Abstracts
A growing number of biogeographical methods have attempted to describe formal means of reconstructing the biogeographical history of the organisms. Whatever the biogeographical method, however, the source of systematic information has to be well worked out. Taxonomic noise is sometimes a true impediment to properly deal with the complexity of life in its three-dimensional aspects, the threefold parallelism represented by form, space and time. This paper argues that historical systematics is a necessary basis for a historical biogeography. Organismal phylogenies or at least hypotheses of monophyly should be taken as the basis for the study of distribution patterns. Whenever a non-monophyletic taxon is misleadingly taken as monophyletic, erroneous interpretations in evolutionary analyses necessarily follow. When the proportion of paraphyletic taxa considered in an analysis is small, a general pattern may be obtained, but the interpretation of the biogeographical evolution of each paraphyletic taxon will be equivocated. The delimitation of areas of endemism also depends on the precision of the recovered phylogenetic information. Indices based on phylogenetic diversity allow the delimitation of areas for conservation of biological diversity. Despite the plethora of current available biogeographical methods, biogeography is not a mess, as was pointed elsewhere. The order in the discipline is subtle: as biogeography intends to comprehend the living world based on the study of the form, space and time, a phylogenetic framework is a basic requirement. The lack of reliable biogeographical primary information - historical taxa - certainly creates severe obstacles for historical biogeography.
Biogeography; cladistics; conservation; endemism; monophyly; phylogeny; species
Um crescente número de métodos biogeográficos tem buscado descrever maneiras formais de reconstruir a história biogeográfica dos organismos. Entretanto, para qualquer método biogeográfico empregado, a fonte de informação sistemática deve ser precisa. Ruído taxonômico é por vezes um verdadeiro obstáculo para se tratar apropriadamente da complexidade da vida no seu aspecto tridimensional, representado pelo triplo paralelismo forma, espaço e tempo. Esse artigo defende que a sistemática é o fundamento necessário para a biogeografia histórica. Filogenias de organismos ou ao menos hipóteses de monofiletismo devem ser a base para o estudo de padrões de distribuição. Táxons não-monofiléticos tomados erroneamente como monofiléticos resultarão em interpretações incorretas nas análises evolutivas. Quando a proporção de táxons parafiléticos considerada em uma análise é pequena, um padrão geral pode ser obtido, mas a interpretação da evolução biogeográfica de cada táxon parafilético será equivocada. A delimitação de áreas de endemismo, da mesma forma, também depende da precisão da informação filogenética. Além disso, índices baseados na diversidade filogenética permitem a delimitação de áreas para a conservação da diversidade biológica. Apesar da pletora de métodos biogeográficos correntes, a biogeografia não é uma confusão, como foi apontado anteriormente. A ordem nessa disciplina é sutil: como a biogeografia pretende compreender o mundo natural baseandose no estudo de forma, tempo e espaço, um arcabouço filogenético é condição essencial. A ausência de informação biogeográfica primária confiável - táxons históricos - cria sérios obstáculos para a biogeografia histórica.
Biogeografia; cladística; conservação; endemismo; espécies; filogenia; monofiletismo
Why biogeographical hypotheses need a well supported phylogenetic framework: a conceptual evaluation
Charles Morphy D. Santos1 1 E-mail: charlesmorphy@pg.ffclrp.usp.br ; Dalton S. Amorim2 2 E-mail: dsamorim@usp.br
Universidade de São Paulo, Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, Departamento de Biologia, Setor de Ecologia e Evolução, Avenida Bandeirantes, 3900, 14040-901, Ribeirão Preto, SP, Brasil
ABSTRACT
A growing number of biogeographical methods have attempted to describe formal means of reconstructing the biogeographical history of the organisms. Whatever the biogeographical method, however, the source of systematic information has to be well worked out. Taxonomic noise is sometimes a true impediment to properly deal with the complexity of life in its three-dimensional aspects, the threefold parallelism represented by form, space and time. This paper argues that historical systematics is a necessary basis for a historical biogeography. Organismal phylogenies or at least hypotheses of monophyly should be taken as the basis for the study of distribution patterns. Whenever a non-monophyletic taxon is misleadingly taken as monophyletic, erroneous interpretations in evolutionary analyses necessarily follow. When the proportion of paraphyletic taxa considered in an analysis is small, a general pattern may be obtained, but the interpretation of the biogeographical evolution of each paraphyletic taxon will be equivocated. The delimitation of areas of endemism also depends on the precision of the recovered phylogenetic information. Indices based on phylogenetic diversity allow the delimitation of areas for conservation of biological diversity. Despite the plethora of current available biogeographical methods, biogeography is not a mess, as was pointed elsewhere. The order in the discipline is subtle: as biogeography intends to comprehend the living world based on the study of the form, space and time, a phylogenetic framework is a basic requirement. The lack of reliable biogeographical primary information - historical taxa - certainly creates severe obstacles for historical biogeography.
Keywords: Biogeography, cladistics, conservation, endemism, monophyly, phylogeny, species.
RESUMO
Um crescente número de métodos biogeográficos tem buscado descrever maneiras formais de reconstruir a história biogeográfica dos organismos. Entretanto, para qualquer método biogeográfico empregado, a fonte de informação sistemática deve ser precisa. Ruído taxonômico é por vezes um verdadeiro obstáculo para se tratar apropriadamente da complexidade da vida no seu aspecto tridimensional, representado pelo triplo paralelismo forma, espaço e tempo. Esse artigo defende que a sistemática é o fundamento necessário para a biogeografia histórica. Filogenias de organismos ou ao menos hipóteses de monofiletismo devem ser a base para o estudo de padrões de distribuição. Táxons não-monofiléticos tomados erroneamente como monofiléticos resultarão em interpretações incorretas nas análises evolutivas. Quando a proporção de táxons parafiléticos considerada em uma análise é pequena, um padrão geral pode ser obtido, mas a interpretação da evolução biogeográfica de cada táxon parafilético será equivocada. A delimitação de áreas de endemismo, da mesma forma, também depende da precisão da informação filogenética. Além disso, índices baseados na diversidade filogenética permitem a delimitação de áreas para a conservação da diversidade biológica. Apesar da pletora de métodos biogeográficos correntes, a biogeografia não é uma confusão, como foi apontado anteriormente. A ordem nessa disciplina é sutil: como a biogeografia pretende compreender o mundo natural baseandose no estudo de forma, tempo e espaço, um arcabouço filogenético é condição essencial. A ausência de informação biogeográfica primária confiável - táxons históricos - cria sérios obstáculos para a biogeografia histórica.
Palavras-chave: Biogeografia, cladística, conservação, endemismo, espécies, filogenia, monofiletismo.
INTRODUCTION
Biogeography is a field with a long, rich and often controversial intellectual heritage, extending back to the very first studies of the natural world (Nelson,1978; Nelson & Platnick, 1981; Mayr, 1982; Humphries & Parenti, 1999; Papavero et al., 2000; Nelson & Ladiges, 2001; Crisci et al., 2003; Lieberman, 2003; Williams & Ebach, 2004; Heads, 2005b). The essential goals of the discipline can be synthesized in two main points, the discovery of the patterns of spatial distribution attained by life on earth - the manner biological groups are displayed on world's geography - and the means by which this distribution was achieved - both in terms of mechanisms and processes (Platnick & Nelson, 1978; Rosen, 1978; Ebach & Humphries, 2003; Ebach et al., 2003; McDowall, 2004). In order to answer the fundamental question "Why are taxa distributed where they are today?" (Platnick & Nelson, 1978:1), for centuries biogeographers have attempted to establish the paths that led to the current spatial distribution of life on earth's surface.
Traditionally, biogeography has been divided in ecological and historical biogeography. Ecological biogeography is often restricted to groups of lower taxonomic rank and smaller geographical areas, and is concerned with processes such as climate or other physical or environmental factors occurring over short temporal scales. Historical biogeography examines events and processes occurring over a long time span, based on causes that existed in the past and with consequences to large sets of organisms (for a review, see Morrone & Crisci, 1995). Such a division, however, has been recently challenged (Lieberman, 2003; Riddle, 2005). It is clear that patterns are neither wholly historical nor wholly ecological. At any time, ecological processes play an important role in shaping the effect of ongoing historical processes. In short, ecological biogeography is also historical, and vice-versa. Herein, biogeography will be always mentioned in its temporal, i.e. historical, perspective.
A well-known metaphor created by Croizat (1964) suggests that the diversity of life is a historical, three-dimensional phenomenon, the three elements being form (similarities and differences in the attributes of the organisms), space and time. The view that biogeography is concerned with the history of biological entities in space is quite widespread. However, the impossibility of fully understanding the biogeography without the other two components is, perhaps, not as well recognized, in spite of the claims that there is no such a thing as an absolute space - space is a relative concept, only comprehensible if considered mutually with the other two sides of Croizat's saying (Croizat, 1964, 1982; Craw et al., 1999; Crisci, 2001; Ghehan, 2001; and other panbiogeograhic studies). If earth and life really evolve together (Croizat, 1958), the patterns of biotic distribution and the connections between biotas are undisputable outputs of the evolutionary process.
Phylogenies are biological hierarchical patterns resulted from descent with modification through time (Hennig, 1966). Obviously, evolution takes place somewhere - life is evolving in space and time. As evolution means nothing without the spatial component, does any spatial component mean anything without evolution? As the natural world has an inherent hierarchy provided by descent with modification, we can use such biological hierarchical patterns in order to recognize historical spatial relationships. As pointed by many authors, particularly those working under a cladistic (or a vicariancist) approach (Nelson & Platnick, 1981; Humphries & Parenti, 1999; Santos, 2005), historical biogeography necessarily must be underlined by phylogenies of organisms to take for grant the consequences of common descent upon the distribution of life over the globe.
In the words of Nelson (1978:269), "biogeography is a strange discipline". This seems a widespread view: biogeography is frequently seen with skepticism and considered one of the most complex disciplines of comparative biology. It is time-consuming, deeply dependent on systematic information, and interrelated to a great sort of evidences provided by other branchs of science, such as geology, geography, paleontology, molecular biology, and ecology. Many things have been told about the apparent "mess of methods" into which the field is immersed, but the claims are not completely unfounded: it is not possible to infer complex histories solely using a straightforward, naïve biogeographical method. Areas, unlike most known taxa, are quite open systems and often have multiple and complicated histories, so there are no simple answers to explain biogeographical patterns (Funk, 2004). The enormous sequences of modifications on the face of the earth in the last 3.8 billion years, with emergence of new terranes, continental drift, coalescence, and fragmentation, and their implication for organismal distribution, with extinction, disjunction, and dispersal, turn "areas" into very complex entities. Crisci (2001:165) accurately synthesizes this intricacy when he says that "probably we will never see it [the biogeographical history] totally revealed".
From a methodological point of view, none of the presently available algorithms for biogeographical analysis are able to deal at the same time with the complete multivariate properties of real data that should be considered during a biogeographical analysis, like directional dispersal, multiple areas with single terminal branchs, failures in vicariance, redundant distributions (paralogy), missing areas caused by extinction, and so on. An alternative in absence of such a unified methodological tool is an integrative approach (Morrone & Crisci, 1995; Donoghue & Moore, 2003), with the application of different methods at each step of the analysis, taking advantages of the merits of each one. Notwithstanding, this approach is still too "loose" and quite close to compromise solutions. For instance, how to define which method is the best for a specific situation, since the answers are not known a priori? Is it possible to create a method that embraces all the multifaceted needs of biogeography in order to turn it into a full-blown autonomic discipline with its own principles? The answer is probably no - at least for the time being. Irrespective of the complexity of the field of research, there is additionally some degree of clash between divergent schools of biogeography. As Malte Ebach points in his debate with Juan Morrone (Ebach & Morrone, 2005:2181), "Biogeography is a field that cannot communicate with one language or one voice because there is a multitude of conflicting ideas and aims".
Anyway, a synthetic or a cut-and-dried recipe is not the solution. The conceptual framework for handling the distribution of organisms and events in space is the phylogenetic perspective. What must remain common to the whole biogeography-ground practice, no matter which kind of scientific heritage is being considered, is phylogeny. As Hennig (1966) pointed out (what has been considered a common place since Wallace's ideas (1858, 1876) about the origin of species), there is a close and recognizable relationship between a species and the space it occupies. The dictum is one of the foundations of modern historical biogeography, and led to the development of a whole new set of practices based on the idea that phylogenetic relationships are biogeographically informative, the so-called cladistic biogeography.
This article intends to advocate for the necessity of phylogenies for historical biogeographical methods, especially regarding the quality of taxonomic data used to recognize areas of endemism and the historical relationships among them. Without reliable delimitation of the groups under inquiry (whatever the taxonomic rank), it is impossible to obtain consistent biogeographical hypotheses. As a corollary, the quality of biogeographical databases is deeply relevant to the identification of priority areas for conservation, which emphasize the meaning of systematics to biogeography.
The value of biogeographical reconstructions relies entirely on available evidence supporting monophyletic lineages in which they are based upon. Thus, misleading information (i.e., non-historical taxa) is a major constraint for the reliable recovery of biogeographical history. Not only is a particular biogeographical reconstruction of non-existing biological entities foolish, but false taxa generate incorrect information in the quest for general biogeographical patterns. It is now well understood that the complexity of historical reconstructions makes any noise in raw data due to the inclusion of non-historical groups a severe handicap to the overall analysis, as we well see further in this essay. Hence, biogeographical tools - techniques, procedures, or even entire methods - should treat their subject under a strictly phylogenetic perspective. Any method or researcher that resists the notion that systematics is the basis for historical biogeography will be absorbed in many difficulties and misunderstanding in addition to those integral of any biogeographical analysis, already hard enough to deal with. Paraphrasing Nelson (1994), after Dobzhansky (1973), nothing in historical biogeography makes sense except in the light of systematics.
Previous organismal phylogenies, or at least hypotheses of monophyly of given taxa, should be taken as the basis for the study of distribution patterns - this statement could be seen as valid not just for cladistic biogeography but also for panbiogeography or any other historical biogeographical method. Once hypotheses of monophyly of different taxa are taken and analyzed together, the hypothesized relationship among the areas derived from the sister-group relationships can be corroborated and explained as a general pattern, abandoned in favor of an alternative explanation, or even considered as a unique event. Phylogenies therefore reveal what genealogical relationships say about the relationships between the areas the organisms occupy.
Monophyly and endemism
The comments above could be considered at first sight obvious and self-evident to biogeographers, but the present state of biogeography literature shows that they are not. The notion that paraphyletic taxa should be recognized and accepted in biological classifications, for example, is still quite pervasive, as can be noted from the manifesto organized by Nordal & Stedje (2005) and some other recent papers on the subject (Brummit, 2006; Hörandl, 2006). Actually, major problems would be minimized if some of the fundamentals presented in the first section of this article were taken for granted prior to biogeographical studies or methodological developments. Let us consider the species problem as an example of the need of sharp systematic tools for biogeography. As pointed by Wheeler & Meier (2000), no matter what kind of scientific responses are offered to the current biodiversity crisis, whether it is the establishment of what groups and how many organisms live on the planet nowadays or the comparison of relative diversity and relationships among two or more taxa or areas, they will always rely upon a general consensus about what a species is. The delimitation of the basic units of biological diversity affects estimates of species diversity, historical-evolutionary analysis of these units, understanding of patterns of gene flow within or among these units, delimitation of areas of endemism and so on (Cracraft, 1997, 2000). Systematic accuracy really does make an enormous difference in the way the natural world is described and understood.
In many cases species as taxa correspond to a set of populations not directly related even though grouped under the same name - sometimes referred to as "paraspecies" (Ackery & Vane-Wright, 1984), "metaspecies" (Donoghue, 1985; Mishler & Brandon, 1987), or "agamospecies" (e.g., Meier & Willmann, 2000; Willmann & Meier, 2000). In other words, names of species do not always correspond to real, historical systems. The inclusion of non-monophyletic species in classifications has been advocated in the literature (e.g., Crisp & Chandler, 1996), but we strongly disagree with this position from a conceptual perspective. Whenever a non-monophyletic taxon, at whatever rank, is incorrectly taken as a valid systematic unity under an evolutionary perspective (homology character evolution, biogeography, conservation), the performed analysis will be flawed.
This seems to be largely overlooked even in part of the scientific community due to the persistence of an idealistic or essentialist understanding of the nature of species. It is a widespread idea that when there is a species name in existing classifications, there is necessarily a corresponding real biological entity-rather than "conglomerations of two or more species [or populations] that we have not yet succeeded in diagnosing" (Wheeler & Platnick, 2000). Notwithstanding, biological classification is real science, and names in a classification are scientific hypotheses. Under some concepts of biological species (e.g., Wheeler & Platnick, 2000), true species without autapomorphies are actually conceivable. But this is not necessarily true the other way around: not all species level taxa without autapomorphies are real entities. Nevertheless, it is not the case to advocate here that individual organisms should be used as the terminal entities in cladistic analysis in order to distinguish monophyletic groups that can be considered real species, rather than supposed interbreeding "groups" of non-related entities, as proposed by Vrana & Wheeler (1992) and by some Phylocode adherents (for a review, see Pickett, 2005). This is an extreme view - the distinction of species is made for and prior to a cladistic analysis (Wheeler & Platnick, 2000).
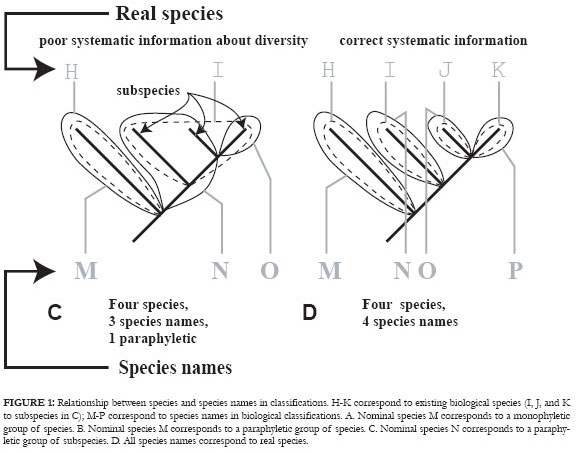
In fact, as stated above, many available species names actually refer to monophyletic groups comprising two or more different species (Figure 1A). Other names correspond to paraphyletic or polyphyletic groups of species (Figure 1B) or populations (Figure 1C). If all species names really corresponded to historical units, some biogeographical methods not based on previous phylogenies would be safe. But the real world is much trickier and the output of systematic activity (the classification at all levels) is subject to failure. For instance, Crisp & Chandler (1996) estimated that "the proportion of paraphyletic and metaphyletic species may be 50% or higher". If true, this would be a tragedy for group studies performed without a truly phylogenetic perspective and for non-phylogenetic methods in biogeography. The example of Drosophila serido (Manfrin & Sene, 2006) is illustrative as to how detailed and extensive systematic effort is necessary (in this case including even chromosomal features) to demonstrate that a "species" is paraphyletic, eliminating an enormous amount of misinformation (about character homology, larvae biology, biogeographical patterns, false jump dispersal events, etc.) prompted by a misconstrued taxon at the species level. The split of D. serido in five different species and the proposal of a phylogenetic hypothesis including all new species and D. borborema, the sister-group of D. serido stricto sensu, allowed a new view of the biogeographical relationships among the components of this group and its evolution (Figure 2).
Some authors, as Rage & Jaeger (1995), understood that paraphyletic taxa can be useful in biogeographical studies, "the only deficiency" being the missing part of the taxa! It is exactly because, unlike monophyletic groups, paraphyletic and polyphyletic taxa do not represent complete hierarchical patterns of character distribution that they are sources of error. The inclusion of such non-monophyletic entities as primary data in biogeographical analyses results in an incorrect depiction of the historical hierarchy among areas (for supraspecific taxa) or an incorrect delimitation of species distribution (for the species level). Monophyletic groups are necessary because they reflect the natural affinity of whatever kinds of biological entities are considered, and the discovery of such natural affinities - i.e. common descent - depends on phylogenetic scrutiny.
Areas of endemism and methods
The phylogenetic perspective in biogeography is also valuable in the conservation effort. Indices based on phylogenetic diversity provide a measure of how species diversity is distributed among monophyletic groups, and allow the scientific delimitation of areas for biological diversity conservation (Nixon & Wheeler, 1992; Wheeler, 1995; Wheeler & Platnick, 2000). Figure 3 presents a hypothetical example (based on a previous discussion by Wheeler & Platnick, 2000) of how such knowledge of phylogenetic diversity is essential to the politics concerning the maintenance of species diversity. Even though there is an equal number of species in both areas A and B, the phylogenetic background clearly shows that the conservation of area B should be prioritized - it has a large number of distantly related monophyletic groups and, hence, more biological diversity, when compared to area A, which contains closely related species of a single monophyletic group.
As species are the basic unities in systematics, so areas of endemism are the basic unities in biogeography. However, despite the crucial role of the concept of areas of endemism in biogeography, since the works of de Candolle (1820) endemism has been defined in a vague manner and the criteria used to identify such areas, far from being well-established, have been developed over centuries (Nelson, 1978; Nelson & Platnick, 1981; Anderson, 1994; Humphries & Parenti, 1999; Hausdorf, 2002; Szumik et al., 2002; Hausdorf & Hennig, 2003; Szumik & Goloboff, 2004). An "area of endemism" means many things to many people, depending on their background and purpose of investigation. Nevertheless, the delimitation of areas of endemism is the very first procedure in many biogeographical studies. How to proceed in this situation? An area of endemism is not an area of distribution (Santos, 2005) and its recognition is much more complex than merely plotting taxonomic occurrences on a map. This position is in accordance to that advocated by Harold & Mooi (1994), to whom an area of endemism is a true historical unity, and to define it accurately biogeographers should necessarily take history into account. Distributional patterns can be considered sources of biogeographical information only in the context of the genealogical relationships within the inhabiting organisms.
As stated by Harold & Mooi (1994), an area of endemism is a geographic region in which are distributed two or more monophyletic taxa that exhibit phylogenetic and distributional congruence, with their respective relatives occurring in other such defined regions. According to them, the delimitation of an area of endemism is a two-step approach: (1) a basic taxonomic/distributional level analysis in which the taxa are chosen, their relationships hypothesized and their distributions delimited; and (2) a testing phase, in which other groups are inserted into the analysis in order to corroborate (or refute) these distributions as indicating areas of endemism, and to test the historical reality of those areas. In short, the establishment of an area of endemism is unable to accomplish without the determination of spatially congruent monophyletic entities.
Historical biogeographical methods aim to (1) recognize spatial homology, (2) identify areas of endemism, (3) formulate hypotheses about area relationships, and (4) recognize the role of past and present events causing spatial distributions (Morrone & Crisci, 1995; McDowall, 2004; Santos, 2005). Among biogeographical procedures, Parsimony Analysis of Endemicity (or PAE) (Rosen, 1988) has been used to delimit areas of endemism based on their shared taxa according to the most parsimonious solution. Sampled localities, usually quadrats drawn over a map, are taken as analogous to shared derived characters (synapomorphies) in phylogenetic analyses. Shared areas in the distribution of species are assumed to indicate shared unique biological history (Rosen, 1988; Rosen & Smith, 1988; Morrone & Crisci, 1995; Brooks & van Veller, 2003). To PAE, phylogenetic relationships of groups are not relevant to the construction of area relationships. PAE is a method that intends to identify areas of endemism and the relationships among them but, as a non-phylogenetic reconstruction procedure, it is unable to distinguish whether an area is historically more closely related to another with regard to a third one (contra, see Nihei, 2006).
According to Morrone (1994), PAE is a helpful tool to establish areas of endemism. However, PAE users only plot taxonomic occurrence on a map in search for congruent distributional patterns of two or more species. Checking congruence in spatial distribution is a trivial procedure and does not provide any sound evidence to determine natural or historical boundaries among the areas of endemism. The recognition of the spatial distribution of a historical entity is not the same thing as the definition of an area of endemism (Harold & Mooi, 1994; Santos, 2005).
Szumik et al. (2002) and Szumik & Goloboff (2004) recognized the correctness of Harold & Mooi's (1994) definition, but pointed out the operational difficulties of the criteria proposed by them, and suggested an optimally criterion that provides a way to assign a score of endemicity to a given area regardless of how that area was firstly hypothesized. As said before, to study the history of connections and disjunctions between areas is a difficult task because even fundamental biogeographical patterns such as endemism are easily obscured. Despite the methodological difficulties, it is quite clear that it is not possible to properly depict the boundaries of areas of endemism without a phylogenetic perspective to correctly identify monophyletic groups. In a brief commentary, Knapp (2005) raised two fundamental benchmarks about biogeography: a careful taxonomic work, and a good definition of areas of endemism. Further: "without good species delimitation and definition, the terminals used in the cladograms or phylogenies are meaningless and without good phylogenetic hypotheses a scientist cannot relate the history of distribution to the history of the organisms" (Knapp, 2005:4).
The same problem of lack of historical connection - in other words, of evolutionary connection - between specimens, populations and species is seen in phylogeography. The method was originally proposed by Avise et al. (1987) and can be defined as "the study of the principles and processes governing the geographic distribution of genealogical lineages at intraspecic level using mitochondrial DNA (mtDNA) in animals and chloroplast DNA (cpDNA) in plants" (Posadas et al., 2006:397). Phylogeography supposedly intends to understand the distributional history of populations and to depict the history of areas in which they inhabit. In phylogeography, individuals are genotyped and assigned to maternal lineages and the resulting branched diagram is related to patterns of geographic distribution. This approach assumes the possibility of vicariance and extinction, but attempts to emphasize the role of dispersal in shaping distributional patterns (Ebach & Humphries, 2002; Morrone, 2002). The definition is alright and it would be nice if the algorithm producing the "phylograms" and biogeographical explanations actually generated evolutionary connections. It is naïve to suppose that because the work is below the species level, the connections between the individuals would be of some other kind. If the connections are not historical (i.e., a phylogeny), what connects individuals and populations? The answer is straightforward: similarity. The fact that the source of data is molecular does not make the problem different from that of morphology. The systematic literature of the 1970s and 1980s has well demonstrated that "similarity" is not a positive indication of kinship relations. Some authors criticize phylogeography considering it as a restricted and blinkered approach in a complex multifaceted discipline (biogeography) which fails to consider its historical context (e.g., Ebach et al., 2003). From a biogeographical point of view, Heads (2005a) is even more incisive in claiming that the whole theory and practice of phylogeography is not centered on hypotheses testing but on reiterating the dispersalistic view of Matthew (1915), of the so-called New York School of Zoogeography (Nelson & Ladiges, 2001), and the phylogenetic biogeography, based on centre of origin and the progression rule (Heads, 2005a; Santos, in press).
On the need for systematics
Nowadays, governmental institutions and professional organizations responsible for developing environmental policies to define areas of conservation have increasingly valued biogeographical studies. Delimitation of areas of endemism, evaluation of diversity and recognition of areas of interface between biomes - typical academic outputs of biogeographical analyses in the past - are now seen as important sources of information for the creation of databases used in the delimitation and maintenance of conservation units. Discussions and debates on biogeography, much more than simple quarrels about semantics or scientific idiosyncrasies, run out of the lab and reach a part of society that is not aware of what goes on within academic walls. There has been some pressure over taxonomy for faster production of species names, with criticisms to whatever could retard this activity - the so-called taxonomic impediment (Carvalho et al., 2005). But of what use is poor systematics for conservation if it results in unreliable source of information? Systematics cannot compromise itself by furnishing such untrustworthy data.
There is a need to put order into biogeography. This does not mean biogeographers should unifying in a single method the current plethora of theories, ideas and procedures of the discipline, all of them with different scopes and intentions. The multiplicity of ways of thinking in biogeography is absolutely salutary to the enrichment of our knowledge of the natural world and to guide our actions toward its protection. The order in the field is more subtle: every biogeographical study requires a phylogenetic framework in order to properly describe aspects of the living world based on the study of the triumvirate space, form and time. As said by Gareth Nelson in a lecture in 1969 (reproduced in Williams & Ebach, 2004:710): "The Recent biota is not simply life, but life ordered and diversified by time and space". During the history of biological systematics, especially after the Darwin-Wallace revolution of the 19th century, there was a search for a method that would truly discover the history of the natural affinities among the organisms in their classifications. After the excited debates in the 1970s and 1980s on the nature of the phylogenetic inquiry, it seems now to be the established paradigm that Hennig's approach is such a method, and phylogenies are the reference systems of comparative biology. Likewise, historical biogeographers who believe that their field is a part of a broad evolutionary research program should consider phylogenies as the reference system to all biogeographical studies. Methods that do not take into account monophyly and congruence among phylogenies of taxa of similar distribution at whatever taxonomic level (as PAE, some phylogeographical procedures or even panbiogeography made without reliable systematic work) will naturally perish.
ACKNOWLEDGMENTS
The authors would like to thank Guilherme Ribeiro for his comments and suggestions. Dr. Malte Ebach, Dr. Peter Cranston and two anonymous referees recommended substantial improvements to the manuscript, for which we are sincerely grateful. Research was supported by CAPES and FAPESP (process 03/12074-9).
REFERENCES
ACKERY, R.R. & VAN-WRIGHT, R.I. 1984. Milkweed butterflies: their cladistic and biology. Cornell University Press, Ithaca, NY.
ANDERSON, S. 1994. Area and endemism. The quarterly review of biology, 69:451-471.
AVISE, J.C.; ARNOLD, J.; BALL, R.M.; BERMING-HAM, E.; LAMB, T.; NEGEL, J.E.; REEB, C.A. & SAUDANERS, N.C. 1987. Intraspecific phylogeography: the mitochondrial DNA bridge between population genetic and systematics. Annual Review of Ecology and Systematics, 18:489-522.
BROOKS, D.R. & VAN VELLER, M.G.P. 2003. Critique of parsimony analysis of endemicity as a method of historical biogeography. Journal of Biogeography, 30:819-825.
BRUMMIT, R.K. 2006. Am I a bony fish? Taxon, 55:268-269.
DE CANDOLLE, A.P. 1820. Essai élémentaire de géographie botanique. In: Dictionaire des sciences naturelles. Flevrault, Strasbourg, v.18.
CARVALHO, M.R.; BOCKMANN, F.A.; AMORIM, D.S.; DE VIVO, M.; DE TOLEDO-PIZA, M.; MENEZES, N.A.; DE FIGUEIREDO, J.L.; CASTRO, R.M.C.; GILL, A.C.; MCEACHRAN, J.D.; COMPAGNO, L.J.V.; SCHELLY, R.C.; BRITZ, R.; LUNDBERG, J.G.; VARI, R.P. & NELSON, G. 2005. Revisiting the taxonomic impediment. Science, 307:353.
CRACRAFT, J. 1997. Species concept in systematics and conservation biology: an ornithological viewpoint. In: Claridge, M.A., Dawah, H.A. & Wilson, M.R. (Eds.), Species: the units of diversity. Chapman & Hall, London, p.325-339.
CRACRAFT, J. 2000. Species concept in theoretical and applied biology: a systematic debate with consequences. In: Wheeler, Q.D. & Meier, R. (Eds.), Species concepts and phylogenetic theory: a debate. Columbia University Press, New York, p.3-14.
CRAW, R.C.; GREHAN, J.R. & HEADS, M.J. 1999. Panbiogeography: tracking the history of life. Oxford University Press, New York.
CRISCI, J.V. 2001. The voice of historical biogeography. Journal of Biogeography, 28:157-168.
CRISCI, J.V.; KATINAS, L. & POSADAS, P. 2003. Historical biogeography: an introduction. Harvard University Press, Cambridge.
CRISP, M.D. & CHANDLER, G.T. 1996. Paraphyletic species. Telopea, 6:813-844.
CROIZAT, L. 1958. Panbiogeography. Published by the author, Caracas.
CROIZAT, L. 1964. Space, time, form: the biological synthesis. Published by the author, Caracas.
CROIZAT, L. 1982. Vicariance, vicariism, panbiogeography, vicariance biogeography, etc. a clarification. Systematic Zoology, 31:291-304.
DOBZHANSKY, T. 1973. Nothing in biology makes sense except in the light of evolution. American Biology Teacher, 35:125-129.
DONOGHUE, M.J. 1985. A critique of the biological species concept and recommendations for a phylogenetic alternative. Bryologist, 88:172-181.
DONOGHUE, M.J.. & MOORE, B.R. 2003. Toward an integrative historical biogeography. Integrative and Comparative Biology, 43:261-270.
EBACH, M.C. & HUMPHRIES, C.J. 2002. Cladistic biogeography and the art of discovery. Journal of Biogeography, 29:427-444.
EBACH, M.C. & HUMPHRIES, C.J. 2003. Ontology of biogeography. Journal of Biogeography, 30:959-962.
EBACH, M.C. & HUMPHRIES, C.J.; & WILLIAMS, D.M. 2003. Phylogenetic biogeography deconstructed. Journal of Biogeography, 30:1385-1396.
EBACH, M.C. & MORRONE, J.J. 2005. Forum on historical biogeography: what is cladistic biogeography? Journal of Biogeography, 32:2179-2187.
FUNK, V.A.. 2004. Revolutions in historical biogeography. In: Lomolino, M.V.; Sax, D.F. & Brown, J.H. (Eds.), Foundations of biogeography: classic papers with commentaries. University of Chicago Press, Chicago and London, p.647-657.
GREHAN, J.H. 2001. Panbiogeografía y la geografía de la vida. In: Llorente, J. & Morrone, J.J. (Eds.), Introducción a la biogeografía en Latinoamérica: teorías, conceptos, métodos y aplicaciones. Las Prensas de Ciencias, Facultad de Ciencias, México, p.181-195.
HÖRANDL, E. 2006. Paraphyletic versus monophyletic taxa-evolutionary versus cladistic classifications. Taxon, 55:564-570.
HAROLD, A.S. & MOOI, R.D. 1994. Areas of endemism: definition and recognition criteria. Systematic Biology, 43:261-266.
HAUSDORF, B. 2002. Units in biogeography. Systematic Biology, 51:648-651.
HAUSDORF, B. & HENNING, C. 2003. Biotic element analysis in biogeography. Systematic Biology, 52:717-723.
HEADS, M. 2005a. Dating nodes on molecular phylogenies: a critique of molecular biogeography. Cladistics, 21:62-78.
HEADS, M. 2005b. The history and philosophy of panbiogeography. In: Morrone, J.J. & Llorente, J. (Eds.), Regionalización Biogeográfica en Iberoamérica y Tópicos Afines. Universidad Nacional Autónoma de México, México, p.67-123.
HENNIG, W. 1966. Phylogenetic systematics. University of Illinois Press, Urbana.
HUMPHRIES, C.J. & PARENTI, L.R. 1999. Cladistic biogeography: interpreting patterns of plant and animal distributions. 2nd edition. Oxford University Press, Oxford.
KNAPP, S. 2005. Biogeography: space, form and time. Journal of Biogeography, 32:3-4.
LIEBERMAN, B.S. 2003. Unifying theory and methodology in biogeography. Evolutionary Biology, 33:1-25.
MATTHEW, W.D. 1915. Climate and evolution. Annals of the New York Academy of Sciences, 24:171-318.
MAYR, E. 1982. The growth of biological thought. Harvard University Press, Cambridge.
MANFRIN, M.H. & SENE, F.M. 2006. Cactophilic Drosophila in South America: a model for evolutionary studies. Genetica, 126:57-75.
MCDOWALL, R.W. 2004. What biogeography is: a place for process. Journal of Biogeography, 31:345-351.
MEIER, R. & WILLIAMANN, R. 2000. The Hennigian species concept. In: Wheeler, Q.D. & Meier, R. (Eds.), Species concepts and phylogenetic theory: a debate. Columbia University Press, New York, p.30-43.
MISHLER, B.D. & BRNADON, R.N. 1987. Individuality, pluralism, and the phylogenetic species concept. Biology & Philosophy, 2:397-414.
MORRONE, J.J. 1994. On the identification of areas of endemism. Systematic Biology, 43:438-441.
MORRONE, J.J. 2002. El espectro del dispersalismo: de los centros de origen a las áreas ancestrales. Revista de la Sociedad Entomológica Argentina, 61:1-14.
MORRONE, J.J. & CRISCI, J.V. 1995. Historical biogeography: introduction to methods. Annual Review of Ecology and Systematics, 26:373-401.
NELSON, G. 1978. From Candolle to Croizat: comments on the history of biogeography. Journal of Historical Biology, 11:269-305.
NELSON, G. 1994. Homology and systematics. In: Hall, B.K. (Ed.), Homology: the hierarchical basis of comparative biology. Academic Press, San Diego, p.101-149.
NELSON, G.& LADIGES, P.Y. 2001. Gondwana, vicariance biogeography and the New York School revisited. Australian Journal of Botany, 49:389-409.
NELSON, G.& PLATNICK, N. 1981. Systematics and biogeography: cladistics and vicariance. Columbia University Press, New York.
NIHEI, S.S. 2006. Misconceptions about parsimony analysis of endemicity. Journal of Biogeography, 33:2099-2106.
NIXON, K.C. & WHEELER, Q.D. 1992. Measures of phylogenetic diversity. In: Novacek, M.J. & Wheeler, Q.D. (Eds.), Extinction and phylogeny. Columbia University Press, New York, p.216-234.
NORDAL, I. & STEDJE, B. 2005. Paraphyletic taxa should be accepted. Taxon, 54:5-6.
PAPAVERO, N.; LLORENTE-BOUSQUETS, J.; ORGANISTA, D.E. & MASCARENHAS, R. 2000. História da biologia comparada: desde o Gênesis até o fim do Império Romano do Ocidente. Holos Editora, Ribeirão Preto.
PICKETT, K.M. 2005. The new and improved PhyloCode, now with types, ranks, and even polyphyly: a conference report from the First International Phylogenetic Nomenclature Meeting. Cladistics, 21:79-82.
PLATNICK, N. & NELSON, G. 1978. A method of analysis for historical biogeography. Systematic Zoology, 27:1-16.
POSADAS, P.; CRISCI, J.V. & KATINAS, L. 2006. Historical biogeography: A review of its basic concepts and critical issues. Journal of Arid Environments, 66:389-403.
RAGE, J.C. & JAEGER, J.J. 1995. The sinking Indian raft: a response to Thewison and McKenna. Systematic Biology, 44:260-264.
RIDDLE, B.R. 2005. Is biogeography emerging from its identity crisis? Journal of biogeography, 32:185-186.
ROSEN, B.R. 1988. From fossils to earth history: applied historical biogeography. In: Myers, A.A. & Giller, P.S. (Eds.), Analytical biogeography: an integrated approach to the study of animal and plant distributions. Chapman and Hall, London, p.437-481.
ROSEN, B.R. & SMITH, A.B. 1988. Tectonics from fossils? Analysis of reef-coral and sea-urchin distributions from late Cretaceous to Recent, using a new method. In: Audley-Charles, M.G. & Hallam, A. (Eds.), Gondwana and Tethys. Série Geological Society Special Publication, n.37. Oxford University Press, Oxford, p.275-306.
ROSEN, D.E. 1978. Vicariant patterns and historical explanation in biogeography. Systematic Zoology, 27:159-188.
SANTOS, C.M.D. 2005. Parsimony analysis of endemicity: time for an epitaph? Journal of Biogeography, 32:1284-1286.
SANTOS, C.M.D. in press [2007]. On basal clades and ancestral nodes. Journal of Biogeography.
SZUMIK, C.A.; CUEZZO, F.; GOLOBOFF, P.A. & CHALUP, A.E. 2002. An optimally criterion to determine areas of endemism. Systematic Biology, 51:806-816.
SZUMIK, C.A. & GOLOBOFF, P.A. 2004. Areas of endemism: an improved optimally criterion. Systematic Biology, 53:968-977.
VRANA, P. & WHEELER, W. 1992. Individual organisms as terminal entities: laying the species problem to rest. Cladistics, 8:67-72.
WALLACE, A.R. 1858. On the tendency of varieties to depart indefinitely from the original type. Proceedings of the Linnean Society of London, 3:53-62.
WALLACE, A.R.1876. The geographical distribution of animals. Macmillan, London.
WHEELER, Q.D. 1995. Systematics, the scientific basis for inventories of biodiversity. Biodiversity and Conservation, 4:476-489.
WHEELER, Q.D. & MEIER, R. (eds.). 2000. Species concepts and phylogenetic theory: a debate. Columbia University Press, New York.
WHEELER, Q.D. & PLATNICK, N. 2000. The phylogenetic species concept (sensu Wheeler and Platnick). In: Wheeler, Q.D. & Meier, R. (Eds.), Species concepts and phylogenetic theory: a debate. Columbia University Press, New York, p.55-69.
WILLIAMS, D.M. & EBACH, M.C. 2004. The reform of palaeontology and the rise of biogeography : 25 years after 'ontogeny, phylogeny, paleontology and the biogenetic law' (Nelson, 1978). Journal of Biogeography, 31:685-712.
WILLMANN, R. & MEIER, R. 2000. A critique from the Hennigian species concept perspective. In: Wheeler, Q.D. & Meier, R. (Eds.), Species concepts and phylogenetic theory: a debate. Columbia University Press, New York, p.101-118.
Recebido em: 09.11.2006
Aceito em: 05.04.2007
Impresso em: 15.06.2007
- ACKERY, R.R. & VAN-WRIGHT, R.I. 1984. Milkweed butterflies: their cladistic and biology. Cornell University Press, Ithaca, NY.
- ANDERSON, S. 1994. Area and endemism. The quarterly review of biology, 69:451-471.
- AVISE, J.C.; ARNOLD, J.; BALL, R.M.; BERMING-HAM, E.; LAMB, T.; NEGEL, J.E.; REEB, C.A. & SAUDANERS, N.C. 1987. Intraspecific phylogeography: the mitochondrial DNA bridge between population genetic and systematics. Annual Review of Ecology and Systematics, 18:489-522.
- BROOKS, D.R. & VAN VELLER, M.G.P. 2003. Critique of parsimony analysis of endemicity as a method of historical biogeography. Journal of Biogeography, 30:819-825.
- BRUMMIT, R.K. 2006. Am I a bony fish? Taxon, 55:268-269.
- DE CANDOLLE, A.P. 1820. Essai élémentaire de géographie botanique. In: Dictionaire des sciences naturelles. Flevrault, Strasbourg, v.18.
- CARVALHO, M.R.; BOCKMANN, F.A.; AMORIM, D.S.; DE VIVO, M.; DE TOLEDO-PIZA, M.; MENEZES, N.A.; DE FIGUEIREDO, J.L.; CASTRO, R.M.C.; GILL, A.C.; MCEACHRAN, J.D.; COMPAGNO, L.J.V.; SCHELLY, R.C.; BRITZ, R.; LUNDBERG, J.G.; VARI, R.P. & NELSON, G. 2005. Revisiting the taxonomic impediment. Science, 307:353.
- CRACRAFT, J. 1997. Species concept in systematics and conservation biology: an ornithological viewpoint. In: Claridge, M.A., Dawah, H.A. & Wilson, M.R. (Eds.), Species: the units of diversity. Chapman & Hall, London, p.325-339.
- CRACRAFT, J. 2000. Species concept in theoretical and applied biology: a systematic debate with consequences. In: Wheeler, Q.D. & Meier, R. (Eds.), Species concepts and phylogenetic theory: a debate. Columbia University Press, New York, p.3-14.
- CRAW, R.C.; GREHAN, J.R. & HEADS, M.J. 1999. Panbiogeography: tracking the history of life. Oxford University Press, New York.
- CRISCI, J.V. 2001. The voice of historical biogeography. Journal of Biogeography, 28:157-168.
- CRISCI, J.V.; KATINAS, L. & POSADAS, P. 2003. Historical biogeography: an introduction. Harvard University Press, Cambridge.
- CRISP, M.D. & CHANDLER, G.T. 1996. Paraphyletic species. Telopea, 6:813-844.
- CROIZAT, L. 1958. Panbiogeography. Published by the author, Caracas.
- CROIZAT, L. 1964. Space, time, form: the biological synthesis. Published by the author, Caracas.
- CROIZAT, L. 1982. Vicariance, vicariism, panbiogeography, vicariance biogeography, etc. a clarification. Systematic Zoology, 31:291-304.
- DOBZHANSKY, T. 1973. Nothing in biology makes sense except in the light of evolution. American Biology Teacher, 35:125-129.
- DONOGHUE, M.J. 1985. A critique of the biological species concept and recommendations for a phylogenetic alternative. Bryologist, 88:172-181.
- DONOGHUE, M.J.. & MOORE, B.R. 2003. Toward an integrative historical biogeography. Integrative and Comparative Biology, 43:261-270.
- EBACH, M.C. & HUMPHRIES, C.J. 2002. Cladistic biogeography and the art of discovery. Journal of Biogeography, 29:427-444.
- EBACH, M.C. & HUMPHRIES, C.J. 2003. Ontology of biogeography. Journal of Biogeography, 30:959-962.
- EBACH, M.C. & HUMPHRIES, C.J.; & WILLIAMS, D.M. 2003. Phylogenetic biogeography deconstructed. Journal of Biogeography, 30:1385-1396.
- EBACH, M.C. & MORRONE, J.J. 2005. Forum on historical biogeography: what is cladistic biogeography? Journal of Biogeography, 32:2179-2187.
- FUNK, V.A.. 2004. Revolutions in historical biogeography. In: Lomolino, M.V.; Sax, D.F. & Brown, J.H. (Eds.), Foundations of biogeography: classic papers with commentaries. University of Chicago Press, Chicago and London, p.647-657.
- GREHAN, J.H. 2001. Panbiogeografía y la geografía de la vida. In: Llorente, J. & Morrone, J.J. (Eds.), Introducción a la biogeografía en Latinoamérica: teorías, conceptos, métodos y aplicaciones. Las Prensas de Ciencias, Facultad de Ciencias, México, p.181-195.
- HÖRANDL, E. 2006. Paraphyletic versus monophyletic taxa-evolutionary versus cladistic classifications. Taxon, 55:564-570.
- HAROLD, A.S. & MOOI, R.D. 1994. Areas of endemism: definition and recognition criteria. Systematic Biology, 43:261-266.
- HAUSDORF, B. 2002. Units in biogeography. Systematic Biology, 51:648-651.
- HAUSDORF, B. & HENNING, C. 2003. Biotic element analysis in biogeography. Systematic Biology, 52:717-723.
- HEADS, M. 2005a. Dating nodes on molecular phylogenies: a critique of molecular biogeography. Cladistics, 21:62-78.
- HEADS, M. 2005b. The history and philosophy of panbiogeography. In: Morrone, J.J. & Llorente, J. (Eds.), Regionalización Biogeográfica en Iberoamérica y Tópicos Afines. Universidad Nacional Autónoma de México, México, p.67-123.
- HENNIG, W. 1966. Phylogenetic systematics. University of Illinois Press, Urbana.
- HUMPHRIES, C.J. & PARENTI, L.R. 1999. Cladistic biogeography: interpreting patterns of plant and animal distributions. 2nd edition. Oxford University Press, Oxford.
- KNAPP, S. 2005. Biogeography: space, form and time. Journal of Biogeography, 32:3-4.
- LIEBERMAN, B.S. 2003. Unifying theory and methodology in biogeography. Evolutionary Biology, 33:1-25.
- MATTHEW, W.D. 1915. Climate and evolution. Annals of the New York Academy of Sciences, 24:171-318.
- MAYR, E. 1982. The growth of biological thought. Harvard University Press, Cambridge.
- MANFRIN, M.H. & SENE, F.M. 2006. Cactophilic Drosophila in South America: a model for evolutionary studies. Genetica, 126:57-75.
- MCDOWALL, R.W. 2004. What biogeography is: a place for process. Journal of Biogeography, 31:345-351.
- MEIER, R. & WILLIAMANN, R. 2000. The Hennigian species concept. In: Wheeler, Q.D. & Meier, R. (Eds.), Species concepts and phylogenetic theory: a debate. Columbia University Press, New York, p.30-43.
- MISHLER, B.D. & BRNADON, R.N. 1987. Individuality, pluralism, and the phylogenetic species concept. Biology & Philosophy, 2:397-414.
- MORRONE, J.J. 1994. On the identification of areas of endemism. Systematic Biology, 43:438-441.
- MORRONE, J.J. 2002. El espectro del dispersalismo: de los centros de origen a las áreas ancestrales. Revista de la Sociedad Entomológica Argentina, 61:1-14.
- MORRONE, J.J. & CRISCI, J.V. 1995. Historical biogeography: introduction to methods. Annual Review of Ecology and Systematics, 26:373-401.
- NELSON, G. 1978. From Candolle to Croizat: comments on the history of biogeography. Journal of Historical Biology, 11:269-305.
- NELSON, G. 1994. Homology and systematics. In: Hall, B.K. (Ed.), Homology: the hierarchical basis of comparative biology. Academic Press, San Diego, p.101-149.
- NELSON, G.& LADIGES, P.Y. 2001. Gondwana, vicariance biogeography and the New York School revisited. Australian Journal of Botany, 49:389-409.
- NELSON, G.& PLATNICK, N. 1981. Systematics and biogeography: cladistics and vicariance. Columbia University Press, New York.
- NIHEI, S.S. 2006. Misconceptions about parsimony analysis of endemicity. Journal of Biogeography, 33:2099-2106.
- NIXON, K.C. & WHEELER, Q.D. 1992. Measures of phylogenetic diversity. In: Novacek, M.J. & Wheeler, Q.D. (Eds.), Extinction and phylogeny. Columbia University Press, New York, p.216-234.
- NORDAL, I. & STEDJE, B. 2005. Paraphyletic taxa should be accepted. Taxon, 54:5-6.
- PAPAVERO, N.; LLORENTE-BOUSQUETS, J.; ORGANISTA, D.E. & MASCARENHAS, R. 2000. História da biologia comparada: desde o Gênesis até o fim do Império Romano do Ocidente. Holos Editora, Ribeirão Preto.
- PICKETT, K.M. 2005. The new and improved PhyloCode, now with types, ranks, and even polyphyly: a conference report from the First International Phylogenetic Nomenclature Meeting. Cladistics, 21:79-82.
- PLATNICK, N. & NELSON, G. 1978. A method of analysis for historical biogeography. Systematic Zoology, 27:1-16.
- POSADAS, P.; CRISCI, J.V. & KATINAS, L. 2006. Historical biogeography: A review of its basic concepts and critical issues. Journal of Arid Environments, 66:389-403.
- RAGE, J.C. & JAEGER, J.J. 1995. The sinking Indian raft: a response to Thewison and McKenna. Systematic Biology, 44:260-264.
- RIDDLE, B.R. 2005. Is biogeography emerging from its identity crisis? Journal of biogeography, 32:185-186.
- ROSEN, B.R. 1988. From fossils to earth history: applied historical biogeography. In: Myers, A.A. & Giller, P.S. (Eds.), Analytical biogeography: an integrated approach to the study of animal and plant distributions. Chapman and Hall, London, p.437-481.
- ROSEN, B.R. & SMITH, A.B. 1988. Tectonics from fossils? Analysis of reef-coral and sea-urchin distributions from late Cretaceous to Recent, using a new method. In: Audley-Charles, M.G. & Hallam, A. (Eds.), Gondwana and Tethys. Série Geological Society Special Publication, n.37. Oxford University Press, Oxford, p.275-306.
- ROSEN, D.E. 1978. Vicariant patterns and historical explanation in biogeography. Systematic Zoology, 27:159-188.
- SANTOS, C.M.D. 2005. Parsimony analysis of endemicity: time for an epitaph? Journal of Biogeography, 32:1284-1286.
- SANTOS, C.M.D. in press [2007]. On basal clades and ancestral nodes. Journal of Biogeography.
- SZUMIK, C.A.; CUEZZO, F.; GOLOBOFF, P.A. & CHALUP, A.E. 2002. An optimally criterion to determine areas of endemism. Systematic Biology, 51:806-816.
- SZUMIK, C.A. & GOLOBOFF, P.A. 2004. Areas of endemism: an improved optimally criterion. Systematic Biology, 53:968-977.
- VRANA, P. & WHEELER, W. 1992. Individual organisms as terminal entities: laying the species problem to rest. Cladistics, 8:67-72.
- WALLACE, A.R. 1858. On the tendency of varieties to depart indefinitely from the original type. Proceedings of the Linnean Society of London, 3:53-62.
- WALLACE, A.R.1876. The geographical distribution of animals. Macmillan, London.
- WHEELER, Q.D. 1995. Systematics, the scientific basis for inventories of biodiversity. Biodiversity and Conservation, 4:476-489.
- WHEELER, Q.D. & MEIER, R. (eds.). 2000. Species concepts and phylogenetic theory: a debate. Columbia University Press, New York.
- WHEELER, Q.D. & PLATNICK, N. 2000. The phylogenetic species concept (sensu Wheeler and Platnick). In: Wheeler, Q.D. & Meier, R. (Eds.), Species concepts and phylogenetic theory: a debate. Columbia University Press, New York, p.55-69.
- WILLIAMS, D.M. & EBACH, M.C. 2004. The reform of palaeontology and the rise of biogeography : 25 years after 'ontogeny, phylogeny, paleontology and the biogenetic law' (Nelson, 1978). Journal of Biogeography, 31:685-712.
- WILLMANN, R. & MEIER, R. 2000. A critique from the Hennigian species concept perspective. In: Wheeler, Q.D. & Meier, R. (Eds.), Species concepts and phylogenetic theory: a debate. Columbia University Press, New York, p.101-118.
Publication Dates
-
Publication in this collection
05 July 2007 -
Date of issue
2007
History
-
Received
09 Nov 2006 -
Accepted
05 Apr 2007