis semanas com adição de bactérias probióticas (Lactobacillus acidophilus, L. casei, e L. rhamnosus) apresentaram alterações na expressão de genes envolvidos na imunidade e outras funções da mucosa (5757 Van Baarlen P, Troost F, Van der Meer C, Hooiveld G, Boekschoten M, Brummer RJ, et al. Human mucosal in vivo transcriptome responses to three lactobacilli indicate how probiotics may modulate human cellular pathways. Proc Natl Acad Sci USA. 2011;108(Suppl 1):4562-9.).
Kadooka e cols., utilizando Lactobacillus gasseri em indivíduos obesos, verificaram que os probióticos comparados ao placebo determinaram redução de adiposidade visceral, além de redução de peso corporal e circunferência da cintura (5858 Kadooka Y, Sato M, Imaizumi K, Ogawa A, Ikuyama K, Akai Y, et al. Regulation of abdominal adiposity by probiotics (Lactobacillus gasseri SBT2055) in adults with obese tendencies in a randomized controlled trial. Eur J Clin Nutr. 2010;64(6):636-43.). Em gestantes, pode-se observar que o uso de probióticos (Lactobacillus rhamnosus e Bifidobacterium lactis) determinou menor risco de diabetes gestacional, além de comprovar segurança da utilização nessa condição, e potencial proteção contra doenças metabólicas futuras (5959 Luoto R, Laitinen K, Nermes M, Isolauri E. Impact of maternal probiotic-supplemented dietary counselling on pregnancy outcome and prenatal and postnatal growth: a double-blind, placebo-controlled study. Br J Nutr. 2010;103(12):1792-9.).
Entretanto, pouco se sabe sobre como esses probióticos poderiam impactar na população bacteriana intestinal autóctone, já que há uma estabilidade desse ecossistema, no qual as bactérias se autorregulam e variáveis de confusão são de difícil controle (6060 McNulty NP, Yatsunenko T, Hsiao A, Faith JJ, Muegge BD, Goodman AL, et al. The impact of a consortium of fermented milk strains on the gut microbiome of gnotobiotic mice and monozygotic twins. Sci Transl Med. 2011;26;3(106):106ra106.). Cepas probióticas de Lactobacillus podem melhorar a integridade da barreira intestinal, porém, leites fermentados contendo vários probióticos não alteraram a composição da microbiota intestinal de camundongos germ free e gêmeos monozigóticos (6060 McNulty NP, Yatsunenko T, Hsiao A, Faith JJ, Muegge BD, Goodman AL, et al. The impact of a consortium of fermented milk strains on the gut microbiome of gnotobiotic mice and monozygotic twins. Sci Transl Med. 2011;26;3(106):106ra106.). Assim, mais estudos são necessários para avaliar de que forma é possível induzir alteração duradoura da microbiota e como essas alterações se associariam com benefícios clínicos no hospedeiro de longo prazo.
CONCLUSÕES
Obesidade, distúrbios do metabolismo glicídico e lipídico e hipertensão arterial, que favorecem a ocorrência de doença cardiovascular aterosclerótica, representam graves problemas de saúde pública em todo o mundo, estando entre as principais causas de mortalidade em países desenvolvidos e em desenvolvimento. A compreensão sobre o papel dos micro-organismos intestinais representa importante caminho de investigação, capaz de contribuir para melhora desse quadro de morbimortalidade.
A microbiota intestinal ganhou atenção da comunidade científica devido à sua participação em mecanismos que favorecem saúde ou doença. Padrões dietéticos interferem na composição da microbiota e têm relevância na modulação metabólica e regulação da adiposidade corporal. Maior proporção de Firmicutes em relação aos Bacteroidetes relaciona-se a obesidade e distúrbios metabólicos. Não apenas as proporções entre filos, mas também a variabilidade de micro-organismos, é relevante na fisiopatogênese de doenças. Identificação de gêneros e espécies em diferentes modelos experimentais tem possibilitado ampliar o conhecimento sobre os efeitos metabólicos da microbiota. Mais recentemente tem sido avaliado o impacto dos enterótipos nos processos fisiológicos e patológicos.
A supressão do FIAF, inibição da AMP-Q ativada, estímulo aos receptores FFAR e SECB e translocação de LPS são alguns mecanismos pelos quais a microbiota participa desses processos. Estes favorecem deposição de ácidos graxos, aumentam a adiposidade corporal, estimulam citocinas inflamatórias e induzem resistência à insulina, deteriorando o perfil cardiometabólico.
Intervenções adequadamente delineadas em fatores dietéticos são necessárias para compreender possíveis relações causais entre dieta e doenças, mediadas pela microbiota intestinal. O uso de probióticos e prebióticos na alimentação pode ser medida preventiva ou terapêutica, por favorecer uma composição saudável e maior funcionalidade da microbiota, diminuindo LPS circulante, portanto, a endotoxemia e a inflamação crônica subclínica. Porém, desconhece-se se em longo prazo poderia impactar na ocorrência de DCNTs.
Estudos em andamento sobre a relação “padrões de dietas, bactérias intestinais e resposta imune-inflamatória” deverão auxiliar na prevenção e no controle das principais doenças crônicas que afetam a humanidade.
REFERÊNCIAS
-
1Instituto Brasileiro de Geografia e Estatística (IBGE). Diretoria de Pesquisas, Coordenação de Trabalho e Rendimento. Pesquisa de Orçamentos Familiares 2008-2009: Despesas, rendimentos e condições de vida. Rio de Janeiro: IBGE; 2010.
-
2Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, et al. The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci USA. 2004;101(44):15718-23.
-
3Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI. Obesity alters gut microbial ecology. Proc Natl Acad Sci USA. 2005;102(31):11070-5.
-
4Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444(7122):1022-3.
-
5Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761-72.
-
6Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM, et al. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia. 2007;50(11):2374-83.
-
7Alwan A, Maclean DR, Riley LM, d’Espaignet ET, Mathers CD, Stevens GA, et al. Monitoring and surveillance of chronic non-communicable diseases: progress and capacity in high-burden countries. Lancet. 2010;376(9755):1861-8.
-
8Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human gut microbiome. Nature. 2011;473(7346):174-80.
-
9Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010;464(7285):59-65.
-
10Penders J, Thijs C, Vink C, Stelma FF, Snijders B, Kummeling I, et al. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics. 2006;118(2):511-21.
-
11Kalliomäki M, Collado MC, Salminen S, Isolauri E. Early differences in fecal microbiota composition in children may predict overweight. Am J Clin Nutr. 2008;87(3):534-8.
-
12Huttenhower C, Gevers D, Knight R, Abubucker S, Badger JH, Chinwalla AT, et al. Structure, function and diversity of the healthy human microbiome. Nature. 2012;486(7402):207-14.
-
13de Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA. 2010;107(33):14691-96.
-
14Angelakis E, Armougom F, Million M, Raoult D. The relationship between gut microbiota and weight gain in humans. Future Microbiol. 2012;7(1):91-109.
-
15Wu GD, Chen J, Hoffmann C, Bittinger K, Chen Y, Sue A, et al. Linking long-term dietary patterns with gut microbial enterotypes. 2011;334(6052):105-8.
-
16Woodmansey EJ. Intestinal bacteria and ageing. J Appl Microbiol. 2007;102(5):1178-86.
-
17Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027-31.
-
18Cani PD, Possemiers S, Van de Wiele T, Guiot Y, Everard A, Rottier O, et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut. 2009;58(8):1091-103.
-
19Vijay-Kumar M, Aitken JD, Carvalho FA, Cullender TC, Mwangi S, Srinivasan S, et al. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science. 2010;328(5975):228-31.
-
20Caricilli AM, Picardi PK, de Abreu LL, Ueno M, Prada PO, Ropelle ER, et al. Gut microbiota is a key modulator of insulin resistance in TLR 2 knockout mice. PLoS Biol. 2011;9(12):e1001212.
-
21Fantuzzi G, Mazzone T. Adipose tissue and atherosclerosis. Arterioscler Thromb Vasc Biol. 2007;27:996-1003.
-
22Tchernof A, Després JP. Pathophysiology of human visceral obesity: an update. Physiol Rev. 2013;93(1):359-404.
-
23Turnbaugh PJ, Bäckhed F, Fulton L, Gordon JI. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe. 2008; 3(4):213-23.
-
24Turnbaugh PJ, Hamady M, Yatsunenko T, Cantarel BL, Duncan A, Ley RE, et al. A core gut microbiome in obese and lean twins. Nature. 2009;457(7228):480-4.
-
25Duncan SH, Lobley GE, Holtrop G, Ince J, Johnstone AM, Louis P, Flint HJ. Human colonic microbiota associated with diet, obesity and weight loss. Int J Obes. 2008;32(11):1720-4.
-
26Schwiertz A, Taras D, Schäfer K, Beijer S, Bos NA, Donus C, et al. Microbiota and SCFA in lean and overweight healthy subjects. Obesity. 2010;18(1):190-5.
-
27Bajzer M, Seeley RJ. Physiology: obesity and gut flora. Nature. 2006;444(7122):1009-10.
-
28Bäckhed F, Manchester JK, Semenkovich CF, Gordon JI. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc Natl Acad Sci USA. 2007;104(3):979-84.
-
29Kola B. Role of AMP-activated protein kinase in the control of appetite. J Neuroendocrinol. 2008;20:942-51.
-
30Muccioli GG, Naslain D, Bäckhed F, Reigstad CS, Lambert DM, Delzenne NM, et al. The endocannabinoid system links gut microbiota to adipogenesis. Mol Syst Biol. 2010;6(392):1-15.
-
31Roberfroid M, Gibson GR, Hoyles L, McCartney AL, Rastall R, Rowland I, et al. Prebiotic effects: metabolic and health benefits. Br J Nutr. 2010;104(Suppl 2):S1-63.
-
32Le Poul E, Loison C, Struyf S, Springael JY, Lannoy V, Decobecq ME, et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J Biol Chem. 2003;278(28):25481-9.
-
33Tazoe H, Otomo Y, Kaji I, Tanaka R, Karaki SI, Kuwahara A. Roles of short-chain fatty acids receptors, GPR41 and GPR43 on colonic functions. J Physiol Pharmacol. 2008;59(Suppl 2):251-62.
-
34Holzer P, Reichmann F, Farzi A. Neuropeptide Y, peptide YY and pancreatic polypeptide in the gut-brain axis. Neuropeptides. 2012;46(6):261-74.
-
35Ulven T. Short-chain free fatty acid receptors FFA2/GPR43 and FFA3/GPR41 as new potential therapeutic targets. Front Endocrinol. 2012;3:111.
-
36Dewulf EM, Ge Q, Bindels LB, Sohet FM, Cani PD, Brichard SM, et al. Evaluation of the relationship between GPR43 and adiposity in human. Nutr Metab. 2013;10(1):11.
-
37Delzenne NM, Cani PD, Neyrinck AM. Modulation of glucagon-like peptide 1 and energy metabolism by inulin and oligofructose: experimental data. J Nutr. 2007;137(11 Suppl):2547S-51S.
-
38Cani PD, Lecourt E, Dewulf EM, Sohet FM, Pachikian BD, Naslain D, et al. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am J Clin Nutr. 2009;90:1236-43.
-
39Delzenne NM, Neyrinck AM, Cani PD. Modulation of the gut microbiota by nutrients with prebiotic properties: consequences for host health in the context of obesity and metabolic syndrome. Microb Cell Fact. 2011;10(Suppl 1):S10.
-
40Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci USA. 2013;110(22):9066-71.
-
41Cani PD, Bibiloni R, Knauf C, Waget A, Neyrinck AM, Delzenne NM, et al. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes. 2008;57(6):1470-81.
-
42Tsukumo DML, Carvalho-filho MA, Carvalheira BC, Prada O, Hirabara SM, Schenka A, et al. Loss-of-function mutation in toll-like receptor 4 prevents diet-induced obesity and insulin resistance. Diabetes. 2007;56:1986-98.
-
43Carvalho BM, Guadagnini D, Tsukumo DM, Schenka AA, Latuf-Filho P, Vassallo J, et al. Modulation of gut microbiota by antibiotics improves insulin signalling in high-fat fed mice. Diabetologia. 2012;55(10):2823-34.
-
44Zhang C, Zhang M, Wang S, Han R, Cao Y, Hua W, et al. Interactions between gut microbiota, host genetics and diet relevant to development of metabolic syndromes in mice. ISME J. 2010;4(2):232-41.
-
45Hemarajata P, Versalovic J. Effects of probiotics on gut microbiota: mechanisms of intestinal immunomodulation and neuromodulation. Ther Adv Gastroenterol. 2013;6(1):39-51.
-
46Mani V, Hollis JH, Gabler NK. Dietary oil composition differentially modulates intestinal endotoxin transport and postprandial endotoxemia. Nutr Metab. 2013;10(1):6.
-
47Goodman AL, Kallstrom G, Faith JJ, Reyes A, Moore A, Dantas G, et al. Extensive personal human gut microbiota culture collections characterized and manipulated in gnotobiotic mice. Proc Natl Acad Sci USA. 2011;108(15):6252-7.
-
48Amar J, Burcelin R, Ruidavets JB, Cani PD, Fauvel J, Alessi MC, et al. Energy intake is associated with endotoxemia in apparently healthy men. Am J Clin Nutr. 2008;87(5):1219-23.
-
49de La Serre CB, Ellis CL, Lee J, Hartman AL, Rutledge JC, Raybould HE. Propensity to high-fat diet-induced obesity in rats is associated with changes in the gut microbiota and gut inflammation. Am J Physiol Gastrointest Liver Physiol. 2010;299(2):G440-8.
-
50Koeth RA, Wang Z, Levison BS, Buffa JA, Org E, Sheehy BT, et al. Intestinal microbiota metabolism of l-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med. 2013;19(5):576-85.
-
51Gibson GR, Probert HM, Loo JV, Rastall RA, Roberfroid MB. Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutr Res Rev. 2004;17:259-75.
-
52Everard A, Lazarevic V, Derrien M, Girard M, Muccioli GG, Muccioli GM, et al. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes. 2011;60(11):2775-86.
-
53Queipo-Ortuño MI, Boto-Ordóñez M, Murri M, Gomez-Zumaquero JM, Clemente-Postigo M, Estruch R, et al. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am J Clin Nutr. 2012;95(6):1323-34.
-
54Reid G, Jass J, Sebulsky MT, McCormick JK. Potential uses of probiotics in clinical practice. Clin Microbiol Rev. 2003;16:658-66.
-
55Devine DA, Marsh PD. Prospects for the development of probiotics and prebiotics for oral applications. J Oral Microbiol. 2009; 1:1-11.
-
56Dewulf EM, Cani PD, Claus SP, Fuentes S, Puylaert PG, Neyrinck AM, et al. Insight into the prebiotic concept: lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut. 2012; 62(8):1112-21.
-
57Van Baarlen P, Troost F, Van der Meer C, Hooiveld G, Boekschoten M, Brummer RJ, et al. Human mucosal in vivo transcriptome responses to three lactobacilli indicate how probiotics may modulate human cellular pathways. Proc Natl Acad Sci USA. 2011;108(Suppl 1):4562-9.
-
58Kadooka Y, Sato M, Imaizumi K, Ogawa A, Ikuyama K, Akai Y, et al. Regulation of abdominal adiposity by probiotics (Lactobacillus gasseri SBT2055) in adults with obese tendencies in a randomized controlled trial. Eur J Clin Nutr. 2010;64(6):636-43.
-
59Luoto R, Laitinen K, Nermes M, Isolauri E. Impact of maternal probiotic-supplemented dietary counselling on pregnancy outcome and prenatal and postnatal growth: a double-blind, placebo-controlled study. Br J Nutr. 2010;103(12):1792-9.
-
60McNulty NP, Yatsunenko T, Hsiao A, Faith JJ, Muegge BD, Goodman AL, et al. The impact of a consortium of fermented milk strains on the gut microbiome of gnotobiotic mice and monozygotic twins. Sci Transl Med. 2011;26;3(106):106ra106.
Datas de Publicação
-
Publicação nesta coleção
Jun 2014
Histórico
-
Recebido
16 Ago 2013 -
Aceito
24 Jan 2014