Resumos
Os lagartos teiús possuem uma dieta generalista, podendo agir como importantes dispersores de sementes em florestas semidecíduas do sudeste do Brasil. Foram estudadas a frugivoria e a dispersão de sementes de lagartos teiús usando animais em cativeiro, através da oferta de frutos de uma floresta semidecídua. Frutos de trinta espécies vegetais foram oferecidos aos lagartos em cativeiro, com diâmetro variando de 0,81 a 10,0 cm. Não foram encontradas diferenças estatísticas na germinação entre as sementes que passaram pelo trato digestivo do lagarto e as controle de Eugenia uniflora (chi²= 0.69, P>0.50), Genipa americana (chi²= 6.4, P>0.975), Cereus peruvianus (chi²= 0.018, P>0.10), e Solanum viarum (chi²= 6.23, P>0.975). O tempo de retenção da semente no tubo digestivo do teiú variou de 22 a 23 h para Solanum lycocarpum e 43 a 44 h para Syagrus romanzoffiana. Nossos resultados indicam que o lagarto teiú tem potencial para agir como um importante dispersor de sementes nos trópicos.
Brasil; Floresta Atlântica; frugivoria; frutos; lagarto; dispersão de sementes; tempo de retenção da semente; Tupinambis
Tegu lizards have a generalist diet and may play an important role as seed dispersers in semideciduous forests in south-east Brazil. We studied the frugivory and seed dispersal of tegu lizards using captive animals and offering wild fruits from a semideciduous forest. Thirty fruit species were eaten by the lizards in captivity, ranging from 0.81 to 10.0 cm (fruit diameter). Even large fruit adapted to dispersal by large mammals were swallowed (ex. Syagrus oleracea). There were no statistical differences in seed germination between seeds that passed through the lizard gut and the control in Eugenia uniflora (chi2 = 0.69, P>0.50), Genipa americana (chi2 = 6.4, P>0.975), Cereus peruvianus (chi2 = 0.018, P>0.10), and Solanum viarum (chi2 = 6.23, P>0.975). Seed retention time in the tegu gut ranged from 2224 h (Solanum lycocarpum) to 4344 h (for Syagrus romanzoffiana). Our results indicate that tegu lizards have a potential to be an important seed dispersers in the Neotropics.
Brazil; Atlantic forest; frugivory; fruits; lizard; seed dispersal; seed retention time; Tupinambis
Frugivoria e dispersão de sementes pelo lagarto teiú Tupinambis merianae (Reptilia: Teiidae)* * O resumo do trabalho foi apresentado no III Congresso Internacional de Frugivoria e Dispersão de Sementes realizado na cidade de São Pedro, SP, Brasil, no ano de 2000.
Frugivory and seed dispersal by the tegu lizard Tupinambis merianae Reptilia: Teiidae
Everaldo Rodrigo de CastroI, III; Mauro GalettiII
IGrupo de Fenologia e Dispersão de Sementes, Universidade Estadual Paulista, Departamento de Ecologia, Caixa Postal 199, 13506-900 Rio Claro, SP, Brasil
IIGrupo de Fenologia e Dispersão de Sementes, Universidade Estadual Paulista, Departamento de Ecologia, Caixa Postal 199, 13506-900 Rio Claro, SP, Brasil. mgaletti@rc.unesp.br
IIIAutor para correspondência: ercastro@rc.unesp.br
RESUMO
Os lagartos teiús possuem uma dieta generalista, podendo agir como importantes dispersores de sementes em florestas semidecíduas do sudeste do Brasil. Foram estudadas a frugivoria e a dispersão de sementes de lagartos teiús usando animais em cativeiro, através da oferta de frutos de uma floresta semidecídua. Frutos de trinta espécies vegetais foram oferecidos aos lagartos em cativeiro, com diâmetro variando de 0,81 a 10,0 cm. Não foram encontradas diferenças estatísticas na germinação entre as sementes que passaram pelo trato digestivo do lagarto e as controle de Eugenia uniflora (c²= 0.69, P>0.50), Genipa americana (c²= 6.4, P>0.975), Cereus peruvianus (c²= 0.018, P>0.10), e Solanum viarum (c²= 6.23, P>0.975). O tempo de retenção da semente no tubo digestivo do teiú variou de 22 a 23 h para Solanum lycocarpum e 43 a 44 h para Syagrus romanzoffiana. Nossos resultados indicam que o lagarto teiú tem potencial para agir como um importante dispersor de sementes nos trópicos.
Palavras-chave: Brasil, Floresta Atlântica, frugivoria, frutos, lagarto, dispersão de sementes, tempo de retenção da semente, Tupinambis.
ABSTRACT
Tegu lizards have a generalist diet and may play an important role as seed dispersers in semideciduous forests in south-east Brazil. We studied the frugivory and seed dispersal of tegu lizards using captive animals and offering wild fruits from a semideciduous forest. Thirty fruit species were eaten by the lizards in captivity, ranging from 0.81 to 10.0 cm (fruit diameter). Even large fruit adapted to dispersal by large mammals were swallowed (ex. Syagrus oleracea). There were no statistical differences in seed germination between seeds that passed through the lizard gut and the control in Eugenia uniflora (c2 = 0.69, P>0.50), Genipa americana (c2 = 6.4, P>0.975), Cereus peruvianus (c2 = 0.018, P>0.10), and Solanum viarum (c2 = 6.23, P>0.975). Seed retention time in the tegu gut ranged from 2224 h (Solanum lycocarpum) to 4344 h (for Syagrus romanzoffiana). Our results indicate that tegu lizards have a potential to be an important seed dispersers in the Neotropics.
Keywords: Brazil, Atlantic forest, frugivory, fruits, lizard, seed dispersal, seed retention time, Tupinambis.
INTRODUÇÃO
Os répteis foram, juntamente com os peixes, os primeiros vertebrados a interagirem com plantas que possuem sementes (Tiffney 1984). Assim, há 200 milhões de anos, os progenitores das atuais Cycas deixaram fósseis com sementes de frutos aparentemente adaptados para o consumo por répteis primitivos (Sporne 1965). Os frutos são parte da dieta de muitas aves, mamíferos e répteis tais como tartarugas e lagartos (Iverson 1985; Jordano 2000). As tartatugas e os lagartos consomem uma variedade de espécies de frutos, além de folhas, botões e flores (Rick & Bowman 1961; Rand 1978; Williams et al. 1993; Figueira et al. 1994; Valido & Nogales 1994; Saez & Traveset 1995; Whiting & Greef 1997; Vitt et al. 1997; Nogales et al. 1998; Lord & Marshall 2001; Cooper & Vitt 2002; Cooper et al. 2002).
Os lagartos são talvez os mais conhecidos répteis frugívoros. Evidências estão sendo lentamente acumuladas de que eles podem ser importantes mutualistas em muitas regiões tropicais e temperadas (Iverson 1985; Whitaker 1987; Losos & Greene 1988; Traveset 1990, 1995; Valido & Nogales 1994; Moll & Jansen 1995; Castilla 2000; Olesen & Valido 2003). Estudos sobre a dieta de lagartos têm aumentado nos últimos quinze anos, fornecendo dados importantes da história natural de muitas espécies (Sylber 1988; Willson et al. 1996; Vitt et al. 1997; Cooper & Vitt 2002). Estes estudos têm também fornecido informações sobre partilha de recurso, relação entre modo de forrageamento, predação e táticas reprodutivas (Pianka 1980; Huey & Pianka 1981; Durtsche 1995, Vitt et al. 1999).
Entre os lagartos, muitas espécies se alimentam de frutos, destacando-se as famílias Gekkonidae, Scincidae, Iguanidae, Lacertidae, Varanidae e Teiidae (Whitaker 1987; Traveset 1990; Valido & Nogales 1994; Castilla 2000; Cooper & Vitt 2002). Na família Teiidae, existem relatos do consumo de frutos por Ameiva, Cnemidophorus, Kentropyx (Vitt et al. 1997, 2001; Cooper et al. 2002) e principalmente Tupinambis (Donadio & Gallardo 1984; Bolkovic et al. 1989; Sazima & Haddad 1992; Williams et al. 1993; Mercolli & Yanosky 1994; Reis 1995). Segundo Bolkovic et al. (1989), Tupinambis rufescens também pode ser dispersor de sementes de espécies vegetais locais, devido ao grande número de sementes e frutos encontrados no sistema digestivo deste animal (Williams et al. 1993).
O gênero Tupinambis se distribui por quase toda a América do Sul, habita a Bacia Amazônica, as áreas costeiras das Guianas, Venezuela, Colômbia e Norte do Brasil e áreas adjacentes, ao Sul do Paraguai e Uruguai e Norte da Argentina (Presch 1973). As espécies desse gênero são caçadas tradicionalmente por alguns povos indígenas para subsistência, além de serem importantes fontes de couro e carne em alguns países como a Argentina (Fitzgerald et al. 1991). Tupinambis merianae ocorre na maioria das florestas da região sudeste do Brasil, possuindo uma dieta onívora que pode incluir invertebrados, vertebrados, ovos e várias espécies de frutos (Presch 1973; Sazima & Haddad 1992), podendo atuar como dispersor de sementes em pequenos fragmentos florestais.
Diante disso, os objetivos do trabalho foram responder às seguintes perguntas: quais as espécies de frutos são consumidas pelo Teiú Tupinambis merianae? Qual o tempo de retenção das sementes no tubo digestivo do animal? Qual o estado das sementes após terem passado pelo sistema digestivo? A passagem das sementes pelo sistema digestivo do teiú altera a capacidade de germinação?
MATERIAL E MÉTODOS
O estudo foi realizado no campus da Universidade Estadual Paulista (Unesp), Rio Claro, São Paulo de dezembro de 1998 a novembro de 1999. Quatro recintos escolhidos ao acaso e com um número de teiús variando de 3-5 por recinto foram utilizados. No cativeiro, os teiús eram alimentados quase que diariamente com uma dieta de carne. Antes dos frutos serem oferecidos aos teiús, eles eram trazidos para o laboratório de Fisiologia Vegetal da Unesp - Rio Claro, onde foram tomadas medidas de comprimento, diâmetro dos frutos e sementes e peso fresco.
A coleta dos frutos foi realizada prioritariamente na Mata São José, onde ocorrem teiús, mas também em alguns pomares da região. A Mata São José está localizada entre as cidades de Rio Claro e Araras, em área de 230 ha, com 630 metros de altitude, (47º28'W e 22º22'S). A vegetação arbórea da área é densa, com dossel variando de 15 a 30 metros de altura em diferentes locais. O clima da região segundo Köeppen (1948) é do tipo "Cwa", com temperatura do mês mais frio variando entre 3-18ºC (Pagano & Leitão Filho 1987). A mata apresenta-se bastante alterada pelo homem, com indícios de desmatamento e caça, sendo que alguns grandes frugívoros como tucanos, antas e bugios já não mais ocorrem na área (C. São Bernardo, dados não publicados).
Experimento 1 - Teve como finalidade verificar se a passagem das sementes pelo sistema digestivo do teiú altera a capacidade de germinação das mesmas. Após o oferecimento dos frutos nos recintos, as fezes dos teiús contendo as sementes foram coletadas e lavadas em água corrente dentro de peneiras com malha de 0,5 x 0,5 mm. Estas sementes passadas pelo sistema digestivo dos animais (tratamento) foram submetidas a testes de germinação, junto com sementes que não passaram pelo sistema digestivo dos animais (controle), na Estufa de Germinação do Laboratório de Fisiologia Vegetal. Para avaliar diferenças na quantidade final das sementes germinadas entre tratamento e controle foi utilizado o teste de qui-quadrado.
Experimento 2 - Teve como finalidade verificar o tempo de retenção das sementes no tubo digestivo do teiú e seu estado após sua eliminação. Quatro teiús foram selecionados para este experimento, um de cada recinto. Foram oferecidos a eles frutos marcados com pequenas tiras de plástico coloridas com a data de oferecimento, ocultas na alimentação (Milton 1981). No momento do oferecimento dos frutos marcados, realizado na primavera e verão (estação de maior atividade dos teiús), os animais foram observados e a hora da ingestão dos frutos anotada. Após a ingestão dos frutos marcados, foram feitas observações a cada 2 horas no recinto à procura das tiras de plástico nas fezes.
RESULTADOS
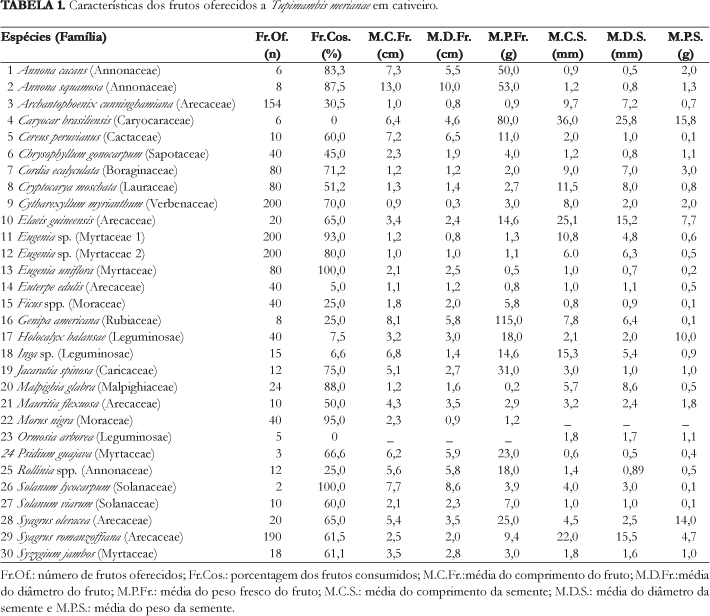
Os frutos com maior taxa de consumo pelos teiús foram: Solanum lycocarpum, Eugenia uniflora, Eugenia sp. e Morus nigra. O tamanho dos frutos consumidos variou de 0,81 a 10,0 cm de diâmetro (Tab. 1). Os frutos pequenos, como os de Eugenia uniflora e Syagrus romanzoffiana, quando oferecidos, foram engolidos por inteiro, enquanto que os de S. oleracea, que possuem uma semente com maior diâmetro, foram só despolpados mas não engolidos. Os grandes frutos, como os de Solanum lycocarpum e Annona cacans foram comidos aos pedaços.
Das espécies vegetais oferecidas ao teiú, duas não foram ingeridas, os frutos de Caryocar brasiliensis (Caryocaraceae) e as sementes de Ormosia arborea (Leguminosae) (Tab. 1). Das que foram ingeridas, praticamente todas as sementes foram encontradas ilesas nas fezes. Para a realização dos testes de germinação, não foi encontrado para alguns frutos um número suficiente de sementes no recinto, talvez devido à perda das sementes causada por pisoteamento pelos teiús, pois o substrato do recinto era composto por grama, dificultando o encontro das sementes. Das sete espécies usadas nos testes de germinação (Eugenia uniflora, Genipa americana, Solanum viarum, Cereus peruvianus, Cordia ecalyculata, Annona squamosa e Malpighia glabra), apenas quatro germinaram (Eugenia uniflora, Genipa americana, Solanum viarum, Cereus peruvianus) e as outras três foram atacadas por fungos (Fig. 1).
No primeiro experimento, não foram encontradas diferenças significativas na quantidade final de sementes germinadas entre tratamento e controle para as seguintes espécies Eugenia uniflora (c2 = 0.69, g.l. = 1, P>0.50), Genipa americana (c2 = 6.4, P>0.975), Cereus peruvianus (c2 = 0.018, P>0,10) e Solanum viarum (c2 = 6.23, P>0.975) (Fig. 1). Em relação à velocidade de germinação, somente as sementes da espécie Genipa americana tiveram um aumento quando passaram pelo sistema digestivo dos teiús. Cerca de 60% das sementes submetidas ao tratamento germinaram em apenas uma semana e 80% delas germinaram em duas semanas (Fig. 2B).
No segundo experimento, foram utilizadas duas espécies de frutos, uma com sementes de diâmetro grande, como o gerivá Syagrus romanzoffiana (Arecaceae) de 15,5 mm e outra com sementes de diâmetro menor como a lobeira Solanum lycocarpum (Solanaceae) de 3 mm. As sementes do gerivá demoraram mais para percorrer o sistema digestivo do teiú, com uma média de 43 a 44 horas. As sementes menores de Solanum lycocarpum, levaram um tempo menor para percorrer o tubo digestivo do animal, com uma média de 22 a 24 horas.
DISCUSSÃO
O lagarto Tupinambis merianae, apesar de possuir uma dieta generalista, poderia atuar como dispersor da maioria das espécies de frutos oferecidos a eles em cativeiro. O lagarto consumiu frutos que apresentavam uma grande variação de tamanho. Em áreas fragmentadas como a Mata São José, onde a maioria dos grandes frugívoros como aves e mamíferos está reduzida, os teiús provavelmente poderiam atuar como dispersores, já que se deslocam por grandes áreas à procura de alimento durante o seu forrageamento (M. Galetti observação pessoal).
Williams et al. (1993) encontraram no tubo digestivo de Tupinambis rufescens frutos de Celtis spinosa, de leguminosas do gênero Prosopis, especialmente P. torquata, de Caparidáceas, em sua maioria Capparis salicifolia, C. tweediana, C. speciosa, C. retusa, e de Zizyphus mistyol. Já Donadio & Gallardo (1984), observaram no sistema digestivo de T. rufescens frutos de Condalia microphylla, Zizyphus mistol (Caparidaceae), Prosopis alba (Leguminosae), Celtis sellowiana e Opuntia quimilo (Cactaceae), dentre outras.
Mercolli & Yanosky (1994) citam várias espécies de frutos encontrados no tubo digestivo de Tupinambis merianae como os de Eugenia uniflora (Myrtaceae), Syagrus romanzoffiana (Arecaceae) e Copernicia alba. Milstead (1961) encontrou no sistema digestivo do teiú T. merianae uma grande quantidade de frutos dos gêneros Vitis e Phylodendron (Williams et al. 1993). Já Reis (1995), na região de Santa Catarina, registrou em campo Tupinambis sp. consumindo frutos de palmito Euterpe edulis (Arecaceae).
A viabilidade das sementes das quatro espécies vegetais testadas na estufa de germinação não foi alterada por terem passado pelo tubo digestivo do animal. Apenas as sementes de Genipa americana mostraram uma alta taxa de germinação nas primeiras semanas quando comparadas com o controle. O efeito da passagem das sementes no tubo digestivo de lagartos é variável. Em algumas espécies de plantas, não houve diferença na capacidade de germinação de sementes passadas pelo sistema digestivo de lagartos (Whitaker 1987; Castilla 2000).
Segundo Traveset (1998), em uma revisão sobre os efeitos dos répteis na capacidade de germinação das sementes, das 41 espécies de plantas consumidas por lagartos, a maioria não foi afetada (56% dos casos) e, quando existia um efeito, era mais positivo (28%) que negativo (16%). Em contraste, os lagartos parecem modificar a velocidade de germinação em muitos casos (63%), acelerando-a mais vezes (47%) que desacelerando-a (16%).
No experimento de tempo de retenção das sementes no sistema digestivo do animal, foi observado que as sementes com diâmetro maior demoraram um maior tempo para passarem pelo tubo digestivo do teiú quando comparadas com as sementes menores. As diferenças no tempo de retenção das sementes no tubo digestivo do animal, assim como a qualidade do tratamento dado a elas, podem alterar o padrão de germinação (Schupp 1993).
O prolongamento do tempo de retenção de sementes de Withania frutescens no tubo intestinal de Podarcis lilfordi não afetou o sucesso germinativo (Castilla 2000). O tempo de retenção das sementes no tubo intestinal do lagarto não apresentou correlação com o sucesso germinativo, não havendo evidência de que o longo tempo de passagem das sementes pelo tubo aumente a sua taxa de germinação.
Segundo Traveset (1998) e Traveset & Verdú (2002), o tempo de retenção do alimento dentro do sistema digestivo não é determinado somente pela morfologia e fisiologia do animal. Vários fatores devem afetar o tempo de passagem pelo tubo digestivo, como o nível nutricional da dieta, consistência do alimento, dureza, água contida ou quantidade de alimento. O tempo de passagem também pode variar dependendo da quantidade de alimento consumido (Traveset 1998).
Os resultados indicam que Tupinambis merianae tem potencial para agir como dispersor de sementes, sendo muito provável ocorrer dispersão e germinação de algumas espécies em ambientes naturais. Estes lagartos se deslocam e forrageiam por áreas abertas (clareiras ou bordas) (Sazima & Haddad 1992), possibilitando a distribuição das sementes em locais propícios a germinação e estabelecimento. Nós sugerimos que maior atenção seja dada ao papel desses animais como dispersores de sementes nas regiões tropicais.
AGRADECIMENTOS
À Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP) pela Bolsa de Iniciação Científica concedida ao primeiro autor e pelo auxílio (96/10464-7 e 01/14463-5); ao Laboratório de Fisiologia Vegetal da Universidade Estadual Paulista pela facilidade de acesso a Estufa de Germinação e ao Prof. Dr. Augusto Abe por permitir o acesso aos recintos dos lagartos localizado no campus da Unesp de Rio Claro. MG recebe bolsa de produtividade de pesquisa do CNPq.
Recebido em: 28.07.2003
Aceito em: 06.02.2004
- Augspurger, C.K. & Kitajima, K. 1992. Experimental studies of seed recruitment from contrasting seed distributions. Ecology, 73:1270-1284.
- Bolkovic, M.L.; Protomastro, J.J. & Caziani, S.M. 1989. Dispersión de semillas por la iguana colerada (Tupinambis rufescens) y el mataco bola (Tolypeustes matacos) en el bosque de quebrachos y mistoles. In: Reunion Argentina Ecológica, 14. Jujuy, Arg.
- Castilla, A.M. 2000. Does passage time through the lizard Podarcis lilfordi's guts affect germination performance in the plant Withania frutescens? Acta Oecologica, 21:119-124.
- Cooper, W.E. & Vitt, L.J. 2002. Distribution, extent, and evolution of plant consumption by lizards. Journal of Zoology, 257:487-517.
- Cooper, W.E.; Caldwell, J.P.; Vitt, L.J.; Perez-Mellado, V. & Baird, T.A. 2002. Food-chemical discrimination and correlated evolution between plant diet and plant-chemical discrimination in lacertiform lizards. Canadian Journal of Zoology, 80:655-663.
- Donadio, O.E. & Gallardo, J.M. 1984. Biologia y conservación de las especies del género Tupinambis (Squamata, Sauria, Teiidae) em la Republica Argentina. Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia". Zoologia, 13:117-127.
- Durtsche, R.D. 1995. Foraging ecology of the fringe-toed lizard, Uma inomata, during periods of high and low food abundance. Copeia, 4:915-926.
- Figueira, J.E.C.; Vasconcellos Neto, J.G.; Garcia, M.A. & Souza, A.L.T. 1994. Saurocory in Melocactus violaceus (Cactaceae). Biotropica, 26:295-301.
- Fitzerald, L.A.; Chiani, J.M. & Donadio, O.E. 1991. Tupinambis lizards in Argentina: implementing management of a traditionally exploited resouce. In: Neotropical wildlife use and conservation University of Chicago, Chicago. p.303-316.
- Huey, H.B. & Pianka, E.R. 1981. Ecological consequences of foraging mode. Ecology, 62:991-999.
- Iverson, J.B. 1985. Lizards as seed dispersers? Journal of Herpetology, 19:292-293.
- Jordano, P. 2000. Fruits and frugivory. In: Fenner, M. (Ed.), Seeds, the ecology of regeneration in plant communities CABI, Wallingford. p.125-165.
- Köeppen, W. 1948. Climatologia. Fondo de Cultura Economica. Mexico, p.478.
- Lord, J.M. & Marshall, J. 2001. Correlations between growth form, habitat, and fruit colour in the New Zealand flora, with reference to frugivory by lizards. New Zealand Journal of Botany, 39:567-576.
- Losos, J.B. & Greene, H.W. 1988. Ecological and evolutionary implications of diet in monitor lizards. Biological Journal of the Linnean Society, 35:379-407.
- Mercolli, C. & Yanosky, A.A. 1994. The diet of adult Tupinambis teguixin (Sauria: Teiidae) in the eastern chaco of Argentina. Herpetological Journal, 4:15-19.
- Milstead, W.W. 1961. Notes on teiid lizards in Southern Brazil. Copeia, 4:493-495.
- Milton, K. 1981. Food choice and digestive strategies of two sympatric primate species. American Naturalist, 117:496-505.
- Moll, D. & Jansen, K.P. 1995. Evidence for a role in seed dispersal by two tropical herbivorous turtles. Biotropica, 27:121-127.
- Nogales, M.; Delgado, J.D. & Medina F.M. 1998. Shrikes, lizards and Lycium intricatum (Solanaceae) fruits: a case of indirect seed dispersal on oceanic island (Alegranza, Canary lslands). Journal of Ecology, 86:866-871.
- Olesen, J.M. & Valido A. 2003. Lizards as pollinators and seed dispersers: an island phenomenon. Trends in Ecology and Evolution, 18:177-181.
- Pagano, S.N. & Leitão Filho, E.F. 1987. Composição florística do estrato arbóreo da mata mesófila semidecídua no município de Rio Claro - Estado de São Paulo. Revista Brasileira de Botânica, 10:37-47.
- Pianka, E.R. 1980. Guild struture in desert lizards. Oikos 35:194-201.
- Presch, W. 1973. A review of the tegu lizards genus Tupinambis (Sauria: Teiidae) from South America. Copeia , 4:740-746.
- Rand, A.S. 1978. Reptilian arboreal folivores. In: Montgomery, G.G. (Ed.), The ecology of arboreal folivores Smithsonian Institute Press, Washington. p.115-122.
- Reis, A. 1995. Dispersão de sementes de Euterpe edulis Martius (Palmae) em uma floresta ombrófila densa Montana da Encosta Atlântica em Blumenau, SC Tese (Doutorado), Universidade Estadual de Campinas, Campinas, SP.
- Rick, C.M. & Bowman, R.I. 1961. Galapagos tomatoes and tortoises. Evolution, 15:407-417.
- Sáez, E. & Traveset, A. 1995. Fruit and nectar feeding by Podarcis lilfordi (Lacertidae) on Cabrera Archipelago (Balearic Islands). Herpetological Review, 26:121-123.
- Sazima, I. & Haddad, C.F.B. 1992. Répteis da Serra do Japi. In: Morellato, L.P.C. (Ed.), História Natural da Serra do Japi: ecologia e preservação de uma área florestal no sudeste do Brasil Universidade Estadual de Campinas/Fundação de Amparo à Pesquisa do Estado de São Paulo, Campinas, SP. p.212-235.
- Schupp, E.W. 1993. Quantity, quality, and the effectiveness of seed dispersal by animals. Frugivory and seed dispersal: ecological and evolutionary aspects. Vegetatio, 107/108:15-29.
- Sporne, K.R. 1965. The Morphology of Gymnosperms. In: The structure and evolution of primitive seed-plants Hutchinson University Library, London.
- Sylber, C. K. 1988. Feeding habits of the lizards Sauromalus varius and S. hispidus in the Gulf of California. Journal of Herpetology, 22:413-424.
- Tiffney, B. H. 1984. Seed size, dispersal syndromes and the rise of the angiosperms: evidence and hypothesis. Annals of the Missouri Botanical Garden, 71:55-576.
- Traveset, A. 1990. Ctenosaura similis Gray (Iguanidae) as a seed disperser in a Central American deciduous forest. American Midland Naturalist, 123:402-404.
- Traveset, A. 1995. Seed dispersal of Cneorum tricoccon L. (Cneoraceae) by lizards and mammals in the Balearic Islands. Acta Oecologica, 16:171-178.
- Traveset, A. 1998. Efect of seed passage through vertebrate frugivores guts on germination: a review. Perspectives in Plant Ecology, Evolution and Systematics, 1/2:151-190.
- Traveset, A. & Verdú, M. 2002. A Meta-analysis of the Effect of Gut Treatment on Seed Germination. In: Levey, D.J.; Silva, W.R. & Galetti, M. (Eds.), Seed dispersal and frugivory: ecology, evolution and conservation CABI, Wallingford, p. 339-350.
- Valido, A. & Nogales, M. 1994. Frugivory and seeds dispersal by the lizard Gallotia galloti (Lacertidae) in a xeric habitat of the Canary lslands. Oikos, 70:403-411.
- Vitt, L.J.; Zani, P.A.; Caldwell, J.P.; Araujo, M.C. & Magnusson W.E. 1997. Ecology of whiptail lizards (Cnemidophorus) in the Amazon region of Brazil. Copeia 4:745-757.
- Vitt, L.J.; Zani, P.A. & Esposito, M.C. 1999. Historical ecology of Amazonian lizards: implications for community ecology. Oikos, 87:286-294.
- Vitt, L.J.; Sartorius, S.S.; Avilla-Pires, T.C.S. & Esposito, M.C. 2001. Life at the river's edge: ecology of Kentropyx altamazonica in Brasilian Amazonia. Canadian Journal of Zoology, 79:1855-1865.
- Whitaker, A.H. 1987. The roles of lizards in New Zealand plant reproductive strategies. New Zealand Journal of Botany, 25:315-328.
- Whiting, M.J. & Greeff, J.M. 1997. Facultative frugivory in the cape flat lizard Platysaurus capensis (Sauria: Cordylidae). Copeia, 4:811-818.
- Williams, J.D.; Donadio, O.E. & Ré, I. 1993. Notas relativas a la dieta de Tupinambis rufescens (Reptilia: Sauria) del noroeste argentino. Neotropica, 39:45-51.
- Willson, M.F.; Sabag, C.; Figueroa, J.; Armesto, J.J. & Caviedes, M. 1996. Seed dispersal by lizards in Chilean rainforest. Revista Chilena de Historia Natural, 69: 339-342.
Datas de Publicação
-
Publicação nesta coleção
25 Ago 2004 -
Data do Fascículo
2004
Histórico
-
Recebido
28 Jul 2003 -
Aceito
06 Fev 2004