Resumos
Dentre os fatores que contribuem para reduzir a produtividade das lavouras de milho estão as pragas, destacando-se a lagarta-do-cartucho, Spodoptera frugiperda, a qual pode reduzir a produção em até 34%. Um dos métodos de controle de pragas, cujo custo é reduzido e não causa efeitos indesejáveis ao ambiente, é o desenvolvimento de cultivares resistentes. Diversas fontes de resistência foram identificadas, linhagens desenvolvidas, registradas e liberadas ao público; entretanto, pouca informação tem sido veiculada a respeito dos mecanismos e causas dessa resistência. O objetivo do presente trabalho foi estudar os mecanismos de resistência, não-preferência e antibiose em genótipos de milho selecionados, com resistência a S. frugiperda. Os ensaios foram conduzidos em laboratório e em casa de vegetação da EMBRAPA/CNPMS. O genótipo CMS 14C apresentou antibiose influindo negativamente na biologia de S. frugiperda. 'CMS 24' e 'CMS 23' apresentaram também esse mecanismo, porém em menor intensidade. Foi constatada não-preferência alimentar das lagartas para 'Zapalote Chico' e 'BR 201'. Não-preferência para oviposição foi constatada em 'CMS 14C' e 'Zapalote Chico'.
Insecta; Zea mays; resistência de plantas; Spodoptera frugiperda
Insect pests is a major factor in limiting maize yields. The fall armyworm, Spodoptera frugiperda has been considered the most important field pest, reducing yields up to 34%. Insect resistance has been considered a low cost method of control, and represents a significant factor for preserving the environment. Several resistant lines have been released, although, the mechanisms of resistance have not yet been determined. This work was undertaken to investigate possible resistance mechanisms such as non-preference and antibiosis of S. frugiperda in some selected lines. The studies were conducted in laboratory and greenhouse at EMBRAPA/Maize and Sorghum National Research Center. The results demonstrated that the genotype CMS 14C has high level of antibiosis and 'CMS 24' and 'CMS 23' medium level. The choice tests indicated feeding non-preference to 'Zapalote Chico' and 'BR 201'. The genotypes CMS 14C and Zapalote Chico were less preferred for oviposition, compared with the remaining genotypes.
Insecta; Zea mays; host plant resistance; Spodoptera frugiperda
FITOSSANIDADE
AVALIAÇÃO DE ANTIBIOSE E NÃO-PREFERÊNCIA EM CULTIVARES DE MILHO SELECIONADOS COM RESISTÊNCIA À LAGARTA-DO-CARTUCHO(1 (1 ) Recebido para publicação em 11 de dezembro de 1998 e aceito em 11 de novembro de1999. )
PAULO AFONSO VIANA(2 (1 ) Recebido para publicação em 11 de dezembro de 1998 e aceito em 11 de novembro de1999. ); MARCOS ROBERTO POTENZA(3 (1 ) Recebido para publicação em 11 de dezembro de 1998 e aceito em 11 de novembro de1999. )
RESUMO
Dentre os fatores que contribuem para reduzir a produtividade das lavouras de milho estão as pragas, destacando-se a lagarta-do-cartucho, Spodoptera frugiperda, a qual pode reduzir a produção em até 34%. Um dos métodos de controle de pragas, cujo custo é reduzido e não causa efeitos indesejáveis ao ambiente, é o desenvolvimento de cultivares resistentes. Diversas fontes de resistência foram identificadas, linhagens desenvolvidas, registradas e liberadas ao público; entretanto, pouca informação tem sido veiculada a respeito dos mecanismos e causas dessa resistência. O objetivo do presente trabalho foi estudar os mecanismos de resistência, não-preferência e antibiose em genótipos de milho selecionados, com resistência a S. frugiperda. Os ensaios foram conduzidos em laboratório e em casa de vegetação da EMBRAPA/CNPMS. O genótipo CMS 14C apresentou antibiose influindo negativamente na biologia de S. frugiperda. 'CMS 24' e 'CMS 23' apresentaram também esse mecanismo, porém em menor intensidade. Foi constatada não-preferência alimentar das lagartas para 'Zapalote Chico' e 'BR 201'. Não-preferência para oviposição foi constatada em 'CMS 14C' e 'Zapalote Chico'.
Palavras-chave: Insecta, Zea mays, resistência de plantas, Spodoptera frugiperda.
ABSTRACT
NON-PREFERENCE AND ANTIBIOSIS OF FALL ARMYWORM FOR SELECTED CORN GENOTYPES
Insect pests is a major factor in limiting maize yields. The fall armyworm, Spodoptera frugiperda has been considered the most important field pest, reducing yields up to 34%. Insect resistance has been considered a low cost method of control, and represents a significant factor for preserving the environment. Several resistant lines have been released, although, the mechanisms of resistance have not yet been determined. This work was undertaken to investigate possible resistance mechanisms such as non-preference and antibiosis of S. frugiperda in some selected lines. The studies were conducted in laboratory and greenhouse at EMBRAPA/Maize and Sorghum National Research Center. The results demonstrated that the genotype CMS 14C has high level of antibiosis and 'CMS 24' and 'CMS 23' medium level. The choice tests indicated feeding non-preference to 'Zapalote Chico' and 'BR 201'. The genotypes CMS 14C and Zapalote Chico were less preferred for oviposition, compared with the remaining genotypes.
Key words: Insecta, Zea mays, host plant resistance, Spodoptera frugiperda.
1. INTRODUÇÃO
A cultura do milho é de grande importância para o Brasil, país onde anualmente são plantados cerca de 13 milhões de hectares, com produção de 34 milhões de toneladas de grãos (TSUNECHIRO et al., 1995). Entretanto, há fatores que contribuem para reduzir a produtividade das lavouras, como as pragas, destacando-se entre elas a lagarta-do-cartucho, Spodoptera frugiperda, a qual pode reduzir a produção em até 34% (CARVALHO, 1970). Grande parte da produção de milho no País é oriunda de pequenas propriedades onde, muitas vezes, devido a fatores econômicos e à falta de informação a respeito de tecnologias disponíveis, os agricultores não executam qualquer medida visando o controle de pragas. Em áreas nas quais a agricultura é desenvolvida, o controle dessa praga está centrado na utilização de inseticidas químicos que aumentam o custo de produção, os riscos de contaminação ambiental e conseqüente intoxicação de animais e de seres humanos.
A literatura demonstra que foram realizados vários estudos acerca da avaliação do ataque da S. frugiperda no milho. Diversas fontes de resistência foram identificadas, linhagens desenvolvidas, registradas e liberadas ao público (WISEMAN e DAVIS, 1979; WISEMAN, 1985; WISEMAN e DAVIS, 1990; OSUNA et al., 1995; WILLIAMS e DAVIS, 1997; VIANA e GUIMARÃES, 1997); entretanto, pouca informação tem sido veiculada acerca dos mecanismos e causas dessa resistência. Embora o desconhecimento do mecanismo, da herança e das causas da resistência não seja limitante para o desenvolvimento de um programa de melhoramento visando resistência a insetos, de acordo com SMITH et al. (1989) a elucidação desses parâmetros seria muito útil ao progresso do programa, contribuindo para a escolha do método de melhoramento adotado, a previsão da duração e a eficiência da resistência e para a ajuda no planejamento de novas linhas de ação a serem seguidas na solução de problemas futuros.
Os mecanismos de resistência têm sido constatados em diferentes genótipos de milho. WISEMAN et al. (1981) e WILLIAMS et al. (1983) encontraram alto nível de antibiose e não-preferência alimentar para o genótipo resistente MpSWCB-4 e baixo nível de antibiose associado à não-preferência alimentar para o 'Antigua 2D-118'. Foi também encontrada antibiose em tecidos foliares liofilizados de milho resistente, prejudicando sensivelmente o desenvolvimento da lagarta, devido à quantidade reduzida de substâncias essenciais na planta (WILLIAMS et al., 1990; WILLIAMS e BUCKEY, 1992). A não-preferência alimentar de lagartas por folhas de milho resistente foi descrita por WISEMAN et al. (1983), WISEMAN e ISENHOUR (1988) e YANG et al. (1993). A causa da resistência foi relacionada ao teor de lipídeos na cutícula das folhas.
O objetivo deste trabalho foi estudar os mecanismos de resistência (antibiose e não-preferência) em genótipos de milho com resistência a S. frugiperda, selecionados através de escala visual de danos.
2. MATERIAL E MÉTODOS
Os ensaios foram conduzidos em laboratório e em casa de vegetação da EMBRAPA Milho e Sorgo, Sete Lagoas (MG). Foram avaliados os genótipos CMS 14C, CMS 23, CMS 24, Zapalote Chico e BR 201. As condições do laboratório foram de 70% de UR, temperatura de 26 ± 1 °C e fotoperíodo de 14L:10E. Na casa de vegetação a temperatura média foi de 27 °C, e não houve controle de fotoperíodo e de UR.
2.1. Testes de preferência
Foram conduzidos ensaios de múltipla escolha e dupla escolha para estudar a atratividade de lagartas recém-eclodidas (com chance de escolha), preferência alimentar (sem chance) e oviposição (com chance). O ensaio de múltipla escolha para atratividade foi conduzido com três estádios de desenvolvimento das plantas, a saber: quatro a cinco folhas, seis a oito folhas e nove a dez folhas. Em placas de Petri, com o fundo recoberto de parafina e uma camada sobreposta de cartolina branca e papel filtro, foram espetados, em disposição circular e oposta, dois alfinetes de cada tratamento, contendo dois discos de folha de 1,5 cm de diâmetro cada um, retirados da região do cartucho da planta. No centro da placa foram liberadas 100 lagartas recém-eclodidas. Para o ensaio de dupla escolha utilizou-se o mesmo método descrito para o de múltipla escolha, diferenciando-se quanto ao número de tratamentos utilizados que, nesse caso, foram dois, dispostos alternadamente em oito alfinetes circundando a placa. A avaliação foi realizada seis horas após a liberação dos insetos, anotando-se o número de lagartas em cada genótipo. Utilizou-se o delineamento experimental inteiramente casualizado, com cinco repetições para o estádio de quatro a cinco folhas, trinta repetições para o estádio de seis a oito folhas e dez repetições para o estádio de nove a dez folhas, sendo as repetições dependentes da disponibilidade de tecido foliar. Para avaliação de preferência alimentar (sem chance de escolha) mediu-se o consumo foliar de 50 lagartas mantidas individualizadas e alimentadas com os referidos genótipos, entre o sexto dia e o final do ciclo larval. O delineamento experimental foi inteiramente casualizado, com 50 repetições.
O ensaio de preferência para oviposição foi conduzido em casa de vegetação. Os genótipos avaliados foram combinados, dois a dois, e semeados alternadamente em vasos de cinco litros de solo. Todos os vasos continham três plantas de cada genótipo, perfazendo o total de seis plantas por vaso, e foram cobertos com uma gaiola cilíndrica de 30 cm x 30 cm, recoberta com voil. Quando as plantas atingiram o estádio de quatro a cinco folhas, foram liberados três casais de mariposas, por gaiola, alimentados com solução de açúcar a 10%. Após cinco dias avaliou-se o número de posturas por planta. O delineamento experimental utilizado foi em blocos incompletos, balanceados, com 20 repetições.
2.2. Testes de antibiose
No ensaio de laboratório utilizou-se o delineamento experimental inteiramente casualizado, com 50 repetições. Lagartas recém-eclodidas foram individualizadas em copos de 50 mL e alimentadas diariamente com pedaços de folhas retiradas da região do cartucho das plantas, com quatro a cinco folhas de cada um dos referidos genótipos. Foram avaliadas a duração e a viabilidade das fases larval e pupal, a massa de pupas e a razão sexual. Paralelamente alimentaram-se outras 50 lagartas por genótipo, as quais foram divididas em dois grupos de 25, para avaliação do comprimento e massa de lagarta, largura da cápsula cefálica, aos 6 e 13 dias respectivamente. O delineamento experimental utilizado foi inteiramente casualizado, com 25 repetições.
Os adultos emergidos do lote de 50 lagartas foram agrupados em dez gaiolas de oviposição (gaiolas de PVC, de 12 cm de diâmetro e 20 cm de altura, revestidas com papel sulfite, e em cada gaiola foi colocado um casal). As extremidades da gaiola foram vedadas com placa de Petri, e os adultos alimentados com solução de açúcar a 10%. Foram avaliados o período de pré-oviposição e de oviposição e o número de posturas e de ovos por fêmea. O delineamento experimental utilizado foi inteiramente casualizado, com dez repetições.
Os parâmetros avaliados foram transformados em ou log(x), de acordo com a natureza dos dados, e submetidos à análise de variância. As médias foram separadas pelo teste de Tukey (P £ 0,05).
ou log(x), de acordo com a natureza dos dados, e submetidos à análise de variância. As médias foram separadas pelo teste de Tukey (P £ 0,05).
3. RESULTADOS E DISCUSSÃO
3.1. Testes de preferência
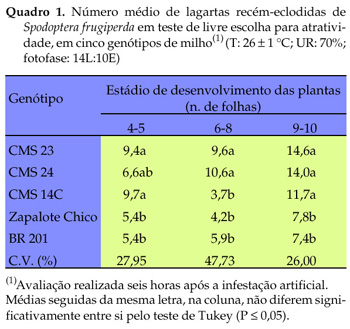
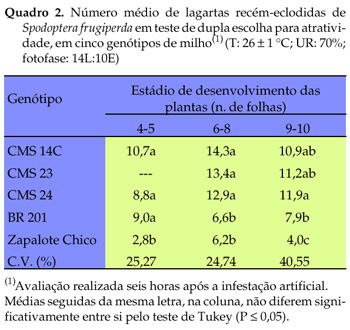
No teste de múltipla escolha para atratividade de lagartas, realizado com plantas no estádio de quatro a cinco folhas observou-se que os genótipos CMS 14C e CMS 23 foram os mais preferidos e 'BR 201' e 'Zapalote Chico', os menos preferidos (Quadro 1). No estádio de seis a oito folhas, observou-se uma inversão em relação a 'CMS 14C', o qual tornou-se menos preferido, juntamente com 'BR 201' e 'Zapalote Chico'. Essa inversão na preferência das lagartas por 'CMS 14C' ocorreu no teste de múltipla escolha, no qual o comportamento da lagarta possivelmente tenha sido afetado pela presença de vários genótipos agrupados, ao passo que na disposição de dupla escolha voltou a prevalecer a atratividade exercida por esse genótipo. Com o estádio de nove a dez folhas, 'BR 201' e 'Zapalote Chico' mantiveram-se como os menos preferidos pelas lagartas. A diferença significativa na preferência das lagartas (entre os genótipos estudados) manteve-se estável nos distintos estádios de desenvolvimento dos genótipos avaliados, exceto para 'CMS 14C', no estádio de seis a oito folhas. No teste de dupla escolha os resultados obtidos confirmam os do teste de múltipla escolha, quanto à maior atratividade das lagartas por 'CMS 14C', 'CMS 23' e 'CMS 24', e menor para 'Zapalote Chico' e 'BR 201' (Quadro 2).
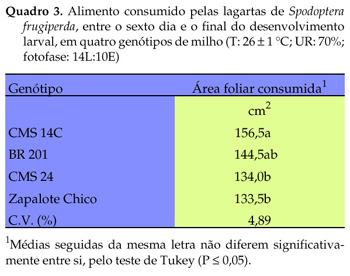
Devido à baixa germinação das sementes (que impossibilitou obter quantidade de folhas suficiente para a alimentação das lagartas, para o teste de preferência alimentar), 'CMS 23' não foi utilizado na avaliação do alimento consumido entre o sexto dia e o final do desenvolvimento larval. As lagartas alimentadas com 'CMS 14C' consumiram maior quantidade de área foliar até o final do ciclo larval, do que as alimentadas com folhas de 'CMS 24' e 'Zapalote Chico' (Quadro 3), indicando que o inseto necessita maior quantidade de alimento desse genótipo para suprir a sua dieta nutricional e, assim, completar o seu ciclo biológico.
Os testes de preferência para oviposição realizados em casa de vegetação, mostraram que as combinações com a presença dos genótipos CMS 23 e BR 201 foram as que obtiveram a maior postura, indicando maior preferência (para oviposição) do inseto por esses materiais (Quadro 4). O menor número de posturas foi encontrado nas combinações com a presença do genótipo CMS 14C, indicando menor preferência para oviposição nesse material. Observou-se, também, um incremento no número de posturas nas combinações do genótipo CMS 14C com 'CMS 23' e 'BR 201', que foram mais preferidos que as combinações de 'CMS 14C' com 'Zapalote Chico' ou com 'CMS 24'. A combinação de 'CMS 23' com 'BR 201' foi a que apresentou o maior número de posturas.
3.2. Testes de antibiose
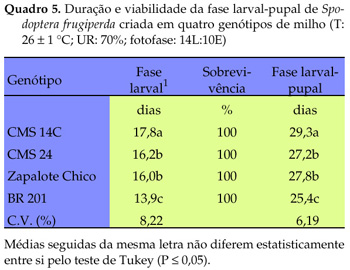
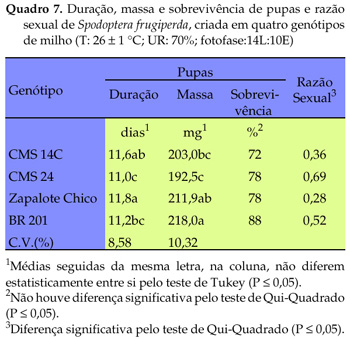
Os resultados obtidos para a fase larval-pupal de S. frugiperda diferiram significativamente entre si, evidenciando que os genótipos utilizados influenciaram as referidas variáveis (Quadro 5). Os insetos criados no genótipo CMS 14C tiveram o período larval mais longo, ao passo que aqueles criados em 'BR 201' apresentaram esse período mais curto, situando-se, de forma intermediária, os insetos criados nos genótipos CMS 24 e Zapalote Chico. A viabilidade larval para os referidos genótipos foi de 100%. Verificou-se que o desenvolvimento larval aos 6 e 13 dias de idade após a eclosão das lagartas, avaliado por três parâmetros, foi influenciado pelos genótipos estudados (Quadro 6). Aos seis dias observaram menor massa e menor comprimento de lagartas criadas nos genótipos CMS 14C, CMS 23, CMS 24 e Zapalote Chico, em relação a 'BR 201'. No genótipo CMS 14C as lagartas tiveram menor largura da cápsula cefálica do que as desenvolvidas em 'Zapalote Chico' e 'BR 201', as quais não diferiram significativamente entre si. Aos 13 dias de idade observou-se que as lagartas criadas em 'CMS 14C' diferiram significativamente quanto à massa, comprimento e largura da cápsula, das lagrtas criadas em 'Zapalote Chico' e 'BR 201', que tiveram os maiores valores para essas variáveis. A cápsula cefálica das lagartas desenvolvidas em 'CMS 23' apresentou valores intermediários aos observados para as desenvolvidas em 'CMS 14C', 'CMS 24', 'Zapalote Chico' e 'BR 201'.
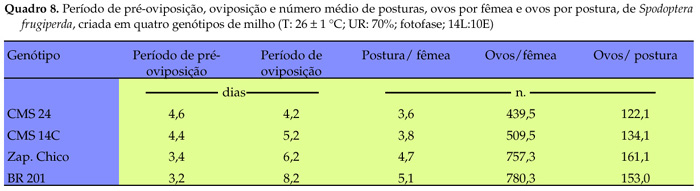
A duração da fase de pupa foi menor para as lagartas criadas nos genótipos CMS 24 e BR 201, em relação a 'Zapalote Chico' (Quadro 7). A massa de pupas foi menor para as criadas em 'CMS 24' e 'CMS 14C', em relação às criadas no híbrido BR 201, indicando que este último proporcionou melhores condições nutricionais para as lagartas, que os demais genótipos. Não houve diferença significativa na viabilidade de pupas provenientes dos insetos alimentados com os genótipos avaliados, havendo, entretanto, na proporção de machos e fêmeas. A menor razão sexual foi verificada em 'Zapalote Chico' (0,28) e a maior foi observada em 'CMS 24' (0,69). Apesar das variáveis período médio de pré-oviposição e oviposição, número médio de posturas por fêmea, ovos por postura e por fêmea não terem sido analisados estatisticamente, devido ao reduzido número de insetos que completaram o ciclo, aparentemente ocorreu maior período de oviposição, número de ovos e de posturas, em 'Zapalote Chico' e 'BR 201', uma vez que os insetos oriundos de 'CMS 24' e 'CMS 14C' apresentaram os menores valores para esses parâmetros (Quadro 8), confirmando os resultados obtidos anteriormente de ação prejudicial sobre a biologia da praga.
Os resultados indicaram que os genótipos mais atrativos e mais consumidos pelas lagartas foram, em geral, os menos adequados à sua biologia, levando a crer que há primeiramente um efeito da planta no comportamento do inseto que difere da adequação de cada genótipo ao balanço nutricional da lagarta.
WILLIAMS e DAVIS (1997) e WISEMAN (1997) referiram-se à possibilidade de ocorrência de antibiose e não-preferência alimentar para alguns genótipos, dificultando a separação desses mecanismos de resistência. Comparando-se tais resultados com os obtidos neste trabalho, verificou-se que, apesar do maior consumo de alimento das lagartas criadas em 'CMS14C', houve aumento do ciclo biológico, redução de massa, de comprimento e de largura da cápsula cefálica da lagarta, indicando que o mecanismo de antibiose ocorreu isoladamente e em maior intensidade neste tratamento. Observou-se em menor intensidade essa mesma tendência para 'CMS 24' e 'CMS 23'. Outras variáveis biológicas mensuradas também indicaram a ocorrência desse mecanismo para 'CMS 24', reduzindo o período de oviposição, número de posturas e de ovos. WISEMAN et al. (1981) detectaram a presença de antibiose em plantas de milho, no estádio de oito a dez folhas, para a lagarta-do-cartucho. Observou-se que lagartas alimentadas com genótipos resistentes apresentavam menor comprimento e menor consumo de alimento, comparadas a lagartas alimentadas em genótipos suscetíveis. WILLIAMS e DAVIS (1997), baseando-se em uma série de ensaios conduzidos para determinar os mecanismos de resistência de milho à Diatraea grandiosella e S. frugiperda, ressaltaram que substâncias tóxicas em alta concentração, características de dureza dos grãos e de outras estruturas nas folhas, são fatores provavelmente envolvidos em linhagens resistentes. Salientaram, ainda, que a ocorrência de um desses fatores pode ser responsável por parte, mas não por toda a base genética para a resistência.
Os testes de atratividade mostraram 'Zapalote Chico' como o genótipo menos preferido, seguido por 'BR 201'. As causas dessa resistência são consideradas complexas por YANG et al. (1991, 1993). Estudos químicos e ultra-estruturais das folhas de milho, conduzidos por esses autores, com outros genótipos de milho, demonstraram que lipídeos na cutícula da folha podem afetar a não-preferência para alimentação da lagarta-do-cartucho. Nos testes de oviposição, verificou-se que 'CMS 14C' e 'Zapalote Chico' foram os menos preferidos. De acordo com NG et al. (1990), em testes de preferência para oviposição as mariposas são capazes de diferenciar entre cultivares resistentes e suscetíveis, depositando número de ovos significativamente menor sobre as folhas de material resistente do que em suscetíveis.
A principal importância da elucidação dos mecanismos de resistência em genótipos selecionados foi enfatizada por ORTEGA et al. (1980). Segundo os autores, o conhecimento dos mecanismos envolvidos na resistência pode facilitar a seleção de plantas com essa característica, aumentando a eficiência do programa de melhoramento. No caso específico dos genótipos avaliados no presente trabalho, essa informação aplica-se ao desenvolvimento de híbridos experimentais com resistência a S. frugiperda que também estão sendo avaliados em algumas regiões para detectar outras características de interesse agronômico.
4. CONCLUSÕES
1) O genótipo CMS 14C apresentou antibiose como o mecanismo de resistência a S. frugiperda. 'CMS 24' e 'CM 23' apresentaram também esse mecanismo, porém em menor intensidade.
2) 'Zapalote Chico' e 'BR 201' apresentaram o mecanismo de não-preferência alimentar.
3) 'Zapalote Chico' e 'CMS 14C' mostraram não-preferência para oviposição.
(2) EMBRAPA - CNPMS, Caixa Postal 151, 35701-970 Sete Lagoas (MG).
(3) Instituto Biológico, Caixa Postal 12.898, 04010-970 São Paulo (SP).
- CARVALHO, R.P.L. Danos, flutuaçőes da populaçăo, controle e comportamento de Spodoptera frugiperda (J.E. Smith 1797), e sua suscetibilidade de diferentes genótipos de milho, em condiçőes de campo. Piracicaba, 1970. 170p. Tese (Doutorado) - ESALQ/USP.
- NG, S.S.; DAVIS, F.M.; WILLIAMS W.P. Ovipositional response of southwestern corn borer (Lepidoptera: Pyralidae) and fall armyworm (Lepidoptera: Noctuidae) to selected maize hybrids. Journal of Economic Entomology, College Park, v.83, n.3, p.1575-1577, 1990.
- ORTEGA, A.; VASAL, S.; MIHM, J.; HERSHEY, C. Breeding for insect resistance in maize. In: MAXWELL, F.G.; JENNINGS, P.R., eds. Breeding plants resistant to insects New York: John Wiley & Sons, 1980. p.371-420.
- OSUNA, J.A.; LARA, F.M.; OLIVEIRA, M.A.P.; TOZETTI, A.D. Avaliaçăo de famílias de meio-irmăos em milho visando resistęncia a Helicoverpa zea (Boddie) e Spodoptera frugiperda Anais da Sociedade Entomológica do Brasil, Londrina, v.24, n.1, p.21-26, 1995.
- SMITH, M.E.; MIHM, J.A.; JEWELL, D.C. Breeding for multiple resistance to temperate, sub tropical, and tropical maize, insect pests at CIMMYT. In: INTERNATIONAL SYMPOSIUM ON METHODOLOGIES FOR DEVELOPING HOST PLANT RESISTANCE TO MAIZE INSECTS, 1989, Mexico. Proceedings Mexico, CIMMYT, 1989. p. 222-234.
- TSUNECHIRO, A.; BESSA JUNIOR, A. de A.; GHILARDI, A.A.; FERREIRA, C.R.R.P.T. et al. Prognóstico agrícola 1995/96: algodăo, amendoim, arroz, feijăo, mandioca, milho, soja. Informaçőes Econômicas, Săo Paulo, v.25, n.9, p.13-71, 1995.
- VIANA, P.A.; GUIMARĂES, P.E.O. Maize resistance to the lesser cornstalk borer and fall armyworm in Brazil. In: INTERNATIONAL SYMPOSIUM HELD AT THE INTERNATIONAL MAIZE AND WHEAT IMPROVEMENT CENTER, 1994, Mexico. Proceedings Mexico: CIMMYT, 1997. p.112-116.
- WILLIAMS, W.P.; BUCKEY, P.M. Growth of fall armyworm (Lepidoptera: Noctuidae) larvae on resistant and susceptible corn. Journal of Economic Entomology, College Park, v.85, n.5, p.2039-2042, 1992.
- WILLIAMS, W.P.; DAVIS, F.M. Mechanisms and bases of resistance in maize to southwestern corn borer and fall armyworm. In: INTERNATIONAL SYMPOSIUM HELD AT THE INTERNATIONAL MAIZE AND WHEAT IMPROVEMENT CENTER. 1994, Mexico. Proceedings Mexico: CIMMYT, 1997. p.29-36.
- WILLIAMS, W.P.; DAVIS, F.M.; WISEMAN, B.R. Fall armyworm resistance in corn and its suppression of larval growth and survival. Agronomy Journal, Madison, v.75, n.5, p.831-832, 1983,
- WILLIAMS, W.P; BUCKEY, P.M.; HEDIN, P.A.; DAVIS, F.M. Laboratory biossay for resistance in corn to fall armyworm (Lepidoptera: Noctuidae) and southwestern corn borer (Lepidoptera: Pyralidae). Journal of Economic Entomology, College Park, v.83, n.4, p.1578-1581, 1990.
- WISEMAN, B.R. Development of resistance in corn and sorghum to a foliar and ear/panicle feeding worm complex. In: ANNUAL CORN AND SORGHUM INDUSTRY RESEARCH CONFERENCE, 40, 1985. Proceedings. Washington: ASTA, 1985. p.108-124.
- WISEMAN, B.R. Mechanisms of maize resistance to corn earworm and fall armyworm. In: INTERNATIONAL SYMPOSIUM HELD AT THE INTERNATIONAL MAIZE AND WHEAT IMPROVEMENT CENTER, 1994, Mexico. Proceedings Mexico: CIMMYT, 1997. p.46-54.
- WISEMAN, B.R.; DAVIS, F.M. Plant resistance to the armyworm. Florida Entomologist, Gainesville, v.62, n.1, p.123-130, 1979.
- WISEMAN, B.R.; DAVIS, F.M. Plant resistance to insects attacking corn and grain sorghum. Florida Entomologist, Gainesville, v.73, n.3, p.446-458, 1990.
- WISEMAN, B.R.; ISENHOUR, D.J. Feeding responses of fall armyworm larvae on excised green and yellow whorl tissue of resistant and susceptible corn. Florida Entomologist, Gainesville, v.71, n.2, p.243-249, 1988.
- WISEMAN, B.R.; DAVIS, F.M.; WILLIAMS, W.P. Fall armyworm: larval density and movement as an indication of nonpreference in resistant corn. Protection Ecology, v.5, n.2, p.135-141, 1983.
- WISEMAN, B.R.; WILLIAMS, W.P.; DAVIS, F.M. Fall armyworm: resistance mechanisms in selected corn. Journal of Economic Entomology, College Park, v.74, n.5, p.622-624, 1981.
- YANG, G.; ISENHOUR, D.J.; ESPELIE, K.E. Activity of maize leaf cuticular lipids in resistance to leaf-feeding by the fall armyworm. Florida Entomologist, Gainesville, v.74, n.2,p.229-236, 1991.
- YANG, G.; WISEMAN, B.R.; ESPELIE, K.E. Movement of neonate fall armyworm (Lepidoptera: Noctuidae) larvae on resistant and susceptible genotypes of corn. Environmental Entomology, College Park, v.22, n.3,p. 547-553, 1993.
Datas de Publicação
-
Publicação nesta coleção
10 Jan 2001 -
Data do Fascículo
2000
Histórico
-
Recebido
11 Dez 1998 -
Aceito
11 Nov 1999