Resumos
Objetivou-se avaliar o crescimento da plântula e o metabolismo de carboidratos associados ao aumento da tolerância à hipoxia exercido pela presença do cálcio no período de germinação e/ou alagamento de plântulas de milho com diferentes idades. O experimento foi desenvolvido na Universidade Federal de Lavras, MG, em 2002. Cariopses de milho da variedade 'Saracura' (BRS-4154) foram germinadas em água ou solução de CaCl2. Após dois e quatro dias, as plântulas foram submetidas ao alagamento em tubos de PVC com tampão (com ou sem CaCl2) por três dias, sendo então avaliadas a sobrevivência, massa seca e as características bioquímicas. O cálcio aumentou a sobrevivência ao alagamento de plântulas com quatro dias, entretanto, não influenciou naquelas com dois dias. O alagamento reduziu a massa seca das plântulas sem prejudicar sua recuperação pós-estresse, entretanto, o cálcio influenciou no acúmulo de massa seca pós-hipoxia somente em plântulas com quatro dias, mostrando uma recuperação mais rápida dos danos causados no período de alagamento, o que ocorre em plântulas de dois dias de germinação, independentemente desse elemento. Em plântulas mais jovens também houve maior mobilização das reservas de amido, assim como, maior translocação e utilização dessas reservas. O alagamento reduziu drasticamente a atividade da invertase ácida do vacúolo, independentemente da presença do cálcio, comprovando sua não participação nos processos comandados por essa enzima. A redução na atividade das invertases favorece menor concentração de hexoses, diminuindo a respiração anaeróbica e, conseqüentemente, a produção de substâncias tóxicas, aumentando a sobrevida das plântulas em condições de hipoxia.
Invertase; alfa-amilase; cálcio; hipoxia
This research evaluated the plantlet growth and carbohydrate metabolism associated with the increase in hypoxia tolerance caused by calcium presence during germination and/or flooding of maize plantlets with different ages. The experiment was conducted at Universidade Federal de Lavras, MG, in 2002. Maize caryopses (var.'Saracura') were germinated in water or CaCl2. After two and four days, the plantlets were submitted to flooding in PVC tubes with buffer (with or without CaCl2) during three days, being evaluated the survival, dry weight and biochemical characteristics. Calcium increased plantlet survival to flooding at four days, although there was no influence on two-day samples. Flooding reduced the plantlet dry weight without affect on the recovery after stress, therefore calcium influenced dry weight accumulation after hypoxia only in plantlets at four days, showing a faster recovery from damages caused during the flooding period, which also occurs in plantlets treated at two days of germination, but independently of this element. Younger plantlets also showed higher mobilization of the starch reserves, as well as higher translocation and utilization of these reserves. Flooding reduced drastically the vacuole acid invertase activity, independent of calcium presence, demonstrating that it has no participation in the processes coordinated by this enzyme. The reduction in the invertase activity favors a lower hexose concentration, decreasing anaerobic respiration and, consequently, the production of toxic substances, thereby increasing the survival of plantlets in hypoxia conditions.
Invertase; alpha-amylase; calcium; hypoxia
ÁREAS BÁSICAS
Crescimento de plântulas do milho 'Saracura' e atividade de a-amilase e invertases associados ao aumento da tolerância ao alagamento exercido pelo cálcio exógeno
Growth of maize 'Saracura' (BRS-4154) and a-amylase and invertases activity associated with the increase of flooding tolerance caused by exogenous calcium
Daniela Deitos FriesI, * * Autora correspondente , V; José Donizeti AlvesI, VI; Nelson Delú FilhoII; Paulo César MagalhãesIII; Patrícia de Fátima Pereira GoulartIV; Marcelo Murad MagalhãesI, VII
ISetor de Fisiologia vegetal, Departamento de Biologia, UFLA, Caixa Postal 3037, 37200-000 Lavras (MG). E-mails: friesdd@ufla.br ; jdalves@ufla.br; mmurad@ufla.br
IIUniversidade Vale do Rio Verde, Av. Castelo Branco, 82, Chácara das Rosas, 37410-000 Três Corações (MG). E-mail: ndelu@unincor.br
IIIEmbrapa Milho e Sorgo, Rodovia MG 424 km 65, 35701-970 Sete Lagoas (MG). E-mail: pcesar@cnpms.embrapa.br
IVUnilavras, Rua Padre José Poggel, 506, Centenário, 37200-000 Lavras (MG). E-mail: patriciagoulart@unilavras.edu.br
VBolsita de Pós-Doutorado da FAPEMIG
VIBolsista do CNPq
VIIBolsista Embrapa/Café
RESUMO
Objetivou-se avaliar o crescimento da plântula e o metabolismo de carboidratos associados ao aumento da tolerância à hipoxia exercido pela presença do cálcio no período de germinação e/ou alagamento de plântulas de milho com diferentes idades. O experimento foi desenvolvido na Universidade Federal de Lavras, MG, em 2002. Cariopses de milho da variedade 'Saracura' (BRS-4154) foram germinadas em água ou solução de CaCl2. Após dois e quatro dias, as plântulas foram submetidas ao alagamento em tubos de PVC com tampão (com ou sem CaCl2) por três dias, sendo então avaliadas a sobrevivência, massa seca e as características bioquímicas. O cálcio aumentou a sobrevivência ao alagamento de plântulas com quatro dias, entretanto, não influenciou naquelas com dois dias. O alagamento reduziu a massa seca das plântulas sem prejudicar sua recuperação pós-estresse, entretanto, o cálcio influenciou no acúmulo de massa seca pós-hipoxia somente em plântulas com quatro dias, mostrando uma recuperação mais rápida dos danos causados no período de alagamento, o que ocorre em plântulas de dois dias de germinação, independentemente desse elemento. Em plântulas mais jovens também houve maior mobilização das reservas de amido, assim como, maior translocação e utilização dessas reservas. O alagamento reduziu drasticamente a atividade da invertase ácida do vacúolo, independentemente da presença do cálcio, comprovando sua não participação nos processos comandados por essa enzima. A redução na atividade das invertases favorece menor concentração de hexoses, diminuindo a respiração anaeróbica e, conseqüentemente, a produção de substâncias tóxicas, aumentando a sobrevida das plântulas em condições de hipoxia.
Palavras-chave: Invertase, a-amilase, cálcio, hipoxia.
ABSTRACT
This research evaluated the plantlet growth and carbohydrate metabolism associated with the increase in hypoxia tolerance caused by calcium presence during germination and/or flooding of maize plantlets with different ages. The experiment was conducted at Universidade Federal de Lavras, MG, in 2002. Maize caryopses (var.'Saracura') were germinated in water or CaCl2. After two and four days, the plantlets were submitted to flooding in PVC tubes with buffer (with or without CaCl2) during three days, being evaluated the survival, dry weight and biochemical characteristics. Calcium increased plantlet survival to flooding at four days, although there was no influence on two-day samples. Flooding reduced the plantlet dry weight without affect on the recovery after stress, therefore calcium influenced dry weight accumulation after hypoxia only in plantlets at four days, showing a faster recovery from damages caused during the flooding period, which also occurs in plantlets treated at two days of germination, but independently of this element. Younger plantlets also showed higher mobilization of the starch reserves, as well as higher translocation and utilization of these reserves. Flooding reduced drastically the vacuole acid invertase activity, independent of calcium presence, demonstrating that it has no participation in the processes coordinated by this enzyme. The reduction in the invertase activity favors a lower hexose concentration, decreasing anaerobic respiration and, consequently, the production of toxic substances, thereby increasing the survival of plantlets in hypoxia conditions.
Key words: Invertase, a-amylase, calcium, hypoxia.
1. INTRODUÇÃO
O excesso de água no solo promove deficiência de oxigênio, tornando o ambiente radicular hipóxico. Como conseqüência da respiração dos órgãos submersos da planta e/ou de microrganismos, o oxigênio é esgotado e esse ambiente torna-se anóxico. Em qualquer dessas situações, o desenvolvimento das plantas é prejudicado. Plantas tolerantes ou que apresentam certo grau de tolerância desenvolvem estruturas ou mecanismos que lhe permitem sobreviver por períodos mais prolongados nessas condições (ALVES et al., 2000; BRAY et al., 2000).
Na falta de oxigênio, o metabolismo é desviado para a via anaeróbica que, além de ter baixo rendimento energético, produz substâncias tóxicas, tais como etanol e ácido lático. Em contrapartida, a expressão de genes responsáveis pela síntese de enzimas do metabolismo de carboidratos (ANDREWS et al., 1994; GUGLIELMINETTI et al., 1995; RICARD et al.; 1998), assim como a formação de aerênquimas e raízes adventíceas (HE et al., 1994; KOLB et al., 1998; ZENG et al., 1999) parecem estar relacionadas aos mecanismos de tolerância em plantas.
Entre as plantas de interesse econômico no Brasil, o milho faz parte das culturas cujo cultivo está sujeito a ambientes encharcados, tendo a produtividade afetada devido à sua baixa tolerância a essas condições. Em 1997, foi lançada pela Embrapa Milho e Sorgo a variedade de milho denominada 'Saracura' (BRS-4154), com alto grau de tolerância ao alagamento em condições de campo (PARENTONNI et al., 1997).
Verificou-se que em ambiente semicontrolado, o milho 'Saracura' foi muito superior na tolerância ao alagamento quando comparado com uma variedade não tolerante (BR-107). Entretanto, com o prolongamento do estresse, observou-se intensa lise celular no mesocótilo, levando ao tombamento e à morte da plântula, características mais precocemente observadas na variedade BR-107 (VITORINO et al., 2001).
Várias estratégias têm sido utilizadas com o objetivo de aumentar a recuperação pós-estresse das culturas submetidas à deficiência de oxigênio, dentre elas, a aplicação exógena de cálcio. O fornecimento de cálcio prolongou a sobrevivência da variedade 'Saracura', aumentando o período de tolerância de dois para quatro dias, sem prejuízo na taxa de sobrevivência (ALVES et al., 2002). A atuação do cálcio endógeno na sobrevivência de plântulas de milho ao estresse anaeróbico também foi evidenciada por SUBBAIAH et al. (1994), que verificaram que a mobilização desse elemento de reservatórios intracelulares até o citoplasma induziu a síntese da desidrogenase alcoólica e sintase da sacarose.
A presença do cálcio no período de germinação promoveu um aumento na sobrevivência à hipoxia de plântulas do milho 'Saracura', entretanto, foi mais eficiente quando esse elemento esteve presente também no alagamento. O alto índice de sobrevivência das plântulas na presença do cálcio se deve à redução do crescimento da plântula exercido pela alta pressão osmótica da solução adicionada no papel de germinação. Essa redução no crescimento ocorreu principalmente quando esse elemento foi adicionado por mais de 24 horas e os maiores índices de sobrevivência estavam relacionados aos tratamentos com maiores reduções no crescimento (ALVES et al., 2002). Ao avaliar plântulas do milho 'Saracura' germinadas por dois e quatro dias na presença de cálcio, FRIES (2003) verificou que a redução no crescimento em função desse elemento foi significativa somente em plântulas mais velhas. Esse menor desenvolvimento esteve relacionado a uma menor utilização de açúcares solúveis pela plântula, uma vez que houve maior degradação das reservas de amido do endosperma e conseqüente translocação na presença do cálcio. Além disso, a menor atividade da invertase ácida do vacúolo, observada tanto em plântulas de dois quanto de quatro dias de germinação, resultou em um menor potencial osmótico nas células promovendo uma redução na elongação celular. O cálcio, por sua vez, pode estar atuando como um modulador da atividade dessa enzima durante o período de germinação (FRIES, 2003).
Em plantas superiores as invertases catalisam a hidrólise irreversível da sacarose em hexoses. Essas enzimas têm papel importante no metabolismo celular, no desenvolvimento do tecido e no crescimento do órgão, por fornecer hexoses que serão utilizadas como fonte de carbono ou como moléculas sinalizadoras. As hexoses produzidas pelas invertases também podem aumentar o potencial osmótico celular (KOCH, 1996; TANG et al., 1999; STURM, 1999; MITSUHASHI et al., 2004). Todavia, a função dessas enzimas no período de germinação e sua participação no desenvolvimento inicial de plântulas ainda é pouco conhecida. Embora a hipoxia exerça importante influência na regulação da relação fonte-dreno entre o endosperma e a plântula, possivelmente pela modulação da expressão de genes relacionados (MITSUHASHI et al., 2004), as invertases parecem não ter uma atuação significativa nessas condições (GUGLIELMINETTI et al., 1995; RICARD et al., 1998; ZENG et al., 1999).
VANTOAI et al. (1995) verificaram que a idade da planta pode interferir na tolerância ao estresse anóxico ou hipóxico. Em seu trabalho, esses autores relataram que plântulas mais novas (dois dias de germinação) foram mais tolerantes à anoxia que plântulas de três dias ou mais e indicaram a operação de mecanismos que permitam a utilização de glicose sob condições anóxicas. Da mesma forma, ANDREWS et al. (1994) afirmaram que plântulas de idades distintas respondiam diferentemente à anoxia. Esses autores observaram que plântulas com até três dias após a embebição induziam a atividade da desidrogenase alcoólica, tanto por hipoxia como por anoxia. Já, plântulas com até cinco dias após a embebição dependiam de um pré-tratamento hipóxico para a indução dessa enzima.
A fim de investigar a influência do cálcio no aumento da tolerância ao alagamento, o presente trabalho teve por objetivo avaliar o crescimento da plântula e o metabolismo de carboidratos, assim como, a atividade de enzimas relacionadas, como a-amilase e invertases, associados à presença do cálcio no período de germinação e/ou alagamento de plântulas de milho com diferentes idades.
2. MATERIAL E MÉTODOS
Para a realização deste trabalho, utilizaram-se plântulas de milho da variedade 'Saracura' (BRS-4154), Ciclo 14, desenvolvida pela Embrapa Milho e Sorgo.
Cariopses de milho foram envolvidas em papel de germinação embebido em água ou solução de cloreto de cálcio (CaCl2) a 0,75% (p/v) e mantidas em câmara de germinação úmida (modelo Mangelsdorss) a 27 ± 2°C no escuro, por dois e quatro dias. Após esses períodos foram realizadas avaliações de massa seca das plântulas, análises bioquímicas e o alagamento. Para as análises bioquímicas, parte aérea, raízes e endospermas foram acondicionados em nitrogênio líquido e armazenados a -20°C.
O alagamento foi realizado em tubos de PVC de 75 mm de diâmetro. Cada tubo continha 100 plântulas e 1,2 L do tampão de alagamento: Tris-hidroximetilaminometano 5 mM, pH 8 com 100 mg.L-1 de ampicilina (SAAB e SACHS, 1996) ou o mesmo tampão acrescido de CaCl2 a 0,75% (p/v). Para tornar o ambiente do tubo hipóxico (3% [v/v] O2 em N2), fez-se o borbulhamento com nitrogênio gasoso por 3 minutos (1 L.min-1). Os tubos foram mantidos no escuro e, após três dias de estresse, avaliou-se a sobrevivência, massa seca e características bioquímicas.
A sobrevivência ao alagamento foi realizada baseando-se no enverdecimento e vigor das plântulas após cinco dias de cultivo em caixas com vermiculita em sala de crescimento (fotoperíodo de 16 horas). A porcentagem da sobrevivência foi calculada em relação a 100% de sobrevivência verificada em plântulas que não foram alagadas (dados não mostrados). No fim desse período avaliou-se também, a massa seca das plântulas.
Os carboidratos solúveis foram extraídos pela homogeneização de 1 g de massa fresca em 4 mL do seguinte tampão: fosfato de potássio 100 mM, pH 7,0, PMSF (Phenylmethyl Sulfonyl Fluoride) 1 mM, DTT (ditiotreitol) 1 mM, ácido ascórbico 20 mM e cloreto de sódio (NaCl) 200 mM, seguido de centrifugação a 9.000 g por 20 minutos a 4 °C e coleta do sobrenadante. O processo foi realizado mais duas vezes e os sobrenadantes combinados.
Para extração do amido, o pellet foi ressuspendido com 8 mL do tampão acetato de potássio 200 mM, pH 4,8 e colocado em banho-maria (100°C) por 5 minutos. Em seguida, foram adicionados 2 mL do preparado da enzima amiloglucosidase, contendo 12,6 unidades, incubando em banho-maria a 40 °C por 2 horas, sob agitação. Após centrifugação a 9.000 g por 20 minutos, o sobrenadante foi coletado e o volume completado para 25 mL. Os açúcares solúveis totais e amido foram quantificados pelo método da Antrona (DISCHE, 1962) e suas concentrações foram expressas em miligramas de glicose por grama de massa fresca (mg.g-1 MF).
A extração da a-amilase foi realizada conforme SKADSEN (1993) com algumas modificações. Um grama de massa fresca de endosperma foi homogeneizado com 4 mL de tampão acetato de sódio 200 mM (pH 5,5), 1 mM de PMSF, 10 mM de CaCl2, 1 mM de DTT e 20 mM de ácido ascórbico, seguido de centrifugação a 9.000 g por 20 minutos a 4 °C. O sobrenadante foi submetido ao banho-maria a 70 °C por 20 minutos, para a inativação da a-amilase e outras enzimas amilolíticas. Após nova centrifugação, o sobrenadante foi utilizado para a incubação em tampão acetato de sódio 200 mM (pH 5,5), 2% de amido solúvel e 5 mM de CaCl2. A incubação ocorreu em banho-maria a 30 °C por 40 minutos. As reações foram paralisadas pela submersão em N2 líquido. Reações a 0 minuto de incubação foram determinadas para eliminar açúcares redutores pré-existentes. Para a quantificação utilizou-se o método do DNS para açúcares redutores (MILLER, 1959). A atividade das enzimas foi obtida após 40 minutos de incubação e abatimento dos valores do tempo zero e expressa em µmol de glicose por grama de matéria fresca por hora (µmol gli.g-1 MF.h-1).
A extração e incubação das invertases solúveis foram realizadas conforme descrito por ZENG et al. (1999) e da invertase ácida da parede celular (IAPC) segundo CAZETTA et al. (1999), com algumas modificações.
A invertase neutra do citosol (INC) e invertase ácida do vacúolo (IAV) foram extraídas pela homogeneização de 1 g de massa fresca em 4 mL do tampão de extração contendo 100 mM de HEPES (pH 7,5), 1 mM de PMSF, 5 mM de MgCl2, 1 mM de DTT e 20 mM de ácido ascórbico, seguida pela centrifugação a 9.000 g por 20 minutos a 4°C. O sobrenadante foi coletado para as análises das invertases solúveis e o pellet ressuspendido e homogeneizado durante 7 minutos com 4 mL de tampão citrato 200 mM (pH 4,8), 1 mM de PMSF, 5 mM de MgCl2, 1 mM de DTT, 20 mM de ácido ascórbico e 1 M de NaCl para a extração da IAPC, seguido de centrifugação a 9.000 g por 20 minutos a 4°C, coletando-se o sobrenadante para a análise da enzima.
A atividade das enzimas foi realizada em tampão fosfato de potássio 100 mM (pH 7,5) para a INC e em tampão citrato de sódio 200 mM (pH 4,8) para a IAV e IAPC, acrescidos de 5 mM de MgCl2 e 200 mM de sacarose. A incubação foi realizada em banho-maria a 30 °C por 40 minutos. A paralisação da reação e quantificação foram feitas da mesma forma que da a-amilase. A atividade da enzima foi expressa em µmol de glicose por grama de matéria fresca por hora (µmol gli.g-1 MF.h-1).
3. RESULTADOS E DISCUSSÃO
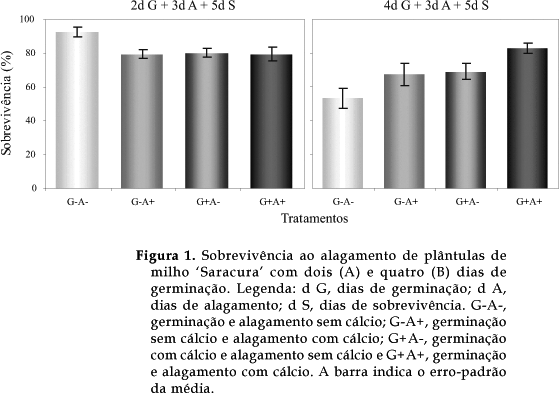
Nas plântulas de dois dias de idade, a presença do cálcio no período de germinação e/ou alagamento não teve efeito benéfico no aumento da sobrevivência como ocorreu nas plântulas de quatro dias (Figura 1). Nestas últimas, observou-se um aumento gradativo na sobrevivência devido ao contato com o cálcio, sendo mais expressivo quando sua presença ocorreu simultaneamente na germinação e no alagamento (G+A+). Neste caso, verificou-se um aumento de 57% na sobrevivência em relação àquela observada na ausência total de cálcio (G-A-).
Esses resultados reafirmam a influência do cálcio na tolerância à hipoxia de plântulas de quatro dias, observada por ALVES et al. (2002) e mostram pela primeira vez, que plântulas de milho 'Saracura' com dois dias de idade são tolerantes a este tipo de estresse, independentemente da presença do cálcio. Maior tolerância em plântulas de milho mais jovens foi observada por VAN TOAI et al. (1995), ao verificar um declínio na sobrevivência de plântulas com três dias ou mais, após 48 horas de anoxia.
Estudando o comportamento de plântulas de milho 'Saracura' de quatro dias de idade à hipoxia, VITORINO et al. (2001) relacionaram a diminuição da sobrevivência ao alagamento à intensa lise celular ocorrida na região do mesocótilo, a qual tornava-se translúcida e flácida com o prolongamento do estresse, formando uma forte constrição, responsável pelo tombamento e morte das plântulas. No presente trabalho, esse sintoma não foi observado em plântulas com dois dias de idade, independentemente da presença do cálcio. Em plântulas com quatro dias, o cálcio no papel de germinação ou no tampão de alagamento (G+ A- e G- A+) contribuiu para a atenuação desse sintoma, o qual não se manifestou quando esse elemento esteve presente em ambas as situações (G+ A+).
De maneira geral, nas plântulas após o período de alagamento havia uma massa seca de parte aérea e raízes menores que aquelas observadas após a germinação (Figura 2). Entretanto, essa perda não prejudicou o ganho de massa seca pós-estresse. Observou-se que as plântulas mais jovens sobreviventes ao estresse experimentaram em média um ganho de massa seca de parte aérea de oito vezes, tomando como base a massa observada no fim do alagamento, enquanto o ganho nas mais velhas foi de três vezes. Ao mesmo tempo, nas raízes, esse ganho alcançou em média sete e duas vezes em plântulas de dois e quatro dias respectivamente.
O cálcio, por sua vez, não afetou o acúmulo de massa seca pós-hipoxia na parte aérea de plântulas com dois dias de germinação, submetidas ao alagamento (Figura 2A). Por outro lado, em plântulas de quatro dias (Figura 2B), observou-se, após o período de sobrevivência, aumento médio em massa seca de 87% na ausência total do cálcio (G- A-) ou quando esse esteve presente somente no alagamento (G- A+). Quando, porém, o cálcio foi fornecido apenas na germinação (G+ A-), ou em ambas as situações (G+ A+), esse aumento foi de 284%.
Nas raízes, a presença do cálcio promoveu aumentos expressivos no acúmulo de massa seca durante a sobrevivência, principalmente em plântulas que foram alagadas com quatro dias de idade (Figura 2C e 2D). Enquanto nas plântulas mais jovens, esses aumentos foram na ordem de 475% na ausência total de cálcio (G-A-) até 938% em sua total presença (G+A+), nas plântulas de quatro dias verificaram-se índices muito inferiores. Apesar de essas plântulas serem maiores no fim do período de alagamento, elas passaram mais tempo se recuperando dos danos causados por esse estresse do que produzindo massa. Dessa forma, observou-se apenas a recuperação da massa seca perdida no período de estresse, quando o cálcio não esteve presente (G- A-) ou quando foi fornecido somente no alagamento (G- A+), sendo um pouco maior no segundo caso, embora não significativo. Quando somente a germinação ocorreu na presença desse elemento (G+ A-) ou quando o cálcio esteve presente em ambas as situações (G+ A+), houve aumentos de 70% e112%, respectivamente, na massa seca de raiz no período de sobrevivência.
A presença do cálcio no tampão de alagamento (G- A+; G+ A+) promoveu maior acúmulo de massa seca nas raízes de plântulas de dois dias (Figura 2C) durante a sobrevivência, quando relacionada aos seus respectivos tratamentos alagados sem cálcio (G- A-; G+ A-). Para as plântulas de quatro dias houve diferença significativa somente entre aquelas que germinaram na presença do cálcio (G+A-; G+A+). Esse maior acúmulo de massa seca pode ser resultado do aparecimento de maior número de raízes adventícias, observado durante o experimento, cuja formação pode estar relacionada à presença do cálcio no período de alagamento. De maneira geral, para diversas culturas, o cálcio induz a formação desse tipo de raízes quando comparadas com aquelas crescidas em solos deficientes nesse elemento (Marschner, 1986).
Esses resultados evidenciam que a presença do cálcio em plântulas de quatro dias, principalmente no período de germinação, promove uma recuperação mais rápida dos danos causados no período de estresse hipóxico, permitindo maior utilização de compostos carbonados no crescimento da plântula. A rápida recuperação e posterior crescimento ocorrem em plântulas de dois dias de germinação, independente mente da presença do cálcio.
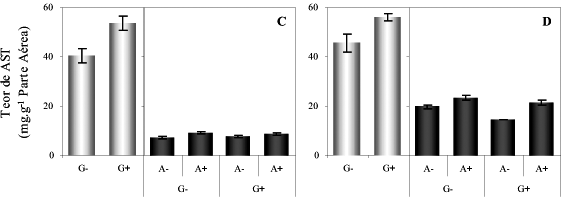
De maneira geral, no fim do período de alagamento, os teores de amido nos endospermas de cariopses foram semelhantes entre si (Figura 3A e 3B). No fim da germinação, observaram-se maiores teores de amido nas plântulas de dois dias do que naquelas de quatro dias, em vista de menor atividade da a-amilase. Pode se inferir, portanto, que, no período de alagamento, ocorreu maior mobilização dessas reservas nas plântulas mais jovens. Apesar disso, a atividade da a-amilase em ambas as plântulas não aumentou, chegando até mesmo a decrescer, não significativamente, quando o cálcio esteve presente (Figura 3C e 3D).
A redução nos teores de amido observada após o alagamento nos endospermas de cariopses de dois dias de germinação, relacionada à baixa atividade da a-amilase, pode ser devido à atividade de outras enzimas amilolíticas como a a-amilase e/ou fosforilase do amido. Essa baixa atividade pode estar relacionada à inibição da síntese de a-amilase durante o estresse, o que foi observado em trigo e cevada sob condições de anoxia (PERATA et al., 1993).
Apesar de não haver diferenças significativas entre os teores de amido, os teores de açúcares solúveis totais, após o período de estresse, foram maiores nos endospermas de cariopses de plântulas de quatro dias de germinação quando comparados aos teores naqueles de plântulas de dois dias (Figura 4A e 4B). Analisando-se esses resultados, supõe-se que, durante o alagamento, plântulas de dois dias estão translocando maior quantidade de reservas de carboidratos que plântulas de quatro dias. Desse modo, o fator de tolerância existente em plântulas de dois dias pode estar relacionado ao transporte de compostos carbonados do endosperma para a plântula, provindos, principalmente da hidrólise do amido.
Em geral, os teores de açúcares solúveis totais, na parte aérea (Figura 4C e 4D) e raiz (Figura 4E e 4F) após o alagamento, foram maiores nas plântulas de quatro dias que de dois dias de idade. Uma vez que, no fim da germinação, o cálcio promoveu um acúmulo desses açúcares na plântula, culminando em menor utilização e redução no crescimento das plântulas (FRIES, 2003), a presença do cálcio no período de alagamento, de maneira geral, não influenciou nos teores desses carboidratos. Assim, se em plântulas de dois dias de germinação está havendo maior translocação de reservas do endosperma para a plântula e não está ocorrendo acúmulo de açúcares na parte aérea e raiz durante o alagamento, essas plântulas, além de seus mecanismos de transporte mais eficientes, conseguem utilizar melhor as reservas para a sua manutenção durante a hipoxia, por isso, ao saírem do período de estresse, recuperam-se mais rapidamente. O aperfeiçoamento na tolerância de plântulas de dois dias, após a adição de glicose, foi observado por VAN TOAI et al. (1995), que sugeriram que tais resultados podem ser interpretados como uma indicação da operação de mecanismos que permitam a utilização de glicose sob condições anóxicas.
O alagamento reduziu a atividade de todas as invertases (Figura 5 e 6), independentemente da presença do cálcio. As invertases neutra do citosol (INC) (Figuras 5A, 5B, 6A e 6B) e ácida da parede celular (IAPC) (dados não mostrados) tiveram reduções menores em sua atividade, enquanto a invertase ácida do vacúolo (IAV) (Figuras 5C, 5D, 6C e 6D) foi a mais fortemente afetada. Em plântulas de dois dias de germinação, a atividade dessa enzima caiu em média 90% na parte aérea e 96% na raiz. Já em plântulas de quatro dias essas reduções chegaram a 98 e 96% respectivamente.
Apesar de a função das invertases no período de germinação ainda ser pouco conhecida, sabe-se que elas clivam sacarose para fornecer hexoses como fonte de energia e carbono, regulando o turgor das células e promovendo a expansão celular, além de gerar um gradiente de concentração de sacarose entre órgãos fonte e dreno (TANG et al., 1999; STURM, 1999; MITSUHASHI et al., 2004; TAIZ e ZEIGER, 2004). Durante a fase de crescimento de órgãos ou células, a invertase ácida do vacúolo é a enzima com maior importância no suprimento desses açúcares (KOCH, 1996; TANG et al., 1999; MITSUHASHI et al., 2004). Os resultados observados com o milho 'Saracura' por FRIES (2003) corroboram com esses autores quando, tanto na parte aérea quanto em raiz, a invertase ácida do vacúolo teve as mais altas atividades. Ao mesmo tempo, a presença do cálcio reduziu a atividade dessa enzima durante a germinação e, conseqüentemente, a diminuição do efeito osmótico exercido pela clivagem da sacarose promoveu menor expansão celular, reduzindo o crescimento da plântula. Esses resultados sugerem, portanto, que existe uma modulação exercida pelo cálcio sobre essa enzima durante esse período. No presente trabalho, o alagamento reduziu fortemente a atividade dessa enzima, tanto em plântulas de dois como de quatro dias de germinação. Em plântulas mais jovens, nas quais se observou alto grau de tolerância, a invertase neutra do citosol (Figuras 5A e 6A) que foi pouco afetada pelo alagamento, principalmente na parte aérea, poderia estar atuando na manutenção metabólica da plântula durante o estresse. Por outro lado, como o cálcio não teve influência significativa na atividade das invertases no período de estresse, pode ser comprovado que não participa dos processos comandados por essas enzimas.
A redução na atividade das invertases no período de alagamento promove menor concentração de hexoses disponíveis para o metabolismo celular, diminuindo, portanto, a respiração anaeróbica e, conseqüentemente, a produção de substâncias tóxicas. Assim, essas plântulas conseguem manter uma sobrevida por um período maior em condições de hipoxia. Por outro lado, novas investigações em relação aos mecanismos de atuação do cálcio no aumento da tolerância ao estresse hipóxico são necessárias.
Ao analisar plântulas de milho de cinco a sete dias de germinação, submetidas a tratamentos com baixo teor de oxigênio, ZENG et al. (1999) observaram uma diminuição brusca na atividade da invertase ácida do vacúolo nas primeiras horas de anoxia ou hipoxia, estabilizando em seguida para níveis mais baixos que sob condições aeróbicas.
O mesmo comportamento ocorreu com a sintase da sacarose, porém após um rápido declínio, essa enzima recuperou sua atividade alcançando níveis próximos ou superiores aos encontrados em aerobiose. Segundo esses autores, a diminuição na atividade da sintase da sacarose pode contribuir no ajustamento das plântulas às condições de baixo oxigênio. Esses resultados confirmam a importância da sintase da sacarose durante a hipoxia de plântulas de milho relatada por RICARD et al. (1998). Ao mesmo tempo, GUGLIELMINETTI et al. (1995) observaram que a razão de atividade entre a sintase da sacarose e a invertase neutra do citosol, em coleóptilos de arroz de oito dias de idade, foi de 5:1 sob condições aeróbicas e 30:1 sob anoxia. Esse dado revela que, em ambas as condições, as duas enzimas estão atuando, porém, sob anoxia a sintase da sacarose tem maior função no compartimento citosólico. Em milho, a preferência pela via da sintase da sacarose foi confirmada pelos menores índices na atividade da invertase ácida do vacúolo (ZENG et al., 1999). Nesse trabalho, o genótipo com a menor atividade da invertase juntamente com a menor capacidade relativa de clivagem da sacarose (atividade da invertase mais atividade da sintase da sacarose) obteve o maior índice de sobrevivência.
4. CONCLUSÕES
1. Plântulas de dois dias são tolerantes à hipoxia "per si". Tal tolerância está relacionada a mecanismos de utilização de carboidratos durante a hipoxia.
2. A presença de cálcio exógeno aumenta a tolerância de plântulas de quatro dias ao estresse hipóxico e promove uma recuperação mais rápida dos danos causados nesse período.
3. A atividade da invertase ácida do vacúolo é fortemente reduzida no período de alagamento e o cálcio não tem qualquer influência sobres essa enzima durante esse período.
Recebido para publicação em 27 de dezembro de 2005 e aceito em 17 de agosto de 2006.
- ALVES, J. D.; MAGALHÃES, M. M.; GOULART, P. F. P.; DANTAS, B. F.; GOUVÊA, J. A.; PURCINO, R. P.; MAGALHÃES, P. C.; FRIES, D. D.; LIVRAMENTO, D. E.; MEYER, L. E.; SEIFERT, M.; SILVEIRA, T. Mecanismos de tolerância da variedade de milho 'Saracura' (BRS-4154) ao alagamento. Revista Brasileira de Milho e Sorgo, Sete Lagoas, v.1, p. 41-52, 2002.
- ALVES, J. D.; MAGALHÃES, M. M.; OLIVEIRA, L. E. M.; VITORINO, P. G. Mecanismo de tolerância de plantas ao alagamento. Universa, Brasília, v. 8, p. 221-242, 2000.
- ANDREWS, D. L.; DREW, M. C.; JOHNSON, J. R.; COBB, B. G. The response of maize seedlings of different ages to hypoxic and anoxic stress. Plant Physiology, Rockville, v. 105, n. 1, p. 53-60, May 1994.
- BRAY, E. A.; BAILEY-SERES, J.; WERETILNYK, E. Responses to abiotic stresses. In: BUCHANAN, B. B.; GRUISSEM, W.; JONES, R. L. Biochemistry & Molecular Biology of Plants. Rockville: American Society of Plant physiologists, 2000. Cap. 22, p. 1158-1249.
- CAZETTA, J. O.; SEEBAUER, J. R.; BELOW, F. E. Sucrose and nitrogen supplies regulate growth of maize kernels. Annals of Botany, London, v. 84, n. 6, p. 747-754, Dec. 1999.
- DISCHE, Z. General color reactions. In: WHISTLER, R. L.; WOLFRAM, M. L. Carbohydrate chemistry. New York: Academic Press, 1962. p. 477-520.
- FRIES, D. D. Comportamento de a-amilase/invertases e mudanças anatômicas associadas ao cálcio exógeno no período de germinação e/ou alagamento de plântulas do milho (Zea Mays L.) 'Saracura' BRS-4154. 2003. 49 f. Dissertação (Mestrado em Agronomia/Fisiologia Vegetal) Universidade Federal de Lavras, Lavras, MG.
- GUGLIELMINETTI, L.; PERATA, P.; ALPI, A. Effect of anoxia on carbohydrate metabolism in rice seedlings. Plant Physiology, Rockville, v. 108, n. 2, p. 735-741, June 1995.
- HE, C.; DREW, M. C.; MORGAN, P. W. Induction of enzimes associated with lysigenous aerenchyma formation in roots of Zea mays L. during hipoxia or nitrogen starvation. Plant Physiology, Rockville, v. 105, n. 3, p. 861-865, July 1994.
- KOCH, K.E. Carbohydrate-modulated gene expression in plants. Annual Review of Plant Physiology and Plant Molecular Biology, Palo Alto, v. 47, p. 509-540, 1996.
- KOLB, R. M.; MEDRI, M. E.; BIANCHINI, E.; PIMENTA, J. A.; GILONI, P. C.; CORREA, G. T. Anatomia Ecológica de Sebastiania commersoniana (Baillon) Smith & Downs (Euphorbiaceae) submetida ao alagamento. Revista Brasileira de Botânica, São Paulo, v. 21, n. 3, p. 261-269, dez. 1998.
- MARSCHNER, H. Mineral Nutrition of Higher Plants. London: Academic Press, 1986. 889 p.
- MILLER, G. L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical Chemistry, Washington, v. 31, n. 3, p. 426-428, Mar. 1959.
- MITSUHASHI, W.; SAZAKI, S.; KANAZAWA, A.; YANG, Y-Y.; KAMIYA, Y.; TOYOMASU, T. Differential expression of acid invertase genes during seed germination in Arabidopsis thaliana Bioscience, Biotechnology and Biochemistry, Tokyo, v. 68, p. 602-608, 2004.
- PARENTONI, S. N.; GAMA, E. E.; LOPES, M. A.; SANTOS, M. X.; GUIMARÃES, P. E. O.; PACHECO, C. A.; SOUZA, I. R. P.; MEIRELES, W.; CORREA, L. A. Seleção para tolerância ao encharcamento na variedade de milho CMS54-Saracura. In: REUNION LATINOAMERICANA, 4.; REUNION DELA ZONA ANDINA DE INVESTIGADORES EM MAIZ, 17., 1997, Cerete Y Cartagena de Ludias, Colômbia. Memória... S.L.: CORPOICA / CIMMYT, 1997. p.368-373.
- PERATA, P.; GESHI, N.; YAMAGUCHI, J.; AKAZAWA, T. Effect os anoxia on the induction of á-amilase in cereal seeds. Planta, Berlin, v. 191, n. 3, p. 402-408, Aug. 1993.
- RICARD, B.; VANTOAI, T.; CHOUREY, P.; SAGLIO, P. Evidence for the critical role of sucrose synthase for anoxic tolerance of maize roots using a double mutant. Plant Physiology, Rockville, v. 116, n. 4, p. 1323-1331, 1998.
- SAAB, I. N.; SACHS, M. M. A flooding-induced xyloglucan endo-transglycosylase homolog in maize is responsive to ethylene and associated with aerenchyma. Plant Physiology, Rockville, v. 112, n. 1, p. 385-391, Sept. 1996.
- SKADSEN, R. W. Aleurones from a barley with low á-amylase activity become highly responsive to gibberellin when detached from the starchy endosperm. Plant Physiology, Rockville, v. 102, n. 1, p. 195-203, May 1993.
- SUBBAIAH, C. C.; ZHANG, J.; SACHS, M. M. Involvement of intracellular calcium in anaerobic gene expression and survival of maize seedlings. Plant Physiology, Rockville, v. 105, n. 1, 369-376, May 1994.
- STURM, A. Invertases. Primary structures, functions, and roles in plant development and sucrose partitioning. Plant Physiology, Rockville, v. 121, p. 1-7, 1999.
- TANG, G-Q.; LÜSCHER, M.; STURM, A. Antisense repression of vacuolar and cell wall invertase in transgenic carrot alters early plant development and sucrose partitioning. The Plant Cell, Rockville, v. 11, p. 177-189, 1999.
- TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 3 ed. Porto Alegre: Artmed, 2004. 719 p.
- VANTOAI, T. T.; SAGLIO, P.; RICARD, B.; PRADET, A. Developmental regulation of anoxic stress tolerance in maize. Plant, Cell and Environment, Oxford, v. 18, n. 8, p. 937-942, Aug. 1995.
- VITORINO, P. G.; ALVES, J. D.; MAGALHÃES, P. C.; MAGALHÃES, M. M.; LIMA, L. C. O.; OLIVEIRA, L. E. M. Flooding tolerance and cell wall alterations in maize mesocotyl during hypoxia. Pesquisa Agropecuária Brasileira, Brasília, v. 36, n. 8, p. 1027-1035, ago. 2001.
- ZENG, Y.; WU, Y.; AVIGNE, W. T.; KOCH, K. E. Rapid repression of maize invertases by low oxygen. Invertase/sucrose synthase balance, sugar signaling potential, and seedling survival. Plant Physiology, Rockville, v. 121, n. 2, p. 599-608, Oct. 1999.
Datas de Publicação
-
Publicação nesta coleção
12 Abr 2007 -
Data do Fascículo
2007
Histórico
-
Aceito
17 Ago 2006 -
Recebido
27 Dez 2005