Resumos
O conhecimento da profundidade ideal de germinação de sementes de plantas daninhas é importante para o desenvolvimento de estratégias de manejo eficientes, seguras e econômicas. Com o objetivo de estudar a emergência de plântulas de Sida rhombifolia L. e Solanum viarum Dunal, em resposta à época (setembro de 2008 e janeiro de 2009) e às profundidades de semeadura (0, 1, 2, 3, 4 e 5 cm), foram realizados dois experimentos em casa de vegetação. Sida rhombifolia mostrou-se sensível às variações de temperatura, em decorrência das épocas de semeadura, e os maiores percentuais de emergência ocorreram nas profundidades entre 1 e 4 cm. Para S. viarum, observou-se forte influência da temperatura sobre a sua emergência, sendo, observado o máximo de emergência, nas profundidades de 1 a 5 cm e sua redução para as sementes locadas na superfície do solo.
Joá-bravo; guanxuma; germinação; biologia reprodutiva
Knowing the ideal depth for germination of weed seed is important for the development of efficient, safe and economical management strategies. In order to study the effects of the seasonality on the seedling emergence of Sida rhombifolia L. and Solanum viarum Dunal, two experiments were conducted in greenhouse conditions during September 2008 and January 2009. Seeds of both species were sown at different depths (0, 1, 2, 3, 4 and 5 cm). Sida rhombifolia was less sensitive to seasonal variations, and the highest percentages of emergence occurred between 1 and 4 cm depths. It was observed strong seasonality influence on the emergence of S. viarum. For this species highest emergency was observed in the greater depths, and the lowest for seeds located on the soil surface.
Tropical soda apple; arrowleaf sida; germination; reproductive biology
FITOSSANIDADE PLANT PROTECTION
Efeito da época sobre a emergência de Sida rhombifolia e Solanum viarum em diferentes profundidades de semeadura
Effects of seasonal variations on the emergence of Solanum viarum and Sida rhombifolia under different sowing depths
Marcelo Claro SouzaI; Mariana Casari ParreiraII; Carita Liberato do AmaralIII; Pedro Luís da Costa Aguiar AlvesIV
IBiólogo, Mestre. Doutorando em Ciências Biológicas no Instituto de Biociências de Rio Claro, Universidade Estadual Paulista Júlio de Mesquita Filho, Av. 24A, 1515, Bela Vista, 13506-900, Rio Claro, São Paulo, Brasil. marcelo_claro@yahoo.com.br
IIEngenheira-Agrônoma, Mestre. Doutoranda do Departamento de Biologia Aplicada à Agropecuária, Universidade Estadual Paulista Júlio de Mesquita Filho, Rod. de Acesso Prof. Paulo Donato Castellane, km 5, 14884-900, Jaboticabal, São Paulo, Brasil. mcparreira@yahoo.com.br
IIIEngenheira-Agrônoma. Mestranda do Departamento de Biologia Aplicada à Agropecuária, Universidade Estadual Paulista Júlio de Mesquita Filho, Rod. de Acesso Prof. Paulo Donato Castellane, km 5, 14884-900, Jaboticabal, São Paulo, Brasil. caritaliberato@gmail.com
IVEngenheiro-Agrônomo, Doutor. Departamento de Biologia Aplicada à Agropecuária, Universidade Estadual Paulista Júlio de Mesquita Filho, Rod. de Acesso Prof. Paulo Donato Castellane, km 5, 14884-900, Jaboticabal, São Paulo, Brasil. plalves@fcav.unesp.br
RESUMO
O conhecimento da profundidade ideal de germinação de sementes de plantas daninhas é importante para o desenvolvimento de estratégias de manejo eficientes, seguras e econômicas. Com o objetivo de estudar a emergência de plântulas de Sida rhombifolia L. e Solanum viarum Dunal, em resposta à época (setembro de 2008 e janeiro de 2009) e às profundidades de semeadura (0, 1, 2, 3, 4 e 5 cm), foram realizados dois experimentos em casa de vegetação. Sida rhombifolia mostrou-se sensível às variações de temperatura, em decorrência das épocas de semeadura, e os maiores percentuais de emergência ocorreram nas profundidades entre 1 e 4 cm. Para S. viarum, observou-se forte influência da temperatura sobre a sua emergência, sendo, observado o máximo de emergência, nas profundidades de 1 a 5 cm e sua redução para as sementes locadas na superfície do solo.
Palavras-chave: Joá-bravo, guanxuma, germinação, biologia reprodutiva.
ABSTRACT
Knowing the ideal depth for germination of weed seed is important for the development of efficient, safe and economical management strategies. In order to study the effects of the seasonality on the seedling emergence of Sida rhombifolia L. and Solanum viarum Dunal, two experiments were conducted in greenhouse conditions during September 2008 and January 2009. Seeds of both species were sown at different depths (0, 1, 2, 3, 4 and 5 cm). Sida rhombifolia was less sensitive to seasonal variations, and the highest percentages of emergence occurred between 1 and 4 cm depths. It was observed strong seasonality influence on the emergence of S. viarum. For this species highest emergency was observed in the greater depths, and the lowest for seeds located on the soil surface.
Key words: Tropical soda apple, arrowleaf sida, germination, reproductive biology.
INTRODUÇÃO
Uma das maiores limitações para a implementação de um programa de manejo integrado de plantas daninhas é a falta de conhecimentos sobre a biologia e ecologia das espécies (Fernández, 1982). Segundo Brighenti et al. (2003), o conhecimento da profundidade da qual a plântula é capaz de emergir pode permitir a adoção de práticas de manejo pertinentes. A profundidade do solo em que uma semente é capaz de germinar e produzir plântula é variável entre as espécies e apresenta importância ecológica e agronômica (Guimarães et al., 2002).
Nos ambientes agrícolas constantemente perturbados, a formação da comunidade de plantas daninhas depende do estabelecimento periódico das plântulas, com base na germinação das sementes presentes no solo. A composição específica e a densidade dependem, dentre outros fatores, do tamanho do banco de sementes, das condições climáticas e da distribuição dessas sementes no perfil do solo (Carmona, 1992; Dyer, 1995).
O banco de sementes e propágulos vegetativos no solo constitui a principal fonte de regeneração de plantas daninhas em áreas agrícolas, sendo a dormência um dos principais mecanismos de preservação de espécies em bancos de sementes, distribuindo a germinação ao longo do tempo, garantindo a sobrevivência das espécies, como sementes, sob condições adversas (Deuber, 1992), sendo a dinâmica, em um banco de sementes, influenciada diretamente pela sucessão de entrada e saída de sementes ao longo do tempo (Simpson et al., 1989).
Espécies silvestres geralmente apresentam mecanismos de dormência, enquanto as cultivadas foram perdendo estes mecanismos, por processo de seleção durante a domesticação, resultando nos cultivares modernos, com pouca ou nenhuma dormência (Carmona, 1992). Dentre as espécies silvestres, encontram-se Sida rhombifolia (guanxuma) e Solanum viarum (joá-bravo).
O joá-bravo é nativo do Brasil e da Argentina, encontrado como planta exótica na América Central, Caribe, África, Índia e Nepal, onde infesta pastagens, culturas, pomares e terrenos baldios. Na América do Norte, especialmente no Estado da Flórida (onde foi involuntariamente introduzido), encontrou condições climáticas e edáficas adequadas (Coile, 1993) e bons agentes de disseminação de suas sementes, tornando-se uma das principais plantas daninhas de pastagens, naquele estado (Medal et al., 1999), e disseminando-se por demais estados norte-americanos. Esta planta pode conter bacterioses que também atacam solanáceas, como tomate, berinjela e outras espécies cultivadas no Brasil.
A guanxuma é uma planta nativa do continente americano, ocorrendo intensamente na América do Sul e, em menor intensidade, no sul dos Estados Unidos. No Brasil, é a espécie mais comum na região Sul, ocorrendo, todavia, em todas as regiões, sendo infestante em diversas culturas, como pastagens e áreas desocupadas, dificultando a colheita mecânica em culturas anuais, por seu caule muito resistente, além de poder servir de hospedeira de um micoplasma, que causa a doença conhecida como "virose das malváceas" (Kissmann & Groth, 2000; Lorenzi, 2000).
Dos fatores que determinam a passagem do estádio fenológico de semente para o de plântulas, a profundidade do diásporo no perfil do solo é um dos mais importantes e, sem dúvida, sua avaliação é essencial para o desenvolvimento de modelos de crescimento populacional da espécie (Canossa et al., 2007). Dessa forma, o conhecimento desse fator subsidia estratégias de manejo eficientes, seguras e econômicas, (Guimarães et al., 2002; Brighenti et al., 2003; Radosevich et al., 2007), possibilita o desenvolvimento de estratégias de redução do potencial do banco de sementes (Souza Filho, 2006), além de auxiliar no entendimento da dinâmica de formação do banco de sementes em solos agrícolas.
Objetivou-se, com este trabalho, avaliar a capacidade de emergência de plântulas de S. viarum e S. rhombifolia em resposta à profundidade de semeadura, em duas épocas, em condições de casa de vegetação.
MATERIAL E MÉTODOS
Este trabalho constou de quatro experimentos, dois para S. viarum (joá-bravo) e dois para S. rhombifolia (guanxuma), conduzidos em casa de vegetação, em duas épocas do ano, em setembro de 2008 e em janeiro de 2009, na cidade de Jaboticabal, Estado de São Paulo, Brasil.
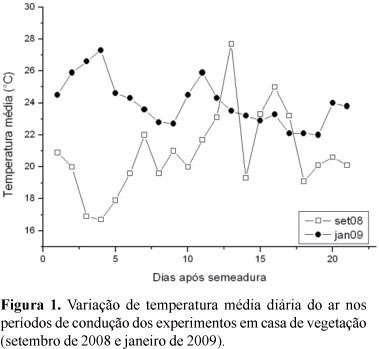
A temperatura média diária (Figura 1), no transcorrer dos experimentos, nas duas épocas de semeadura, foi de 20,8 °C, em setembro de 2008, e 24,0 °C, em janeiro de 2009. O pico máximo de temperatura (27,7 °C), para setembro, ocorreu no 13° DAS e, em janeiro (27,3 °C), no 4° DAS. Os picos mínimos, observados em setembro (16,7 °C) e janeiro (22,0 °C), ocorreram no 4° e 19° DAS, respectivamente
Como parcelas experimentais, foram utilizados vasos plásticos com capacidade de 5 L, preenchidos com terra coletada da camada arável de um Latossolo Vermelho distrófico (Embrapa, 2006), com granulometria de 380 g kg-1 de argila, 50 g kg-1 de silte e 570 g kg-1 de areia. A terra foi seca à sombra e, em seguida, passada em peneira de malha de 5 mm, antes de ser acondicionada nos vasos.
As sementes das duas espécies de plantas daninhas foram coletadas de plantas que apresentavam bom estado sanitário, nos meses de agosto e dezembro de 2008, ambas em Jaboticabal. As coletas ocorreram com antecedência máxima de 15 dias da instalação de cada experimento, visando a evitar que as sementes entrassem em processo de dormência. Em cada vaso, foram depositadas 50 sementes, nas profundidades em estudo.
O delineamento experimental adotado em todos os experimentos foi inteiramente casualizado, com seis tratamentos, em cinco repetições. Os tratamentos constaram de seis profundidades de semeadura: 0, 1, 2, 3, 4 e 5 cm. As plântulas foram consideradas emergidas quando a protrusão dos cotilédones tornou-se visível (Machado Neto & Pitelli, 1988). No caso das sementes localizadas na superfície do solo, a emergência foi considerada quando as plântulas apresentavam início de abertura dos cotilédones.
Diariamente, e até o final do período de emergência, as plântulas foram cortadas próximo ao solo e contadas. Com os dados diários de emergência, calculou-se o índice de velocidade de emergência (IVE) para cada profundidade de semeadura, segundo a fórmula proposta por Maguire (1962):
IVE = E1/D1 + E2/D2 + ... En/Dn (E1, E2, ...,
em que:
En = nº de plântulas emergidas, observadas no intervalo da 1ª, 2ª, ..., última contagem (após 20 dias);
D1, D2, ..., Dn = nº de dias da semeadura à 1ª, 2ª, ..., última contagem.
Também foi calculado o percentual de germinação para cada época avaliada, sendo estas transformadas para Arc sen ( ) (Gomes, 1984 adaptado). Os dados obtidos foram submetidos à análise de variância (teste F) e as médias foram comparadas por meio do teste de Tukey, a 5% de probabilidade.
) (Gomes, 1984 adaptado). Os dados obtidos foram submetidos à análise de variância (teste F) e as médias foram comparadas por meio do teste de Tukey, a 5% de probabilidade.
RESULTADOS E DISCUSSÃO
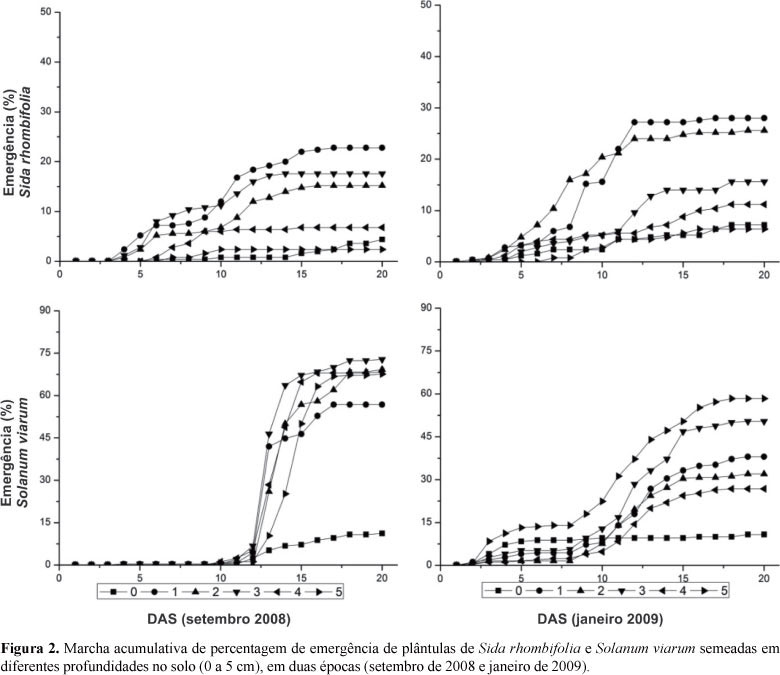
As plântulas de guanxuma emergiram de forma semelhante, nas duas épocas avaliadas, entre o 3º e 17º dias após semeadura (DAS), enquanto o joá-bravo mostrou ser mais susceptível à temperatura. Em setembro de 2008, a emergência de joá-bravo deu-se entre o 10º e 17º DAS e, em janeiro de 2009, sua emergência foi mais gradativa e por um período mais prolongado, ocorrendo a partir do 3º DAS (Figura 2). Estas variações na velocidade de germinação e emergência são particulares da biologia reprodutiva de cada espécie. Em experimento, avaliando emergência de Bidens pilosa L. em diferentes profundidades de semeadura, por exemplo, foram observadas variações na emergência das plântulas, sendo estas atribuídas a variações sazonais decorrentes dos diferentes períodos avaliados (Souza et al., 2009). Muniz Filho et al. (2004) verificaram que a velocidade de emergência de picão-preto foi reduzida significativamente com o aumento da profundidade de semeadura.
Em relação ao percentual de plântulas emergidas e às variações observadas nos IVE (Tabela 1), houve variações significativas nos dois períodos para guanxuma e joá-bravo, de modo que, para guanxuma observaram-se menores percentuais nas maiores profundidades, concordando com os resultados obtidos por Smith et al. (1992), em que as maiores percentagens de emergência ocorreram entre 0,5 cm e 2,0 cm de profundidade, não ocorrendo emergência a partir de 5,0 cm de profundidade de semeadura. Pacheco et al. (2009), trabalhando com Brachiaria brizantha (Hochst. ex A. Rich.) Stapf em diferentes profundidades de semeadura, observou sensível redução na velocidade de emergência, com o aumento da profundidade de deposição da semente. Para joá-bravo, observou-se redução de emergência na superfície, como observaram Mullahey & Cornell (1994) quando constataram emergência de 63% de plântulas a 3,6 cm de profundidade.
Ao se avaliar a biologia reprodutiva de S. viarum (Akanda et al., 1996) e de outras espécies, como Stachytarpheta cayennensis (Rich.) Vahl, Ipomoea asarifolia (Desr.) Roem. & Schult. (Dias Filho, 1996), Xanthium strumarium L. (Toledo et al., 1993), verificou-se que algumas espécies germinam somente na superfície ou nos primeiros milímetros do solo, embora outras sejam capazes de germinar a 10 ou 15 cm de profundidade. No caso específico do S. viarum, profundidades superiores a 6 cm reduzem significativamente sua emergência, sendo a profundidade de 12 cm mais prejudicial que a própria superficialidade das sementes no solo (Akanda et al., 1996). Para Pearson et al. (2003) e Alberguini & Yamashita (2010), o tamanho das sementes possui relação direta com os processos germinativos. Assim as plântulas podem apresentar dificuldade de emergir, quando enterrada em maiores profundidades. Essa capacidade de germinar, ou não, em maiores profundidades pode determinar as espécies que predominarão nos diferentes sistemas de produção, em função principalmente dos métodos de preparo das áreas para o plantio (direto ou convencional), constituindo assim importante estratégia de sobrevivência das espécies daninhas.
As plantas daninhas, em especial, dependem diretamente da germinação e emergência para infestar e competir com as espécies cultivadas (Roberts, 1999). Deste modo, a germinação das espécies invasoras pode ser afetada por condições de luz, temperatura, ação de fitormônios e umidade, que são variáveis durante o período de formação das sementes (Takahoshi, 1995).
Segundo Akanda et al. (1996), a temperatura responsável pelo maior percentual de germinação de S. viarum é de 30°C. Dessa forma, a temperatura pode ter sido um dos fatores responsáveis pelas alterações observadas na emergência de joá-bravo. De acordo com Gasparim et al. (2005), a temperatura do solo é um dos fatores mais importantes para germinação de sementes e as temperaturas nas proximidades da superfície do solo são muito semelhantes, sendo significativamente atenuadas apenas após 5 cm de profundidade. Para guanxuma, aparentemente, as variações de temperatura não influenciaram na germinação e emergência das plântulas, pois dentro do período de emergência (5º a 15º DAS) a temperatura variou entre 20°C e 25°C, estando essa margem de oscilação dentro da faixa de maior emergência, observada por Smith et al., (1992), em experimento envolvendo S. rhombifolia e S. spinosa. Segundo esses mesmos autores, a luminosidade não é fator limitante à germinação de S. rhombifolia. Dessa forma, explicam-se em parte, as semelhanças observadas para o percentual de emergência e IVE, nas profundidades de 5 cm e superficial, nas duas épocas avaliadas.
A distribuição vertical de sementes ao longo do perfil do solo apresenta estreita correlação com o tipo de preparo e afeta as condições ambientais a que as sementes estão sujeitas (Carmona & Villas Boas, 2001), uma vez que a luz é fortemente atenuada, à medida que a profundidade no solo aumenta (Canossa et al., 2007), e as espécies com sementes pequenas, geralmente, requerem luz para a germinação (Vieira, 2007). Na superfície do solo, além da maior quantidade de luz, a alternância de temperaturas também pode estimular a germinação de um grande número de espécies (Carmona & Villas Boas, 2001).
CONCLUSÕES
A emergência de Sida rhombifolia foi influenciada pelas duas épocas (setembro de 2008 e janeiro 2009) e os maiores percentuais de emergência ocorreram para sementes locadas entre 1 e 4 cm de profundidade, independentemente da época de semeadura.
Para Solanum viarum, observou-se que a época de semeadura apresentou forte relação com a emergência, nas profundidades entre 1 e 5 cm, sendo observado redução de emergência apenas para as sementes locadas na superfície do solo.
Recebido para publicação em 04/04/2011 e aprovado em 10/10/2011
- Akanda RU, Mullahey JJ & Shilling DG (1996) Environmental factors affecting germination of tropical soda apple (Solanum viarum). Weed Science, 44:570-574.
- Alberguini AL & Yamashita OM (2010) Profundidade de semeadura e presença de palha afetam a emergência de plântulas de Vernonia ferruginea Planta Daninha, 28:1005-1013.
- Brighenti AM, Voll E & Gazziero DLP (2003) Biologia e manejo do Cardiospermum halicacabum Planta Daninha, 21:229-237.
- Canossa RS, Oliveira JR RS, Constantin J. Biffe DF, Alonso DG & Franchini LHM (2007) Profundidade de semeadura afetando a emergência de plântulas de Alternanthera tenella Planta Daninha, 25:719-725.
- Carmona R & Villas Boas HDC (2001) Dinâmica de sementes de Bidens pilosa no solo. Pesquisa Agropecuaria Brasileira, 36:457-463.
- Carmona R (1992) Problemática e manejo de bancos de sementes de invasoras em solos agrícolas. Planta Daninha, 10:5-16.
- Coile NC (1993) Tropical soda apple, Solanum viarum Dunal: the plant from hell (Solanaceae). Florida: Florida Department of Agriculture e Consumer Services, 4p. (Botany Circular, 27)
- Deuber R (1992) Botânica das plantas daninhas. In: Deuber R (Ed.) Ciências das plantas daninhas. Jaboticabal, FUNEP. p.31-73.
- Dias Filho MB (1996) Germination and emergence of Stachytarpheta cayennensis and Ipomoea asarifolia Planta Daninha, 14:118-126.
- Dyer WE (1995) Exploiting weed seed dormancy and germination requirements through agronomic practices. Weed Science, 43:498-503.
- Embrapa (2006) Centro Nacional de Pesquisa Agropecuária de Solos. Sistema brasileiro de classificação de solo. Rio de Janeiro, Embrapa Solos. 306p.
- Fernández OA (1982) Manejo integrado de malezas. Planta Daninha, 5:69-79.
- Gasparim E, Ricieri RP, Silva SL, Dallacort R & Gnoatto E (2005) Temperatura no perfil do solo utilizando densidades de cobertura e solo nu. Acta Scientiarum. Agronomy, 27:107-115.
- Guimarães SC, Souza IF & Pinho EVRV (2002) Emergência de Tridax procumbens em função de profundidade de semeadura, do conteúdo de argila no substrato e da incidência de luz na semente. Planta Daninha, 20:413-419.
- Gomes FP (1984) A estatística moderna na pesquisa agropecuária. Piracicaba, POTAFOS. 160p.
- Kissmann KG & Groth D (2000) Plantas infestantes e nocivas. São Paulo, BASF. Tomo II. 725p.
- Lorenzi H (2000) Plantas daninhas do Brasil: terrestres, aquáticas, parasitas, tóxicas e medicinais. 3Ş ed. Nova Odessa, Instituto Plantarum. 608p.
- Machado Neto JG & Pitelli RA (1988) Profundidade de semeadura na emergência de amendoim-bravo. Pesquisa Agropecuaria Brasileira, 23:1203-1208.
- Maguire JD (1962) Speed of germination aid selection and evaluation for seedling emergence and vigor. Crop Science, 2:176-177.
- Medal JC, Pitelli RA, Santana A, Gandolfo D, Gravena R & Habeck DH (1999) Host specificity of Metriona elatior, a potential biological control agent of tropical soda apple, Solanum viarum, in the USA. BioControl, 44:421-436.
- Mullahey JJ & Cornell J (1994) Biology of tropical soda apple (Solanum viarum) an introduced weed in Florida. Weed Technology, 8:465-469.
- Muniz Filho A, Carneiro PT, Cavalcanti MLF & Albuquerque RC (2004) Capacidade de emergência de picão-preto em diferentes profundidades de semeadura. Revista de Biologia e Ciência da Terra, 4:1. Disponível em: <http://eduep.uepb.edu.br/rbct/sumarios/pdf/picaopreto.pdf>. Acessado em: 15 de fevereiro de 2011.
- Pacheco LP, Pires FR, Monteiro FP, Procópio SO, Assis RL, Silva GP, Cargnelutti Filho A, Carmo ML & Petter FA (2009) Emergência e crescimento de plantas de cobertura em função da profundidade de semeadura. Semina: Ciências Agrárias, 30:305-314.
- Pearson TRH, Burslem DFRP, Mullins CE & Dalling JW (2003) Functional significance of photoblastic germination in neotropical pioneer trees: a seed'seye view. Functional Ecology, 17:394-402.
- Radosevich SR, Holt JS & Ghersa CM (2007) Ecology of weeds and invasive plants: Relationship to agriculture and natural resource management. 3Ş ed. Hoboken, John Wiley & Sons. 207p.
- Roberts EH (1999) A search for pattern and form. Seed Science Research, 9:181-208.
- Simpson RL, Leck MA & Parker VT (1989) Seed banks: General concepts and methodological issues. In: Leck MA, Parker VT & Simpson RL (Eds.) Ecology of soil seed banks. London, Academic Press. p.3-8.
- Smith CA, Shaw DR & Newsom LJ (1992) Arrowleaf sida (Sida rhombifolia) and prickly sida (Sida spinosa): germination and emergence. Weed Research, 32:103-109.
- Souza Filho APS (2006) Interferência potencialmente alelopática do capim-gengibre (Paspalum maritimum) em áreas de pastagens cultivadas. Planta Daninha, 24:451-456.
- Souza MC, Pitelli RA, Simi LD & Oliveira MCJ (2009) Emergência de Bidens pilosa em diferentes profundidades de semeadura. Planta Daninha, 27:29-34.
- Takahashi N (1995) Physiology of dormancy. In: Matsuo T, Kumazawa K, Ishii R, Ishihara K & Hirata H (Eds.) Science of the rice plant. Tokyo, Food and Agriculture Policy Research Center. p.45-65.
- Toledo REB, Kuva MA & Alves PLCA (1993) Fatores que afetam a germinação e a emergência de Xanthium strumarium L.: dormência, qualidade da luz e profundidade de semeadura. Planta Daninha, 11:15-20.
- Vieira EA (2007) Tamanho de sementes e sobrevivência de plântulas em áreas de pastagens degradadas. Dissertação de Mestrado. Universidade Federal de Mato Grosso do Sul, Campo Grande, 68p.
Datas de Publicação
-
Publicação nesta coleção
17 Jan 2012 -
Data do Fascículo
Dez 2011
Histórico
-
Aceito
10 Out 2011 -
Recebido
04 Abr 2011