Resúmenes
El esqueleto axial es una estructura compleja de origen cuticular, formada a partir de las láminas esclerotizadas de los esternitos y pleuritos torácicos así como por sus invaginaciones internas. Los caracteres de la parte ventral del esqueleto axial, que se reflejan externamente en las líneas de sutura del esternón torácico, son criterios importantes en el actual sistema de supragenerico de Trichodactylidae: (Trichodactylinae y Dilocarcininae (Dilocarcinini + Valdiviini)). Con el objetivo de probar esa hipótesis de clasificación fue realizado el estudio anatómico del esqueleto axial de 43 individuos adultos, tanto machos como hembras, pertenecientes a 17 especies de 10 géneros de Trichodactylidae. Para la preparación del material, los individuos fueron diseccionados y sus esqueletos sumergidos, por un periodo de 24 a 48 horas, en una solución caliente de Hidróxido de Potasio (KOH). El análisis anatómico permitió describir un grupo de variaciones en la forma, orientación y conformación del esternón torácico, techo pleural, fragmas ventrales y dorsales, así como de la lámina de unión. De acuerdo con esas características las especies estudiadas fueron reunidas en tres grupos, siendo esas agrupaciones consistentes con la actual propuesta de clasificación de Trichodactylidae en subfamilias y tribus. El presente trabajo resalta la importancia informativa de la morfología del esqueleto axial para la taxonomía de Trichodactylidae.
Trichodactylidae; cangrejos de agua dulce; esqueleto axial; lámina de unión; sella turcica.
The axial skeleton is a complex structure of cuticular origin, formed by sclerotic plates of thoracic sternite and pleurites as well as its internal invaginations. The ventral characters of the axial skeleton, reflected externally in the suture lines of the thoracic sternum, are an important classification criteria for the current suprageneric system of Trichodactylidae: (Trichodactylinae & Dilocarcininae (Dilocarcinini + Valdiviini)). In order to test this hypothesis, the internal and external characters of the axial skeleton of 42 adult crabs belonging to 17 species and 10 genera were examined. For the material preparation the crabs were dissected and their skeletons submerged in a hot solution of potassium hydroxide (KOH) for a period of 24 to 48 hours. Variations in the shape, orientation and conformation of the thoracic sternum, pleural roof, ventral and dorsal phragmas and junction plate were described from anatomical preparations. Based on these characteristics, the species were separated into three groups, which are consistent with the proposed classification of Trichodactylidae into subfamilies and tribes. This study highlights the importance of the morphological characteristics of the axial skeleton for Trichodactylidae taxonomy.
Trichodactylidae; freshwater crabs; axial skeleton; junction plate; sella turcica.
INTRODUCCIÓN
A diferencia de otros grupos de crustáceos, en los decápodos se destaca la presencia de un complejo sistema esquelético formado a partir de placas esclerotizadas de origen cuticular. Funcionalmente esta armazón provee puntos de anclaje para la inserción de la musculatura de los apéndices torácicos (Drach, 1950Drach, P. 1950. Les étapes évolutives de l'endosquelette chez lês Crustacés Décapodes. Comptes rendus hebdomadaires des séances de l'Académie des Sciences, 231(25):1563-1565.; Bitsch & Bitsch, 2002Bitsch, C. & Bitsch, J. 2002. The endoskeletal structures in arthropods: citology, morphology and evolution. Arthropod Structure & Development, 30:159-177.). En conjunto esta estructura recibe el nombre de esqueleto axial (Secretan, 1980Secretan, S. 1980. Le plan de base du "squelette axial" d'un Crustacé décapode macroure et sa terminologie. Comptes rendus hebdomadaires des seánces de l'Académie des Sciences, série D, 291:877-880.; Secretan-Rey, 2002Secretan-Rey, S. 2002. Monographie du squelette axial de Nephrops norvegicus (Linné, 1758). Zoosystema, 24(1):81-176.; Guinot et al., 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.). En Brachyura, conforme con el proceso de "carcinisación" y reorganización de los órganos internos, las estructuras del esqueleto axial sufrieron marcadas modificaciones tanto en su orientación como en su forma de unión, dando origen a un esqueleto central rígido y metamerizado en el que todos sus elementos están fusionados (Drach 1950Drach, P. 1950. Les étapes évolutives de l'endosquelette chez lês Crustacés Décapodes. Comptes rendus hebdomadaires des séances de l'Académie des Sciences, 231(25):1563-1565.; Secretan 1998, 2002Secretan, S. 2002. La lame de jonction, base de l'organisation phragmale des Décapodes Brachyoures. Crustaceana, 75(3-4):637-641.).
A medida que los estudios sobre la morfología del esqueleto axial aumentaron, se evidenció la importancia de esta estructura como herramienta en la sistemática de los braquiuros (Drach, 1950Drach, P. 1950. Les étapes évolutives de l'endosquelette chez lês Crustacés Décapodes. Comptes rendus hebdomadaires des séances de l'Académie des Sciences, 231(25):1563-1565.; Secretan, 1998Secretan, S. 1998. The sella turcica of crabs and the endophragmal system of decapods. Journal of Natural History, 32:1753-1767., Secretan, 2002Secretan, S. 2002. La lame de jonction, base de l'organisation phragmale des Décapodes Brachyoures. Crustaceana, 75(3-4):637-641.; Secretan-Rey, 2002Secretan-Rey, S. 2002. Monographie du squelette axial de Nephrops norvegicus (Linné, 1758). Zoosystema, 24(1):81-176.; Guinot, 1979Guinot, D. 1979. Données nouvelles sur la morphologie, la phylogenèse et la taxonomie des Crustacés Décapodes Brachyoures. Mémories du Muséum National d'Historie Naturelle, série A, Zoologie, 112: 1-354.; Guinot et al., 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.). De acuerdo con (Guinot 1979Guinot, D. 1979. Données nouvelles sur la morphologie, la phylogenèse et la taxonomie des Crustacés Décapodes Brachyoures. Mémories du Muséum National d'Historie Naturelle, série A, Zoologie, 112: 1-354.:264): "... le plan général de la charpente endophragmale évolue dans une direction déterminée à l'intérieur de chaque lignée et, chose fort intéressante pour le taxonomiste, il semble relativement peu affecté par les particularités du mode de vie de charque espèce. Une restructuration complète ne paraît pas être possible par la seule influence des phénomènes adaptatifs.".
A nivel externo, la configuración del esqueleto axial es parcialmente reflejada en las líneas de sutura que atraviesan la superficie ventral del esternón torácico. (Magalhães & Türkay 1996Magalhães, C. & Türkay M. 1996. Taxonomy of the Neotropical freshwater crab family Trichodactylidae I. The generic system with description of some new genera (Crustacea: Decapoda: Brachyura). Senckenbergiana biologica, 75:63-95.) emplearon caracteres de estas líneas de sutura como criterio taxonómico en su propuesta de clasificación supragenerica en Trichodactylidae.
Trichodactylidae es una familia de braquiuros que habitan ambientes exclusivamente de agua dulce en el Neotrópico. Actualmente esta familia comprende 49 especies y subespecies (Ng et al., 2008Ng, P.K.L.; Guinot, D.& Davie, P.J.F. 2008. Systema Brachyurorum: Part 1. An annotated checklist of extant brachyuran crabs of the world. The Raffles Bulletin of Zoology, 17:1-286.), con distribución en México y América del Sur, en las planicies fluviales y costeras de los ríos Amazonas, Paraguay, Paraná, Orinoco, Magdalena y Maracaibo (Rodríguez, 1992Rodríguez, G. 1992. The freshwater crabs of America. Family Trichodactylidae and supplement to the family Pseudothelphusidae. Faune Tropicale, 31:1-189.).
El primer sistema de clasificación taxonómica de la familia fue propuesto por (Rathbun 1906Rathbun, M.J. 1906. Le crabs d'eau douce (Potamidae). Nouvelles Archives du Museum d'Histoire Naturelle de Paris, 8(4):33-122.), quien utilizo como criterio de clasificación la forma del caparazón. Posteriormente y de forma independiente, (Pretzmann 1968Pretzmann, G. 1968. Die Familie Trichodactylidae (Milne Edwards 1853) Smith 1870 (Vorläufige Mitteilung). Entomologisches Nachrichtenblatt, 15(7-8):70-76.) y (Bott 1969Bott, R. 1969. Die Süsswasserkrabben Süd-Amerikas und ihre Stammesgeschichte. Eine Revision der Trichodactylidae und der Pseudothelphusidae östlich der Anden (Crustacea, Decapoda). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 518:1-94.) propusieron modificaciones a esta clasificación con base en caracteres del primer gonopodo masculino, sin embargo sus interpretaciones diferentes originaron clasificaciones confusas y ambiguas (Magalhães, 2003Magalhães, C. 2003. Famílias Pseudothelpusidae e Trichodactylidae. In: Melo, G.A.S. Manual de identificação dos Crustácea Decápode de água doce do Brasil. São Paulo, Editora Loyola. p. 143-287.). (Rodríguez 1992Rodríguez, G. 1992. The freshwater crabs of America. Family Trichodactylidae and supplement to the family Pseudothelphusidae. Faune Tropicale, 31:1-189.) propuso el primer sistema supragenerico para la familia, basado en caracteres del primer apéndice sexual masculino y del abdomen; en él se reconocen 10 géneros agrupados en tres tribus y dos subfamlias. Posteriormente, (Magalhães & Türkay 1996Magalhães, C. & Türkay M. 1996. Taxonomy of the Neotropical freshwater crab family Trichodactylidae I. The generic system with description of some new genera (Crustacea: Decapoda: Brachyura). Senckenbergiana biologica, 75:63-95.) propusieron una nueva clasificación taxonómica en la que agrupan las especies de la familia en 15 géneros, dos tribus y dos subfamilias: (Trichodactylinae + Dilocarcininae (Valdiviini + Dilocarcinini)). A diferencia de (Rodríguez 1992Rodríguez, G. 1992. The freshwater crabs of America. Family Trichodactylidae and supplement to the family Pseudothelphusidae. Faune Tropicale, 31:1-189.), los autores no reconocen la tribu Holtuisini, transfiriendo a Silviocarcinus para Valdiviini, y a Zilchiopsis de Dilocarcinini para Valdiviini. Actualmente este es el sistema de clasificación aceptado (Sternberg, 1997Sternberg, R. von. 1997. Cladistics of the freshwater crab family Trichodactylidae (Crustacea: Decapoda): appraising the reappraisal. Journal of Comparative Biology, 2(1):49-62.; Ng et al., 2008Ng, P.K.L.; Guinot, D.& Davie, P.J.F. 2008. Systema Brachyurorum: Part 1. An annotated checklist of extant brachyuran crabs of the world. The Raffles Bulletin of Zoology, 17:1-286.).
Desde el trabajo de (Magalhães & Türkay 1996Magalhães, C. & Türkay M. 1996. Taxonomy of the Neotropical freshwater crab family Trichodactylidae I. The generic system with description of some new genera (Crustacea: Decapoda: Brachyura). Senckenbergiana biologica, 75:63-95.), otros autores han realizado observaciones sobre el esqueleto axial en Trichodactyidae. (Sternberg & Cumberlidge 2003Sternberg, R. von. & Cumberlidge, N. 2003. Autapomorphies of the endophragmal system in trichodactilyd freshwater crabs (Crustacea: Decapoda: Eubrachyura). Journal of Morphology, 256:23-28.) describen un grupo de autapomorfias, siendo la más particular de ellas, la fuerte reducción de la sella turcica. (Juares 2008Juares, A. 2008. Conexões morfológicas entre carapaça e esqueleto endofragmal nos caranguejos de água doce Trichodactylidae Milne-Edwards, 1853 (Crustacea: Decapoda: Brachyura). Informe final de la pasantía curricular para la obtención del titulo de grado en ciencias biológicas. São José do Rio Preto, SP, Universidade Estadual Paulita "Júlio de Mesquita Filho") estudió las uniones morfológicas del esqueleto axial con el caparazón, encontrando un sistema de aseguramiento similar entre las especies de Trichodactylidae y algunas especies representantes de Portunoidea.
En el presente trabajo son descritos aspectos de la morfología interna del esqueleto axial de Trichodactylidae y se evalúa la hipótesis de clasificación de subfamilias y tribus propuesta por (Magalhães & Türkay 1996Magalhães, C. & Türkay M. 1996. Taxonomy of the Neotropical freshwater crab family Trichodactylidae I. The generic system with description of some new genera (Crustacea: Decapoda: Brachyura). Senckenbergiana biologica, 75:63-95.) en base a los elementos anatómicos de esta estructura.
Elementos de morfología
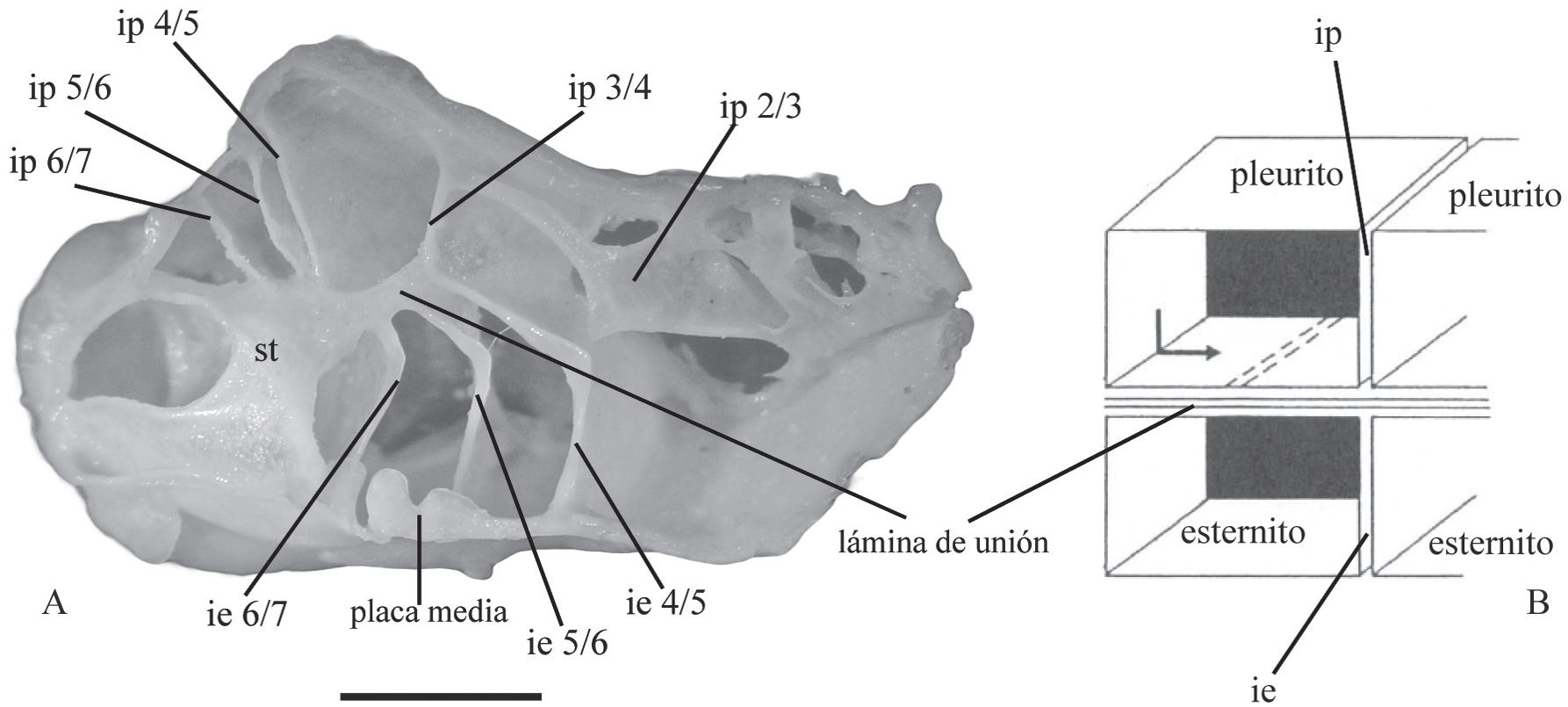
En Brachyura la armazón del esqueleto axial se conforma a partir de los escleritos ventrales (esternitos), laterales (pleuritos), y de las placas resultado de la invaginación de estos escleritos en el límite de dos somitos torácicos adyacentes (Fig. 1A, B) (Guinot, 1979Guinot, D. 1979. Données nouvelles sur la morphologie, la phylogenèse et la taxonomie des Crustacés Décapodes Brachyoures. Mémories du Muséum National d'Historie Naturelle, série A, Zoologie, 112: 1-354.; Secretan, 1998Secretan, S. 1998. The sella turcica of crabs and the endophragmal system of decapods. Journal of Natural History, 32:1753-1767.). Esas placas reciben el nombre de fragmas, y son nombradas con una doble denominación de acuerdo a los somitos que los originan. Cuando el fragma es formado por dos esternitos, es denominado interesternito, y cuando se forma entre dos pleuritos es denominado interpleurito (Fig. 1A, B) (Tavares & Secretan, 1993Tavares, M. & Secretan, S. 1993. La notion de thelycum et de spermathèque chez les Crustaces Décapodes. Comptes Rendus de l'Académie des sciences, 316(2):133-138., Secretan-Rey, 2002Secretan-Rey, S. 2002. Monographie du squelette axial de Nephrops norvegicus (Linné, 1758). Zoosystema, 24(1):81-176.).

A-B. Corte sagital del esqueleto axial de Dilocarcinus pagei Stimpson, 1861, macho adulto, en vista mesial. A, fotografía del esqueleto axial. B, representación esquemática de la constitución de la lámina de unión (modificado de Secretan, 1998). Abreviaciones: ie, interesternito; ip, interpleurito; st, sella turcica reforzada. Escala, 5 mm.
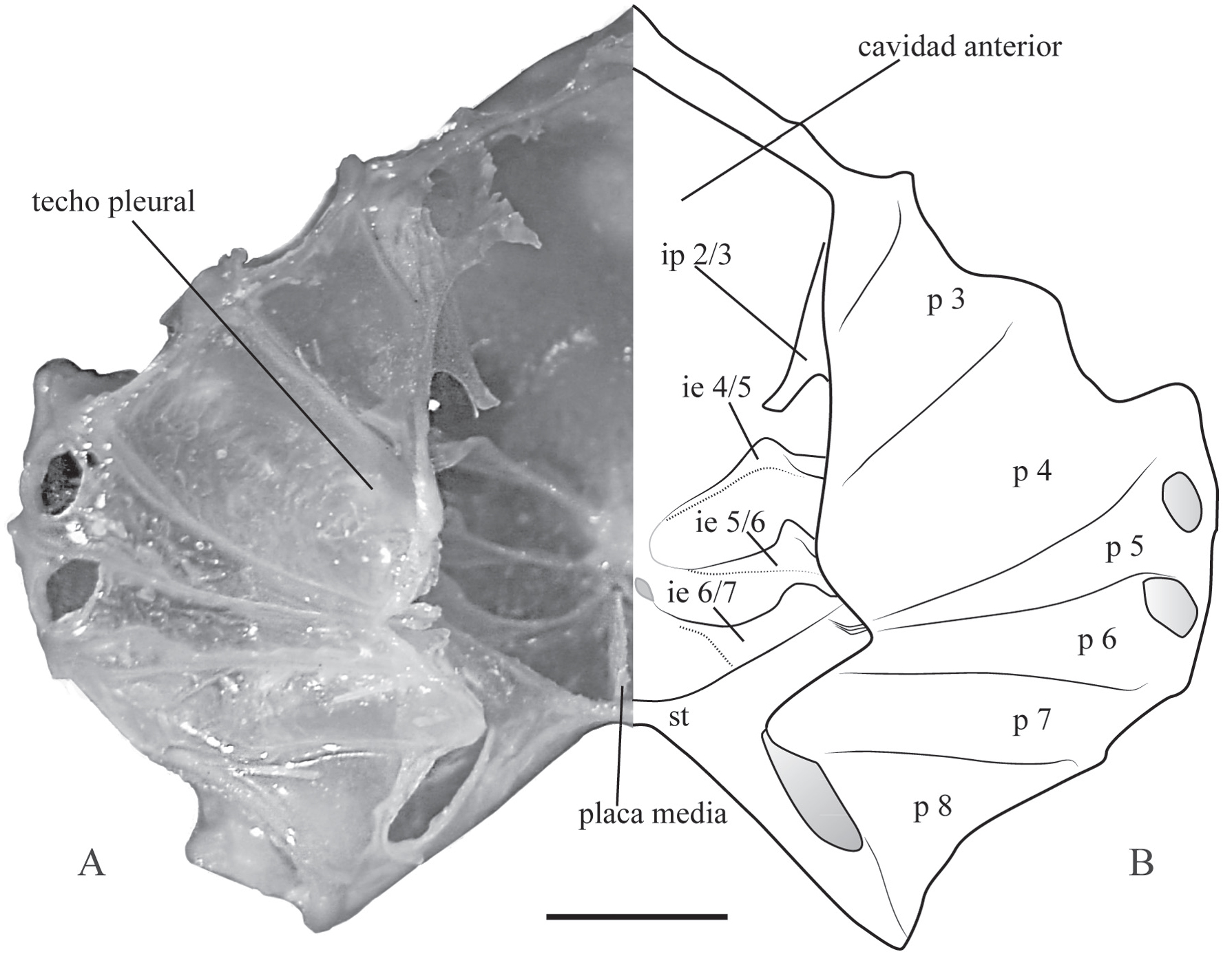
Los interesternitos pueden atravesar completamente la extensión transversal del esqueleto o estar interrumpidos en el medio y como consecuencia el fragma queda divido en dos placas laterales (Fig. 2A, B) (Guinot, 1979Guinot, D. 1979. Données nouvelles sur la morphologie, la phylogenèse et la taxonomie des Crustacés Décapodes Brachyoures. Mémories du Muséum National d'Historie Naturelle, série A, Zoologie, 112: 1-354.; Guinot et al., 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.).

A-B. Esqueleto axial de Trichodactylus fluviatilis Latreille, 1828, macho adulto, en vista dorsal (caparazón removido). A, fotografía de la mitad izquierda. B, representación esquemática de "A". Abreviaciones: ie, interesternito; ip, interpelurito; p, pleurito; st, sella turcica reforzada. Escala, 5 mm.
En la parte ventral los esternitos en conjunto forman el esternón torácico; mientras que en la región dorsal los pleuritos forman una estructura denominada "techo pleural" (Fig. 2A, B). La cohesión de los elementos del esqueleto axial en una sola armazón rígida es posible gracias a la existencia de dos estructuras que surgen como novedades evolutivas en el esqueleto axial de Eubrachyura. La primera, denominada lámina de unión, es una placa orientada horizontalmente, paralela al esternón torácico, formada partir de la fusión de los interesternitos e interpleuritos en cada extremo del esqueleto (Fig. 1A, B); esta lámina permite dividir el esqueleto en dos cámaras sobrepuestas: la cámara ventral compuesta por los esternitos, interesternitos y placa media; y la cámara dorsal formada por los pleuritos e interpleuritos (Guinot, 1979Guinot, D. 1979. Données nouvelles sur la morphologie, la phylogenèse et la taxonomie des Crustacés Décapodes Brachyoures. Mémories du Muséum National d'Historie Naturelle, série A, Zoologie, 112: 1-354.; Vidal-Gadea et al., 2008Vidal-Gadea, A.G.; Rinehart, M.D. & Belanger, J.H. 2008. Skeletal adaptations for forwards and sideways walking in three species of decapods crustaceans. Arthropod Structure & Development, 37:95-108.). La segunda estructura denominada sella turcica reforzada, es considerada como un fragma intertagmal de transición entre el tórax y el abdomen (Figs. 1, 2), que tiene origen a partir de la invaginación del octavo segmento torácico y el primero abdominal (Secretan-Rey, 2002Secretan-Rey, S. 2002. Monographie du squelette axial de Nephrops norvegicus (Linné, 1758). Zoosystema, 24(1):81-176.). Aunque la presencia de un fragma intertagmal es común a los crustáceos decápodos, la fusión de esta estructura con el interesternito 7/8 es una condición sinapomórfica de Brachyura, y la fusión adicional con el interpleurito 7/8 ocurre exclusivamente en Eubrachyura, formando la denominada sella turcica reforzada (Guinot et al., 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.). A partir de la región mediana de la sella turcica se extienden antero-lateralmente dos láminas denominados "alas" de la sella turcica que se continúan con la lámina de unión. De acuerdo a (Secretan 1998Secretan, S. 1998. The sella turcica of crabs and the endophragmal system of decapods. Journal of Natural History, 32:1753-1767., Secretan 2002Secretan, S. 2002. La lame de jonction, base de l'organisation phragmale des Décapodes Brachyoures. Crustaceana, 75(3-4):637-641.), la sella turcica forma una fuerte conexión transversal entre las dos mitades del esqueleto preservando así su unidad estructural (Fig. 2).
En la cámara ventral del esqueleto, además de la presencia de los interesternitos, existe una lámina denominada placa media, orientada perpendicular a la sella turcica, pero continua con esta, que se extiende longitudinalmente en el plano sagital del organismo (Figs. 1, 2). Esta placa es formada por la invaginación mediana del esternón torácico y externamente es representada por la línea media (Guinot, 1979Guinot, D. 1979. Données nouvelles sur la morphologie, la phylogenèse et la taxonomie des Crustacés Décapodes Brachyoures. Mémories du Muséum National d'Historie Naturelle, série A, Zoologie, 112: 1-354.; Guinot et al., 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.).
MATERIALES Y MÉTODOS
Material
Para el estudio morfológico fueron examinados 43 especímenes adultos, pertenecientes a 17 especies y 10 géneros representativos de las tribus y subfamilias de Trichodactylidae. De cada especie fue examinado por lo menos un macho y una hembra adulto. El material fue obtenido en donación de las siguientes instituciones: Instituto de Biociências da Universidade de São Paulo (IBUSP); Instituto de Ciencias Naturales de la Universidad Nacional de Colombia (ICN); Instituto Nacional de Limnología (INALI); Universidade Federal de Rio Grande do Sul (UFRGS), Museu de Zoologia da Universidade de São Paulo (MZUSP); Universidad Nacional Autónoma de México (UNAM) y el Instituto Nacional de Pesquisas da Amazônia (INPA).
Los géneros Fredilocarcinus, Melocarcinus, Poppiana, Rodriguezia y Rotundovaldivia no se incluyeron en el estudio, por no contar en las colecciones con material disponible para ser diseccionado. Las preparaciones resultantes de las disecciones fueron depositadas en la colección de crustáceos del INPA, con el numero de registro (INPA-1837).
Métodos
Las preparaciones del esqueleto axial fueron realizadas de acuerdo al siguiente protocolo modificado de (Mocquard 1883Mocquard, F. 1883. Recherches anatomiques sur l'estomac des crustacès podophthalmaires. Annales des Sciences Naturelles Zoologie et Paleontologie, 16:1-311.): (i) registro de las medidas de ancho y largo del caparazón; (ii) extracción de los pereiópodos 1 al 5 mediante corte a la altura de la coxa; (iii) sumersión en una solución de hidróxido de Potasio (KOH) al 30% a aproximadamente 90°C, por un periodo de 24 a 48 horas dependiendo del tamaño del espécimen; (iv) descarte del caparazón y abdomen; (v) lavado y limpieza manual del esqueleto. Las armazones esqueléticas resultantes de este procedimiento fueron preservados en etanol al 70%.
El análisis de las estructuras fue realizado con auxilio de un estereoscopio marca Wild M-8. Las ilustraciones definitivas fueron realizadas con el programa Adobe Illustrator CS5. Las fotografías fueron adquiridas con la cámara Panasonic Lumix modelo DMC-FZ18.
Las estructuras fueron analizadas en vistas dorsal, mesial y sagital, siguiendo la terminología de Guinot et al. (2013)Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.. El corte sagital fue realizado siguiendo el eje medio antero-posterior del cuerpo, teniendo la precaución de preservar la placa media.
RESULTADOS
Caracterización del esqueleto axial de Trichodactylidae
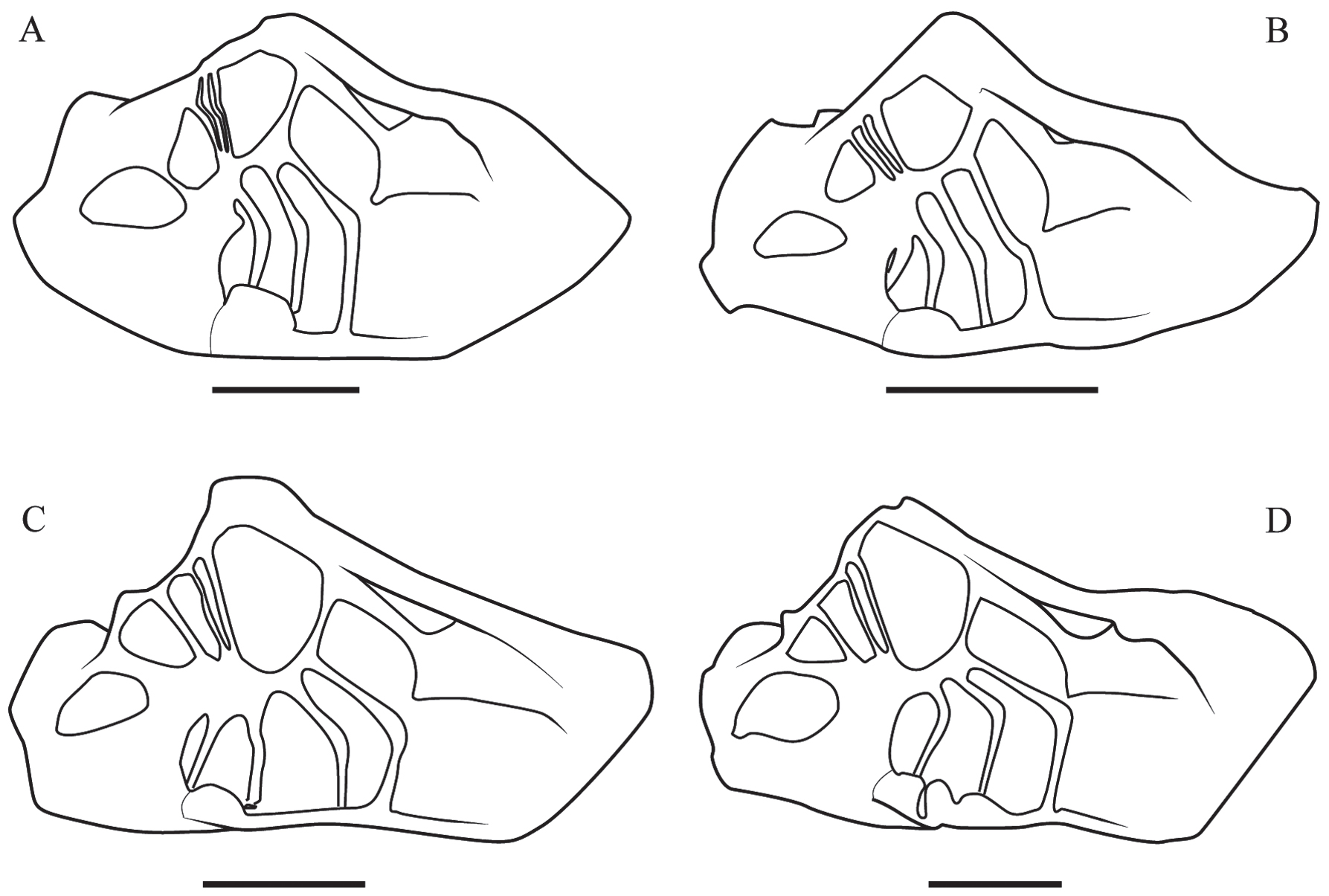
En vista dorsal la característica más sobresaliente del esqueleto axial de Trichodactylidae es la forma de la sella turcica. Esta estructura es fuertemente reducida en el medio (Figs. 2, 3), extendiéndose lateralmente a través de las "alas de la sella turcica" hasta unirse, junto con el interpelurito e interesternito 7/8 en la lámina de unión (Figs. 1, 4). La placa media tiene longitud variable en las especies de la familia, en Dilocarcinini se prolonga hasta el límite anterior del somito 5 (Fig. 4D) mientras que en Trichodactilinae y Valdiviini llega hasta el límite anterior del somito 7 (Fig. 4A-C). En las especies de Dilocarcinini es posible observar interrupciones en la extensión de la placa en forma de muescas profundas, externamente esas muescas son reflejadas en la línea media. Al igual que la sella turcica, la placa media es notablemente corta en su extensión vertical.

A-D. Representación esquemática del esqueleto axial en vista dorsal (caparazón removido). A, Trichodactylus fluviatilis Latreille, 1828, macho adulto (Trichodactylinae). B, Avotrichodactylus constrictus (Pearse, 1911), macho adulto (Trichodactylinae). C, Valdivia serrata White, 1847, macho adulto (Valdiviini). D, Bottiella medemi (Smalley & Rodríguez, 1972) macho adulto (Dilocarcinini). Note la conformación del techo pleural, la forma de la sella turcica y el grado de interrupción de los interesternitos. Escalas, 5 mm.

A-D. Representación esquemática del esqueleto axial en vista sagital. A, Trichodactylus fluviatilis Latreille, 1828, macho adulto (Trichodactylinae). B, Avotrichodactylus constrictus (Pearse, 1911), macho adulto (Trichodactylinae). C, Valdivia serrata White, 1847, macho adulto (Valdiviini). D, Dilocarcinus pagei Stimpson, 1861, macho adulto, (Dilocarcinini). Note los lugares de convergencia de los interesternitos e interpeluritos en la lámina de unión, la prolongación y continuidade de la placa mediana, y la extención vertical de los intesternitos. Escalas, 5 mm.
La lámina de unión está orientada de forma horizontal extendiéndose paralela al esternón torácico, en el extremo anterior está delimitada por la confluencia del interesternito 4/5 con el interpleurito 3/4, en la parte posterior es continua con el interpleurito 7/8 (Fig. 1A, B). En vista mesial fueron observadas diferencias evidentes respecto al lugar de unión de los interpleuritos e interesternitos en la lámina de unión. En las especies de Trichodactylinae los interpleuritos 4/5, 5/6 y 6/7 convergen en el mismo punto en la lámina de unión, siendo que este punto esta alineado o es anterior al lugar de unión del interesternito 6/7 (Fig. 4A-B). En Dilocarcininae solo los interpleuritos 4/5 y 5/6 convergen en el mismo punto en la lámina de unión mientras que los interpleuritos 6/7 y 7/8 se unen en posiciones más posteriores, el punto de convergencia del interesternito 6/7 en la lámina de unión es evidentemente mas posterior que la unión de los interpleuritos 4/5 y 5/6 (Fig. 4C-D).
El interpleurito 2/3 corresponde a una lámina de forma subtriangular orientada mesio-ventralmente, y se destaca por ser la única estructura del esqueleto que no está fusionada directamente con la lámina de unión (Fig. 1A, B).
En el techo pleural, todos los pleuritos son de forma sub-triangular, siendo mas amplios hacia la parte lateral del esqueleto. Mientras que los pleuritos 1 y 2 son reducidos y están fusionados anteriormente, el pleurito 4 se destaca por ser la lámina de mayor tamaño en el techo pleural (Fig. 2).
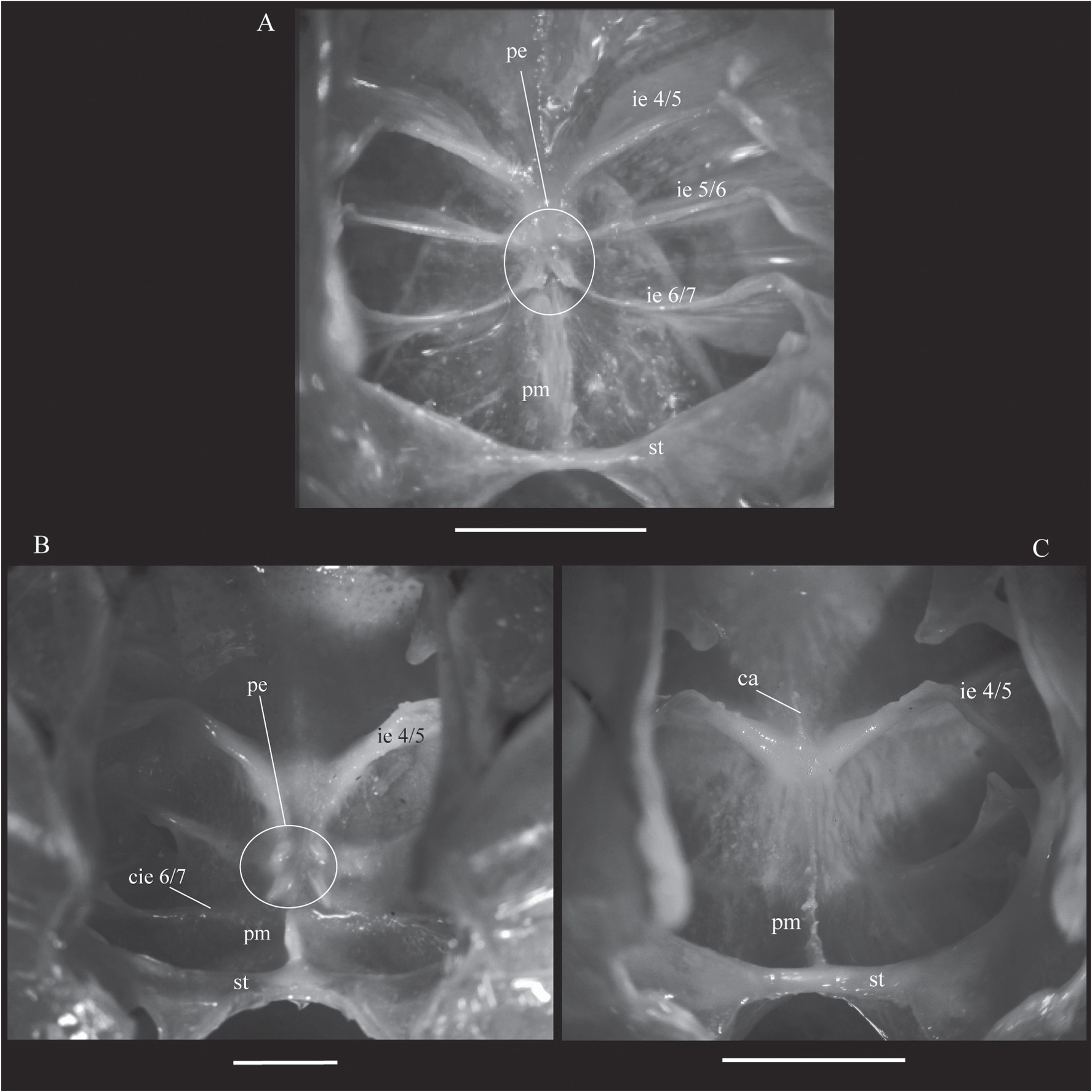
En relación a los elementos de la cámara ventral, todos los interesternitos son interrumpidos mesialmente, aunque el grado de interrupción pude variar de acuerdo a cada subfamilia. El interesternito 4/5 tiene un grado de interrupción constante en todas las especies de la familia examinadas, este fragma se puede considerar como poco interrumpido mesialmente porque se prolonga después de la cobertura de la lámina de unión hasta casi el centro del esternón torácico (Fig. 3). Los interesternitos 5/6 y 6/7 se extienden hasta el centro sobrepasando la cobertura de la lámina de unión en Trichodactylinae (Fig. 3A, B), mientras que en Dilocarcininae están replegados lateralmente bajo cobertura de esta lámina (Fig. 3C, D). No obstante, en algunas especies (e.g., Valdivia serrata) a pesar de poder calificar a los internesternitos como interrumpidos por estar replegados bajo la lámina de unión, es posible observar una cresta baja a manera de vestigio del fragma que se prolonga hasta cerca del centro del esternón (Figs. 3C, 5B). Así mismo, en algunos casos particulares (e.g.,Trichodactylus fluviatilis), es posible observar unas leves elevaciones internas localizada en el centro del esternón del torácico. Esas elevaciones son denominadas aquí de "prominencias externales", y se observaron en el extremo mesial de la prolongación del interesteresternito 6/7 o de la extensión de su cresta (Figs. 3C, 5B).

A-C. Fotografía del esternón torácico, vista desde el interior del esqueleto axial. A, Trichodactylus fluviatilis Latreille, 1828, macho adulto (Trichodactylinae). B, Zilchiopsis collastinensis (Pretzmann, 1968Pretzmann, G. 1968. Die Familie Trichodactylidae (Milne Edwards 1853) Smith 1870 (Vorläufige Mitteilung). Entomologisches Nachrichtenblatt, 15(7-8):70-76.), macho adulto, (Valdiviini). C, Dilocarcinus septemdentatus (Herbst, 1783), macho adulto, (Dilocarcinini). Note el grado de interrupción de los interesternitos, la extensión de las cristas de los interesternitos, las proeminecias externales y la configuración dorsal de la sella turcica. Abreviaciones: ca, cresta anterior; cie, cresta interesternito; ie, interesternito; pe, prominencias esternales; pm, placa media; st, sella turcica reforzada. Escalas, 5 mm.
El grado de interrupción del interesternito 7/8 es constante en todas las especies de la familia. Este fragma se encuentra fuertemente interrumpido, siempre replegado lateralmente bajo la cobertura del ala de la sella turcica, a tal punto que no es posible observarlo en la vista dorsal. Aunque en la mayoría de las especies examinadas el fragma está totalmente fusionado con la lámina de unión (Fig. 6B-D), en un pequeño grupo de especies de Trichodactylinae la porción superior mesial está separada de la lámina de unión (Fig. 6A).

A-D. Fotografía del corte sagital del esqueleto axial mostrando la lámina de unión en vista antero-mesial. A, Trichodactylus fluviatilis Latreille, 1828, macho adulto (Trichodactylinae). B, Avotrichodactylus constrictus (Pearse, 1911), macho adulto (Trichodactylinae). C, Zilchiopsis collastinensis (Pretzmann, 1968Pretzmann, G. 1968. Die Familie Trichodactylidae (Milne Edwards 1853) Smith 1870 (Vorläufige Mitteilung). Entomologisches Nachrichtenblatt, 15(7-8):70-76.), macho adulto, (Valdiviini). D, Dilocarcinus Stimpson, 1861, macho adulto, (Dilocarcinini). Note los lugares de convergencia de los interesternitos e interpeluritos en la lámina de unión y la unión mesial del interesternito 7/8 a la lámina de unión. Abreviaciones: ie, interesternito; ip, interpelurito; lu, lámina de unión; st, sella turcica reforzada. Escalas, 2 mm.
Otra notable singularidad del esqueleto axial de Trichodactylidae es la amplia separación de las dos mitades laterales del esqueleto respecto al eje medio del cuerpo (Fig. 3). Esto implica una mayor prolongación horizontal de los interesternitos 4/5, 5/6 y 6/7, cuando sobrepasan la cobertura de la lámina de unión. Verticalmente esos fragmas se proyectan de manera perpendicular respecto al esternón torácico, y cambian de orientación aproximadamente en la mitad de su extensión, orientándose en dirección diagonal a la parte posterior del cuerpo hasta unirse a la lámina de unión. En el lugar donde ocurre la curvatura se forma un lóbulo más o menos amplio dependiendo de la intensidad de la inflexión (Fig. 4).
Relaciones internas de Trichodactylidae
Si bien las especies de Trichodactylidae presentan un grupo de particularidades generales en la configuración de su esqueleto axial que permite caracterizar las especies de la familia, un análisis detallado evidencia un grupo de variaciones tanto en el esternón torácico (forma de los episternitos) como en las estructuras de la cámara ventral (grado de curvatura e interrupción mesial de los interesternitos 5/6 y 6/7, unión del internesternito 7/8 con el ala de la sella turcica, longitud de la placa media) y en las estructuras de la cámara dorsal (lugar de unión de los interpleuritos 4/5, 5/6 y 6/7 en la lámina de unión, extensión de la margen el borde mesial del techo pleural) que permitió agrupar a las especies de acuerdo a las características comunes del esqueleto. Esos agrupamientos coinciden con lo propuesto en el sistema de clasificación de Magalhães & Türkay (1996).
La caracterización de los grupos de acuerdo a las particularidades del esqueleto axial es la siguiente:
Trichodactylinae (Avotrichodactylus constrictus (Pearse, 1911), Trichodactylus fluviatilis Latreille, 1828, T. dentatus H. Milne-Edwards, 1853, T. panoplus (von Martens, 1869), T. petropolitanus (Göldi, 1886), T. boreallianus Nobili, 1896):
Se caracteriza por un esternón torácico amplio, casi tan ancho como largo, los episternitos son pequeños y de forma subtriangular. Los interesternitos 4/5, 5/6 y 6/7 están poco interrumpidos y se prolongan horizontalmente hasta cerca del centro del esternón (Fig. 3A, B), levemente curvados en su extensión vertical, con lóbulos estrechos en la región de la curvatura (Fig. 4A, B). En Trichodactylus dentatus, T. panoplus, T. fluviatilis y Avotrichodactylus constrictus se observan prominencias extérnales en el extremo mesial del interesternito 6/7 pero solo en T. fluviatilis y A. constrictus se observan prominencias también en el extremo mesial del interesternito 5/6 (Figs. 3A, B, 5A). La placa media es corta y continua extendiéndose transversalmente entre los somitos 7 y 8 (Fig. 4A, B). En Trichodactylus fluviatilis, T. petropolitanus, T. dentatus la margen del interesternito 7/8 está separada en su porción distal de la lámina de unión (Fig. 6A), mientras que en el resto de las especies de la subfamilia la fusión de estas dos estructuras es total (Figs. 4B, 6B). Los interpleuritos 4/5, 5/6, 6/7 y 7/8, se disponen muy cerca uno del otro en su extremo mesial convergiendo todos en el mismo punto en la lámina de unión, siendo que este punto es alineado o anterior con el punto de unión del interesternito 6/7 (Figs. 4A, B, 6A). La margen mesial anterior del techo pleural en vista dorsal se extiende aproximadamente hasta la porción media del esternito 6 (Fig. 6A, B).
De acuerdo a nuestras observaciones existen marcadas diferencias en características como la espesura de la lámina de unión, la extensión vertical de los interesternitos y la fusión del interesternito 7/8 con la lámina de unión, que permiten subdividir la especies de esta subfamilia de la siguiente forma:
Subgrupo A: (Trichodactylus fluviatilis, T. petropolitanus, T. dentatus): especies de porte mayor (hasta 50 mm de ancho de caparazón), caracterizadas por interesternitos extensamente prologados y levemente curvados verticalmente, el interesternito 7/8 esta separado mesialmente de la lámina de unión (Figs. 3A, 4A, 5A, 6A).
Subgrupo B: (Trichodactylus panoplus, T. borellianus y Avotrichodacytlus constrictus): especies de porte menor (hasta 30 mm de ancho de caparazón), caracterizadas por el interesternito 6/7 notablemente corto verticalmente, siendo que en A. constrictus es vestigial; interesternito 7/8 fusionado mesialmente a la lámina de unión; margen anterior mesial del techo pleural y de la lámina de unión más grueso en relación al subgrupo anterior, interpleurito 2/3 parcialmente integrado al techo pleural (Figs. 3B, 4B, 6B).
Valdiviini (Sylviocarcinus devillei H. Milne Edwards, 1853, S. pictus (H. Milne Edwards, 1853), Zilchiopsis collastinensis (Pretzmann, 1968Pretzmann, G. 1968. Die Familie Trichodactylidae (Milne Edwards 1853) Smith 1870 (Vorläufige Mitteilung). Entomologisches Nachrichtenblatt, 15(7-8):70-76.), Forsteria venezuelensis (Rathbun, 1905) y Valdivia serrata White, 1847):
Al igual que en el grupo anterior el esternón torácico es casi tan ancho como largo y los episternitos son pequeños y de forma subtriangular. El interesternito 5/6 es bien interrumpido mesialmente y se repliega lateralmente bajo de la cobertura de la lámina de unión (Fig. 3C). El interesternito 6/7 también está replegado lateralmente, pero a diferencia del interesternito 5/6, en este fragma se observa la extensión de una cresta que se prolonga hasta el medio del esternón (Figs. 3C, 5B). En todas las especies existen prominencias extérnales en la cresta del interesternito 6/7, pero solo en Silviocarcinus devillei, S. pictus y Zilchiopsis collastinensis también se encuentran prominencias en la extensión del interesternito 5/6 (Figs. 3C, 5B). Los interesternitos 4/5, 5/6 y 6/7 son curvados en la extensión dorso-ventral, los lóbulos sobre esta curvatura son ligeramente mas amplios respecto a lo observado en Trichodactylinae (Fig. 4C). Los interpleuritos 4/5 y 5/6 convergen en el mismo lugar en la lámina de unión, siendo este punto alineado o anterior al lugar de unión del interesternito 6/7 (Figs. 4C, 6B). La margen mesial de los interpleuritos 4/5 y 6/7 es recta (Fig. 4C). La placa media es corta y continua, y se extiende entre los somitos 7 y 8 (Fig. 4C).
Dilocarcinini (Bottiella medemi (Smalley & Rodríguez, 1972), Goyazana castelnaui (H. Milne Edwards, 1853), Dilocarcinus pagei Stimpson, 1861, D. septemdentatus (Herbst, 1783), Moreirocarcinus emarginatus (H. Milne Edwards, 1853) y M. laevifrons (Moreira, 1901)):
El esternón torácico es más largo que ancho, los episternitos son amplios y de forma subcircular. En Bottiella medemi, Goyazana castelnaui y Dilocarcinus septemdentatus se observa una cresta débil, orientada en el eje sagital, en la porción media de los somitos 2 y 3 (Fig. 3D).
En este grupo, en general las estructuras correspondientes a la cámara dorsal están localizadas más posteriormente en relación a los otros grupos. Eso es evidente porque, en vista dorsal, la margen mesial anterior del techo pleural se prolonga hasta la porción posterior de la sella turcica y el pleurito 4 tiene una orientación mas inclinada en relación a los otros grupos (Figs. 3D, 4D).
En la cámara ventral los interesternitos 5/6 y 6/7 son interrumpidos mesialmente y están replegados bajo la cobertura de la lámina de unión (Fig. 3D). En ninguna de las especies del grupo fueron observadas prominencias esternales. Los interesternitos 4/5, 5/6 y 6/7 son marcadamente curvados sobre su extensión dorso-ventral y tiene lóbulos amplios (Fig. 4D). La placa media se prolonga hasta el límite anterior del somito 5, presentando discontinuidades en su extensión, mientras que en Bottiella medemi, Goyazana castelnaui, Moreirocarcinus emarginatus y M. laevifrons esta placa tiene una sola muesca en la posición correspondiente a la prolongación del interesternito 5/6, en Dilocarcinus pagei y D. septemdentatus la placa tiene dos muescas en las posiciones correspondientes a las prolongaciones de los interesternitos 5/6 y 6/7 (Fig. 4D). Estas discontinuidades son también reflejadas en la línea media. La apariencia de la margen mesial de los interpleurito 4/5, 5/6 y 6/7 y la forma como convergen en la lámina de unión es similar a lo descrito para Valdiviini, sin embargo a diferencia de este grupo, el lugar de unión de esos fragmas en la lámina de unión es anterior a la unión del interesternito 6/7, llegando a estar alineada con en el interesternito 7/8 (Fig. 4D).
DISCUSIÓN
El estudio detallado de las estructuras esqueléticas en los braquiuros ha permitido en los últimos años determinar patrones y establecer relaciones de homología, soportando y auxiliando aquellas observaciones basadas en caracteres más tradicionales como el caparazón o los apéndices torácicos (Guinot, 1979Guinot, D. 1979. Données nouvelles sur la morphologie, la phylogenèse et la taxonomie des Crustacés Décapodes Brachyoures. Mémories du Muséum National d'Historie Naturelle, série A, Zoologie, 112: 1-354.; Guinot et al., 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.). A partir de nuestro trabajo fue posible realizar un grupo de observaciones que permitieron caracterizar el esqueleto axial de Trichodactylidae, relacionadas tanto con estructuras del esternón torácico como del techo pleural, la lámina de unión y la sella turcica.
Respecto a las singularidades del esqueleto axial de los tricodactilideos, nuestros resultados coincidieron con las observaciones de (Sternberg & Cumberlidge 2003Sternberg, R. von. & Cumberlidge, N. 2003. Autapomorphies of the endophragmal system in trichodactilyd freshwater crabs (Crustacea: Decapoda: Eubrachyura). Journal of Morphology, 256:23-28.), en lo que respecta a considerar la fuerte reducción de la sella turcica y de la placa media como una particularidad de la familia. Sin embargo, es discutible su interpretación referida a la reducción del interesternito 7/8, aunque este carácter sirve como criterio para distinguir entre Trichodactylidae y Pseudothelphusidae (Cumberlidge & Ng, 2009Cumberlidge, N. & Ng, P. 2009. Systematics, evolution, and biogeography of freshwater crabs. In: Martin, J.; Crandall, K. & Felder, D. Decapod Crustacean Phylogenetics. Florida, CRC Press Taylor & Francis Group. p. 491-508.); el grado de interrupción y la disposición de este fragma en Trichodactylidae es similar a lo observado en especies de Portunoidea de los géneros Carcinus e Chaceon.
Las observaciones de (Sternberg & Cumberlidge 2003Sternberg, R. von. & Cumberlidge, N. 2003. Autapomorphies of the endophragmal system in trichodactilyd freshwater crabs (Crustacea: Decapoda: Eubrachyura). Journal of Morphology, 256:23-28.) también son imprecisas al afirmar que mesialmente el pleurito 7 es del mismo tamaño que el pleurito 4, ya que según nuestras observaciones el pleurito 4 es notablemente más amplio que todos los demás.
A pesar de la fuerte reducción mesial de la sella turcica descrita para la familia, es evidente la fusión del interesternito y el interpleurito 7/8 a este fragma intertagmal, permitiendo determinar el estado característico de la sella turcica "reforzada", que de acuerdo a (Guinot et al. 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.) está presente en los Eubrachyura.
Independiente de que el grado de interrupción de los interesternitos 5/6 y 6/7 varia de acuerdo a cada subfamilia de Trichodactylidae, la característica de poseer los interesternitos 4/5 a 7/8 interrumpidos mesialmente es constante para todas las especies de la familia. De acuerdo a esta característica es posible caracterizar el esqueleto axial de Trichodactylidae dentro del "patrón 5" de la clasificación propuesta por (Guinot et al. 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.), que agrupa los braquiuros que poseen los interesternitos 4/5 a 7/8 incompletos. Este patrón abarca una amplia variedad de eubraquiuros Heterotramata y a todos los Thoracotremata. A su vez los autores proponen una clasificación interna dentro del "patrón 5" determinada por la extensión de la placa media. Como fue descrito en los resultados, existen dos patrones respecto a la extensión de la placa media en las especies de la familia. En Dilocarcinini, la placa media es larga, prolongada hasta el limite anterior del somito 4 e interrumpida en su extención por un par de muescas profundas, mientras que en Valdiviini y Trichodactylinae la placa media es corta, continua y se extiende hasta el limite anterior del somito 5. Siguiendo la clasificación de (Guinot et al. 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.), Dilocarcinini corresponde al sub-patrón 5c y Valdiviini y Trichodactylinae al 5e. Esta caracterización del esqueleto axial en base a los patrones de las líneas de sutura del esternón torácico, que fue inicialmente propuesta en (Guinot 1979Guinot, D. 1979. Données nouvelles sur la morphologie, la phylogenèse et la taxonomie des Crustacés Décapodes Brachyoures. Mémories du Muséum National d'Historie Naturelle, série A, Zoologie, 112: 1-354.) y complementada con nuevas informaciones en (Guinot et al. 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.), no pretende reflejar las relaciones filogenéticas de los grupos, mas si un patrón general de evolución del esqueleto axial en los braquiuros. De acuerdo a esto, es coherente que todas las especies de Trichodactylidae estén agrupadas en un mismo grupo "patrón 5" junto con otros Heterotremata. La forma particular de la placa media de Dilocarcinini parece ser una sinapomorfia del grupo, lo que refleja su clasificación en un subgrupo diferente al de las otras especies de la familia. De acuerdo a (Guinot et al. 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.), una disposición similar a la placa media de Dilocarcinini, no solo respecto a su longitud sino también a su continuidad, está presente en especies del genero Ucides de la familia Ocypodidae.
Respecto a las relaciones internas de la familia, los resultados de este trabajo fueron consistentes con la clasificación supragenerica propuesta por (Magalhães & Türkay 1996Magalhães, C. & Türkay M. 1996. Taxonomy of the Neotropical freshwater crab family Trichodactylidae I. The generic system with description of some new genera (Crustacea: Decapoda: Brachyura). Senckenbergiana biologica, 75:63-95.), los caracteres relacionados las líneas de sutura del esternón torácico, descritos por estos autores, corresponden con las observaciones realizadas por nosotros respecto al grado de interrupción de los interesternitos y extensión de la placa media.
La subfamilia Trichodactylinae, compuesta por los géneros Trichodactylus, Rodriguezia y Avotrichodactylus (Magalhães & Türkay, 1996Magalhães, C. & Türkay M. 1996. Taxonomy of the Neotropical freshwater crab family Trichodactylidae I. The generic system with description of some new genera (Crustacea: Decapoda: Brachyura). Senckenbergiana biologica, 75:63-95.), está bien delimitada respecto al esqueleto axial. Sus principales características son: la extensión de los interesternitos 5/6 y 6/7 hasta el centro del esternón; la forma redondeada del borde mesial superior de los interpleuritos 4/5, 5/6 y 6/7 y la convergencia de esos fragmas en un mismo punto en la lámina de unión.
No obstante los resultados también sugieren el agrupamiento de Trichodactylus borellianus, T. panoplus y Avotrichodactylus constrictus como un subgrupo de Trichodactylinae. (Rodríguez 1992Rodríguez, G. 1992. The freshwater crabs of America. Family Trichodactylidae and supplement to the family Pseudothelphusidae. Faune Tropicale, 31:1-189.) designo el género Mikrotrichodactylus para agrupar a T. borellinus y T. panoplus, sin embargo, (Magalhães & Türkay 1996Magalhães, C. & Türkay M. 1996. Taxonomy of the Neotropical freshwater crab family Trichodactylidae I. The generic system with description of some new genera (Crustacea: Decapoda: Brachyura). Senckenbergiana biologica, 75:63-95.) no reconocieron este género. De las características que agrupan estas dos especies la más significativa es la unión mesial del interesternito 7/8 a la lámina de unión, junto a este carácter también se observo la cobertura parcial del surco peniano por el episternito 7 descrito por (Rodríguez 1992Rodríguez, G. 1992. The freshwater crabs of America. Family Trichodactylidae and supplement to the family Pseudothelphusidae. Faune Tropicale, 31:1-189.). Estas características podrían significar la revalidación de Mikrotrichodactylus, así como la eventual subdivisión a nivel de la tribu de Trichodactylinae. Esta hipótesis solo podrá ser validada después de la realización de estudios morfológicos más amplios que involucren todas las especies del grupo.
Respecto a Dilocarcininae, sus dos tribus también fueron claramente delimitadas por características del esqueleto axial, sin embargo, a diferencia de Dilocarcinini que tiene características propias y bien definidas (como la extensión de la placa media), la mayoría de los caracteres que definen a Valdiviini son compartidos por especies de alguno de los otros dos grupos. Considerando que existe un grupo de caracteres externos, como la fusión de los somitos abdominales, la presencia de cicatrices pos-gástricas (postgastric pits) y la morfología del primer gonopodo, que permiten una clara delimitación de Dilocarcininae, este taxón se puede definir por las siguientes características del esqueleto axial: (a) la marcada interrupción mesial de los interesternitos 5/6 y 6/7; (b) el borde superior mesial recto de los interesternitos 4/5 y 6/7; (c) el interpleurito 6/7 está unido posteriormente a la convergencia de los interpleuritos 4/5 y 5/6 a la lámina de unión. Al contrario de lo mencionado por (Magalhães & Türkay 1996Magalhães, C. & Türkay M. 1996. Taxonomy of the Neotropical freshwater crab family Trichodactylidae I. The generic system with description of some new genera (Crustacea: Decapoda: Brachyura). Senckenbergiana biologica, 75:63-95.) referente a que Dilocarcininae tiene el esternón torácico más estrecho que Trichodactylinae, en las especies analizadas en este trabajo, Trichodactylinae y Valdiviini fueron más cercanos entre sí respecto a este carácter, pues en ambos grupos el esternón torácico es amplio con episternitos subtriangulares, mientras que en Dilocarcinini el esternón es más estrecho y con episternitos semicirculares mas amplios.
De acuerdo a (Guinot 1979Guinot, D. 1979. Données nouvelles sur la morphologie, la phylogenèse et la taxonomie des Crustacés Décapodes Brachyoures. Mémories du Muséum National d'Historie Naturelle, série A, Zoologie, 112: 1-354.) y (Guinot et al. 2013Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.), la condición primitiva en el esqueleto axial generalmente está determinada por el grado de interrupción mesial de los interesternitos. Según este criterio, el esqueleto axial de Trichodactylinae presentaría la condición más plesiomórfica dentro de la familia, dado que es el grupo con los interesternitos menos interrumpidos. Además de la interrupción de los interesternitos, la separación de la porción distal de la margen mesial del interesternito 7/8 con el ala de la sella turcica observada en algunas especies de esta subfamilia (subgrupo A), es una condición que comparte con especies de las otras familias de Eubrachyura (Pseudothelphusidae, Geryonidae y Carcinidae). Con estos criterios se puede concluir que Trichodactylinae es el grupo con características más primitivas respecto a los otros dos; mientras que Dilocarcinini, dada las particularidades de su esqueleto, especialmente la longitud de la placa media y la amplitud del esternón torácico, sería el grupo más derivado. Estas observaciones son consistentes con los cladogramas presentados por (Rodríguez 1992Rodríguez, G. 1992. The freshwater crabs of America. Family Trichodactylidae and supplement to the family Pseudothelphusidae. Faune Tropicale, 31:1-189.) y (Sternberg 1997Sternberg, R. von. 1997. Cladistics of the freshwater crab family Trichodactylidae (Crustacea: Decapoda): appraising the reappraisal. Journal of Comparative Biology, 2(1):49-62.).
A partir de los resultados de nuestro estudio es posible concluir que aunque el esqueleto axial es una estructura compleja y con un patrón de organización bien establecido en los eubraquiuros, puede presentar particularidades a nivel de familia, subfamilia e incluso tribu. Este estudio destaca la importancia de la morfología de las estructuras internas del esqueleto axial como herramienta taxonómica en los diferentes grupos de Brachyura.
AGRADECIMIENTOS
Los ejemplares fueron obtenidos gracias a las donaciones de las siguientes investigadores: Sérgio Luiz de Siqueira Bueno, del Instituto de Biociências da Universidade de São Paulo (IBUSP), Martha Helena Rocha de Campos, del Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá, Colombia (ICN), Paulo Collins, Instituto del Nacional de Limnología, Santo Tomé, Argentina (INALI), Georgina Bond-Buckup, de la Universidade Federal de Rio Grande do Sul (UFRGS), Porto Alegre, Brasil y Fernando Álvarez, de la Universidad Nacional Autónoma de México, Ciudad de México (UNAM). Los autores agradecen al CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnólogico) por la beca de estudio otorgara mediante el programa PEC-PG 2008 (MP) y por la beca de investigación (CM, proc. nº 303837/2012-6; MT, proc nº 301806/2010-1) y a la FAPESP por la beca de doctorado proc. nº 2012/01334-7 (MP).
- Bitsch, C. & Bitsch, J. 2002. The endoskeletal structures in arthropods: citology, morphology and evolution. Arthropod Structure & Development, 30:159-177.
- Bott, R. 1969. Die Süsswasserkrabben Süd-Amerikas und ihre Stammesgeschichte. Eine Revision der Trichodactylidae und der Pseudothelphusidae östlich der Anden (Crustacea, Decapoda). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 518:1-94.
- Cumberlidge, N. & Ng, P. 2009. Systematics, evolution, and biogeography of freshwater crabs. In: Martin, J.; Crandall, K. & Felder, D. Decapod Crustacean Phylogenetics. Florida, CRC Press Taylor & Francis Group. p. 491-508.
- Drach, P. 1950. Les étapes évolutives de l'endosquelette chez lês Crustacés Décapodes. Comptes rendus hebdomadaires des séances de l'Académie des Sciences, 231(25):1563-1565.
- Guinot, D. 1979. Données nouvelles sur la morphologie, la phylogenèse et la taxonomie des Crustacés Décapodes Brachyoures. Mémories du Muséum National d'Historie Naturelle, série A, Zoologie, 112: 1-354.
- Guinot, D.; Tavares, M. & Castro, P. 2013. Significance of the sexual openings and supplementary structures on the phylogeny of brachyuran crabs (Crustacea, Decapoda, Brachyura), with new nomina for higher-ranked podotreme taxa. Zootaxa, 3665(1):1-414.
- Juares, A. 2008. Conexões morfológicas entre carapaça e esqueleto endofragmal nos caranguejos de água doce Trichodactylidae Milne-Edwards, 1853 (Crustacea: Decapoda: Brachyura). Informe final de la pasantía curricular para la obtención del titulo de grado en ciencias biológicas. São José do Rio Preto, SP, Universidade Estadual Paulita "Júlio de Mesquita Filho"
- Magalhães, C. 2003. Famílias Pseudothelpusidae e Trichodactylidae. In: Melo, G.A.S. Manual de identificação dos Crustácea Decápode de água doce do Brasil. São Paulo, Editora Loyola. p. 143-287.
- Magalhães, C. & Türkay M. 1996. Taxonomy of the Neotropical freshwater crab family Trichodactylidae I. The generic system with description of some new genera (Crustacea: Decapoda: Brachyura). Senckenbergiana biologica, 75:63-95.
- Mocquard, F. 1883. Recherches anatomiques sur l'estomac des crustacès podophthalmaires. Annales des Sciences Naturelles Zoologie et Paleontologie, 16:1-311.
- Ng, P.K.L.; Guinot, D.& Davie, P.J.F. 2008. Systema Brachyurorum: Part 1. An annotated checklist of extant brachyuran crabs of the world. The Raffles Bulletin of Zoology, 17:1-286.
- Pretzmann, G. 1968. Die Familie Trichodactylidae (Milne Edwards 1853) Smith 1870 (Vorläufige Mitteilung). Entomologisches Nachrichtenblatt, 15(7-8):70-76.
- Rathbun, M.J. 1906. Le crabs d'eau douce (Potamidae). Nouvelles Archives du Museum d'Histoire Naturelle de Paris, 8(4):33-122.
- Rodríguez, G. 1992. The freshwater crabs of America. Family Trichodactylidae and supplement to the family Pseudothelphusidae. Faune Tropicale, 31:1-189.
- Secretan, S. 1980. Le plan de base du "squelette axial" d'un Crustacé décapode macroure et sa terminologie. Comptes rendus hebdomadaires des seánces de l'Académie des Sciences, série D, 291:877-880.
- Secretan, S. 1998. The sella turcica of crabs and the endophragmal system of decapods. Journal of Natural History, 32:1753-1767.
- Secretan, S. 2002. La lame de jonction, base de l'organisation phragmale des Décapodes Brachyoures. Crustaceana, 75(3-4):637-641.
- Secretan-Rey, S. 2002. Monographie du squelette axial de Nephrops norvegicus (Linné, 1758). Zoosystema, 24(1):81-176.
- Sternberg, R. von. 1997. Cladistics of the freshwater crab family Trichodactylidae (Crustacea: Decapoda): appraising the reappraisal. Journal of Comparative Biology, 2(1):49-62.
- Sternberg, R. von. & Cumberlidge, N. 2003. Autapomorphies of the endophragmal system in trichodactilyd freshwater crabs (Crustacea: Decapoda: Eubrachyura). Journal of Morphology, 256:23-28.
- Tavares, M. & Secretan, S. 1993. La notion de thelycum et de spermathèque chez les Crustaces Décapodes. Comptes Rendus de l'Académie des sciences, 316(2):133-138.
- Vidal-Gadea, A.G.; Rinehart, M.D. & Belanger, J.H. 2008. Skeletal adaptations for forwards and sideways walking in three species of decapods crustaceans. Arthropod Structure & Development, 37:95-108.
Fechas de Publicación
-
Publicación en esta colección
2015
Histórico
-
Recibido
04 Feb 2015 -
Acepto
31 Mar 2015