Resumos
Apresentamos uma investigação comparativa da morfologia das terminálias masculina e feminina de gêneros da família Rhagionidae (Diptera, Brachycera, Tabanomorpha) com distribuição neotropical. Partindo do plano básico de Brachycera, hipóteses de homologias entre as peças reprodutivas foram analisadas em um contexto comparativo. Os resultados sugerem que as condições presentes em Rhagionidae são no geral muito modificadas quando comparadas com o ancestral comum mais recente de Brachycera. Este trabalho apresenta uma hipótese filogenética heurística cujos resultados podem servir como base para o entendimento da grande diversificação morfológica das terminálias masculina e feminina dos ragionídeos, apontando para a solução de algumas controvérsias a respeito da morfologia das estruturas reprodutivas do grupo.
Brachycera; Diptera; Morfologia; Neotropical; Rhagionidae; Terminália
We present a comparative investigation of both male and female terminalia of the Neotropical genera of Rhagionidae (Diptera, Brachycera, Tabanomorpha). Based on Brachyceran bauplan, homology hypotheses between the terminalia parts were analyzed in a comparative context. The results suggest that the conditions observed in Rhagionidae are in general very modified when compared to the common ancestor of Brachycera. Herein we show a heuristic phylogenetic hypothesis which may serve as a basis for the understanding of the great morphological diversification of the male and female genitalia of rhagionids, pointing to a solution to some controversies about the morphology of the reproductive structures in the group.
Brachycera; Diptera; Morphology; Neotropical; Rhagionidae; Terminalia
Morfologia comparada das terminálias masculina e feminina dos rhagionidae (Diptera, Tabanomorpha) neotropicais
Daniel D.D. CarmoI, II; Charles Morphy D. SantosI, III
IUniversidade Federal do ABC, Centro de Ciências Naturais e Humanas. Rua Santa Adélia, 166, Bairro Bangu, CEP 09210-170, Santo André, SP, Brasil
IIE-mail: dandorndias@gmail.com
IIIE-mail para correspondência: charles.santos@ufabc.edu.br
ABSTRACT
We present a comparative investigation of both male and female terminalia of the Neotropical genera of Rhagionidae (Diptera, Brachycera, Tabanomorpha). Based on Brachyceran bauplan, homology hypotheses between the terminalia parts were analyzed in a comparative context. The results suggest that the conditions observed in Rhagionidae are in general very modified when compared to the common ancestor of Brachycera. Herein we show a heuristic phylogenetic hypothesis which may serve as a basis for the understanding of the great morphological diversification of the male and female genitalia of rhagionids, pointing to a solution to some controversies about the morphology of the reproductive structures in the group.
Key-words: Brachycera; Diptera; Morphology; Neotropical; Rhagionidae; Terminalia.
RESUMO
Apresentamos uma investigação comparativa da morfologia das terminálias masculina e feminina de gêneros da família Rhagionidae (Diptera, Brachycera, Tabanomorpha) com distribuição neotropical. Partindo do plano básico de Brachycera, hipóteses de homologias entre as peças reprodutivas foram analisadas em um contexto comparativo. Os resultados sugerem que as condições presentes em Rhagionidae são no geral muito modificadas quando comparadas com o ancestral comum mais recente de Brachycera. Este trabalho apresenta uma hipótese filogenética heurística cujos resultados podem servir como base para o entendimento da grande diversificação morfológica das terminálias masculina e feminina dos ragionídeos, apontando para a solução de algumas controvérsias a respeito da morfologia das estruturas reprodutivas do grupo.
Palavras-chave: Brachycera; Diptera; Morfologia; Neotropical; Rhagionidae; Terminália.
INTRODUÇÃO
A ordem Diptera (Hexapoda, Holometabola) é dividida em "Diptera Inferiores" e Brachycera (Yeates & Wiegmann, 2005). Em Brachycera, há quatro infraordens: Muscomorpha, Stratiomyomorpha, Xylophagomorpha e Tabanomorpha (Yeates, 2002) - alguns autores aceitam a existência de uma quinta infraordem, Vermileonomorpha (Stuckenberg, 2001). Os Tabanomorpha são constituídos tradicionalmente por cinco famílias - Vermileonidae, Pelecorhynchidae, Athericidae, Tabanidae e Rhagionidae, esta última foco do presente trabalho.
Rhagionidae contém alguns dos mais antigos membros de Diptera, tendo se diversificado provavelmente há cerca de 175 milhões de anos (Santos, 2008; Kraemer & Nel, 2009). A família é constituída por 22 gêneros viventes, com aproximadamente 500 espécies descritas (Santos, 2008). Seu monofiletismo ainda é incerto, uma vez que existem poucos caracteres morfológicos para suportar as relações entre os seus gêneros e muitas das sinapomorfias listadas possuem caráter dúbio. As primeiras hipóteses das relações de parentesco entre os ragionídeos foram feitas sem uma análise filogenética formal (Nagatomi, 1982; Stuckenberg, 2001). Kerr (2010) é a análise cladística mais extensa feita até o momento para a família, combinando dados morfológicos e moleculares. Ela suporta a hipótese de que Rhagionidae é um grupo monofilético, mas sua amostragem taxonômica é relativamente pequena, com apenas 18 gêneros de um total de 49 aceitos (Santos, 2008).
Rhagionidae distribui-se por todo o globo e, de acordo com Santos (2008), apenas cinco de seus gêneros são encontrados na Região Neotropical: Atherimorpha White, 1915, Sierramyia (Neorhagio) (Lindner) Kerr, 2010, Rhagio Fabricius, 1775, Pelecorhynchus Macquart, 1850 e Chrysopilus Macquart, 1826. Dentre eles, Chrysopilus constitui a maioria absoluta da diversidade de ragionídeos neotropicais (Santos & Amorim, 2007). Não há indicativos de que exista um subgrupo monofilético de Rhagionidae endêmico dessa região, dado que há compartilhamento de vários gêneros presentes nesta e em outras regiões biogeográficas, notadamente no componente Gondwânico (como proposto anteriormente por Santos, 2008, e Amorim et al., 2009).
Nenhum dos poucos estudos acerca da morfologia comparada das terminálias masculina e feminina de Brachycera (Nagatomi & Iwata, 1976; Nagatomi, 1984; Sinclair et al., 1994) propôs-se a investigar especificamente os gêneros de Rhagionidae com distribuição na Região Neotropical e nenhum deles o fez em um contexto filogenético explícito. O objetivo do presente trabalho é cobrir parte dessa lacuna. Aqui, as terminálias masculina e feminina foram ilustradas e descritas de forma comparativa, o que permite reconstruir parte da história evolutiva das peças genitais de Rhagionidae desde o plano básico de Brachycera (Sinclair et al., 1994), apontando possíveis homologias e constituindo a base para uma análise filogenética mais ampla. Além disso, uma hipótese filogenética preliminar foi obtida com o intuito de verificar como as características de terminália variam entre os Rhagionidae neotropicais.
MATERIAL E MÉTODOS
Os espécimes examinados pertencem às coleções do Museu de Zoologia da Universidade de São Paulo, Brasil (MZUSP), do Museu de Zoologia "Alfonso L. Herrera" da Facultad de Ciencias de la Universidad Nacional Autônoma del México (MZFC) e do American Museum of Natural History, EUA (AMNH). A lista completa dos espécimes encontra-se na Tabela 1. As ilustrações das terminálias foram feitas a partir de lâminas permanentes e não permanentes. Para as lâminas não permanentes, os espécimes foram montados temporariamente em glicerina, em lâminas escavadas, que foram observadas com auxílio de microscópio com câmera clara. A lista de atributos foi elaborada no programa DELTA (Dallwitz et al., 2000) e funcionou como base para a codificação e a construção da matriz de táxons versus caracteres. Além da observação direta dos espécimes, também foram levantados dados a partir da literatura (e.g., Nagatomi & Iwata, 1976; Nagatomi, 1984; Sinclair et al., 1994; Santos, 2006). A matriz foi analisada no software WINCLADA (Nixon, 1999) através das opções busca heurística com 1.000 réplicas e otimização do tipo ACCTRAN.
A terminologia adotada segue as propostas de Santos (2005, 2006) e Cumming & Wood (2009).
RESULTADOS
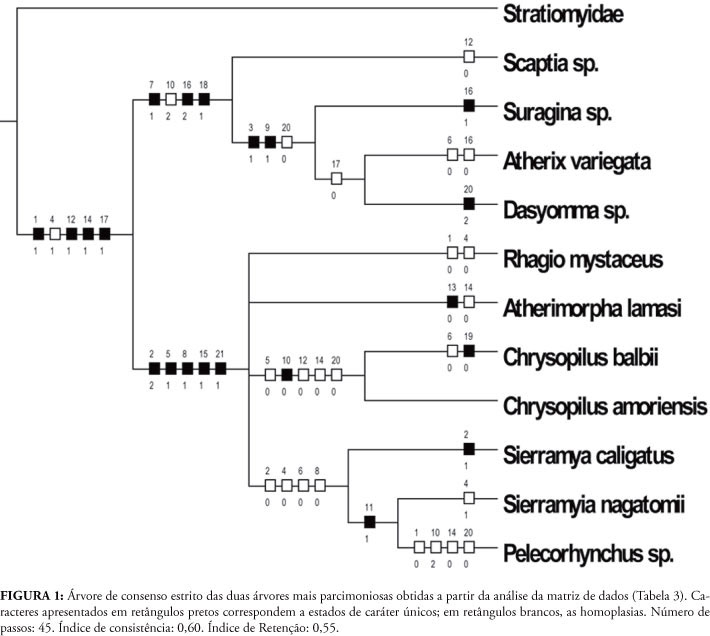
Foram examinados 31 exemplares da infraordem Tabanomorpha (Tabela 1). As descrições morfológicas das terminálias masculina e feminina dos gêneros de Rhagionidae distribuídos na Região Neotropical basearam-se em uma série de atributos estruturais, que constituíram a base para a codificação dos caracteres, listados na Tabela 2 e compilados na matriz de 21 caracteres versus 12 táxons terminais, que incluem gêneros de Rhagionidae, grupos-externos escolhidos entre outras famílias de Tabanomorpha e também um Stratiomyidae (Tabela 3).
A análise filogenética da matriz resultou em duas árvores igualmente parcimoniosas, cujo consenso estrito tem 45 passos, índice de consistência global 0,60 e índice de retenção global 0,55. Como o objetivo do presente trabalho foi o de fazer uma análise comparativa das terminálias masculina e feminina de ragionídeos neotropicais, esse cladograma deve ser considerado apenas uma hipótese heurística a partir da qual podem ser discutidos aspectos importantes da morfologia das terminálias, que correspondem a um conjunto de dados com alto sinal filogenético entre os Insecta, em geral, e entre Diptera, em particular (e.g., Almeida & Ale-Rocha, 2010; Song & Bucheli, 2010).
A despeito da limitada amostragem de táxons terminais no cladograma da Figura 1, o padrão obtido é congruente com algumas hipóteses levantadas para explicar a evolução de determinados aspectos morfológicos dos Rhagionidae (como discutido, por exemplo, em Santos, 2006 e Kerr, 2010). Isso demonstra que os caracteres de terminália masculina e feminina são fundamentais para se reconstruir as relações de parentesco e determinar os passos da história evolutiva dos ragionídeos.
DISCUSSÃO
Sobre a terminália masculina
As terminálias dos Rhagionidae neotropicais foram comparadas com as de gêneros de Athericidae (Figs. 2A, B) e Tabanidae (Figs. 2C, D), devido a proximidade filogenética desses grupos com os ragionídeos, além do bem estabelecido monofiletismo do clado constituído por esses dois táxons (Wiegmann et al., 2000; Yeates, 2002; Kerr 2010). Foram também feitas comparações com o plano básico de Brachycera elaborado por Sinclair et al. (1994). O plano-básico da terminália masculina de Brachycera é caracterizado pela presença de um edeago composto por um apódema ejaculatório, processo endoedeagal e um saco espermático proeminente, além de um par de parâmeros que constituem uma placa sobre o conjunto formado pelas peças constituintes do edeago. O saco espermático encontra-se, em geral, na porção medial dos segmentos da terminália masculina, próximo à região distal do apódema ejaculatório, dentro da placa parameral, e é circundado por dois escleritos que variam de tamanho, denominados processos ejaculatórios laterais. O apódema ejaculatório insere-se na margem anterior do saco espermático, no interior da base da placa parameral. Na porção mais distal do apódema ejaculatório encontra-se o processo endoedeagal. O tergito IX encontra-se separado do esternito IX.
Na base de Brachycera, o esternito IX forma uma placa triangular separada dos gonocoxitos (Matile, 1990; Sinclair et al., 1994). Essa característica se modifica tanto em Athericidae (aqui representado por Atherix variegata Walker, 1848) quanto em Tabanidae (Scaptia molesta Wiedemann, 1828), mostrados respectivamente nas Figs. 2A e 2C. A separação entre o esternito IX e os gonocoxitos mantém-se na maioria dos Rhagionidae estudados. No entanto, em Chrysopilus e em Pelecorhynchus ocorre a fusão total ou parcial dos gonocoxitos com o esternito IX (Figs. 3A, C e 4A, B). Apesar de compartilharem essa condição, ela provavelmente teve surgimento independente, uma vez que Chrysopilus e Pelecorhynchus não compõem, juntos, um grupo monofilético (Fig. 1), como discutido por Kerr (2010). Além disso, a fusão dos gonocoxitos com o esternito IX aparece em outros grupos de Diptera, como Eremoneura (Sinclair & Cumming, 2006) e Asilidae (Dikow, 2009).
Em Athericidae, observa-se a presença de um par de extensões endofálicas (tradução para aedeagal tines) muito alongadas (Fig. 2A), condição esta também observada em Tabanidae (Fig. 2C). De fato, a presença de extensões endofálicas longas pode ser vista como sinapomorfia de Athericidade + Tabanidae (Sinclair et al., 1994). Zloty et al. (2005) sugeriram que a presença de extensões endofálicas longas suportava o clado Oreoleptidae + Athericidade + Tabanidae. Dentro dos Rhagionidae neotropicais, essas estruturas sofrem grande redução em Rhagio e Sierramyia (Figs. 5C e 6A, C, respectivamente), e desaparecem em Chrysopilus, Atherimorpha e Pelecorhynchus (Figs. 3A, C; 4A, B; e 5A). Isso já havia sido discutido por Stuckenberg (1973). Para Kerr (2010), no entanto, não existem extensões endofálicas em Sierramyia ou Rhagio. Ele acredita ser falha a hipótese de Santos (2006) de que as estruturas presentes nesses dois gêneros são homólogas àquelas presentes na terminália de Athericidade + Tabanidae (e Bolbomyia, gênero tradicionalmente posicionado entre os Rhagionidae), apesar de declarar não ter analisado nenhum espécime de Sierramyia. Aqui, seguindo a interpretação de Stuckenberg (1973), Sinclair et al. (1994) e Santos (2006), as extensões endofálicas de Athericidae, Tabanidae e Rhagionidae são consideradas homólogas, com a hipótese filogenética apontando para uma redução do comprimento dessa estrutura entre os ragionídeos.
Em Athericidae + Tabanidae, o tergito IX encontra-se fundido ao tergito X e é sinapomorfia inequívoca do clado (Zloty et al., 2005), não ocorrendo entre os Rhagionidae, nos quais estas placas encontram-se separadas. Ainda nos Athericidae, os apódemas gonocoxais, quando presentes, encontram-se reduzidos e profundamente inseridos no gonocoxitos, ou ausentes. Essa última condição não ocorre nem entre os Tabanidae, nem entre os ragionídeos, nos quais o apódema gonocoxal geralmente se estende até a margem proximal do gonocoxito ou mesmo além dela (Figs. 2C; 3A, B; 4A, B; 5A, C; 6A, C). Embora Stuckenberg (1973) considere presentes os apódemas gonocoxais em Athericidae, as observações feitas aqui demonstram que as estruturas apontadas por ele são, de fato, processos ejaculatórios laterais bastante alongados, ultrapassando o limite distal dos gonocoxitos (Fig. 2A).
Os apódemas paramerais são robustos em Athericidae (Fig. 2A). Esse estado se mantém em Rhagio e Atherimorpha, mas há redução desses apódemas em Chrysopilus e Sierramyia (Figs. 3A, C; 4A, C; 6A, C). Em Pelecorhynchus os apódemas estão fundidos medialmente, uma condição autapomórfica do grupo (Fig. 4A).
Em Sierramyia e Atherimorpha observa-se a presença de uma placa dorsal ao edeago, denominada de guia edeagal (Nagatomi, 1984). Essa estrutura está ausente na descrição do plano básico de Brachycera feita por Sinclair et al. (1994), bem como em Stratiomyidae (Fig. 7A) mas aparece nos dois outros grupos externos estudados (Athericidae e Tabanidae). Isso sugere que a guia edeagal seja uma convergência entre Sierramyia, Atherimorpha, Athericidae e Tabanidae, hipótese que só pode ser testada através de uma análise filogenética ampla de Tabanomorpha, utilizando uma grande amostragem de gêneros de ragionídeos e das outras famílias da infraordem.
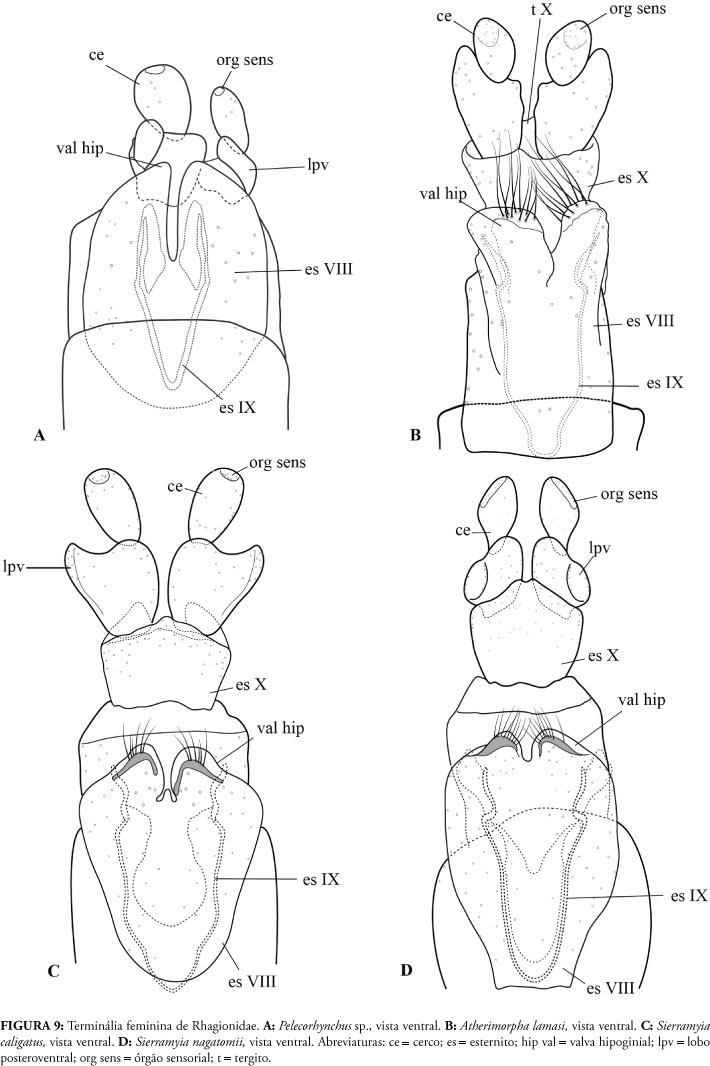
Dentro de Rhagionidae, o grupo que apresenta o maior número de modificações em relação ao plano-básico de Brachycera é sem dúvida Pelecorhynchus, o que tem levado vários autores (e.g., Woodley, 1989 e Kerr, 2010) a considerá-lo como gênero pertencente a uma família separada, Pelecorhynchidae. Em Pelecorhynchus, os apódemas gonocoxais encontram-se muito modificados e adquirem uma forma mais alongada e afilada (Fig. 4A), diferentemente de outros grupos de Rhagionidae. O saco espermático é muito reduzido e os apódemas paramerais não podem ser diferenciados dos parâmeros, o que pode significar a provável ocorrência de fusão dessas estruturas durante a evolução do grupo. Da mesma forma, não há gonóstilos robustos quando comparados com outros ragionídeos (Fig. 4A). Apesar das modificações apresentadas por Pelecorhynchus terem levado alguns autores a propor que gênero seja deslocado para a família Pelecorhynchidae, estudos recentes apontam para a reinserção de Pelecorhynchus em Rhagionidae (como Stuckenberg, 2001 e Santos, 2006), especialmente com base na estrutura da sua terminália feminina, como discutido abaixo.
Sobre a terminália feminina
É comum se considerar a existência de poucas variações na terminália feminina de Diptera. Aqui, quando Rhagionidae é comparado com os grupos-externos (Stratiomyidae, Athericidae e Tabanidae), pode-se notar que há caracteres da terminália feminina úteis para a identificação de gêneros (como é o caso de Pelecorhynchus, Fig. 9A) e espécies, devendo ser incluídos em análises filogenéticas de diferentes níveis.
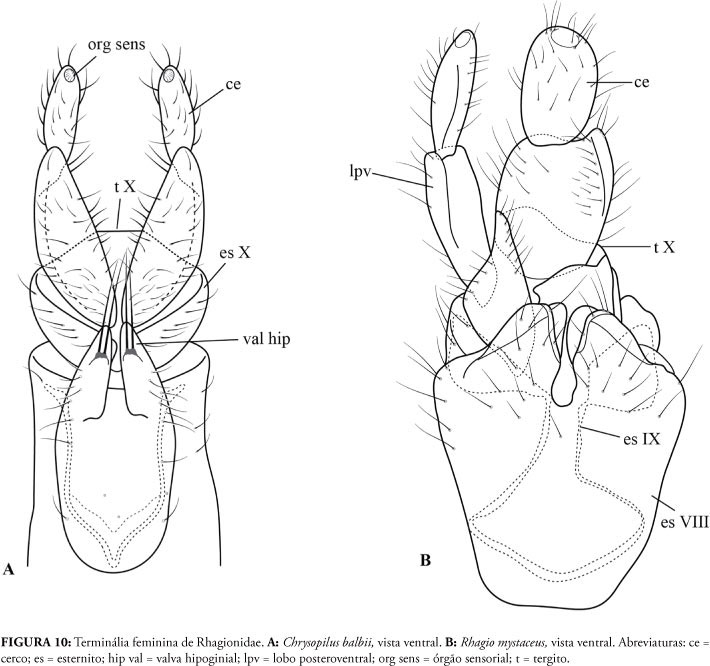
Tanto em Athericidae quanto em Tabanidae os cercos da terminália feminina possuem apenas um artículo (Fig. 8A-C), ao passo que em todos os grupos de Rhagionidae investigados, os cercos possuem dois artículos (Figs. 9A-C; 10A, B), o que vale também para outros gêneros não distribuídos na Região Neotropical (Santos, 2006). A condição "cerco biarticulado" é provavelmente plesiomórfica e presente no plano básico de Diptera (Cumming & Wood, 2009), mantendo-se na base da infraordem Tabanomorpha e se modificando no clado Athericidae + Tabanidae.
A valva hipoginial encontra-se presente nos gêneros Suragina (Athericidade) e Scaptia (Tabanidae) (Fig. 8A, B) e em todos os ragionídeos estudados, exceto em Pelecorhynchus. Assim, a sua presença pode ser interpretada como condição ancestral em Rhagionidae (como demonstrado também por Nagatomi & Iwata, 1976), desaparecendo em Pelecorhynchus. Nas valvas de Chrysopilus amoriensis (Bromley) Kerr, 2010 e Chrysopilus balbii Santos & Amorim, 2007, observa-se a presença de cerdas pronunciadas (Fig. 10A), características do gênero e úteis na identificação de suas espécies, como é o caso de C. balbii, em que são observadas apenas duas longas cerdas em cada uma das valvas (Santos & Amorim, 2007). Outra estrutura de importância filogenética é a furca genital - como apontam Nagatomi & Iwata (1976), Santos (2005, 2006), Santos & Amorim (2007) e Kerr (2010) (Figs. 8A-C; 9A-D; 10A, B). Em alguns grupos de Diptera, a furca genital tem importante papel funcional uma vez que mantém os ductos espermáticos em sua posição anatômica (Lewis & Dice, 1988).
Os autores que defendem o posicionamento de Pelecorhynchus na família Pelecorhynchidae apontam como autapomorfia da família a presença de uma evaginação do primeiro artículo do cerco, denominada lobo póstero-ventral (Woodley, 1989), provavelmente relacionada ao direcionamento dos excrementos (Stuckenberg, 2001). No entanto, apesar de muito pronunciado em Pelecorhynchus, o lobo póstero-ventral não é exclusivo do gênero, podendo ser observado em todos os Rhagionidae neotropicais (Figs. 9A-D; 10A, B). Santos (2006) já havia discutido a respeito dessa estrutura em outros gêneros da família, o que indica que ela está presente desde o ancestral comum de todos os Rhagionidae, com reduções em alguns grupos mais derivados.
CONSIDERAÇÕES FINAIS
A análise das terminálias masculina e feminina dos Rhagionidae neotropicais demonstra que a família possui um grande número de modificações com relação ao plano básico de Brachycera. Isso é interessante uma vez que o registro fóssil dos ragionídeos é antigo (Mostovski & Jarzembowski, 2000; Kraemer & Nel, 2009), o que os colocaria muito próximos à origem dos Diptera, apesar de Rhagionidae ser um grupo bastante derivado quando consideradas a sua morfologia, principalmente das terminálias masculina e feminina, e as suas relações de parentesco com os demais dípteros. Assim, duas interpretações são possíveis: ou os Diptera diversificaram-se de maneira explosiva no início do Mesozóico (no Triássico Inferior, há pelo menos 250 milhões de anos) ou eles se originaram muito tempo antes, ainda no Paleozóico, no Permiano (entre 251 e 299 milhões de anos) ou mesmo no Carbonífero (entre 359 e 299 milhões de anos) (Amorim & Silva, 2002; Santos, 2008), apesar de os fósseis indicarem idades mais recentes. De acordo com Wiegmann et al. (2011), os Diptera teriam sofrido três episódios de rápida radiação: Diptera inferiores (220 milhões de anos), Brachycera inferiores (180 milhões de anos) e Schizophora (65 milhões de anos).
Na Região Neotropical houve uma grande diversificação entre os Rhagionidae, com ênfase no gênero Chrysopilus, encontrado desde o sul da América do Sul até as fronteiras com a região Neártica, ao norte do México. A grande diversidade morfológica neotropical impede que o trabalho se esgote sem antes se fazer uma revisão aprofundada de todos os gêneros de Rhagionidae, algo fora do escopo do presente artigo. Assim, para resolver outras questões filogenéticas entre os Rhagionidae, e dessa família com os demais grupos da infraordem Tabanomorpha, é necessária uma análise filogenética robusta, envolvendo um número muito maior de táxons e de caracteres. No entanto, o estudo da morfologia comparada da terminálias, tanto masculina quanto feminina, como proposto aqui, pode funcionar como base para análises mais amplas, provenientes não só das peças reprodutoras, mas também de outros conjuntos de dados morfológicos com forte sinal filogenético (como peças bucais, escleritos torácicos e venação alar).
AGRADECIMENTOS
Os autores agradecem à Drª. Vanesssa Verdade (UFABC), à Msc. Sarah Oliveira (USP-RP) e ao Msc. Renato Capellari (USP-RP) pela leitura e sugestões em uma versão preliminar desse artigo. O trabalho foi financiado pelo CNPq (474511/2009-0), FAPESP (2010/07006-6) e pelo Programa de Iniciação Científica da Universidade Federal do ABC (PIC/UFABC). Parte dos resultados foi apresentada no VII Congresso Internacional de Dipterologia, ocorrido em agosto de 2010 em San José, Costa Rica.
Recebido em: 21.07.2011
Aceito em: 05.10.2011
Impresso em: 16.12.2011
- ALMEIDA, J.C. & ALE-ROCHA, R. 2010. Comparative morphology of the male terminalia of the subtribe Rhinotorina (Diptera, Heleomyzidae, Rhinotorini). Zootaxa, 2736:44-56.
- AMORIM, D.S. & SILVA, V.C. 2002. How far advanced was Diptera evolution in Pangea. Annales de la Societé Entomologique de France, 38:177-200.
- AMORIM, D.S.; SANTOS, C.M.D. & OLIVEIRA, S.S. 2009. Allochronic taxa as an alternative model to explain circumantartic disjunctions. Systematic Entomology, 34:2-9.
- CUMMING, J.M. & WOOD, D.M. 2009. Adult morphology and terminology. In: Brown, B.V.; Borkent, A.; Cumming, J.M.; Wood, D.M.; Woodley, N.E. & Zumbado, M.A. (Eds.). Manual of Central American Diptera. Volume 1. NRC-CNRC Research Press, Ottawa, p. 2-50.
- DALLWITZ, M.J.; PAINE, T.A. & ZURCHER, E.J. 2000. Principles of interactive keys. Disponível em: http://delta-intkey.com Acesso em: 02/set./2011.
- DIKOW, T. 2009. Phylogeny of Asilidae inferred from morphological characters of imagines (Insecta: Diptera: Brachycera: Asiloidea). Bulletin of the American Museum of Natural History, 319:1-175.
- KERR, P.K. 2010. Phylogeny and classification of Rhagionidae, with implications for Tabanomorpha (Diptera: Brachycera). Zootaxa, 2592:1-133
- KRAEMER, M.M.S. & NEL, A. 2009. First recorded evidence in the fossil record of snipe flies (Diptera: Rhagionidae) in Cretaceous amber, France. Cretaceous Research, 30:1367-1375.
- LEWIS, D.J. & DYCE, A. 1988. Taxonomy of the Australian Phlebotominae (Diptera: Psychodidae) with revision of genus Sergentomyia from region. Invertebrate Systematics, 2:755-804.
- MATILE, L. 1990. Recherches sur la systématique et l'évolution des Keroplatidae (Diptera, Mycetophiloidea). Mémoires du Muséum national d'Histoire naturelle Paris (A), 148:1-682.
- MOSTOVSKI, M.B. & JARZEMBOWSKI, E.A. 2000. The first brachycerous flies (Diptera: Rhagionidae) from the Lower Jurassic of Gondwana. Paleontological Journal, 34:367-369.
- NAGATOMI, A. 1982. The genera of Rhagionidae (Diptera). Journal of Natural History, 16:31-70.
- NAGATOMI, A. 1984. Male genitalia of the Lower Brachycera (Diptera). Beiträge zur Entomologie, 34:99-157.
- NAGATOMI, A. & IWATA, K. 1976. Female Terminalia of the Lower Brachycera (Diptera). Beiträge zur Entomologie, 26:5-47.
- NAGATOMI, A. & IWATA, K. 1978. Female terminalia of lower Brachycera - II. Beiträge zur Entomologie, 28:263-293.
- NIXON, K.C. 1999. Winclada (BETA) ver. 0.9.9. Publicado pelo autor, Ithaca, NY. Disponível em: http://www.cladistics.com/about_winc.htm Acesso em 02/set./2011.
- SANTOS, C.M.D. 2005. First record of genus Atherimorpha (Diptera:Rhagionidae) in Brazil, with description of new species. Zootaxa, 1021:37-43.
- SANTOS, C.M.D. 2006. Description of two new species of Neorhagio (Diptera, Tabanomorpha, Rhagionidae), and remarks of a controversial female character. Zootaxa, 1174:49-62.
- SANTOS, C.M.D. 2008. Geographical distribution of Tabanomorpha (Diptera, Brachycera): Athericidae, Austroleptidae, Oreoleptidae, Rhagionidae, and Vermileonidae. Entomobrasilis, 1(3):43-50. Disponível em: http://www.periodico.ebras.bio.br/ojs/index.php/ebras/article/view/25/44 Acesso em: 02/set./2011.
- SANTOS, C.M.D. & AMORIM, D.S. 2007. Why biogeografical hypotheses need a well supported phylogenetic framework: a conceptual evaluation. Papéis Avulsos de Zoologia, 47(4):63-73.
- SINCLAIR, B.J. & CUMMING, J.M. 2006. The morphology, higher-level phylogeny and classification of the Empidoidea (Diptera). Zootaxa, 1180:1-172.
- SINCLAIR, B.J.; CUMMING, J.M. & WOOD, D.M. 1994. Homology and phylogenetic implications of male genitalia in Diptera - Lower Brachycera. Entomologica Scandinavica, 24:407-432.
- SONG, H. & BUCHELI, S.R. 2010. Comparison of phylogenetic signal between male genitalia and non-genital characters in insect systematics. Cladistics, 26:23-35.
- STUCKENBERG, B.R. 1973. The Athericidae, a new family in the lower Brachycera (Diptera). Annals of the Natal Museum, 21:649-673.
- STUCKENBERG, B.R. 2001. Prunning the tree: a critical review of classifications of the Homeodacytla (Diptera, Brachycera), with new perspectives and an alternative classification. Studia Dipterologica, 8:3-42.
- WIEGMANN, B.M.; TRAUTWEIN, M.D.; WINKLER, I.S.; BARRA, N.B.; KIM, J-W.; LAMBKIN, C.; BERTONE, M.A.; CASSEL, B.K.; BAYLESS, K.M.; HEIMBERG, A.M.; WHEELER, B.M.; PETERSON, K.J.; PAPE, T.; SINCLAIR, B.J.; SKEVINGTON, J.H.; BLAGODEROV, V.; CARAVAS, J.; KUTTY, S.N.; SCHMIDT-OTT, U.; KAMPMEIER, G.E.; THOMPSON, F.C.; GRIMALDI, D.A.; BECKENBACH, A.T.; COURTNEY, G.W.; FRIEDRICH, M.; MEIER, R. & YEATES, D.K. 2011. Episodic radiations in the fly tree of life. Proceedings of the National Academy of Sciences, 108:5690-5695.
- WIEGMANN, B.M.;. TSAUR, S-C.; WEBB, D.W.; YEATES, D.K. & CASSEL, B.K. 2000. Monophyly and relationships of the Tabanomorpha (Diptera: Brachycera) based on 28S ribosomal gene sequences. Annals of the Entomological Society of America, 93:1031-1038.
- WOODLEY, N.E. 1989. Phylogeny and classification of the "orthorrhaphous" Brachycera. In: McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (Coords.). Manual of Nearctic Diptera. Volume III. Research Branch. Agriculture Canada, Ottawa, p. 1371-1395.
- YEATES, D.K. 2002. Relationships of the Lower Brachycera (Diptera): a quantitative synthesis of morphological characters. Zoologica Scripta, 31:105-121.
- YEATES, D.K. & WIEGMANN, B.M. 2005. The evolutionary biology of flies. Columbia University Press.
- ZLOTY, J.; SINCLAIR, B.J. & PRITCHARD, G. 2005. Discovered in our backyard: a new genus and species of a new family from the Rocky Mountains of North America (Diptera, Tabanomorpha). Systematic Entomology, 30:248-266.


Datas de Publicação
-
Publicação nesta coleção
29 Nov 2011 -
Data do Fascículo
2011
Histórico
-
Aceito
05 Out 2011 -
Recebido
21 Jul 2011