Abstracts
Despite their wide use in ornithological surveys, point counts and mist nets follow protocols developed in temperate regions, with little attention to possible modifications for tropical systems. Using these methods on a 3-month basis from December 2009-January 2011 in two forest fragments in southeastern Brazil, we wished to evaluate how long these locations needed to be surveyed with point counts for a relatively complete avifaunal inventory (at least 90% of all species and contacts), and if mist net hourly captures can equally detect numbers of species and individuals. Daily counting with four 20-min points during five consecutive days in a rain forest (MC) detected 90% of the estimated species richness after 20 h (60 20-min point counts), while 17 h (51 20-min point counts) did not detect 90% of the estimated species richness in a semideciduous forest (IT). The first 5 min of point counting in MC (63% of all species) and in IT (65%) detected significantly more species than the remaining minutes, but it took 15 min to accumulate 86% of all contacts in both forests. Consecutive 5-day mist netting (~ 9 h/day) resulted in 70.5 net-h/m² (MC) and 74.8 net-h/m² (IT) of sample effort, but 80-85% of the estimated number of species was obtained. Although accumulation curves showed no tendency towards stabilization of the number of observed species, the estimated number of species began to stabilize after the first 20 h in both forests. There was no significant difference in capture rates for both species richness and abundance among hourly net checks, but a trend in which these parameters were highest between the second and fourth checks of the day was observed. A 3-day (43.8 and 63.3 net-h/m²) mist netting section was enough to record 90% of the species captured during five days in MC and IT, respectively, while precise enough not to jeopardize species richness estimation. The number of individuals, however, decreased order 34% in MC and 38% in IT under the same conditions. Considering the number of net checks, 90% of the estimated species richness was captured until the 1100 h check in both remnants, while 67% of all individuals were captured until this same hour. Our results demonstrate that surveying the avifauna in these locations will require unique approaches, which must be tested before the beginning of point counting or net opening.
Avian abundance; Bird species richness; Bird surveying methods; Method efficiency; Neotropical forests
Embora muito utilizado com a finalidade de estimar a abundância de espécies de aves, pontos de escuta e redes de neblina seguem protocolos desenvolvidos em regiões temperadas, com pouca atenção para modificações para sistemas tropicais. Para averiguar por quanto tempo é necessária amostragem por pontos de escuta para o registro da maior parte da avifauna (ao menos 90% de todas as espécies e indivíduos), assim como para determinar se as capturas com redes de neblina em intervalos de 1 h detectam igualmente números de espécies e indivíduos, ambas as metodologia foram utilizadas a cada três meses entre dezembro de 2009 e janeiro de 2011 em dois fragmentos florestais do sudeste do Brasil. Quatro pontos de escuta de 20 min conduzidos durante cinco dias consecutivos acumularam 90% da riqueza estimada após 20 h (60 pontos de 20 minutos) em uma mata ombrófila densa (MC), enquanto 17 h (51 pontos de 20 minutos) foram insuficientes para o registro da mesma porcentagem de espécies em uma mata semidecidual (IT). Os primeiros 5 min dos pontos de escuta detectaram significativamente mais espécies em MC (63% do total de espécies) e em IT (65%) em comparação com os minutos restantes, mas foram necessários 15 min para o registro de 86% do total de contatos em ambas as florestas. Cinco dias consecutivos (~ 9 h/dia) com redes de neblina abertas resultaram em 70,5 horas-rede/m² (MC) e 74,8 horas-rede/m² (IT) de esforço amostral, de modo que 80 a 85% do número estimado de espécies foram capturados. Embora curvas de acumulação não tenham apresentado tendência à estabilização do número de espécies observado, o número de espécies estimado demonstrou assíntota a partir das primeiras 20 h em ambas as florestas. Não houve diferença significativa na captura de espécies ou indivíduos entre horários de revisões a cada hora, mas notou-se uma tendência na qual tais parâmetros mostraram-se mais elevados entre as segundas e quartas revisões do dia. Redes de neblina abertas durante três dias (43,8 e 63,3 horas-rede/m² em MC e IT, respectivamente) foram suficientes para o registro de 90% das espécies capturadas. Essa diminuição do esforço amostral não prejudicou a estimativa do número de espécies, ao passo que o número de indivíduos capturados diminuiu em 34% em MC e 38% em IT. As revisões até as 1100 h capturaram 90% de todas as espécies registradas com redes de neblina em ambos os fragmentos, porém 67% de todos os indivíduos foram capturados até este horário. Nossos resultados demonstram que o inventário e a estimativa de abundância da avifauna nessas localidades requerem delineamentos únicos, os quais devem ser testados antes do início da coleta de dados com pontos de escuta ou redes de neblina.
Abundância da avifauna; Eficiência de métodos; Florestas neotropicais; Métodos de amostragem de aves; Riqueza de espécies de aves
Evaluation of methodological protocols using point counts and mist nets: a case study in southeastern Brazil
Vagner CavarzereI,II,2 2 . Corresponding author, e-mail: cavarzere@usp.br. ; Fernanda AlvesI,II; Érika MachadoII; Marco Antônio RegoII; Luís Fábio SilveiraII; Marcos Mendonça Costa1 1 . Rua Amadeu Geraldo, nº 210, CEP 13211-683, Jundiaí, SP, Brasil. ; Andrés Calonge-Méndez1 1 . Rua Amadeu Geraldo, nº 210, CEP 13211-683, Jundiaí, SP, Brasil.
IPós-Graduação, Depto. Zoologia, Universidade de São Paulo. Rua do Matão, Travessa 14, nº 101, CEP 05508-900, São Paulo, SP, Brasil
IISeção de Aves, Museu de Zoologia, Universidade de São Paulo. Caixa Postal 42.494, 04218-970, São Paulo, SP, Brasil
ABSTRACT
Despite their wide use in ornithological surveys, point counts and mist nets follow protocols developed in temperate regions, with little attention to possible modifications for tropical systems. Using these methods on a 3-month basis from December 2009-January 2011 in two forest fragments in southeastern Brazil, we wished to evaluate how long these locations needed to be surveyed with point counts for a relatively complete avifaunal inventory (at least 90% of all species and contacts), and if mist net hourly captures can equally detect numbers of species and individuals. Daily counting with four 20-min points during five consecutive days in a rain forest (MC) detected 90% of the estimated species richness after 20 h (60 20-min point counts), while 17 h (51 20-min point counts) did not detect 90% of the estimated species richness in a semideciduous forest (IT). The first 5 min of point counting in MC (63% of all species) and in IT (65%) detected significantly more species than the remaining minutes, but it took 15 min to accumulate 86% of all contacts in both forests. Consecutive 5-day mist netting (~ 9 h/day) resulted in 70.5 net-h/m2 (MC) and 74.8 net-h/m2 (IT) of sample effort, but 80-85% of the estimated number of species was obtained. Although accumulation curves showed no tendency towards stabilization of the number of observed species, the estimated number of species began to stabilize after the first 20 h in both forests. There was no significant difference in capture rates for both species richness and abundance among hourly net checks, but a trend in which these parameters were highest between the second and fourth checks of the day was observed. A 3-day (43.8 and 63.3 net-h/m2) mist netting section was enough to record 90% of the species captured during five days in MC and IT, respectively, while precise enough not to jeopardize species richness estimation. The number of individuals, however, decreased order 34% in MC and 38% in IT under the same conditions. Considering the number of net checks, 90% of the estimated species richness was captured until the 1100 h check in both remnants, while 67% of all individuals were captured until this same hour. Our results demonstrate that surveying the avifauna in these locations will require unique approaches, which must be tested before the beginning of point counting or net opening.
Key-words: Avian abundance; Bird species richness; Bird surveying methods; Method efficiency; Neotropical forests.
RESUMO
Embora muito utilizado com a finalidade de estimar a abundância de espécies de aves, pontos de escuta e redes de neblina seguem protocolos desenvolvidos em regiões temperadas, com pouca atenção para modificações para sistemas tropicais. Para averiguar por quanto tempo é necessária amostragem por pontos de escuta para o registro da maior parte da avifauna (ao menos 90% de todas as espécies e indivíduos), assim como para determinar se as capturas com redes de neblina em intervalos de 1 h detectam igualmente números de espécies e indivíduos, ambas as metodologia foram utilizadas a cada três meses entre dezembro de 2009 e janeiro de 2011 em dois fragmentos florestais do sudeste do Brasil. Quatro pontos de escuta de 20 min conduzidos durante cinco dias consecutivos acumularam 90% da riqueza estimada após 20 h (60 pontos de 20 minutos) em uma mata ombrófila densa (MC), enquanto 17 h (51 pontos de 20 minutos) foram insuficientes para o registro da mesma porcentagem de espécies em uma mata semidecidual (IT). Os primeiros 5 min dos pontos de escuta detectaram significativamente mais espécies em MC (63% do total de espécies) e em IT (65%) em comparação com os minutos restantes, mas foram necessários 15 min para o registro de 86% do total de contatos em ambas as florestas. Cinco dias consecutivos (~ 9 h/dia) com redes de neblina abertas resultaram em 70,5 horas-rede/m2 (MC) e 74,8 horas-rede/m2 (IT) de esforço amostral, de modo que 80 a 85% do número estimado de espécies foram capturados. Embora curvas de acumulação não tenham apresentado tendência à estabilização do número de espécies observado, o número de espécies estimado demonstrou assíntota a partir das primeiras 20 h em ambas as florestas. Não houve diferença significativa na captura de espécies ou indivíduos entre horários de revisões a cada hora, mas notou-se uma tendência na qual tais parâmetros mostraram-se mais elevados entre as segundas e quartas revisões do dia. Redes de neblina abertas durante três dias (43,8 e 63,3 horas-rede/m2 em MC e IT, respectivamente) foram suficientes para o registro de 90% das espécies capturadas. Essa diminuição do esforço amostral não prejudicou a estimativa do número de espécies, ao passo que o número de indivíduos capturados diminuiu em 34% em MC e 38% em IT. As revisões até as 1100 h capturaram 90% de todas as espécies registradas com redes de neblina em ambos os fragmentos, porém 67% de todos os indivíduos foram capturados até este horário. Nossos resultados demonstram que o inventário e a estimativa de abundância da avifauna nessas localidades requerem delineamentos únicos, os quais devem ser testados antes do início da coleta de dados com pontos de escuta ou redes de neblina.
Palavras-chave: Abundância da avifauna; Eficiência de métodos; Florestas neotropicais; Métodos de amostragem de aves; Riqueza de espécies de aves.
INTRODUCTION
Determining avian abundance is important for understanding both the ecology and conservation needs of species. Two principal methodologies have been used to obtain this information: mist nets and audio-visual counts (Blake & Loiselle, 2001). Both methods have been compared as survey techniques (Gram & Faaborg, 1997; Poulin et al., 2000), offering benefits and costs (Wallace et al., 1996; Dunn & Ralph, 2004), while providing different perspectives on community structure (Whitman et al., 1997; Blake & Loiselle, 2000). Decades ago, several authors noted that mist nets do not provide trustworthy relative abundance estimates among different species, estimates of absolute density of the same species in the same habitat, or the inability of sampling vertical strata, especially those with canopies above net height (MacArthur & MacArthur, 1974; Karr, 1981). Capture probabilities based on observed values (rather than estimated values) may also result in differences among age classes or sex among species, and even among individuals of the same species in different habitats (Remsen & Good, 1996).
Recent studies have demonstrated that unbiased relative abundances of different species (and/or of the same species among different habitats), age classes or sex can be estimated with the use of mist nets through sampling designs and data analyses that take into account the heterogeneity in capture probabilities among species (and/or the same species among habitats). An example of such approach is Pollock's robust design, more recently detailed by Williams et al. (2002). Since density is defined by number of individuals/area, it is hard to conceive that mist nets can provide density estimates at all because it is hard to define horizontal areas covered by them. However, it is possible to obtain population size estimates based on capture-recapture with the state-space formulation of the robust design version of the Cormack-Jolly-Seber model, which was shown to be quite robust (Chase et al., 1997). Evaluation studies have also shown that abundance indices derived from mist net sampling often compare well to independent data on the parameters of interest. Several comparisons have been made between long-term trends in abundance indices based on netting data and trends from independent sources; correlations were strongest when statistical techniques were used that compensated for variation in detections (Dunn & Ralph, 2004 and references therein).
Similarly, audio-visual methods are known to have deficiencies in species detection, as well as in providing accurate avian density measures (Verner, 1985). Distance sampling by transect or point counts is probably the best known technique to estimate density without capture-recaptures because it explicitly takes into account imperfect detection. Cimprich (2009), for example, found distance sampling point counting to be a good abundance estimate of the Black-capped Vireo Vireo atricapilla, but he did stress the importance of count duration. In other words, studies have shown that the problem was not with techniques, but with the way empirical data was used. This motivated the development of sampling designs that allow for detection probabilities to vary among species or habitats, such as the occupancy modeling approach (Mackenzie et al., 2002).
While comparison between methods is fairly common (Pagen et al., 2002; Martin et al., 2010; Arizaga et al., 2011), an aspect that has received less attention is that these methods are widely used in tropical regions based on protocols primarily designed for temperate regions, and not properly tested to guarantee maximum efficiency in tropical systems. Seasonality, for example, does not seem to influence the detection of the number of species and individuals with point counts in semideciduous forests in southern Brazil (Volpato et al., 2009), or transect counts in a cerrado landscape in São Paulo (Cavarzere, 2013). Seven- to 10-min point counts, rather than 20-min point counts, are enough to detect most species, including threatened and/or endemic species in semideciduous and rain forests (Develey, 2004; Betini, 2001). Thus, the design of the study and nature of variability in bird detection will determine whether increasing the number of points (or the number of visits per point) will have greater effect in detecting a population trend (Thompson et al., 2002). Many researchers have tested the efficiency of mist nets (e.g., Jenni et al., 1996; Rappole et al., 1998; Lövei et al., 2001; Ralph & Dunn, 2004; Whitman, 2004), but these evaluations, except for capture rates with different mesh sizes (Piratelli, 2003) and variations in capture rates between early and late hours of the day (Mallet-Rodrigues & Noronha, 2003), remain to be deciphered in Brazil.
Here, we address the following procedural issues surrounding avian census methods in Neotropical regions. It is expected that a survey will not detect most species when the time at a point count is longer than the time for which the species detection rate is maximized or when this time is extended beyond the point at which a representative percentage (90%) of the predicted species diversity has been detected (Vergara et al., 2010). Therefore, our first goal was to evaluate how much time is needed to survey two forests with point counts to record at least 90% of the estimated species richness. As birds become more active during the first hours of the day (Blake, 1992), it would be expected that capture rates are greatest during the first net checks. Thus, our second aim was to examine if time of day, measured as hourly net checks, affects the number of both species and individuals captured. Our null hypothesis was that hourly checks could equally detect species and individuals. We did not wish to compare methods nor temperate versus tropical systems.
MATERIALS AND METHODS
Study areas
We conducted this study in two localities: Fazenda Entre Rios and Fazenda Montes Claros (Fig. 1). Fazenda Entre Rios (23º16'S, 48º26'W; 680 m) is located in the municipality of Bofete, in the plateaus of the interior of the state of São Paulo, southeastern Brazil. Bofete lies on the boundary of three other municipalities: Angatuba, Itatinga and Pardinho.
According to Köppen's classification, climate is humid subtropical (Cfa), seasonal, with annual rainfall of 1,400 mm (Viani & Rodrigues, 2007). We surveyed one remnant (319 ha) in this location, hereafter IT, composed of seasonal semideciduous forest surrounded by Eucalyptus sp. plantations. Fazenda Montes Claros (23º02'S, 46º01'W; 690 m) is located in the municipality of São José dos Campos, São Paulo, approximately 250 km from IT, abutting the municipalities of Sapucaí-Mirim and Camanducaia in Minas Gerais state, and Joanópolis, Piracaia, Ugaratá, Jacareí, Jambeiro, Caçapava and Monteiro Lobato, in São Paulo. The city lies in the Paraíba do Sul River Valley between the Serra do Mar and Serra da Mantiqueira mountain ranges of the Atlantic forest. We surveyed a mature 1,150 ha forest fragment (hereafter MC) with predominance of bamboo (Merostachys sp.) thickets; Eucalyptus sp. plantations also surround the matrix habitat.
Sampling design and bird counts
We surveyed birds visually, with the help of binoculars, as well as aurally, during four seasons (visiting locations every 3 months), from December 2009-January 2011. We positioned 20 mist nets (12 m × 3 m × 30 mm) on two separate 120 m trails ca. 1 km apart in each study area. At the beginning and end of each mist net line we determined one point count and visited all four points (two on each mist net line, two lines in each counting station) during five consecutive days. Point counting, with a 100 m radius of detection, lasted for 20 min and started 10 min before sunrise (Vielliard & Silva, 1990; Bibby et al., 2000); we varied the sequence of starting points among days. On some occasions we also annotated species and number of contacts into 5-min intervals. The same individual was not considered for subsequent intervals if already recorded in a former interval, but the same species was annotated again in a subsequent interval if it represented unequivocally a distinct individual from a preceding one. Mist nets touched bottom and were open during five consecutive days from 0600-1400 h. Point counts and mist nets were conducted simultaneously and a 5-day sampling was carried out four times, once during each season. Birds were banded with unique metallic rings provided by the Centro Nacional de Pesquisa para Conservação das Aves Silvestres (CEMAVE), measured, weighed, photographed and then released. Individuals that casually died are housed in the Museu de Zoologia da Universidade de São Paulo (MZUSP).
Analyses
A One-Way Repeated Measures ANOVA was used to examine differences in numbers of species and contacts (= number of individuals) among 5-min counting intervals and hourly variations with mist net captures. Tukey pairwise tests were used to determine differences within intervals and net checks. Each campaign was considered a replica, in which case the number of seasons corresponded to the number of samples (N = 4). We are aware that there may be differences in bird detections among seasons and one could argue that seasons may represent pseudoreplication. However, our objectives were simpler and aimed to determine the variations in bird detections in spite of the months we conducted these surveys. In order to investigate if interactions between hour of the day and season might occur, we would need to present different sampling designs (such as temporal replicas acquired along at least three years), an approach we do not possess at the moment.
We did not pool seasonal values of either species richness or abundance, so each season (sample) consisted of empirical (observed) numbers of species and individuals. Points resulted in four classes of minutes (0-5, 6-10, 11-15 and 16-20 min), whereas hourly variations corresponded to net checks. Mist netting analyses were performed separately for three and five consecutive days so we could evaluate cut off points in a sense that longer periods of mist netting would (or would not) be necessary. The first net check started at 0600 h and continued at regular 1 h intervals. The number of species and individuals was not pooled and the analyses included observed values. Sample-based species accumulation curves with order randomized 100 times, as well as the Michaelis-Menten species richness estimator (MMMean; Keating & Quinn, 1998), which performed best compared to other estimators (Herzog et al., 2002), were produced with EstimateS 8.2 (Colwell, 2009). Mist net effort was calculated as the number of open nets (net-h) divided by m2 (Bibby et al., 2000). Statistical analyses were performed with SPSS 13.0 (SPSS, 2004) and the significance level adopted was α = 0.05.
RESULTS
Point counts
We obtained 3,641 contacts of 180 bird species over a total of 124 point counts. We accumulated 52 point counts in IT (17 h) recording 113 species and 867 contacts, and carried out 72 point counts in MC (24 h), acquiring 2,774 contacts of 132 species (Appendix Appendix ). Surveying during the first 23 h accounted for > 90% of all observed contacts in MC, whereas 90% of all contacts were recorded after 16 h of point counting in IT. The MMMean estimator predicted 132 and 140 species for IT and MC, respectively, meaning that as many as 19 species may have gone unnoticed in IT and eight species were not detected in MC. Accumulation curves showed no tendency towards stabilization, but while it took 20 h (60 20-min points) to record 90% of the predicted number of species in MC, only 85% of the predicted number of species was detected in IT after the total surveying hours in this site (Fig. 2).
On occasions in which we assigned birds into 5-min intervals we accumulated 28 points in IT (9.3 h, 309 contacts of 75 species) and 20 points in MC (6.7 h, 361 contacts of 70 species). The first 5 min accounted for 63% and 65% of all species recorded in MC and IT, respectively, but 86% of all contacts were detected only after 10-min counts in both study sites. The number of accumulated species increased constantly with increasing surveying effort, but the number of estimated species did not increase accordingly. The number of species and contacts recorded were significantly different among intervals in IT (Frichness3-18 = 42.42, = 8.64, P = 0.001) and in MC P = 0.001; Fabundance3-18 = 12.73, P = = 15.98, (Frichness3-18 0.000; Fabundance3-18 P = 0.000), being greatest during the first 5 min in both study sites (Table 1). Despite a trend in decreasing species richness, the remaining time intervals were not significantly different among each other (Fig. 3).
Mist nets
At the end of 20 days, we captured 1,404 individuals of 108 bird species over a total of 145.3 net-h/ m2. We accumulated 70.5 net-h/m2 in IT (600 individuals of 80 species) and another 74.8 net-h/m2 in MC, capturing 804 individuals of 64 species (Appendix Appendix ). Although the observed number of species did not show a tendency towards stabilization, curves showing species richness predicted by the MMMean estimator began to stabilize during the first 20 h of mist netting in both sites (Fig. 4). Five days accounted for 96 expected species in IT, but captures conducted during 3-day mist netting sections (63.3 net-h/m2) estimated 93 species while capturing 436 individuals (38% less than captured during five days). In MC, the MMMean estimator predicted 73 species along 5-day surveys; when analyzing the accumulation of 3-day mist netting sections (43.8 net-h/m2) we recorded 598 individuals (-34%) of 60 species, while estimating 72 species. The species netted in IT only after three days were: Amazilia versicolor, Antrhacothorax nigricollis, Crypturellus parvirostris, Empidonomus varius, Myiobius atricaudus, Sporophila angolensis, Thalurania glaucopis and Xenops rutilans; and those in MC were Pteroglossus bailoni, Myiarchus tyrannulus, Sirystes sibilator and Vireo olivaceus.
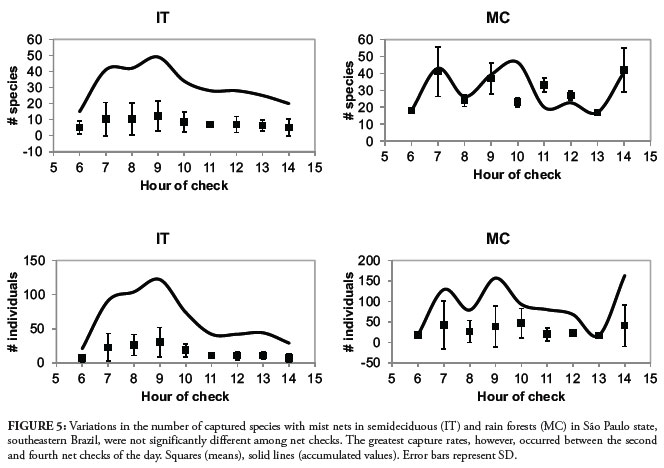
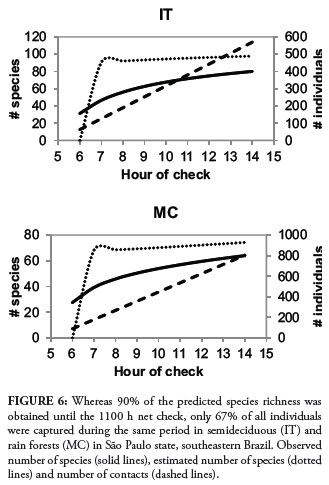
There were no significant differences among the numbers of hourly captures in species richness (F7-21 = 1.56, P = 0.203) or abundance (F7-21 = 1.85, P = 0.131) in IT or MC (Frichness5-10 = 0.78, P = 0.588; Fabundance5-10 = 0.76, P = 0.598). However, the total number of captures was highest between the second and forth checks, tended to drop until noon, and fluctuated unpredictably during the last two hours examined (Fig. 5). Although 90% of all species was captured until the 1100 h check, the number of individuals captured until this same hour corresponded to 67% of all netted birds (Fig. 6).
DISCUSSION
We found that point counting in these two Neotropical forests would not necessarily record most species and contacts when surveying birds with a set of four 20-min counts per day. Nearly 15% of all species and as many as 19 species were missed in IT after 17 h of censuses, while more than 90% of the observed species were detected in MC after 20 h. MMMean estimates did not stabilize in either site. As a consequence, the estimated number of species could increase with further sampling, altering the proportions of the estimated species richness. The fact that the first 5 min accounted for at least 63% of all species recorded during 20-min counts suggests that the length of a point count means a trade-off between the number of points carried out and the number of species detected at that point. As four points conducted per day covered little area, it took us four days to record more than 93% of all observed species. This problem could be controlled by increasing the number of point counts while reducing the length of the count. In a semideciduous forest fragment in Paraná state, southern Brazil, five 15-min point counts carried out during 3 days were enough to detect more than 90% of the observed number of species (Anjos, 2007). Esquivel & Peris (2008) detected 87% and 93% of the observed bird species richness with 5- and 10-min point counts, respectively, conducting seven point counts in a Paraguayan Atlantic forest.
For our study areas, the first 5 min of point counting detected significantly higher numbers of species and contacts. In addition, 5 min would detect 68% of the estimated species in MC and only 53% in IT. Esquivel & Peris (2008) also observed this trend in which the first 5 min detect more birds but are insufficient for surveying most Atlantic forest endemics. The decrease in detections during the last intervals was a common pattern as well. We suggest that to record most species it would be more fruitful to conduct more, albeit shorter, point counts, as also concluded by Anjos et al. (2010). This question, however, remains to be tested with specific sampling designs. Individual abundance will probably need longer counts as 86% of all contacts were detected only after 15 min. It is fortuitous to remember that longer counts will probably overestimate abundance because the chance of counting the same individual (and violating the assumption of "one individual = 1 contact") increases with time (Cimprich, 2009). It appears there is a trade-off between detecting most individuals and avoiding double counting.
In the United States, Siegel et al. (2001) concluded that a single visit would be enough to detect even high conservation-value species with point counts. This was partly true for our surveys. We recorded several endangered species according to the threatened avifauna of the state of São Paulo (Silveira et al., 2009) during one sample only, either during the breeding (Geotrygon violacea), or nonbreeding season (Pyroderus scutatus), or during two consecutive breeding seasons (Biatas nigropectus, Sporophila frontalis, S. falcirostris). Sporophila bamboo mast-seeding followers were overwhelmingly abundant during one breeding season but much less abundant or absent during the remaining seasons. Small samples in time and space may limit some inferences and counting on these occasions would compromise individual analyses when establishing conservation priorities due to the substantial fluctuation of abundance values throughout the seasons (Siegel et al. 2001). We therefore suggest sampling sites more than just once.
The predicted number of captured species was almost identical in IT and MC, even with lesser mist netting effort in the latter study site. Thus, a 3-day mist netting section (at least 9 h/day = 5 net-h/m2) was enough to accurately estimate species richness in these forests (Faaborg et al., 2004). The number of individuals, however, was reduced by at least 34% compared to five days of mist netting, compromising abundance estimation, which has also been described for small birds in Iberia (Arizaga et al., 2011). Mist net sampling detects secretive or rarely vocal species that are ineffectively sampled by visual-auditory censuses, and of non-territorial species for which some census techniques are inappropriate (Karr, 1981). This was true for several species exclusively recorded by nets, such as Geotrygon violacea. Nonetheless, only one secretive, non-endemic and not threatened species (M. atricaudus) was detected exclusively during 3-day mist netting sections, suggesting that a 3-day sampling effort will not detect conservation-value species in our study sites (Whitman et al., 1997; Derlindati & Caziani, 2005; Estades et al., 2006).
As species become more active during the first hours of the day (Blake, 1992), it would be expected that capture rates be greatest during the first net checks. However, although not significantly, it was possible to notice a trend in which the total number of species and individuals were highest between the second and fourth net checks. Peaks in capture rates until 1100 h have also been suggested for Atlantic forest birds (Mallet-Rodrigues & Noronha, 2003). As bird capture by nets is directly dependent on movement, poor capture rates during the first hours indicate birds remain still, singing in their perches. As we have no data on capture rates after 1400 h we believe the 0700-0900 h net checks is the best time of day to survey birds with mist nets in these locations (but see below), capturing most species and individuals. Early morning singing and a trend in greatest capture rates after 0700 h suggest birds sing conspicuously during the beginning of the day and start to move around a few hours after dawn. This is consistent with the inefficient foraging hypothesis, which states that the timing of dawn song is related to light availability (Kacelnik, 1979). As a result, birds become active at twilight when light levels are insufficient for foraging, yet adequate for social communication as well as predator avoidance. Only then do they start to move (Berg et al., 2006). Since it is known that birds have a second activity peak during the last hours of daylight (Blake, 1992), our results could have been different if we had closed our nets after 1700 h, for example.
Although net checks until 1100 h cumulatively provided 90% of all species captured, we believe this method requires too much effort (considering logistics and number of personnel for banding), to be used for only 5 h a day. This is further corroborated by the fact that these first five checks accounted for 67% of all captured individuals. As long as 3 days are sampled we suggest that nets should be kept open from 0600 h to at least 1400 h. It is possible that even after 8 h of mist netting some new species may still be captured, which has led some to advocate a combination of methods to inventory bird communities (Rappole et al., 1998; Stiles & Rosseli, 1998; Somenzari et al., 2011). Protocols must be defined in order to accomplish the expected results without compromising survey quality (Whitman, 2004; Banks-Leite et al., 2011) and studies should be carefully thought out before nets are set up, to ensure that the sampling design and the estimated sample size would allow study objectives to be met (Ralph et al., 2004). The information needed to make that decision would likely vary from place to place, so it should be examined in a variety of systems. For the locations in which we carried out bird surveys, we found that 5-min point counting would record most species than the following minutes and mist netting for 5 h during at least 3 consecutive days would capture most species and individuals, both without compromising species richness estimation.
The fact that we considered seasons as replicas could have masked the effects of time of day. Although only a long-term study with at least three temporal replicas would reliably account for seasonal patterns in bird detection, there is no reason to expect birds would start to move unpredictably (such as more captures at 1200 h in cold winter days as opposed to hot summer days) after the first morning hours. The peak in which birds are tangled in mist nets may vary a few hours according to each season simply because summer days have more hours of light than winter days or due to lower temperatures in winter mornings. It is of special interest to know the times of day (early mornings or late afternoons) in which capture rates are greatest. Thus, known peaks in captures may motivate researchers to use mist nets more frequently, or even to investigate if some taxonomic groups or guilds of birds are more likely to be netted during a specific hour of the day. It is also imperative to know if there is seasonal effect on capture rates and, more importantly, if there are interactions between time of day and season.
ACKNOWLEDGEMENTS
We thank Suzano Papel e Celulose for allowing access to the facilities at Fazenda Entre Rios and Fazenda Montes Claros. CEMAVE supplied our banding rings, and IdeaWild supported us with field equipment. LFS is supported by a productivity fellowship from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). This work would have been impossible without several ornithologists who helped us with bird banding during our field trips: B. Rennó, F. Bocalini, G. Del-Rio, L. Moreira-Lima and R. Marcondes. Three anonymous reviewers considerably improved the first versions of this manuscript.
REFERENCES
Anjos, L. 2007. A eficiência do método de amostragem por pontos de escuta na avaliação da riqueza de aves. Revista Brasileira de Ornitologia, 15:239-243.
Anjos, L.; Volpato, G.H.; Mendonça, L.B.; Serafini, P.; Lopes, E.V.; Boçon, R.; Silva, E.S. & Bisheimer, M.V. 2010. Técnicas de levantamento quantitativo de aves em ambiente florestal: uma análise comparativa baseada em dados empíricos. In: Matter, S.; Straube, F.C.; Accordi, I.; Piacentini, V.Q.; Cândido-Jr, J.F. (Orgs.). Ornitologia e Conservação: ciência aplicada, técnicas de pesquisa e levantamento. Technical Books Editora, Rio de Janeiro. p. 63-76.
Arizaga, J.; Deán, J.I.; Vilches, A.; Alonso, D. & Mendiburu, A. 2011. Monitoring communities of small birds: a comparison between mist-netting and counting. Bird Study, 58:291-301.
Banks-Leite, C.; Ewers, R.M.; Pimentel, R.G. & Metzger, J.P. 2011. Decision on temporal sampling protocol influence the detection of ecological patterns. Biotropica, 44:378-385.
Berg, K.S; Brumfield, R.T. & Apanius, V. 2006. Phylogenetic and ecological determinants of the Neotropical dawn chorus. Proceedings of the Royal Society B. Biological Sciences, 273:999-1005.
Betini, G.S. 2001. Amostragem de aves por pontos numa floresta estacional semidecidual, São Paulo, Brasil. (Dissertação de Mestrado). Escola Superior de Agricultura Luiz de Queiroz, Piracicaba.
Bibby, C.J.; Burgess, N.D.; Hill, D.A. & Mustoe, S.H. 2000. Bird census techniques. Academic Press, London.
Blake, J.G. 1992. Temporal variation in point counts of birds in a lowland wet forest in Costa Rica. Condor, 94:265-275.
Blake, J.G. & Loiselle, B.A. 2000. Diversity of birds along an elevational gradient in the Cordillera Central, Costa Rica. The Auk, 117:663-686.
Blake, J.G. & Loiselle, B.A. 2001. Bird assemblages in second growth and old-growth forests, Costa Rica: perspectives from mist-nets and point counts. The Auk, 118:304-326.
Cavarzere, V. 2013. Does the reproductive season account for more records of birds in a marked seasonal climate landscape in the state of São Paulo, Brazil? Papéis Avulsos de Zoologia, 53:253-260.
Cavarzere, V.; Moraes, G.P.; Dalbeto, A.; Maciel, F.G. & Donatelli, R.J. 2011. Birds from cerradão woodland, an overlooked forest of the Cerrado Region, Brazil. Papéis Avulsos de Zoologia, 51:259-273.
CBRO Comitê Brasileiro de Registros Ornitológicos. 2011. Lista das aves do Brasil. Available at <www.cbro.org.br>. Access In: 25/May/2013.
Chase, M.K.; Nur, N. & Geupel, G.R. 1997. Survival, productivity, and abundance in a Wilson's Warbler population. The Auk, 114:354-366.
Cimprich, D.A. 2009. Effect of count duration on abundance estimates of Black-capped Vireos. Journal of Field Ornithology, 80:94-100.
Colwell, R.K. 2009. EstimateS: statistical estimation of species richness and shared species from samples. Version 8.2. User's Guide and application. Available at: http://viceroy.eeb.uconn.edu/EstimateS. Access in 06/20/2010.
Derlindati, E.J. & Caziani, S.M. 2005. Using canopy and understory mist nets and point counts to study bird assemblages in Chaco forests. Wilson Bulletin, 117:92-99.
Develey, P.F. 2004. Efeitos da fragmentação e do estado de conservação da floresta na diversidade de aves de Mata Atlântica. (Ph.D. dissertation). Universidade de São Paulo, São Paulo. Dunn, E.H. & Ralph, J. 2004. Use of mist nets as a tool for bird population monitoring. Studies in Avian Biology, 6:1-6.
Esquivel, M.A. & Peris, S. 2008. Influence of time of day, duration and number of counts in point count sampling of birds in an Atlantic Forest of Paraguay. Ornitologia Neotropical, 19:229-242.
Estades, C.F.; Escobar, M.A.H.; Tamasevib, J.A.; Vukasovic, M.A. & Páez, M. 2006. Mist nets versus point counts in the estimation of forest bird abundances in south-central Chile. Ornitologia Neotropical, 17:203-212.
Faaborg, J.; Arendt, W.J. & Dugger, K.M. 2004. Bird population studies in Puerto Rico using mist nets: general patterns and comparisons with point counts. Studies in Avian Biology, 29:144-150.
Gram, W.K. & Faaborg, J. 1997. The distribution of Neotropical migrant birds wintering in the El Cielo Biosphere Reserve, Tamaulipas, Mexico. Condor, 99:658-670.
Herzog, S.K.; Kessler, M. & Cahill, T.M. 2002. Estimating species richness of tropical bird communities from rapid assessment data. The Auk, 119:749-769.
Jenni, L.; Leuenberger, M. & Rampazzi, F. 1996. Capture efficiency of mist-nets with comments on their role in the assessment of passerine habitat use. Journal of Field Ornithology, 67:263-274.
Kacelnik, A. 1979. The foraging efficiency of Great Tits (Parus major L.) in relation to light intensity. Animal Behavior, 27:237-241.
Karr, J.R. 1981. Surveying birds with mist nets. Studies in Avian Biology, 6:62-67.
Keating, K.A. & Quinn, J.F. 1998. Estimating species richness: The Michaelis-Menten model revisited. Oikos, 81:411-416.
Lövei, G.L.; Csörgõ, T. & Miklay, G. 2001. Capture efficiency of small birds by mist nets. Ornis Hungarica, 11:19-25.
MacArthur, R.H. & MacArthur, A.T. 1974. On the use of mist nets for population studies of birds. Proceedings of the National Academy of Sciences USA, 71:3230-3233.
Mackenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A. & Langtimm, C. 2002. Estimating site occupancy rates when detection probabilities are less than one. Ecology, 83:2248-2255.
Mallet-Rodrigues, F. & Noronha, M.L.M. 2003. Variação na taxa de captura de passeriformes em um trecho de Mata Atlântica de encosta, no sudeste do Brasil. Ararajuba, 11:111-118.
Martin, T.E.; Blackburn, G.A. & Simcox, W. 2010. An assessment of the effectiveness of two methods in describing a Neotropical cloud forest bird community. Ornitologia Neotropical, 21:131-147.
Pagen, R.W.; Thompson, F.R. & Burhans, D.E. 2002. A comparison of point count and mist-net detections of songbirds by habitat and time of season. Journal of Field Ornithology, 73:53-59.
Parker, T.A.; Stotz, D.F. & Fitzpatrick, J.W. 1996. Ecological and distributional databases. In: Stotz, D.F.; Fitzpatrick, J.W.; Parker, T.A. & Moskovits, D.K. (Eds.). Neotropical birds: ecology and conservation. The University of Chicago Press, Chicago. p. 116-436.
Piratelli, A.J. 2003. Mesh size and bird capture rates in Mato Grosso do Sul state, Brazil. Brazilian Journal of Biology, 63:105-111.
Poulin, B.; Lefebvre, G. & Pilard, P. 2000. Quantifying the breeding assemblage of reedbed passerines with mistnet and point count surveys. Journal of Field Ornithology, 71:443-454.
Ralph, C.J. & Dunn, E.H. 2004. Monitoring bird populations using mist nets. Studies in Avian Biology, 29:1-211.
Ralph, C.J.; Dunn, E.H.; Peach, W.J. & Handel, C.M. 2004. Recommendations for the use of mist nets for inventory and monitoring of bird populations. Studies in Avian Biology, 29:187-196.
Rappole, J.H.; Winker, K. & Powell, G.V.N. 1998. Migratory bird habitat use in southern Mexico: mist nets versus point counts. Journal of Field Ornithology, 69:63-643.
Remsen Jr., J.V. & Good, D.A. 1996. Misuse of data from mistnet captures to assess relative abundance in bird populations. The Auk, 113:381-398.
Siegel, R.B.; Desante, D.F. & Nott, M.P. 2001. Using point counts to establish conservation priorities: how many visits are optimal? Journal of Field Ornithology, 72:228-235.
Silva, J.M.C. 1995. Birds of the Cerrado region, South America. Steenstrupia, 21:69-92.
Silveira, L.F.; Benedicto, G.A.; Schunck, F. & Sugieda, A.M. 2009. Aves. In: Bressan, P.M.; Kierulff, M.C. & Sugieda, A.M. (Orgs.). Fauna ameaçada de extinção no Estado de São Paulo: Vertebrados. São Paulo, Fundação Parque Zoológico de São Paulo e Secretaria do Meio Ambiente.
Somenzari, M.; Silveira, L.F.; Piacentini, V.Q.; Rego, M.A.; Schunck, F. & Cavarzere, V. 2011. Birds of an Amazonia- Cerrado ecotone in southern Pará, Brazil, and the efficiency of associating multiple methods in avifaunal inventories. Revista Brasileira de Ornitologia, 19:260-275.
SPSS Statistical Package for Social Sciences. 2004. Version 13.0. SPSS Inc, Chicago, IL.
Stiles, F.G. & Rosselli, L. 1998. Inventario de las aves de un bosque alto andino: comparación de dos métodos. Caldasia, 20:29-43.
Thompson, F.R.; Burhans, D.E. & Root, B. 2002. Effects of point count protocol on bird abundance and variability estimates and power to detect population trends. Journal of Field Ornithology, 73:141-150.
Vergara, P.M.; Jiménez, J.E. & Schllater, R.P. 2010. Effective point-count duration for estimating bird species richness in Chilean forests. Zoological Studies, 49:381-391.
Verner, J. 1985. Assessment of counting techniques. Current Ornithology, 2:247-302.
Viani, R.A.G. & Rodrigues, R.R. 2007. Sobrevivência em viveiro de mudas de espécies nativas retiradas da regeneração natural de remanescente florestal. Pesquisa Agropecuária Brasileira, 48:1067-1075.
Vielliard, J.M.C. & Silva, W.R. 1990. Nova metodologia de levantamento quantitativo da avifauna e primeiros resultados no interior do Estado de São Paulo, Brasil. In: Encontro Nacional dos Anilhadores de Aves, 4º. Anais. Recife. p. 117-151.
Volpato, G.H.; Lopes, E.V.; Mendonça, L.B.; Boçon, R.; Bisheimer, M.V.; Serafini, P.P. & Anjos, L. 2009. The use of the point count method for bird survey in the Atlantic Forest. Zoologia, 26:74-78.
Wallace, G.E.; Alonso, H.G.; McNicholl, H.G.M.K.; Batista, D.R.; Prieto, R.O.; Sosa, A.L.; Oria, B.S. & Wallace, E.A.H. 1996. Winter surveys of forest-dwelling Neotropical migrant and resident birds in three regions of Cuba. Condor, 98:745-768.
Williams, B.K.; Nichols, J.D. & Conroy, M.J. 2002. Analysis and management of animal populations. Academic Press, San Diego.
Whitman, A.A.; Hagan III, J.M. & Brokaw, N.V.L. 1997. A comparison of two bird survey techniques used in a subtropical forest. Condor, 99:955-965.
Whitman, A.A. 2004. Use of mist nets for study of Neotropical bird communities. Studies in Avian Biology, 29:161-167
Aceito em: 28/05/2013
Impresso em: 30/09/2013
APPENDIX
Bird species recorded with point counts (PC) and mist nets (MN) in two Atlantic forest fragments of São Paulo state, southeastern Brazil. IT = semideciduous forest, MC = rain forest. Atl: Atlantic forest endemic species (Parker et al., 1996, except for those in Cavarzere et al., 2011), cer: cerrado endemic species (Silva, 1995). Species taxonomy is according to the Comitê Brasileiro de Registros Ornitológicos (CBRO, 2011).
Appendix
- Anjos, L. 2007. A eficięncia do método de amostragem por pontos de escuta na avaliaçăo da riqueza de aves. Revista Brasileira de Ornitologia, 15:239-243.
- Anjos, L.; Volpato, G.H.; Mendonça, L.B.; Serafini, P.; Lopes, E.V.; Boçon, R.; Silva, E.S. & Bisheimer, M.V. 2010. Técnicas de levantamento quantitativo de aves em ambiente florestal: uma análise comparativa baseada em dados empíricos. In: Matter, S.; Straube, F.C.; Accordi, I.; Piacentini, V.Q.; Cândido-Jr, J.F. (Orgs.). Ornitologia e Conservaçăo: cięncia aplicada, técnicas de pesquisa e levantamento. Technical Books Editora, Rio de Janeiro. p. 63-76.
- Arizaga, J.; Deán, J.I.; Vilches, A.; Alonso, D. & Mendiburu, A. 2011. Monitoring communities of small birds: a comparison between mist-netting and counting. Bird Study, 58:291-301.
- Banks-Leite, C.; Ewers, R.M.; Pimentel, R.G. & Metzger, J.P. 2011. Decision on temporal sampling protocol influence the detection of ecological patterns. Biotropica, 44:378-385.
- Berg, K.S; Brumfield, R.T. & Apanius, V. 2006. Phylogenetic and ecological determinants of the Neotropical dawn chorus. Proceedings of the Royal Society B. Biological Sciences, 273:999-1005.
- Betini, G.S. 2001. Amostragem de aves por pontos numa floresta estacional semidecidual, Săo Paulo, Brasil. (Dissertaçăo de Mestrado). Escola Superior de Agricultura Luiz de Queiroz, Piracicaba.
- Bibby, C.J.; Burgess, N.D.; Hill, D.A. & Mustoe, S.H. 2000. Bird census techniques. Academic Press, London.
- Blake, J.G. 1992. Temporal variation in point counts of birds in a lowland wet forest in Costa Rica. Condor, 94:265-275.
- Blake, J.G. & Loiselle, B.A. 2000. Diversity of birds along an elevational gradient in the Cordillera Central, Costa Rica. The Auk, 117:663-686.
- Blake, J.G. & Loiselle, B.A. 2001. Bird assemblages in second growth and old-growth forests, Costa Rica: perspectives from mist-nets and point counts. The Auk, 118:304-326.
- Cavarzere, V. 2013. Does the reproductive season account for more records of birds in a marked seasonal climate landscape in the state of Săo Paulo, Brazil? Papéis Avulsos de Zoologia, 53:253-260.
- Cavarzere, V.; Moraes, G.P.; Dalbeto, A.; Maciel, F.G. & Donatelli, R.J. 2011. Birds from cerradăo woodland, an overlooked forest of the Cerrado Region, Brazil. Papéis Avulsos de Zoologia, 51:259-273.
- CBRO Comitę Brasileiro de Registros Ornitológicos. 2011. Lista das aves do Brasil. Available at <www.cbro.org.br>. Access In: 25/May/2013.
- Chase, M.K.; Nur, N. & Geupel, G.R. 1997. Survival, productivity, and abundance in a Wilson's Warbler population. The Auk, 114:354-366.
- Cimprich, D.A. 2009. Effect of count duration on abundance estimates of Black-capped Vireos. Journal of Field Ornithology, 80:94-100.
- Colwell, R.K. 2009. EstimateS: statistical estimation of species richness and shared species from samples. Version 8.2. User's Guide and application. Available at: http://viceroy.eeb.uconn.edu/EstimateS Access in 06/20/2010.
- Derlindati, E.J. & Caziani, S.M. 2005. Using canopy and understory mist nets and point counts to study bird assemblages in Chaco forests. Wilson Bulletin, 117:92-99.
- Develey, P.F. 2004. Efeitos da fragmentaçăo e do estado de conservaçăo da floresta na diversidade de aves de Mata Atlântica. (Ph.D. dissertation). Universidade de Săo Paulo, Săo Paulo.
- Dunn, E.H. & Ralph, J. 2004. Use of mist nets as a tool for bird population monitoring. Studies in Avian Biology, 6:1-6.
- Esquivel, M.A. & Peris, S. 2008. Influence of time of day, duration and number of counts in point count sampling of birds in an Atlantic Forest of Paraguay. Ornitologia Neotropical, 19:229-242.
- Estades, C.F.; Escobar, M.A.H.; Tamasevib, J.A.; Vukasovic, M.A. & Páez, M. 2006. Mist nets versus point counts in the estimation of forest bird abundances in south-central Chile. Ornitologia Neotropical, 17:203-212.
- Faaborg, J.; Arendt, W.J. & Dugger, K.M. 2004. Bird population studies in Puerto Rico using mist nets: general patterns and comparisons with point counts. Studies in Avian Biology, 29:144-150.
- Gram, W.K. & Faaborg, J. 1997. The distribution of Neotropical migrant birds wintering in the El Cielo Biosphere Reserve, Tamaulipas, Mexico. Condor, 99:658-670.
- Herzog, S.K.; Kessler, M. & Cahill, T.M. 2002. Estimating species richness of tropical bird communities from rapid assessment data. The Auk, 119:749-769.
- Jenni, L.; Leuenberger, M. & Rampazzi, F. 1996. Capture efficiency of mist-nets with comments on their role in the assessment of passerine habitat use. Journal of Field Ornithology, 67:263-274.
- Kacelnik, A. 1979. The foraging efficiency of Great Tits (Parus major L.) in relation to light intensity. Animal Behavior, 27:237-241.
- Karr, J.R. 1981. Surveying birds with mist nets. Studies in Avian Biology, 6:62-67.
- Keating, K.A. & Quinn, J.F. 1998. Estimating species richness: The Michaelis-Menten model revisited. Oikos, 81:411-416.
- Lövei, G.L.; Csörgő, T. & Miklay, G. 2001. Capture efficiency of small birds by mist nets. Ornis Hungarica, 11:19-25.
- MacArthur, R.H. & MacArthur, A.T. 1974. On the use of mist nets for population studies of birds. Proceedings of the National Academy of Sciences USA, 71:3230-3233.
- Mackenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A. & Langtimm, C. 2002. Estimating site occupancy rates when detection probabilities are less than one. Ecology, 83:2248-2255.
- Mallet-Rodrigues, F. & Noronha, M.L.M. 2003. Variaçăo na taxa de captura de passeriformes em um trecho de Mata Atlântica de encosta, no sudeste do Brasil. Ararajuba, 11:111-118.
- Martin, T.E.; Blackburn, G.A. & Simcox, W. 2010. An assessment of the effectiveness of two methods in describing a Neotropical cloud forest bird community. Ornitologia Neotropical, 21:131-147.
- Pagen, R.W.; Thompson, F.R. & Burhans, D.E. 2002. A comparison of point count and mist-net detections of songbirds by habitat and time of season. Journal of Field Ornithology, 73:53-59.
- Parker, T.A.; Stotz, D.F. & Fitzpatrick, J.W. 1996. Ecological and distributional databases. In: Stotz, D.F.; Fitzpatrick, J.W.; Parker, T.A. & Moskovits, D.K. (Eds.). Neotropical birds: ecology and conservation. The University of Chicago Press, Chicago. p. 116-436.
- Piratelli, A.J. 2003. Mesh size and bird capture rates in Mato Grosso do Sul state, Brazil. Brazilian Journal of Biology, 63:105-111.
- Poulin, B.; Lefebvre, G. & Pilard, P. 2000. Quantifying the breeding assemblage of reedbed passerines with mistnet and point count surveys. Journal of Field Ornithology, 71:443-454.
- Ralph, C.J. & Dunn, E.H. 2004. Monitoring bird populations using mist nets. Studies in Avian Biology, 29:1-211.
- Ralph, C.J.; Dunn, E.H.; Peach, W.J. & Handel, C.M. 2004. Recommendations for the use of mist nets for inventory and monitoring of bird populations. Studies in Avian Biology, 29:187-196.
- Rappole, J.H.; Winker, K. & Powell, G.V.N. 1998. Migratory bird habitat use in southern Mexico: mist nets versus point counts. Journal of Field Ornithology, 69:63-643.
- Remsen Jr., J.V. & Good, D.A. 1996. Misuse of data from mistnet captures to assess relative abundance in bird populations. The Auk, 113:381-398.
- Siegel, R.B.; Desante, D.F. & Nott, M.P. 2001. Using point counts to establish conservation priorities: how many visits are optimal? Journal of Field Ornithology, 72:228-235.
- Silva, J.M.C. 1995. Birds of the Cerrado region, South America. Steenstrupia, 21:69-92.
- Silveira, L.F.; Benedicto, G.A.; Schunck, F. & Sugieda, A.M. 2009. Aves. In: Bressan, P.M.; Kierulff, M.C. & Sugieda, A.M. (Orgs.). Fauna ameaçada de extinçăo no Estado de Săo Paulo: Vertebrados. Săo Paulo, Fundaçăo Parque Zoológico de Săo Paulo e Secretaria do Meio Ambiente.
- Somenzari, M.; Silveira, L.F.; Piacentini, V.Q.; Rego, M.A.; Schunck, F. & Cavarzere, V. 2011. Birds of an Amazonia- Cerrado ecotone in southern Pará, Brazil, and the efficiency of associating multiple methods in avifaunal inventories. Revista Brasileira de Ornitologia, 19:260-275.
- SPSS Statistical Package for Social Sciences. 2004. Version 13.0. SPSS Inc, Chicago, IL.
- Stiles, F.G. & Rosselli, L. 1998. Inventario de las aves de un bosque alto andino: comparación de dos métodos. Caldasia, 20:29-43.
- Thompson, F.R.; Burhans, D.E. & Root, B. 2002. Effects of point count protocol on bird abundance and variability estimates and power to detect population trends. Journal of Field Ornithology, 73:141-150.
- Vergara, P.M.; Jiménez, J.E. & Schllater, R.P. 2010. Effective point-count duration for estimating bird species richness in Chilean forests. Zoological Studies, 49:381-391.
- Verner, J. 1985. Assessment of counting techniques. Current Ornithology, 2:247-302.
- Viani, R.A.G. & Rodrigues, R.R. 2007. Sobrevivęncia em viveiro de mudas de espécies nativas retiradas da regeneraçăo natural de remanescente florestal. Pesquisa Agropecuária Brasileira, 48:1067-1075.
- Vielliard, J.M.C. & Silva, W.R. 1990. Nova metodologia de levantamento quantitativo da avifauna e primeiros resultados no interior do Estado de Săo Paulo, Brasil. In: Encontro Nacional dos Anilhadores de Aves, 4ş. Anais. Recife. p. 117-151.
- Volpato, G.H.; Lopes, E.V.; Mendonça, L.B.; Boçon, R.; Bisheimer, M.V.; Serafini, P.P. & Anjos, L. 2009. The use of the point count method for bird survey in the Atlantic Forest. Zoologia, 26:74-78.
- Wallace, G.E.; Alonso, H.G.; McNicholl, H.G.M.K.; Batista, D.R.; Prieto, R.O.; Sosa, A.L.; Oria, B.S. & Wallace, E.A.H. 1996. Winter surveys of forest-dwelling Neotropical migrant and resident birds in three regions of Cuba. Condor, 98:745-768.
- Williams, B.K.; Nichols, J.D. & Conroy, M.J. 2002. Analysis and management of animal populations. Academic Press, San Diego.
- Whitman, A.A.; Hagan III, J.M. & Brokaw, N.V.L. 1997. A comparison of two bird survey techniques used in a subtropical forest. Condor, 99:955-965.
- Whitman, A.A. 2004. Use of mist nets for study of Neotropical bird communities. Studies in Avian Biology, 29:161-167
Appendix

Publication Dates
-
Publication in this collection
25 Sept 2013 -
Date of issue
2013
History
-
Received
28 May 2013 -
Accepted
30 Sept 2013