Abstracts
The currently accepted albatross taxonomy, based on characters of external morphology, plumage patterns, tail shape, bill size and coloration, organization of the plates of the bill, and, more recently, molecular data such as cytochrome-b gene sequences, resulted in a division of the family Diomedeidae into four genera: Diomedea, comprising the great albatrosses; Phoebastria, the North Pacific albatrosses; Thalassarche, the mollymawks; and Phoebetria, the sooty mollymawks. However, there are only a few, old studies on albatross osteology, which focused mostly on supra-generic relationships. Research on the group's taxonomy and anatomy is important in order to establish a secure basis for the identification of each species, including the differences between males, females and specimens of different ages, and also to verify anatomic characters which might be found useful for phylogenetic analysis based on morphological markers. In the present study, 63 skulls of Diomedea and Thalassarche albatrosses were analyzed and compared, thus establishing topographic correspondences and determining primary homologies, these resulting in: (a) the finding of no pattern of anatomical variation related to sex and age for both T. melanophris and for T. chlororhynchos; (b) the assessment of eight cranial landmarks separating the genera Diomedea and Thalassarche; (c) the recognition of 13 cranial landmarks differing among T. melanophris, T. chlororhynchos and T. cauta; (d) the re-identification of several specimens based on skull characters. The characters here presented for the genera and species, along with further anatomical research on the skull of the Diomedeidae, including the genera Phoebetria and Phoebastria, may help to enlighten relationships within the family.
Albatross; cranial osteology; Diomedea; Thalassarche
A atual taxonomia dos albatrozes consiste na divisão da família Diomedeidae em quatro gêneros: Diomedea, que inclui os grandes albatrozes; Phoebastria, formado pelos albatrozes do norte do Pacífico; Thalassarche, para os chamados "mollymawks" e Phoebetria, para os "mollymawks fuliginosos" e foi baseada em caracteres da morfologia externa, padrões de plumagem, forma da cauda, tamanho e coloração do bico e a organização das placas da ranfoteca e, mais recentemente, seqüências do citocromo-b. Entretanto, existem poucos e antigos estudos sobre a osteologia dos albatrozes que estudaram apenas as relações supra-genéricas. A pesquisa na taxonomia e na anatomia do grupo é importante para se criar uma base segura para a identificação das espécies, incluindo aí as diferenças existentes entre machos, fêmeas e de exemplares em diferentes idades, além de se verificar caracteres anatômicos cranianos que podem revelar-se úteis para análises filogenéticas com base em marcadores morfológicos. No presente trabalho, 63 crânios de representantes dos gêneros Diomedea e Thalassarche foram analisados e comparados, estabelecendo correspondências topográficas e determinando homologias primárias, o que resultou em: (a) a descoberta da ausência de padrões de distinção anatômica relacionados ao sexo e à idade para T. melanophris e para T. chlororhynchos; (b) levantamento de oito caracteres do crânio que separam os gêneros Diomedea e Thalassarche; (c) reconhecimento de 13 caracteres cranianos que diferem entre T. melanophris, T. chlororhynchos e T. cauta; (d) re-identificação de vários espécimes baseados em caracteres cranianos. Os caracteres aqui apresentados para os gêneros e espécies, juntamente com futuras pesquisas na anatomia do crânio de Diomedeidae, incluindo os gêneros Phoebetria e Phoebastria, podem ajudar na elucidação das relações de parentesco dentro da família.
Albatroz; osteologia craniana; Diomedea; Thalassarche
Cranial osteology and taxonomy of albatrosses of genus Dimedea linneaus, 1758 and Thalassarche reichenbach, 1853 (procellariformes: Diomeidae)
Francisco V. Dénes1 1 E.mail: fvdenes@gmail.com ; Luís Fábio Silveira2 2 E.mail: lfsilvei@usp.br
Departamento de Zoologia, Instituto de Biociências, Universidade de São Paulo, Caixa Postal 11.461, CEP 05422-970, São Paulo, SP, Brasil and Museu de Zoologia, Universidade de São Paulo, Caixa Postal 42.494, CEP 04128-970, São Paulo, SP, Brasil
ABSTRACT
The currently accepted albatross taxonomy, based on characters of external morphology, plumage patterns, tail shape, bill size and coloration, organization of the plates of the bill, and, more recently, molecular data such as cytochrome-b gene sequences, resulted in a division of the family Diomedeidae into four genera: Diomedea, comprising the great albatrosses; Phoebastria, the North Pacific albatrosses; Thalassarche, the mollymawks; and Phoebetria, the sooty mollymawks. However, there are only a few, old studies on albatross osteology, which focused mostly on supra-generic relationships. Research on the group's taxonomy and anatomy is important in order to establish a secure basis for the identification of each species, including the differences between males, females and specimens of different ages, and also to verify anatomic characters which might be found useful for phylogenetic analysis based on morphological markers. In the present study, 63 skulls of Diomedea and Thalassarche albatrosses were analyzed and compared, thus establishing topographic correspondences and determining primary homologies, these resulting in: (a) the finding of no pattern of anatomical variation related to sex and age for both T. melanophris and for T. chlororhynchos; (b) the assessment of eight cranial landmarks separating the genera Diomedea and Thalassarche; (c) the recognition of 13 cranial landmarks differing among T. melanophris, T. chlororhynchos and T. cauta; (d) the re-identification of several specimens based on skull characters. The characters here presented for the genera and species, along with further anatomical research on the skull of the Diomedeidae, including the genera Phoebetria and Phoebastria, may help to enlighten relationships within the family.
Keywords: Albatross, cranial osteology, Diomedea, Thalassarche.
RESUMO
A atual taxonomia dos albatrozes consiste na divisão da família Diomedeidae em quatro gêneros: Diomedea, que inclui os grandes albatrozes; Phoebastria, formado pelos albatrozes do norte do Pacífico; Thalassarche, para os chamados "mollymawks" e Phoebetria, para os "mollymawks fuliginosos" e foi baseada em caracteres da morfologia externa, padrões de plumagem, forma da cauda, tamanho e coloração do bico e a organização das placas da ranfoteca e, mais recentemente, seqüências do citocromo-b. Entretanto, existem poucos e antigos estudos sobre a osteologia dos albatrozes que estudaram apenas as relações supra-genéricas. A pesquisa na taxonomia e na anatomia do grupo é importante para se criar uma base segura para a identificação das espécies, incluindo aí as diferenças existentes entre machos, fêmeas e de exemplares em diferentes idades, além de se verificar caracteres anatômicos cranianos que podem revelar-se úteis para análises filogenéticas com base em marcadores morfológicos. No presente trabalho, 63 crânios de representantes dos gêneros Diomedea e Thalassarche foram analisados e comparados, estabelecendo correspondências topográficas e determinando homologias primárias, o que resultou em: (a) a descoberta da ausência de padrões de distinção anatômica relacionados ao sexo e à idade para T. melanophris e para T. chlororhynchos; (b) levantamento de oito caracteres do crânio que separam os gêneros Diomedea e Thalassarche; (c) reconhecimento de 13 caracteres cranianos que diferem entre T. melanophris, T. chlororhynchos e T. cauta; (d) re-identificação de vários espécimes baseados em caracteres cranianos. Os caracteres aqui apresentados para os gêneros e espécies, juntamente com futuras pesquisas na anatomia do crânio de Diomedeidae, incluindo os gêneros Phoebetria e Phoebastria, podem ajudar na elucidação das relações de parentesco dentro da família.
Palavras-chave: Albatroz, osteologia craniana, Diomedea, Thalassarche.
INTRODUCTION
The albatrosses are large seabirds belonging to the family Diomedeidae, with the species distributed in the genera Diomedea, Phoebastria, Thalassarche and Phoebetria (Nunn et al., 1996). Of the 21 species of albatrosses currently considered, nine are classified as Endangered, two of which considered Critically Endangered. The main threat to these species is long-line fishing, operated on a large scale in the southern oceans since the 1950's, with ever-increasing seabird by-catch (BirdLife International 2004; Favero et al. 2003, Neves et al. 2003, Tuck et al. 2003).
The number of albatross genera has varied notably during the group's taxonomic history. All known species were described in the genus Diomedea by Linnaeus (1758), these being reviewed only in 1853, when Reichenbach, through an analysis of external morphology,billme asurements and wing-tail proportions,erected the genera Phoebetria, Phoebastria and Thalassarche.
Coues (1866), based on characters of the hind limbs and nares, established the present families Diomedeidae and Procellariidae. In so doing, Coues (op. cit.) rejected two of the genera proposed by Reichenbach (1853) (Phoebastria and Thalassarche), subsuming them into Diomedea. Other authors, however, disagreeing with the treatment presented by Coues (op. cit.), proposed new genera for the albatrosses: Baird et al. (1884 apud Nunn et al., 1996), based on characters of the bill, created the genus Thalassageron; Mathews (1912 apud Nunn et al., 1996) described Diomedella and Nealbatrus, also accepting Thalassarche Reichenbach, 1853; and Murphy (1917) erected Rothonia.
Mathews (1934), in a revision of the entire family, admitted the existence of eight genera. Later, Mathews and Hallstrom (1943 apud Nunn et al., 1996) created a monotypic genus, Julieata, comprising only J. irrorata, an equatorial species. In 1948 Mathews himself rejected his arrangement of the albatross genera, and subsumed all the known species in the genus Diomedea Linnaeus. In the following year, Boetticher (1949 in Jouanin & Mougin, 1979) described three new genera, Galapagornis, Laysanornis and Penthirenia for the taxa Diomedea irrorata, Diomedea immutabilis and Diomedea nigripes, respectively. The story became even more complicated when, in 1965, Alexander et al., reestablished Coues' (1866) original proposition, accepting as valid only the genera Phoebetria and Diomedea. It is important to note that all cited authors based their conclusions on characters of external morphology, comparing plumage patterns, tail shape, bill size and coloration, besides the organization of the plates of the bill.
In 1996 Nunn et al. presented a first phylogeny of the family, based on cytochrome-b sequences, thus redefining the genera Diomedea and Phoebetria and restoring Phoebastria and Thalassarche. This work has been largely accepted, and even corroborated (Penhallurick, 2004). To date there has been no change in the generic level, nor any family revision based on morphological data (see Table 1 for a historic of the albatross taxonomy).
Despite being a fairly well-studied group, especially as to behavioral aspects, including migratory movements, there are only a few, studies on albatross osteology. These in the majority focused mostly on supra-generic relationships (e.g. Beddard 1898 and Alexander et al. 1965). Utilizing only one or two species of the family, these studies dedicated little attention to intra and/or interspecific variations, and since then there has been no analysis of the anatomical characters present in the Diomedeidae for inferring phylogenies. The two exceptions are an old and detailed work on the osteology of the Order Procellariiformes, by Pycraft (1899) and a study by Wilkinson (1969), in which the author described a fossil species, Diomedea thyridata, based on a bill fragment from an Upper Miocene deposit in Australia. This description was accompanied by a brief comparison of the bill structure of the fossil with several extant albatrosses.
In view of the delicate situation in which several albatross species are found, especially regarding their conservation, research on the group's taxonomy and anatomy is important, in order to establish a securebasis for identification of each species, including the differences between males, females and specimens of different ages, and also to verify anatomic characters which might be found useful for phylogenetic analysis based on morphological markers. These are the objectives of the present study.
MATERIALS AND METHODS
The osteological material used in the present study consists of 63 skulls of Diomedeidae from the following species (number of specimens between brackets): Diomedea dabbenena (4), D. epomophora (1),D. exulans (7), D. sanfordi (1), Thalassarche cauta (1), T. chlororhynchos (12), and T. melanophris (37) (see Appendix II for a detailed list of the material). This material belongs to the ornithological collections of the Museu de Zoologia da Universidade de São Paulo (MZUSP), the Departamento de Zoologia do Instituto de Biociências, Universidade de São Paulo (AZ), the Fundação Universidade do Rio Grande (FURG), and the Museu Oceanográfico do Vale do Itajaí, Universidade do Vale do Itajaí (MOVI). See also the section Re-identifications.
The skull of an adult specimen of Thalassarche melanophris was studied and then compared with other skulls of the same species, assorted according to sex and age class. These were then compared with the skulls of other species of the same genus. When possible, these were also separated according to sex and age (e.g., T. chlororhynchos). Finally, the comparison was extended to skulls of species of the genus Diomedea.
Photographs were taken by FVD using Sony Mavica and Canon PowerShot S70 digital cameras, and processed with Adobe Photoshop CS2.
The nomenclature used for the species of Diomedeidae follows Robertson & Nunn (1998) and Burg & Croxall (2004). For the osteological components we followed Baumel & Witmer (1993) and Zusi & Livezey (2000).
RESULTS
Ossa cranii: os frontale, os parietale, os lacrimale, os ectethmoidale, os mesethmoidale
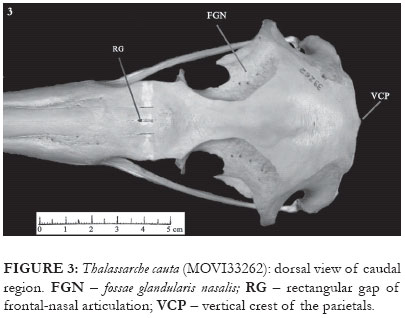
Thalassarche melanophris and T. cauta present a rectangular gap in the contact region between the frontal and nasal bones (RG: Figs. 1-3). This gap presents a variable size, and in some specimens (e.g. MZUSP 83, MZUSP 107) may even be occluded, being little conspicuous in T. chlororhynchos. In representatives of the genus Diomedea this fissure is absent. The medium portion of the frontal bone shows a clear narrowing, in which paired, deep and extensive fossae glandularis nasalis are present (FGN: Figs. 1 and 2). Between these fossae, in T. melanophris and T. chlororhynchos, there is a longitudinal groove with a similar extension. In general the larger the distance between the fossae the shallower the groove. In T. cauta the distance between the fossae is notably larger, although the groove is still evident (Fig. 3). In the skull of Diomedea, on the other hand, this groove is nearly absent.
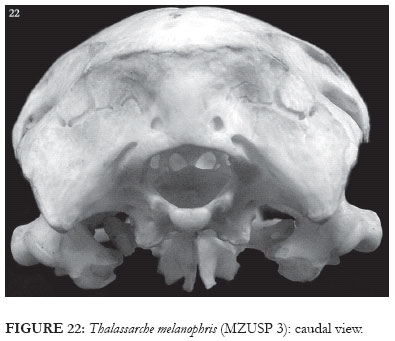
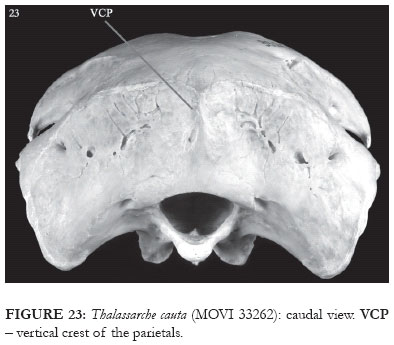
In T. cauta, T. chlororhynchos and in the representatives of Diomedea, the caudal portion of the fossa musculorum temporalium is slightly rounded (Fig. 23), whereas in T. melanophris it forms an acute angle with the crista nuchalis transversa (Fig. 22). Additionally, in T. cauta there is a vertical crest over part of the prominentia cerebellaris, this originating from the crista nuchalis transversa (VCP: Figs. 3 and 23).

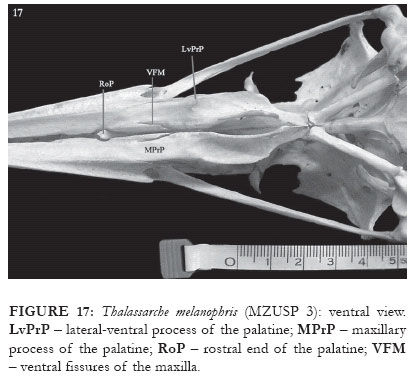
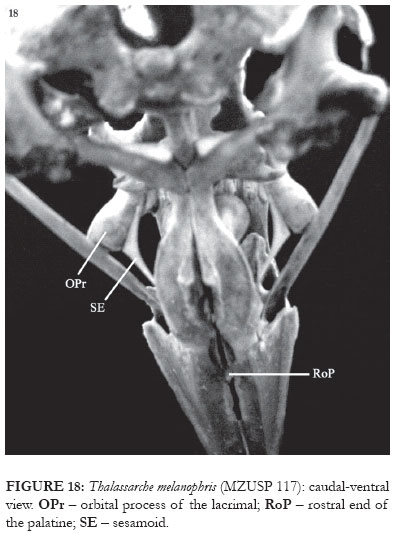
In the representatives of Thalassarche, the supraorbital process of the lacrimal is wide and robust, with the lateral edge being slightly rounded. In its lateral portion, medially there is a discrete crest (CrSOPr: Fig. 27), not occuring in Diomedea. In T. melanophris the orbital process of the lacrimal is long and much pneumatized, its ventral portion being free, not contacting the ectethmoid but contacting the jugal bar (OPr: Figs. 6 and 18). In its proximal portion there are two crests, superior (not always conspicuous) and inferior. The inferior crest presents a discrete groove along its length (ICrOPr: Fig. 27). Articulating with this groove there is a cylindrical-shaped sesamoid bone (SE: Fig. 18). The superior crest is much reduced in representatives of Diomedea and in T. chlororhynchos, the inferior crest being absent (the sesamoid then articulates through a recess in the rostral edge of the process).
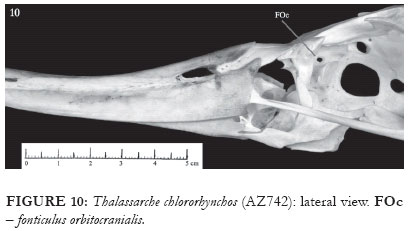
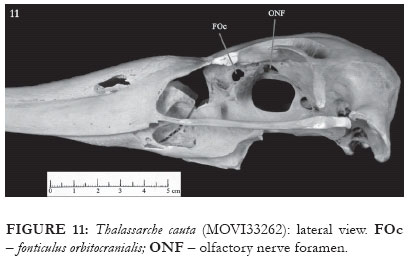
The ectethmoid contacts medially with the rostral portion of the interorbital septum. The facies nasalis presents a horizontal projection, perpendicular to the plane formed by the bone, and which runs through the whole extent of the ectethmoid. The facies also presents a round, shell-shaped crest in its distal portion (E: Fig. 7). The facies orbitalis is plain, and, in the representatives of genus Thalassarche, the distal end has a sharp, caudally-pointed projection (E: Figs. 4-5 and 8), which is absent in the representatives of Diomedea. In some of the skulls of T. chlororhynchos (AZ 749, PM 225), in T. cauta and in the representatives of Diomedea, the mesethmoid presents the fonticuli orbitocraniales (FOc: Figs. 9-14), whose aperture is widely variable. This structure is occluded by a bony wall in T. melanophris.
Os parasphenoidale, os basisphenoidale, os laterosphenoidale, os squamosum
In the species of the genus Thalassarche, the tuba auditiva forms an open fissure in almost its entire lenght, being covered (thus forming a bony tube) only at the caudal end. In representatives of genus Diomedea (except D. sanfordi) some pneumatic foramina are present at the proximal end of the structure.
The lamina parasphenoidalis has, in all of the analyzed species, a slightly triangular shape, with the lateral process occupying the two caudal vertices. Round-shaped basal tubercles are also present but not conspicuously (BT: Figs. 15-16). The crest in the median region of lamina parasphenoidalis, present in T. melanophris and T. chlororhynchos, is reduced in T. cauta and in some specimens of Diomedea.
In Thalassarche there is a small crest on the median part of the laterosphenoid. This crest begins rostrally at the caudal edge of the foramen opticum and extends towards the orbit's superior edge, where it then turns and continues to the postorbital process region (CrLS: Fig. 19). In the contact region between the laterosphenoid and the frontal bones, there is a cleft that begins close to the caudal edge of the olfactory nerve foramen and follows to the crest described above (ClLS: Fig. 19). In representatives of the genus Diomedea, the olfactory nerve foramen is partially or completely enclosed by a bony plate that extends to the ectethmoid, whereas the dorsal cleft of the laterosphenoid begins from a foramen that communicates with the fossa glandularis nasalis (ClLS: Fig. 20).
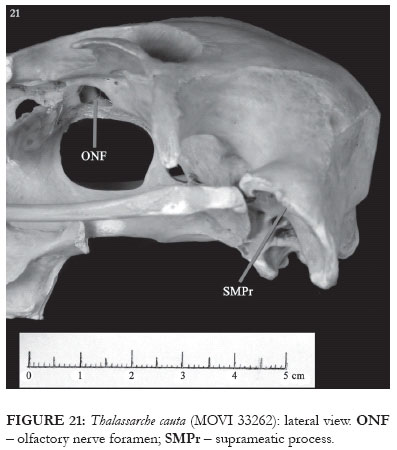
In T. melanophris, caudally to the squamosal cotyla, the suprameatic process of the squamosal, a small rounded protuberance, forms part of the laterocaudal wall of the cotyla itself (SMPr: Figs. 4-6). In T. cauta and T. chlororhynchos, the suprameatic process is notably smaller (SMPr: Figs. 9 and 21). This structure did not vary in the other studied specimens.
Os basioccipitale, os exoccipitale, os supraoccipitale
Along the fossa subcondylaris of the basioccipital, Thalassarche cauta presents a discrete, rostrally oriented crest which is absent in all the other studied specimens. In T. melanophris and T. cauta there is a process lateral to the occipital condyle, located between the fossa parabasalis and the fossa subcondylaris (Figs. 15-16 and 22). This structure shows a high degree of variation in size among the studied specimens; however, this variation does not follow any sexual or age pattern. In T. chlororhynchos this structure is reduced or absent, being also absent in representatives of Diomedea.
Ossa faciei: os nasale, os premaxillare, os maxilare
The size of the nare openings is highly variable in all the species, without any correlation as to sex or age of the specimens. In T. cauta and T. chlororhynchos these openings are, in general, smaller that those present in T. melanophris (Figs. 9-12).
In the representatives of Thalassarche, in the caudoventral portion of the maxilla, in the contact region with the palatine bone, two small, blade-edged fissures, forming a "V" shaped cleft, are present (VFM: Figs. 15-17). These blades are curved medium-dorsally, forming a tubular space in the bill region (CBM: Figs. 7-8). In this region there are four openings formed by the ventral portion of the maxilla, the lateral-ventral region of the nasal bone and the rostrodorsal portion of the palatine (COM: Fig. 24). The two most lateral openings are vertically aligned, with the jugal bar extending between them (Fig. 24). The superior opening is elliptic shaped, with the longer axis in the vertical position, while the inferior opening is of an irregular shape. The other two openings are medial, the inferior one presenting an obliterated bottom, while the superior one, along with the other two lateral openings, gives access to the pneumatized chamber inside the bill (Fig. 24). In the caudal-ventral end of the upper jaw there is a thin process that extends caudally over the rostral portion of the palatine (CPrM: Figs. 5 and 8).
On the other hand, in representatives of Diomedea the openings present a different size and alignment. The assemblage is slightly rotated to the medial region, in such a way that the openings are in lateral, superior, medial and inferior positions, corresponding to the lateral inferior, lateral superior, medial superior and medial inferior openings in T. melanophris. The lateral superior opening, in T. melanophris, is the smallest, whereas in the Diomedea it is the most developed, being separated from the medial superior opening by a small bone column (absent in some specimens, e.g. MOVI 06610, MOVI 10335 and MOVI 13335; Figs. 25-26).
Os palatinum, os pterygoideum, os jugale, os vomer
In T. melanophris the palatine articulates rostroventrally with the upper jaw through its maxillary process in a flat contact region (MPrP: Fig. 15-17). Still in the ventral region, in the medial portion the palatine presents a conspicuous lateral-ventral process (LvPrP: Figs. 15-17). Also evident is the ventral crest, which delimits the edge of the fossa choanalis, followed laterally by the fossa medialis (Figs. 15-17). Dorsally to the lateral crest there is the lamella choanalis, which is a conspicuous, shell shaped, lateral expansion of pars choanalis that almost reaches the ectethmoid (LaCho: Figs. 5 and 7-8). This dorsal expansion of the palatine covers the entire ventral surface of the interorbital septum, and extends inwards to the upper maxilla. This expansion also bends ventrally, and its rostral end, similar to a small process, is visible through the palatine fenestra (RoP: Figs. 15-18).
In T. cauta the lateral-ventral process of the palatine is highly reduced. In T. melanophris the dorsal extension of the palatine presents a flat profile when viewed in cross-section. In T. cauta this structure is much deeper, assuming a "V" shaped profile. The vomer is absent in all the species studied.
Ossa mandibulae
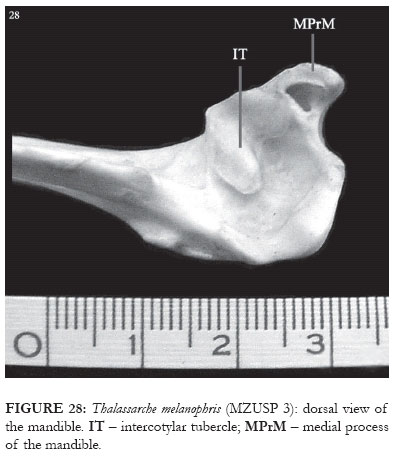
Medial and lateral cotylae are visible in the articular region of the mandible in species of the genus Thalassarche. The intercotylar tubercle is present in a slightly rectangular shape (IT: Fig. 28). Also present is the medial mandibular process, with a foramem pneumaticum (MPrM: Fig. 28).
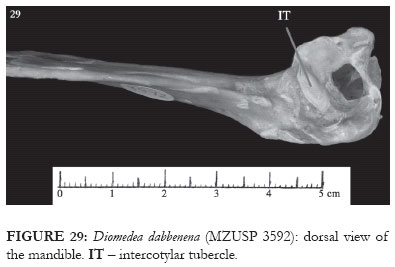
In representatives of Diomedea there is a large pneumatic opening in the medial cotyla (Fig. 29). The intercotylar tubercle is of irregular shape, with its medial end placed rostrally in relation to the lateral end (IT: Fig. 29). The pneumatic foramen of the medial mandibular process, present in the skull of the species of Thalassarche, is absent in the representatives of Diomedea.
Re-identifications
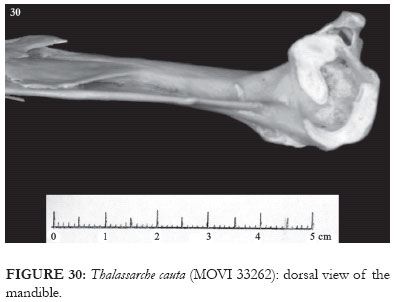
As this study was being developed, it was realized that one of the D. exulans skulls (MOVI 33262) had, when compared to others of the same species, differences related both to size (being notably smaller) and morphology. The skull presents a series of characters, such as the shape and position of the maxilla's caudal portion openings, the non-occluded olfactory nerve foramen (ONF: Figs. 11-12 and 21), and the absence of the pneumatic foramen at the caudal end of the mandible Fig. 30), all of which indicate a greater proximity to the representatives of the genus Thalassarche than to those belonging to the genus Diomedea.
Granted that the specimen in question is a Thalassarche albatross, the size of the skull suggests that it does not belong to either T. melanophris, T. chrysostoma, or T. chlororhynchos, whose skulls are notably smaller. However, T. cauta, a species that occurs in Brazilian waters, is of a larger size, similar to Diomedea exulans (Harrison, 1987; Murphy, 1936). Double et al. (2003), utilize ten body measurements to compare the species T. cauta and T. steadi. Due to the study being focused on the species conservation, the authors chose to use only measurements capable of being taken from live specimens, thus making it difficult to compare them to those taken from the skull. In spite of this fact, the measurements "skull width" (in reference to the distance between the postorbital processes) and "total bill length" (from the rostral end of the culmen to the posterior margin of the thickening that marks the caudal edge of the culmenicornium), both corresponding to "maximum head width" and "culmen length", were taken from the skull in question, and fit within the interval proposed by Double et al. op. cit.) (Attachment I). Unfortunately, the skull of T. cauta is absent from Brazilian collections, so a crucial, thorough comparison could not be done. However, the re-identification of this specimen as Thalassarche cauta is recommended.
The only T. chrysostoma specimen analyzed has a fully dark ramicornium, while both the culmenicornium and the unguicornium are yellowish (Fig. 31). This coloration is different from the pattern described for T. chrysostoma, where the ramicornium always presents light parts when the culmenicornium and the unguicornium are yellowish, though this fits in perfectly with the T. chlororhynchos' pattern, where the ramicornium is always dark. Additionally, the capture coordinates of this specimen (Attachment II), well inside the T. chlororhynchos distribution area, are outside that of T. chrysostoma, a more southern species (Harrison, 1983). Thus, specimen MZUSP 95, previusly attributed to T. chrysostoma, was re-identified as Thalassarche chlororhynchos.
Finally, it was realized that one specimen (MZUSP 16098), identified by Pinto (1938) as Diomedea epomophora, that was later re-identified by Grantsau (1995) as D. exulans, is in fact a D. epomophora albatross. This new identification is based on the shape of the external opening of the nare (elliptic in D. exulans and round in D. epomophora; Fig. 32), and on the presence of a black line on the edge of the upper maxilla, characteristic of D. epomophora (Fig. 33). Plumage characters also corroborate Pinto's (op. cit.) initial diagnosis, these being that this specimen presents white upper coverts on the wing's leading edge, a black tail and an absence of black on the lower coverts in the carpal joint region, which diagnoses for D. sanfordi (Fig. 34) (Harrison, 1987; Murphy, 1936).
DISCUSSION AND CONCLUSIONS
Anatomy
Pycraft (1899) analyzed the skeleton of several representatives of the Procellariiformes, in a detailed study on their osteology, seeking to enlighten the relationships between the families, as well as the position of the Order within the class Aves. As regards the albatrosses, the author studied the complete skeletons of two species of the genus Diomedea (sensu Coues, 1866), D. exulans and D. melanophris, and the skulls of representatives of the genus Thalassogeron and that of Phoebetria, and suggests a systematic arrangement of the family, comprising the genera cited above. In general, the descriptions provided by Pycraft (op. cit.) agree with those of the present study, despite nomenclatural differences of some of the osteological components. Such structures will here be properly corresponded, following the sequence presented in Results.
The paired "supraorbital grooves" in the interorbital region of the frontal, described by Pycraft (1899), correspond to the paired fossae glandularis nasalis. Pycraft (op. cit.) is especially interested in the outer wall of these fossae which, according to the author, is well developed in Diomedeidae (in contrast with other representatives of the Order). No pattern of distinction has been found in the present study for this particular structure, among representatives of the different genera. Pycraft (op. cit.) also describes, in the median line of the frontal-parietal region, a groove that indicates the position of the palial cerebral fissure, and that probably corresponds to the groove between the fossae glandularis nasalis described above.
In the region of the parietal bone, Pycraft (1899) terms as
fossa temporalis the
fossa musculorum temporalium, this description being concordant with that offered in the present study. Pycraft's
(op. cit.) description of the lacrimal also generally concordant with that presented herein. However, this author describes the absence of the "anterior limb of the lacrimal", which apparently refers to the bone's anterior region, attributing it to
Diomedea melanophris. No resemblance to the above described condition was found for the skulls of
Thalassarche melanophris in the present study, suggesting that the specimen analyzed by the author (in this case, a single complete skeleton), was damaged. In addition, Pycraft
(op. cit.) describes the structure of an "ossiculum lachrymo-palatinum or os crochu", corresponding to the sesamoid bone. According to the author, in the Procellariiformes this bone is most developed in the representatives of Diomedeidae. Its presence is, however, of no taxonomic value, since it is also present in several non-related groups.
The ectethmoid is termed by Pycraft (1899) as "the aliethmoid, ectoethmoidal ossification that forms the antorbital plate", its description being concordant with that offered in the present study, except for the representatives of genus Phoebetria. The author does not mention the sharp, caudally pointed, projection observed in the representatives of genus Thalassarche.
In his description of the base of the skull, Pycraft (1899) cites a "basitemporal plate of the parasphenoid", that corresponds to lamina parasphenoidalis. Special attention is given to the tuba auditiva, termed "Eustachian groove", especially to the structure's pneumatic foramen. According to the author, the opening found within this "groove" (that corresponds to the pneumatic foramen observed in the representatives of genus Diomedea), is located and directed differently in Diomedeidae than in the rest of the Order. Pycraft (op. cit.) also notes that in Diomedea exulans there is a second opening, lateral to the parasphenoidal rostrum. In Thalassogeron and in Phoebetria, "the Eustachian groove is shallower, wider and without the foramina penumatica". As regards the rest of this structure, both descriptions are concordant, though the lateral processes of the lamina are termed by Pycraft (op. cit.) as "mammillary processes". In addition the author cites "a well marked tubercle lying between the mammillary processes", which corresponds to the basal tubercles. There is no reference to the crest present in the lamina's medial region.
Pycraft's (1899) description of the lateral portion of the skull, including the region of cavitas tympanica, the squamosal bone and the orbita, concurs with that observed in the present study, despite the fact that the author uses the term "orbitosphenoid" for part of the region occupied by the laterosphenoid, which in turn is not mentioned. As regards the facial skeleton, Pycraft (op. cit.) stresses the resemblance of the region to that of the representatives of the Order Ciconiiformes. In the description of the maxillar, caudo-ventral region, the author terms as "maxillopalatine processes" those laminar structures associated with the V-shaped fissures of the upper jaw. The pneumatized chamber inside the bill is referred to as the "antrum of Highmore", whose posterior openings are also described. The main difference between the descriptions is that Pycraft (op. cit.) does not recognize the inferior opening, which is understandable, seeing that is has an obliterated bottom. Thus, only one medial, inner opening and two lateral openings are described. Additionally, for D. exulans, the author describes the large widening of the medial opening (termed, in the present study, lateral superior opening in Thalassarche and superior opening in Diomedea), as observed in Diomedea, the opposite for Phoebetria.
Wilkinson (1969) presents a brief comparison of the bill structure of some of the species of the family, including the two representatives of the genus Phoebetria, Diomedea exulans, Thalassarche cauta, T. chlororhynchos, T. melanophris, and T. chrysostoma (the last four species were treated as still belonging to the genus Diomedea). Using the nomenclature proposed by Pycraft (1899), the author uses the differences of size of the posterior inner opening of the "antrum of Highmore" to separate the genus Diomedea (sensu Alexander, 1965) from that of Phoebetria, and to place a fossil species, D. thyridata. It's interesting that, seeking to elucidate the kinship of D. thyridata among the representatives of Diomedea, Wilkinson (op. cit.), divides the genus into two groups, termed the D. melanophris group (whose species coincide with those of the current genus Thalassarche) and D. exulans group (current genera Diomedea and Phoebetria), based on several characters of the bill (see below). However, the author does not recognize the differences in the posterior openings of the "antrum", clearly verified between the representatives of Diomedea and Thalassarche.
Wilkinson (1969) also creates a new term, "culmenal ridge" to describe the structure formed by the fusion of the "nasal processes of the premaxilla", which was not described by Pycraft (1899). This structure corresponds to the frontal process of the premaxilla.
Finding the correspondence between Pycraft's(1899) description and that of the present study, is most difficult for the region that includes the palatine, vomer and pterygoid. With the exception of the rostral portion, the descriptions of the palatine are fairly the same. This author terms the region of the palatine that extends inside the maxilla, and whose rostrum is visible through the palatine fenestra, as the vomer (which in the present study was considered absent), and stresses the resemblance of the structure in the Procellariiformes to that of the Ciconiiformes and Anseriiformes, as regards the extent of the fusion of os palatinum. The caudal limit of the vomer, where it merges with the palatine is, according to Pycraft (op. cit.), right below the rostral expansion of the esethmoid,where there is a notch in the palatine.
The skulls of Mycteria americana (AZ 1074) and of Cygnus buccinator (AZ 565), representatives of the Orders Ciconiiformes and Anseriformes, respectively, with size comparable to that of the species of Diomedeidae, were studied to verify the "great resemblance" suggested by Pycraft (1899) as regards the vomer observed by him in the Procellariiformes. Nonetheless in these skulls the vomer is clearly visible as a thin, rostrocaudally oriented blade that originates from the caudal end of os palatinum, quite differently from the structure appointed as "similar" by Pycraft (op. cit.) in Diomedeidae, which is compressed dorsoventrally and originates from the rostro-dorsal end of os palatinum.
Pycraft (1899), besides studying the skeleton of adult specimens of several species of Procellariiformes, also analyzed the bone structure of two Oceanodroma leucorrhoa (Hydrobatidae) nestlings. His description of the vomer region in these specimens, as "a pair of elongated, flattened laminae, anteriorly united in the median line, whose free posterior ends are received by the anterior borders of the hemipterygoids, and that are bounded on either side by an inwardly turned scroll of bone from the dorsal border of the palatine (here in reference to the lamella choanalis, observed in the present study)", does not help in understanding the structure observed in the adult skull of representatives of Diomedeidae, which, according to Pycraft himself, is "peculiar" in the general context of the Order.
Considering the differences between the descriptions presented, the observations of the skull structure of representatives of the Ciconiiformes and the Anseriformes, and the relative distance between the families Diomedeidae and Hydrobatidae within the Order Procellariiformes in recent phylogenies (Kennedy & Page, 2002), it is safe to conclude that the real homology of this structure will only be clarified with the study of Diomedeidae nestlings, whose skulls, lacking the intensified bone fusion characteristic of the adult, will most likely be decisively informative.
The description of the mandible of Diomedeidae, as presented by Pycraft (1899), is relatively simple in comparison to that of the other structures mentioned above. The "posterior lateral vacuity, which pierces the posterior end of the supra-angular", probably corresponds to fenestra mandibulae caudalis.
Variations of the skull related to sex and age
No pattern of anatomical distinction between the skulls of individuals of different sexes was found either for T. melanophris or for T. chlororhynchos. This might have been so determined by the small number of skulls in the analysis, since most of the material lacks sexual identification. Thus, an analysis with more, duly identified skulls is necessary to confirm this result. There was also no pattern of distinction between individuals of different ages. Here, however, the situation is different, since the determination of age classes, in the skulls where this information was absent, was made possible by observation of ramphotheca coloration, still present in most of the material. Nevertheless, as the age classes were not perfectly represented (most skulls belonging to juvenile or subadult specimens), it is possible that in an analysis with more skulls, a different condition is to be found.
Variations of the skull between the genera and species of diomedeidae
At present, delimitation of the Diomedeidae genera is based on biogeography, morphometry, external morphology and, more recently, on mitochondrial DNA sequences. Having the relatively scarce number of publications about the family's compared osteology as an example, it is evident that the search for anatomical evidence to corroborate the present taxonomy was neglected.
Pycraft (1899) notes osteological differences between the Procellariiformes, and elaborates a series of identification keys for the order's families and genera, considering for Diomedeidae the genera Diomedea (sensu Coues, 1866), Thalassogeron and Phoebetria. The author refers only to the species Diomedea exulans and "Diomedea melanophrys", the species representing the other two genera in that study being unknown. At that time Thalassogeron was composed of T. chlororhynchos,T. cauta, T. layardi (at present subsumed into T. cauta), T. salvini and T. culminatus (at present subsumed into T. chrysostoma). The characters that distinguish the genus Diomedea from the genus Thalassogeron in the key presented by Pycraft, are: the ratio between the width of the interorbital region of the frontal and the width of the same on the line between the lacrimals, larger in Thalassogeron than in Diomedea; and the shape of the caudal region of the palatine, wider in Diomedea than in Thalassogeron.
The characters observed in the present study that suggest the separation of representatives of the genus Diomedea from those of the genus Thalassarche are (see also table 2): 1. the groove between the fossae glandularis nasalis is either not conspicuous or absent; 2. the absence of the median crest on the supraorbital process of the lacrimal; 3. the fonticulus orbitocranialis is always present (in the skull of Thalassarche melanophris this is absent); 4. the presence of pneumatic openings in the rostral region of the tuba auditiva, withthe exception of the D. sanfordi skull; 5. occlusion of the olfactory nerve foramen; 6. the different shape, size and position of the posterior openings of the caudal vacuity of the maxilla; 7. the presence of a large pneumatic opening on the cotyla medialis of thecaudal region of the mandible; 8. the absence, in most cases, of the pneumatic foramen on the medial process of the mandible.
The conspicuity of the groove between the fossae glandularis nasalis is possibly related to the distance between the same, in a way that one might establish a relationship between this and the first character presented by Pycraft (1899) the separation by the relative width of the orbital region of the frontal bone if one considers that the specimen of Thalassogeron used in that study is representative of Thalassarche. Among the species of Thalassarche analyzed, the only one wherein the distance between the fossae is significantly larger than that verified for the species of Diomedea, is T. cauta, which might indicate that this was the species most probably employed by Pycraft to represent Thalassogeron.
Pycraft (1899) mentions the presence of pneumatic foramina on the tuba auditiva for D. exulans. Since the author includes T. melanophris in the genus Diomedea, his decision to exclude this character from the genus diagnosis is understandable. Curiously, these structures, as observed in the skull of D. epomophora, are absent in D. sanfordi, which clearly shows the need of further studies on representatives of the genus Diomedea.
Characters also absent from Pycraft's (1899) identification keys are the differences of shape, size, and position of the posterior openings of the caudal vacuity of the maxilla, and of the pneumatic foramina of the caudal region of the mandible, despite being present in the author's anatomical description of the skull (again probably due to the inclusion of T. melanophris in the genus Diomedea). Wilkinson (1969), adopting a different taxonomic arrangement, considers the posterior openings of the maxilla to position the fossil species D. thyridata, but does not describe the differences between these openings when commenting on the "groups" within the genus Diomedea. The observation of the condition of these openings in the genus Phoebastria, not included in the present study, might help in understanding this author's decision. The other characters were not commented on by either Pycraft (1899) or Wilkinson (1969).
The only character varying between species of the genus Diomedea was the absence of a pneumatic foramina on the rostral portion of the tuba auditiva, for D. sanfordi. There is no reference to this specific situation in the osteological literature. Since the number of Diomedea skulls analyzed is relatively small, it is possible that this result will not coincide with that of a more comprehensive study.
For the genus Thalassarche, the characters that differ from those observed in T. melanophris are (see also table 3): 1. a fissure in the region of the articulation between the nasal and frontal bones, generally smaller in T. chlororhynchos and T. cauta; 2. the distance between the fossae gladularis nasalis is greater in T. cauta; 3. the shape of the caudal region of fossa musculorum temporalium is different in T. chlororhynchos and T. cauta; 4. the presence of a vertical crest on part of the prominentia cerebellaris in T. cauta; 5. the inferior crest on the supraorbital process of the lacrimal is reduced in T. chlororhynchos; 6. the presence of the fonticulus orbitocranialis in some of the skulls of T. chlororhynchos and T. cauta; 7. the crest on the lamina parasphenoidalis is reduced in T. cauta; 8. the suprameatic process of the squamosal is reduced in T. chlororhynchos and T. cauta; 9. the crest on fossa subcondylaris is present in T. cauta; 10. the lateral process on the condylus occipitalis is reduced or absent in T. chlororhynchos; 11. the size of the external nare openings is generally smaller in T. chlororhynchos and T. cauta; 12. the lateroventral process of the palatine is highly reduced in T. cauta; 13. the cross-section profile of the dorsal expansion of the palatine is "V"-shaped in T. cauta.
Based on the number of differences between representatives of T. melanophris and T. chlororhynchos, one might suggest that these species are more closely related to each other than to T. cauta, although a study including other species of the genus, and employing methods of phylogenetic systematics, is necessary to support such a statement. The characters here presented for the genera and species studied might contribute significantly to enlightening kinship relationships within the family Diomedeidae.
ACKNOWLEDGMENTS
We thank Fábio Olmos, Caio José Carlos, Guilherme Renzo Rocha Brito, Renato Gaban Lima, Tatiana Neves and two anonymous referees for providing us with comments, advice or literature relevant to the manuscript. We are indebted to Jules Soto (MOVI) and Carolus M. Vooren (FURG) for allowing access to specimens, and to Projeto Albatroz for donating specimens to the MZUSP. FVD was supported by a Scientific Initiation scholarship from The State of São Paulo Research Foundation (FAPESP).
REFERENCES
ALEXANDER, W.B.; FALLA, R.A.; JOUANIN, C.; MURPHY, R.C.; SALOMONSEN, F.; VOOUS, K.H.; WATSON, G.E.; BOURNE, W.R.P.; FLEMING, A.C.; KURODA, N.H.; ROWAN, M.K.; SERVENTY, D.L.; TICKELL, W.L..N; WARHAM, J. & WITERBOTTOM, J.M. 1965. The families and genera of petrels and their names. Ibis, 107:401-405.
BAUMEL, J.J. & WITMER, L.M. 1993. Osteologica. In: Baumel, J.J.; King, A.S.; Breazile, J.E.; Evans, H.E. & Berge, J.C.V. (Eds.), Handbook of Avian Anatomy: Nomina Anatomica Avium. Nutall Ornithological Club. Cambridge.
BEDDAR, F.E. 1989. The Structure and classification of birds. Longmans, Green and Co., London.
BIRDLIFE INTERNATIONAL 2004. Threatened birds of the world 2004. Cambridge, BirdLife International. 1 CD-ROM.
BURG, T.M. & CROXALL, J.P. 2004. Global population structure and taxonomy of the wandering albatross species complex. Molecular Ecology, 13:2345-2355.
COUES, E. 1866. Critical review of the Family Procellariidae. Part V. Embracing the Diomedeinae and the Halodrominae. Proceedings of the Academy of Natural Sciences of Philadelphia, 18:172-197.
DOUBLE, M.C.; GALES, R.; REID, T.; BROTHERS, N. & ABBOT, C.L. 2003. Morphometric comparison of Autralian Shy and New Zealand White-capped Albatrosses. Emu, 103:287-294.
FAVERO, M.; KHATACHIKIAN, C.E.; ARIAS, A.; RODRIGUEZ, M.P.s.; CAÑETE, G. & MARIANO-JELICICH, R. 2003. Estimates of seabird by-catchalongthePatagonianShelf byArgentinelonglinefishing vessels, 1999-2001. Bird Conservation International, 13:273-281.
HARRISON, P. 1983. Seabirds: An identification guide. Houghton Mifflin, Boston.
HARRISON, P. 1987. Seabirds of the World: A photographic guide. Princeton University Press, Princeton.
GRANTSAU, R. 1995. Os Albatrozes (Diomedeidae, Procellariiformes) do Atlântico e suas ocorrências na costa brasileira e uma chave de identificação. Boletim do Centro de Estudos Ornitológicos, 12:20-31.
JOUANIN, C. & MOUGIN, J.L. 1979. Order Procellariiformes. In: Mayr, E. & Cottrel, G.W. (Eds.), Check-list of birds of the World. 2nd ed. Harvard University Press, Cambridge, Massachusetts, v.1, p. 48-121.
KENNEDY, M. & PAGE, R.D.M. 2002. Seabird Supertrees: Combining partial estimates of procellariiform phylogeny. The Auk, 119:88-108.
LINNAEUS, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species cum characteribus, differentiis, synonymis, locis. Lautentii Salvii, Stockholm.
MATHEWS, G.M. 1934. Remarks on albatrosses and mollymawks. Ibis, 13:807-816.
MATHEWS, G.M. 1948. Systematic notes on the petrels. Bulletin of the British Ornithologists Club, 68:155-170.
MATHEWS, G.M. & HALLSTROM, J.L. 1943. [Apud Nunn, G.B. et al]. 1996. Notes on the order Procellariiformes. The Federal Capital Press of Australia Limited, Canberra.
MURPHY, R.C. 1917. A new albatross from the west coast of South America. Bulletin of the American Museum of Natural History, 37:861-864.
MURPHY, R.C. 1936. Oceanic birds of South America. MacMillan, New York.
NEVES, T. DA S.; OLMOS, F. & PEPPES, F.V. 2003. Plano de Ação Nacional para a Conservação de Albatrozes e Petréis. Available at: <http://www.projetoalbatroz.com.br>.
NUNN, G.B.; COOPER, J.; JOUVENTIN, P.; ROBERTSON, C.J.R. & ROBERTSON G.G. 1996. Evolutionary relationships among extant albatrosses (Procellariiformes: Diomedeidae) established from complete cytochrome-b gene sequences. The Auk, 113:784-801.
PENHALLURICK, J. & WINK, M. 2004. Analysis of the taxonomy and nomenclature of the Procellariiformes based on complete nucleotide sequences of the mithocondrial cytochrome-b gene. Emu, 104:125-147.
PINTO, O.M. de o. 1938. Catálogo das Aves do Brasil e Lista dos Exemplares que as representam no Museu Paulista. Revista do Museu Paulista, tomo XXII.
PYCRAFT, W.P. 1899. Contributions to the Osteology of Birds. Part III. Tubinares. Proceedings of the Zoological Society of London, 1899: 381-411.
REICHENBACH, L. 1853. Avium Systema Naturelle. Leipzig, Germany.
ROBERTSON, C.J.R. & NUNN, G.B. 1998. Towards a new taxonomy for the albatrosses. In: Robertson, G. & Gales, R.(Eds.), Albatross Biology and Conservation. Surrey Beatty & Sons, Chipping Norton, NSW, Australia, p.13-19.
TUCK, G.N.; POLACHECK, T. & BULMAN, C.M. 2003. Spacio-temporal trends of longline fishing effort in the Southern Ocean and implications for seabird bycatch. Biological Conservation, 114:1-27.
WILKINSON, H.E. 1969. Description of an Upper Miocene albatross from Beaumaris, Victoria, Australia, and a review of the fossil Diomedeidae. Memoirs of the National Museum of Victoria, 29:41-51.
ZUSI, R.L. & LIVEZEY, B.C. 2000. Homology and phylogenetic implications of some enigmatic cranial features in galliform and anseriform birds. Annals of Carnegie Museum, 69:157-193.
Recebido em: 21.11.2006
Aceito em: 13.03.2007
Publicado em: 15.06.2007

Clique aqui para ampliar


Clique aqui para ampliar

- ALEXANDER, W.B.; FALLA, R.A.; JOUANIN, C.; MURPHY, R.C.; SALOMONSEN, F.; VOOUS, K.H.; WATSON, G.E.; BOURNE, W.R.P.; FLEMING, A.C.; KURODA, N.H.; ROWAN, M.K.; SERVENTY, D.L.; TICKELL, W.L..N; WARHAM, J. & WITERBOTTOM, J.M. 1965. The families and genera of petrels and their names. Ibis, 107:401-405.
- BAUMEL, J.J. & WITMER, L.M. 1993. Osteologica. In: Baumel, J.J.; King, A.S.; Breazile, J.E.; Evans, H.E. & Berge, J.C.V. (Eds.), Handbook of Avian Anatomy: Nomina Anatomica Avium. Nutall Ornithological Club. Cambridge.
- BEDDAR, F.E. 1989. The Structure and classification of birds Longmans, Green and Co., London.
- BIRDLIFE INTERNATIONAL 2004. Threatened birds of the world 2004 Cambridge, BirdLife International. 1 CD-ROM.
- BURG, T.M. & CROXALL, J.P. 2004. Global population structure and taxonomy of the wandering albatross species complex. Molecular Ecology, 13:2345-2355.
- COUES, E. 1866. Critical review of the Family Procellariidae. Part V. Embracing the Diomedeinae and the Halodrominae. Proceedings of the Academy of Natural Sciences of Philadelphia, 18:172-197.
- DOUBLE, M.C.; GALES, R.; REID, T.; BROTHERS, N. & ABBOT, C.L. 2003. Morphometric comparison of Autralian Shy and New Zealand White-capped Albatrosses. Emu, 103:287-294.
- FAVERO, M.; KHATACHIKIAN, C.E.; ARIAS, A.; RODRIGUEZ, M.P.s.; CAÑETE, G. & MARIANO-JELICICH, R. 2003. Estimates of seabird by-catchalongthePatagonianShelf byArgentinelonglinefishing vessels, 1999-2001. Bird Conservation International, 13:273-281.
- HARRISON, P. 1983. Seabirds: An identification guide. Houghton Mifflin, Boston.
- HARRISON, P. 1987. Seabirds of the World: A photographic guide. Princeton University Press, Princeton.
- GRANTSAU, R. 1995. Os Albatrozes (Diomedeidae, Procellariiformes) do Atlântico e suas ocorrências na costa brasileira e uma chave de identificação. Boletim do Centro de Estudos Ornitológicos, 12:20-31.
- JOUANIN, C. & MOUGIN, J.L. 1979. Order Procellariiformes. In: Mayr, E. & Cottrel, G.W. (Eds.), Check-list of birds of the World 2nd ed. Harvard University Press, Cambridge, Massachusetts, v.1, p. 48-121.
- KENNEDY, M. & PAGE, R.D.M. 2002. Seabird Supertrees: Combining partial estimates of procellariiform phylogeny. The Auk, 119:88-108.
- LINNAEUS, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species cum characteribus, differentiis, synonymis, locis Lautentii Salvii, Stockholm.
- MATHEWS, G.M. 1934. Remarks on albatrosses and mollymawks. Ibis, 13:807-816.
- MATHEWS, G.M. 1948. Systematic notes on the petrels. Bulletin of the British Ornithologists Club, 68:155-170.
- MATHEWS, G.M. & HALLSTROM, J.L. 1943. [Apud Nunn, G.B. et al]. 1996. Notes on the order Procellariiformes The Federal Capital Press of Australia Limited, Canberra.
- MURPHY, R.C. 1917. A new albatross from the west coast of South America. Bulletin of the American Museum of Natural History, 37:861-864.
- MURPHY, R.C. 1936. Oceanic birds of South America MacMillan, New York.
- NEVES, T. DA S.; OLMOS, F. & PEPPES, F.V. 2003. Plano de Ação Nacional para a Conservação de Albatrozes e Petréis. Available at: <http://www.projetoalbatroz.com.br>.
- NUNN, G.B.; COOPER, J.; JOUVENTIN, P.; ROBERTSON, C.J.R. & ROBERTSON G.G. 1996. Evolutionary relationships among extant albatrosses (Procellariiformes: Diomedeidae) established from complete cytochrome-b gene sequences. The Auk, 113:784-801.
- PENHALLURICK, J. & WINK, M. 2004. Analysis of the taxonomy and nomenclature of the Procellariiformes based on complete nucleotide sequences of the mithocondrial cytochrome-b gene. Emu, 104:125-147.
- PINTO, O.M. de o. 1938. Catálogo das Aves do Brasil e Lista dos Exemplares que as representam no Museu Paulista. Revista do Museu Paulista, tomo XXII.
- PYCRAFT, W.P. 1899. Contributions to the Osteology of Birds. Part III. Tubinares. Proceedings of the Zoological Society of London, 1899: 381-411.
- REICHENBACH, L. 1853. Avium Systema Naturelle Leipzig, Germany.
- ROBERTSON, C.J.R. & NUNN, G.B. 1998. Towards a new taxonomy for the albatrosses. In: Robertson, G. & Gales, R.(Eds.), Albatross Biology and Conservation Surrey Beatty & Sons, Chipping Norton, NSW, Australia, p.13-19.
- TUCK, G.N.; POLACHECK, T. & BULMAN, C.M. 2003. Spacio-temporal trends of longline fishing effort in the Southern Ocean and implications for seabird bycatch. Biological Conservation, 114:1-27.
- WILKINSON, H.E. 1969. Description of an Upper Miocene albatross from Beaumaris, Victoria, Australia, and a review of the fossil Diomedeidae. Memoirs of the National Museum of Victoria, 29:41-51.
- ZUSI, R.L. & LIVEZEY, B.C. 2000. Homology and phylogenetic implications of some enigmatic cranial features in galliform and anseriform birds. Annals of Carnegie Museum, 69:157-193.
Publication Dates
-
Publication in this collection
05 July 2007 -
Date of issue
2007
History
-
Accepted
13 Mar 2007 -
Received
21 Nov 2006