ABSTRACT
The objective of this study was to evaluate the responses of Raphanus sativus to the application of the herbicide tepraloxydim by analyzing photosynthetic and biometric characteristics of the plants. The experiment was conducted in a greenhouse and treatments were instituted when the plants had five expanded leaves. The herbicide tepraloxydim was applied at doses of 0, 75, 100, and 125 g of active ingredient per hectare (g a.i. ha-1). The herbicide dose of 125 g a.i. ha-1 led to a small decrease in the photosynthetic rate, water use efficiency, effective quantum yield of PSII, rate of electron transport, and the concentration of chloroplastidic pigments in the leaves of R. sativus. On the other hand, there was a small increase in CO2 concentration in the substomatal chamber. The number of branches, leaves, flowers, plant height and dry mass of the stem and flowers were reduced more prominently in response to herbicide doses than the photosynthetic characteristics. Therefore, treatment with tepraloxydim inhibits the growth and formation of the leaves, branches and flowers of R. sativus at the stage of development analyzed. However, doses of 75, and 100 g a.i. ha-1 neither compromise the photosynthetic apparatus nor the stability of cell membranes.
Keywords:
acetyl-coenzyme A carboxylase; herbicide; photosynthesis; Raphanus sativus; selectivity
INTRODUCTION
The forage turnip (Raphanus sativus L.), which belongs to the Brassicaceae family is usually used in crop rotation since it has high potential in recycling nutrients, mainly phosphorus and nitrogen (Crusciol et al., 2005Crusciol CAC, Cottica RL, Lima EDV, Andreotti M, Moro E & Marcon E (2005) Persistência de palhada e liberação de nutrientes do nabo forrageiro no plantio direto. Pesquisa Agropecuária Brasileira, 40:161-168.; Heinz et al., 2011Heinz R, Garbiate MV, Viegas Neto AL, Mota LHDS, Correia AMP & Vitorino ACT (2011) Decomposition and nutrient release of crambe and fodder radish residues. Ciência Rural, 41:1549-1555.). This species also tolerates adverse environmental conditions, such as low precipitation and high temperatures (Abdel et al., 2014Abdel CG (2014) Leaf water relation of four radish (Raphanus sativus L. var. sativus) cultivars grown in controlled cabinets under varying temperatures and irrigation levels. International Journal of Agricultural Policy and Research, 02:421-443.; Chen et al., 2014Chen WL, Yang WJ, Lo HF & Yeh DM (2014) Physiology, anatomy, and cell membrane thermostability selection of leafy radish (Raphanus sativus var. oleiformis Pers.) with different tolerance under heat stress. Scientia Horticulturae, 179:367-375. ).
Considering that about 36% of the dry matter of the forage turnip seed is composed of lipids (de Souza et al., 2010de Souza ADV, Fávaro SP, Ítavo LCV & Roscoe R (2010) Caracterização química de sementes e tortas de pinhão-manso, nabo forrageiro e crambe. Pesquisa Agropecuária Brasileira , 44:1328-1335.), this species has is a potential source of raw material for the production of biodiesel. The cost of producing biodiesel from the forage turnip is lower than the costs from other traditional crops, as for example, the soybean (Chammoun et al., 2013Chammoun N, Geller DP & Das KC (2013) Fuel properties, performance testing and economic feasibility of Raphanus sativus (Oilseed radish) biodiesel. Industrial Crops and Products, 45:155-159.). In addition, the by-product of forage turnip obtained from the extraction of oil from its seeds may be used in animal feed (de Souza et al., 2010de Souza ADV, Fávaro SP, Ítavo LCV & Roscoe R (2010) Caracterização química de sementes e tortas de pinhão-manso, nabo forrageiro e crambe. Pesquisa Agropecuária Brasileira , 44:1328-1335.). Thus, the fore mentioned characteristics demonstrate that the cultivation of forage turnip can be promising and guarantee profitability to the farmer.
In general, the maximum productivity potential of crops depends on weed management because competition with the weed community limits access to water, nutrients, and light, causing productivity losses (Rigoli et al., 2008Rigoli RP, Agostinetto D, Schaedler CE, Dal Magro T & Tironi S (2008) Relative competitive ability of wheat (Triticum aestivum) intercropped with Italian ryegrass (Lolium multiflorum) or wild radish (Raphanus raphanistrum). Planta Daninha , 26:93-100.). Although it has great potential in the agricultural scenario, the crop of the forage turnip is still little explored and to date there are no registered herbicides for forage turnip cultivated, only for the management of the genotype that is weed.
As a dicotyledonous, an alternative for the management of the weed community in the production of forage turnip may be the use of graminicides of the aryloxyphenoxypropionate (APP) and cyclohexanedione (CHD) classes (Belkebir & Benhassaine-Kesri et al., 2013Belkebir A & Benhassaine-Kesri G (2013) Sethoxydim treatment inhibits lipid metabolism and enhances the accumulation of anthocyanins in rape (Brassica napus L.) leaves. Pesticide Biochemistry and Physiology, 107:120-126.), which both inhibit Acetyl-Coenzyme A Carboxylase (ACCase) (Kaundun, 2014Kaundun SS (2014) Resistance to acetyl-CoA carboxylase-inhibiting herbicides. Pest Management Science, 70:1405-1417.). However, several species among the dicotyledons are susceptible to ACCase inhibitors, such as colza and crambe (Belkebir & Benhassaine-Kesri, 2013Belkebir A & Benhassaine-Kesri G (2013) Sethoxydim treatment inhibits lipid metabolism and enhances the accumulation of anthocyanins in rape (Brassica napus L.) leaves. Pesticide Biochemistry and Physiology, 107:120-126.; Concenço et al., 2014Concenço G, Ferreira EA, Marques RF, Nunes TC, Santos AS, Palharini WG, Marschall IR, Alves MES & Mendonça CG (2014) Características fisiológicas de Crambe abyssinica sob aplicação de herbicidas. Revista de Ciências Agrárias, 37:361-369.). These species, as well as the forage turnip, belong to the Brassicaceae family. Studies with soybean plants (Belkebir et al., 2006Belkebir A, Paepe RD, Trémolières A, Aïd F & Benhassaine-Kesri G (2006) Sethoxydim affects lipid synthesis and acetyl-CoA carboxylase activity in soybean. Journal of Experimental Botany, 57:3553-3562. ), peanut (Fayez et al., 2014Fayez KA, Radwan DEM, Mohamed AK & Abdelrahman AM (2014) Fusilade herbicide causes alterations in chloroplast ultrastructure, pigment content and physiological activities of peanut leaves. Photosynthetica, 52:548-554.), melon and cucumber (Vidal et al., 2000Vidal RA, Kruse ND, Fleck NG & Merotto Jr A (2000) Selectivity of the herbicide fluazifop-p-buthyl for cucurbitaceae. Planta Daninha , 18:413-417.), and some species of the family Geraniaceae also demonstrated the sensitivities of these species to this class of herbicides (Christopher & Holtum, 2000Christopher JT & Holtum JA (2000) Dicotyledons lacking the multisubunit form of the herbicide-target enzyme acetyl coenzyme A carboxilase may be restricted to the family Geraniaceae. Functional Plant Biology, 27:845-850.).
Plant susceptibility to ACCase inhibitor herbicides can be observed by physiological changes, such as decreased photosynthetic rate and water use efficiency and disturbances in electron transport at the photochemical stage of photosynthesis and membrane permeability (Dayan & Watson, 2011Dayan FE & Watson SB (2011) Plant cell membrane as a marker for light-dependent and light-independent herbicide mechanisms of action. Pesticide Biochemistry and Physiology , 101:182-190.; Dayan & Zaccaro, 2012Dayan FE & Zaccaro MLM (2012) Chlorophyll fluorescence as a marker for herbicide mechanisms of action. Pesticide Biochemistry and Physiology , 102:189-197.; Concenço et al., 2014Concenço G, Ferreira EA, Marques RF, Nunes TC, Santos AS, Palharini WG, Marschall IR, Alves MES & Mendonça CG (2014) Características fisiológicas de Crambe abyssinica sob aplicação de herbicidas. Revista de Ciências Agrárias, 37:361-369.). ACCase is responsible for the formation of malonyl-CoA, which is required in the synthesis of fatty acids and secondary metabolites, such as suberins and flavonoids (Kaundun, 2014Kaundun SS (2014) Resistance to acetyl-CoA carboxylase-inhibiting herbicides. Pest Management Science, 70:1405-1417.). Two forms of ACCase are generally found in plants: one heteromeric form in plastids and the other homomeric form in the cytoplasm (Sasaki & Nagano, 2004Sasaki Y & Nagano Y (2004) Plant acetyl-CoA carboxylase: structure, biosynthesis, regulation, and gene manipulation for plant breeding. Bioscience, Biotechnology, and Biochemistry, 68:1175-1175.).
In this way, enzyme inhibitor herbicides can be used safely with dicotyledons because these herbicides inhibit only the homomeric form of ACCase. The opposite occurs with the species of the Poaceae family since they have only the homomeric form of ACCase, both in the cytoplasm and in the plastids. Since about 80% of the ACCase activity in the leaves occurs in plastids, inhibition of this enzyme leads to the death of species of the family Poaceae (Délye, 2005Délye C (2005) Weed resistance to acetyl coenzyme A carboxylase inhibitors: an update. Weed Science, 53:728-746.).
The herbicide tepraloxydim (an ACCase inhibitor), belongs to the chemical group of CHD - (EZ)-(RS) -2-{1-[(2E)-3-chloroallyloxymino] propil}-3-hydroxy-5-perhydropyran-4-ylcyelohex-2-em-1-one, has a mechanism of systemic action. This herbicide is recommended in post-emergence for some dicotyledons, such as soybeans, cotton, and common beans (maximum dose of 0.5 liter per hectare) (Rodrigues & Almeida, 2011Rodrigues BN & Almeida FS (2011) Guia de herbicidas. 6ª ed. Londrina, Edição dos autores. 697p.). However, the occurrence of sensitivity in dicotyledon species to ACCase inhibitors (Belkebir & Benhassaine-Kesri, 2013Belkebir A & Benhassaine-Kesri G (2013) Sethoxydim treatment inhibits lipid metabolism and enhances the accumulation of anthocyanins in rape (Brassica napus L.) leaves. Pesticide Biochemistry and Physiology, 107:120-126.; Concenço et al., 2014Concenço G, Ferreira EA, Marques RF, Nunes TC, Santos AS, Palharini WG, Marschall IR, Alves MES & Mendonça CG (2014) Características fisiológicas de Crambe abyssinica sob aplicação de herbicidas. Revista de Ciências Agrárias, 37:361-369.) reinforces the need for studies on the selectivity of the crop of interest for proper management of grasses before specific herbicides can be recommended and thus avoid losses for farmers (Vidal et al., 2000Vidal RA, Kruse ND, Fleck NG & Merotto Jr A (2000) Selectivity of the herbicide fluazifop-p-buthyl for cucurbitaceae. Planta Daninha , 18:413-417.).
Considering that the forage turnip is a member of the Brassicaceae, as are the dicotyledons crambe and colza, which have been shown to be sensitive to ACCase inhibitors (Belkebir & Benhassaine-Kesri, 2013Belkebir A & Benhassaine-Kesri G (2013) Sethoxydim treatment inhibits lipid metabolism and enhances the accumulation of anthocyanins in rape (Brassica napus L.) leaves. Pesticide Biochemistry and Physiology, 107:120-126.; Concenço et al., 2014Concenço G, Ferreira EA, Marques RF, Nunes TC, Santos AS, Palharini WG, Marschall IR, Alves MES & Mendonça CG (2014) Características fisiológicas de Crambe abyssinica sob aplicação de herbicidas. Revista de Ciências Agrárias, 37:361-369.), we evaluated the hypothesis that the herbicide tepraloxydim, representative of the chemical group of CHD, in the doses of 75, 100, and 125 g a.i. ha-1 may cause damage the photosynthetic and biometric characteristics of R. sativus plants. Therefore, the objective of this study was to evaluate the responses of R. sativus plants to the application of the herbicide tepraloxydim by analyzing photosynthetic characteristics, membrane permeability, and biometric measurements.
MATERIAL AND METHODS
Plant Material and Experimental Conditions
The experiment was conducted under a randomized block design in a greenhouse. Seeds of Raphanus sativus (cultivar CATIAL 1000) were sown in a mixture of two parts of dystroferric red latosol with one part fine sand, with the purpose of having one plant per pot. Each pot containing 4 dm3 of substrate, 2 g of dolomitic limestone were added, with a relative neutralization power of 100%. Subsequently, fertilization was carried out with 2.8 g of urea, 1.7 g of mono-ammonium phosphate, 2 g of potassium chloride, 0.9 g of magnesium sulfate, 0.02 g of copper sulfate, 0.14 g of zinc sulfate, and 0.03 g of boric acid. The liming and adduction were performed based on the results of the chemical and physical analyses of the substrate, which presented the following composition: pH H2O - 5.8; P = 0.9 mg dm-3; K = 9 mg dm-3; Ca = 0.59 cmolcdm-3; Mg = 0.17 cmolcdm-3; Al = 0.05 cmolcdm-3; H + Al = 1.8 cmolcdm-3; S = 0.8 mg dm-3; B = 0.1 mg dm-3; Cu = 0.5 mg dm-3; Fe = 118 mg dm-3; Mn= 16.7 mg dm-3; Zn = 0.2 mg dm-3; Na = 1.8 mg dm-3; base saturation= 30%; cation exchange capacity = 2.6 cmolcdm-3; organic matter = 6.2%; clay = 38.5%; silt = 7.5%; and sand = 54%.
The herbicide tepraloxydim was applied at 26 days after sowing R. sativus (4 to 5 leaves). The treatments consisted of the following doses: 0 (only water), 75, 100, and 125 grams of active ingredient per hectare (g a.i. ha-1). These doses correspond to 0, 75, 100, and 125%, respectively; of the recommended dose of the product for the bean, cotton, and soybean crops, as well as R. sativus, all of which are dicotyledons. Mineral oil was used in the spray mixture at the concentration of 0.5%. For application, a sprayer was used with constant pressure, maintained by compressed CO2, with a bar containing four spray nozzles and nozzle series. The volume of the spray mixture used was 200 L.ha-1.
The gas exchanges were measured at 3, 7 and 11 days after application of the herbicide (DAAH). The chlorophyll a fluorescence image and the concentration of chloroplastidic pigments were analyzed at 11 DAAH, whereas the biometric characteristics of the plants were determined at 12 DAAH.
Gas exchange
Gas exchange from R. sativus plants was measured in fully expanded leaves to determine the net photosynthetic rate (A, µmol CO2 m-2 s-1), stomatal conductance (g s , mol H2O m-2 s-1), transpiration rate (E, mmol H2O m-2 s-1) and ratio between internal (C i ) and external (C a ) CO2 concentrations (C i /C a ). The water use efficiency (WUE) was calculated as the ratio between A and E. The parameters were measured using an infrared gas analyzer (IRGA, model LI6400xt, Li-Cor, Nebraska, EUA). The measurements of A, g s, E, and C i /C a were performed between 8:00 and 11:00 am under constant photosynthetically active radiation (PAR, 1000 µmol photons m-2 s-1) and temperature (25 °C).
Chlorophyll a fluorescence
The data and images of chlorophyll a fluorescence were measured and obtained in the same leaf of the photosynthesis using a modulated imaging-PAM fluorometer (Heinz, Walz, Effeltrich, Germany). For the measurements, leaves were initially dark-acclimated for 40 min so that the reaction centers were fully opened to obtain the minimal (F 0) and maximal chlorophyll fluorescence (F M). From these values, the potential quantum yield of PSII [F V/F M = (F M - F 0 )/F M] was calculated according to Genty et al. (1989Genty B, Briantais JM & Baker NR (1989) The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimicaet Biophysica Acta, 990:87-92.). After sample illumination, saturation pulses were applied to determine the light-acclimated variables: the quantum yield of photochemical energy conversion in PSII (Y II), the quenching of regulated (Y NPQ), and the non-regulated (Y NO) non-photochemical dissipation. The Y II was also used to estimate the apparent electron transport rate (ETR = Y II x PAR X A leaf x 0.5) (Bilger et al., 1995Bilger W, Schreiber U & Bock M (1995) Determination of the quantum efficiency of photosystem II and of non-photochemical quenching of chlorophyll fluorescence in the field. Oecologia, 102:425-432.), where PAR is the photon flux (µmol m-2 s-1) on the leaves, A leaf is the amount corresponding to the fraction of incident light absorbed by the leaves, and 0.5 is the excitation energy fraction directed to the PSII (Laisk & Loreto, 1996Laisk A & Loreto F (1996) Determining photosynthetic parameters from leaf CO2 exchange and chlorophyll fluorescence (ribulose-1, 5-bisphosphate carboxylase/oxygenase) specificity factor, dark respiration in the light, excitation distribution between photosystems, alternative electron transport rate, and mesophyll diffusion resistance. Plant Physiology, 110:903-912.).
Photosynthetic pigments
Chlorophyll a, chlorophyll b, total chlorophyll, and carotenoids were measured according to Costa et al. (2014Costa AC, Rosa M, Megguer CA, Silva FG, Pereira FD & Otoni WC (2014) A reliable methodology for assessing the in vitro photosynthetic competence of two Brazilian savanna species: Hyptis marrubioides and Hancornia speciosa. Plant Cell, Tissue and Organ Culture (PCTOC), 117:443-454.). Three leaf discs (0.6 cm2) were immersed in 5 mL of a dimethyl sulfoxide (DMSO) solution saturated with calcium carbonate and incubated at 65ᴼC for 24 hours. Absorbance was measured at 665.1, 649.1, and 480.0 nm in a UV-VIS spectrophotometer (model 60S, Thermo Scientific, Madison, USA), and the concentrations were calculated according to Wellburn (1994Wellburn AR (1994) The Spectral Determination of Chlorophylls a and b, As Well As Total Carotenoids, UsingVarious Solvents with Spectrophotometers of Different Resolution. Journal Plant Physiology , 144:307-313.) and expressed by the leaf area.
Rate of electrolyte leakage
Leaf discs (0.6 cm-2) were immersed in ultrapure water, and the initial conductivity was measured after 24 h. The samples were then kept at 100 °C for 1 h, and the final conductivity was evaluated. The electrical conductivity of the leaf sample was measured using a conductivity meter (CD-850 model, Instrutherm, Brazil), and the rate of electrolyte leakage (%) was calculated according to Silva et al., (2014Silva FB, Costa AC, Alves RRP & Megguer CA (2014) Chlorophyll fluorescence as an indicator of cellular damage by glyphosate herbicide in Raphanus sativus L. plants. American Journal of Plant Sciences, 05:2509-2519.).
Biometric analyses
The plants were measured to determine their height (H, m), number of leaves (NL), number of nodes (NN), number of branches (NB), number of flowers (NF), and stem diameter (SD, mm). Leaves, stems and flowers were separated, packed in paper bags and dried in a forced-air-circulation oven (65 °C) for 48 h to obtain leaf dry matter (LDM, g), stem dry matter (SDM, g), and flowers dry matter (FDM, g).
Statistical analyses
The obtained data were submitted to analysis of variance and to the adjustment of the logistic nonlinear regression model, represented by the following formula: Ŷ = a / (1+ (x / b) c), where a = value of the analyzed variable in the lowest dose of the herbicide, b = dose of the herbicide responsible for the 50% decrease of the analyzed variable (I50), and c = slope of the curve around the I50 (de Souza et al., 2000de Souza AP, Ferreira FA, da Silva AA, Cardoso AA & Ruiz HA (2000) Uso da equação logística no estudo de dose-resposta de glyphosate e imazapyr por bioensaios. Planta Daninha, 18:17-28.). For the variables that did not fit the logistic model, a linear model was used. Action Stat 3 software was used for the statistical analysis of the data, and the graphics were made using the software SigmaPlot V.10 (SPSS Inc., USA).
RESULTS AND DISCUSSION
The application of the herbicide tepraloxydim at the lowest doses did not cause visual damage to the leaves of R. sativus plants (Figures 1A, B, and C). However, the dose of 125 g a.i. ha-1 affected the formation of new leaves, evidenced by the leaf wrinkling symptoms (Figure 1D). These symptoms of wrinkling on young leaves, observed at seven DAAH, suggest that the synthesis of lipids, which are essential constituents of membranes, was limited. Thus, heteromeric ACCase probably did not supply all the acetyl-CoA carboxylation to malonyl-CoA that was required, compromising leaf growth zones by inadequate formation of cell membranes and organelles (Délye, 2005Délye C (2005) Weed resistance to acetyl coenzyme A carboxylase inhibitors: an update. Weed Science, 53:728-746.).

Symptoms on leaves of Raphanus sativus 7 days after application of the herbicide tepraloxydim at doses of 0 (A), 75 (B), 100 (C), and 125 g a.i. ha-1 (D).
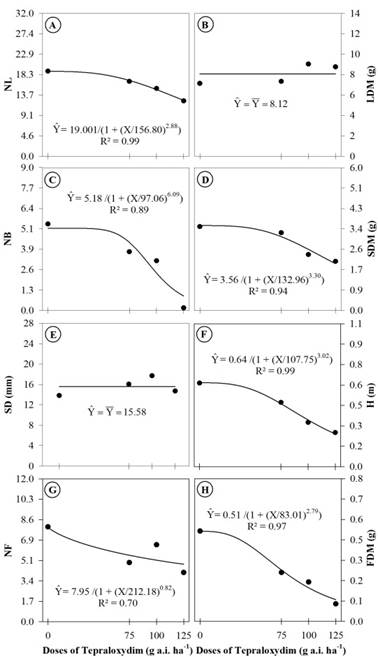
The probable inhibition of the homomeric ACCase by the herbicide tepraloxydim also compromised the growth and development of R. sativus. Therefore, the formation of new structures, such as branches and flowers, and the height of the plants were inhibited by the application of the different doses of the herbicide (Figure 2).

Number of leaves (NL) (A), leaf dry matter (LDM) (B), number of branches (NB) (C), stem dry matter (SDM) (D), stem diameter (SD) (E), plant height (H) (F), number of flowers (NF) (G), and flowers dry matter of (FDM) (H) of Raphanus sativus plants treated with different doses of the herbicide tepraloxydim. Data are means of (n = 6).
The numbers of leaves (NL), branches (NB) and flowers (NF) as well as the dry matter weights of the stems (SDM), flowers (FDM), and the plant height (H) all decreased as a function of the doses of the herbicide tepraloxydim (Figure 2A, C, G, D, H, and F). As indicated by the lower values of I50, which represent the dose of the herbicide responsible for the 50% decrease of the analyzed variable (de Souza et al., 2000de Souza AP, Ferreira FA, da Silva AA, Cardoso AA & Ruiz HA (2000) Uso da equação logística no estudo de dose-resposta de glyphosate e imazapyr por bioensaios. Planta Daninha, 18:17-28.), that the most sensitive biometric characteristics were FDM (83.01 g a.i. ha-1), NB (97.06 g a.i. ha-1), and H (107.75 g a.i. ha-1). However, it was not possible to adjust leaf dry matter (LDM) to the regression model and R. sativus stem diameter (SD) did not differ as a function of the herbicide dose (Figure 2B and E). In this sense, Vidal et al. (2000Vidal RA, Kruse ND, Fleck NG & Merotto Jr A (2000) Selectivity of the herbicide fluazifop-p-buthyl for cucurbitaceae. Planta Daninha , 18:413-417.) also observed reductions in the biometric characteristics of cucumber and melon plants. The leaf area and total dry matter of the plants were reduced in response to application of the ACCase inhibitor herbicide fluazifop-p-butyl (Vidal et al., 2000Vidal RA, Kruse ND, Fleck NG & Merotto Jr A (2000) Selectivity of the herbicide fluazifop-p-buthyl for cucurbitaceae. Planta Daninha , 18:413-417.). The decrease of LDM (approximately 50%) in Brassica napus plants treated with the herbicide sethoxydim was also reported by Belkebir & Benhassaine-Kesri (2013Belkebir A & Benhassaine-Kesri G (2013) Sethoxydim treatment inhibits lipid metabolism and enhances the accumulation of anthocyanins in rape (Brassica napus L.) leaves. Pesticide Biochemistry and Physiology, 107:120-126.). However, in this work with R. sativus plants, there was no change in LDM.
There were a reduction in the chlorophyll and carotenoid content in R. sativus in response to the doses of the herbicide tepraloxydim (Figure 3). This supports the hypothesis that this herbicide restricted the biosynthesis of lipids, which are essential constituents of membranes, including chloroplast membranes. Corroborating this hypothesis, Fayez et al. (2014Fayez KA, Radwan DEM, Mohamed AK & Abdelrahman AM (2014) Fusilade herbicide causes alterations in chloroplast ultrastructure, pigment content and physiological activities of peanut leaves. Photosynthetica, 52:548-554.) reported that the herbicide fluazifop-p-butyl also reduced the concentration of chloroplastidic pigments from leaves of newly formed peanut plants. Similarly, the chlorophyll content of leaves of B. napus was reduced in response to treatment with the herbicide sethoxydim (Belkebir & Benhassaine-Kesri, 2013Belkebir A & Benhassaine-Kesri G (2013) Sethoxydim treatment inhibits lipid metabolism and enhances the accumulation of anthocyanins in rape (Brassica napus L.) leaves. Pesticide Biochemistry and Physiology, 107:120-126.). Although there was a reduction in the chloroplast pigment content in R. sativus in this study, this was not enough to trigger chlorosis and foliar necrosis, which was observed in peanut plants (Fayez et al., 2014Fayez KA, Radwan DEM, Mohamed AK & Abdelrahman AM (2014) Fusilade herbicide causes alterations in chloroplast ultrastructure, pigment content and physiological activities of peanut leaves. Photosynthetica, 52:548-554.).

Concentrations of chlorophyll a (A), chlorophyll b (B), carotenoids (C), and total chlorophyll (D) of Raphanus sativus plants submitted to different doses of the herbicide tepraloxydim. Data are means of (n = 6).
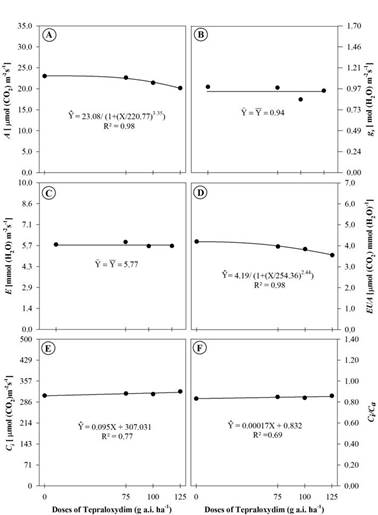
In addition to the biometric variables and the chloroplast pigment content, the parameters of gas exchanges photosynthetic rate (A), instantaneous water use efficiency (WUE), internal concentration of CO2 (C i ), and the ratio between the internal and external CO2 concentration (C i /C a ) differed as a function of the doses of tepraloxydim (Figure 4A, D, E, and F). There was also a difference as a function of the DAAH factor since the measurements were performed at 3, 7, and 11 DAAH. These variations in A, g s , WUE and transpiration rate (E) are due exclusively to the environmental conditions at the time of analysis and to the differentiation of the plant material as a function of the phenological stage change (Figure 5A, B, C, and D). Variables A and WUE decreased in response to increased herbicide rates and C i , and C i /C a increased. However, these changes were not of high order, with the decrease and increase more accentuated only at the higher dose of 125 g a.i. ha-1.

Photosynthetic rate (A) (A), stomatal conductance (g s ) (B), transpiration rate (E) (C), water use efficiency (WUE) (D), CO2 internal concentration (C i ) (E), and ratio between internal and external CO2 concentration (C i /C a ) (F) of Raphanus sativus plants submitted to different doses of the herbicide tepraloxydim. Data are means of (n = 18).

Photosynthetic rate (A) (A), stomatal conductance (g s ) (B), water use efficiency (WUE) (C), and transpiration rate (E) (D) of Raphanus sativus plants at 3, 7, and 11 days after application of the herbicide tempraloxydim. The bars represent the standard error of the means (n = 24).
The reduction in A in young leaves of R. sativus in response to ACCase inhibitor action was also reported in Crambe abyssinica plants treated with the herbicides sethoxydim, fluazifop-p-butyl, and clethodim (Concenço et al., 2014Concenço G, Ferreira EA, Marques RF, Nunes TC, Santos AS, Palharini WG, Marschall IR, Alves MES & Mendonça CG (2014) Características fisiológicas de Crambe abyssinica sob aplicação de herbicidas. Revista de Ciências Agrárias, 37:361-369.). Already Xia et al. (2006Xia XJ, Huang YY, Wang L, Huang LF, Yu YL, Zhou YH & Yu JQ (2006) Pesticides-induced depression of photosynthesis was alleviated by 24-epibrassinolide pretreatment in Cucumis sativus L. Pesticide Biochemistry and Physiology , 86:42-48) reported a marked decrease (up to 63%) in CO2 assimilation in cucumber plants treated with the fluazifop-p-butyl and haloxyfop-p-methyl herbicides at doses of about 40 and 27 g a.i. ha-1, respectively. Considering that ACCase inhibitors block lipid synthesis, the action of these herbicides on the photosynthetic process is indirect. This indirect action can be observed in this study, because it required a high dose of the herbicide tepraloxydim to reduce A by 50% in R. sativus, according to the I50 of 220.77 g a.i. ha-1.
The reduction in the WUE in this study is mainly due to the decrease in CO2 assimilation since E did not differ in response to herbicide doses (Figure 4D and C). Corroborating these results, the accumulation of CO2 in the substomatal chamber in response to the herbicide, as evidenced by the small increase in C i and in the C i /C a ratio, demonstrates that the limitation in A was due, at least in part, to the impairment of CO2 assimilation by inhibition of the Calvin-Benson cycle.
The chlorophyll a fluorescence measures are good indicators of herbicide-promoted stresses to the photosynthetic apparatus (Silva et al., 2014Silva FB, Costa AC, Alves RRP & Megguer CA (2014) Chlorophyll fluorescence as an indicator of cellular damage by glyphosate herbicide in Raphanus sativus L. plants. American Journal of Plant Sciences, 05:2509-2519.; Lima et al., 2017Lima DA, Müller C, Costa AC, Batista PF, Dalvi VC & Domingos M (2017) Morphoanatomical and physiological changes in Bauhinia variegata L. as indicators of herbicide diuron action. Ecotoxicology and environmental safety, 141:242-250.). In this study, the chlorophyll a fluorescence parameters of minimal fluorescence (F 0), relative electron transport rate (ETR), and the effective quantum yield of PSII (Y II) of R. sativus differed according to the increasing doses of the herbicide tepraloxydim (Figure 6A-B, E-F, and G-H). However, the potential quantum yield of PSII (F V/F M), quantum yield of regulated energy dissipation of PSII (Y NPQ), and quantum yield of non-regulated energy dissipation of PSII (Y NO) did not differ (Figure 6C-D, I-J, and K-L). Thus, the F V/F M values, which remained around 0.8, show that there was no photoinhibition (Maxwell & Johnson, 2000Maxwell K & Johnson GN (2000) Chlorophyll Fluorescence: A Practical Guide. Journal Experimental Botany, 51:659-668.). Similar values for this parameter of F V/F M were visualized by Silva et al. (2014) and Moura et al. (2018Moura LMF, Costa AC, Müller C, Silva-Filho RO, Almeida GM, Vital RG, Castro JN & Teixeira MB (2018) Drought tolerance in potential oilseed plants for biofuel production. Australian Journal of Crop Science, 12:289-298.) in R. sativus plants, when compared to the control plants, proving that these were ideal values for this species. The increase of F 0 as a function of the increase of the doses of the herbicide tepraloxydim, even in a non-high order, is an indication of the limitations in the transfer of the excitation energy from the antenna pigments to the reaction center (Lima et al., 2017; Batista et al., 2018Batista PF, Costa AC, Müller C, Silva-Filho RO, Silva FB, Merchant A, Mendes GC & Nascimento KJT (2018) Nitric oxide mitigates the effect of water deficit in Crambe abyssinica. Plant Physiology and Biochemistry, 129:310-322.).

Minimal fluorescence (F 0) (A and B), potential quantum yield of PSII (F V/F M) (C and D), relative eléctron transport rate (ETR) (E and F), effective quantum yield of PSII (Y II) (G and H), quantum yield of regulated energy dissipation of PSII (Y NPQ) (I and J), and quantum yield of non-regulated energy dissipation of PSII (Y NO) (K and L) of Raphanus sativus plants submitted to different doses of the herbicide tepraloxydim. Data are means of (n = 6).
The reduction of A values is related to the reduction of Y II and ETR, since the energy destined for the photochemical dissipation decreased as a function of the increment of the doses of the herbicide tepraloxydim. Thus, the reduction of ATP and NADPH formation was probably the limiting factor for the Calvin-Benson cycle, since there was no stomatal limitation, as evidenced by the g s values. Unlike Y II, the Y NPQ and Y NO remained unchanged, even with increasing Y NPQ at the dose of 125 g a.i. ha-1, this could not be adjusted to the regression model. Y NPQ and Y NO have the function of dissipating the energy absorbed in the form of heat by the xanthophyll cycle, which is activated by the protonation of the thylakoid lumen (Ruban, 2016Ruban AV (2016) Nonphotochemical chlorophyll fluorescence quenching: mechanism and effectiveness in protecting plants from photodamage. Plant Physiology , 170:1903-1916. ). This protonation leads to the de-epoxidation of vialaxanthin, forming anteraxanthin, which forms zeaxanthin (Kromdijk et al., 2016Kromdijk J, Głowacka K, Leonelli L, Gabilly ST, Iwai M, Niyogi KK & Long SP (2016) Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science, 354:857-861.; Mathur et al., 2018Mathur S, Jain L & Jajoo A (2018) Photosynthetic efficiency in sun and shade plants. Photosynthetica , 56:354-365.). This dissipation of energy is a way of protecting the photosynthetic apparatus against the excess energy of excitation in general and the formation of reactive oxygen species in particular (Cardona et al., 2018Cardona T, Shao S & Nixon PJ (2018) Enhancing photosynthesis in plants: the light reactions. Essays in biochemistry, 62:85-94.).
The rate of electrolyte liberation (REL) of R. sativus plants was not altered by the herbicide tepraloxydim (Figure 7). This is an indication that there was no damage at the membrane level (Duke & Kenyon, 1993Duke SO & Kenyon WH (1993) Peroxidizing activity determined by cellular leakage. In: Böger P & Sandmann G (Ed.) Target assays for modern herbicides and related phytotoxic compouds. Boca Raton, CRC Press. p.61-66.). Thus, corroborating the chlorophyll a fluorescence measurements, in which there was no reduction of the F V/F M values, thereby indicating the stability of PSII and the chloroplast membranes.

Rate of electrolyte leakage (REL) of Raphanus sativus plants submitted to different doses of the herbicide tepraloxydim. Data are means of (n = 6).
CONCLUSION
The doses of 125 g a.i. ha-1 of the herbicide tepraloxydim mainly inhibited the growth and formation of leaves, branches and flowers of R. sativus at the development stage analyzed in this study. However, doses of 75 and 100 g a.i. ha-1 do not compromise the photosynthetic apparatus and the stability of cell membranes.
ACKNOWLEDGMENTS
The authors are grateful to the CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico, Brazil), CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil), FAPEG (Fundação de Amparo à Pesquisa do Estado de Goiás), and Instituto Federal Goiano - Campus Rio Verde for financial support. In addition, the authors inform that there is no conflict of interest in carrying the research and publishing the manuscript.
REFERENCES
- Abdel CG (2014) Leaf water relation of four radish (Raphanus sativus L. var. sativus) cultivars grown in controlled cabinets under varying temperatures and irrigation levels. International Journal of Agricultural Policy and Research, 02:421-443.
- Batista PF, Costa AC, Müller C, Silva-Filho RO, Silva FB, Merchant A, Mendes GC & Nascimento KJT (2018) Nitric oxide mitigates the effect of water deficit in Crambe abyssinica Plant Physiology and Biochemistry, 129:310-322.
- Belkebir A & Benhassaine-Kesri G (2013) Sethoxydim treatment inhibits lipid metabolism and enhances the accumulation of anthocyanins in rape (Brassica napus L.) leaves. Pesticide Biochemistry and Physiology, 107:120-126.
- Belkebir A, Paepe RD, Trémolières A, Aïd F & Benhassaine-Kesri G (2006) Sethoxydim affects lipid synthesis and acetyl-CoA carboxylase activity in soybean. Journal of Experimental Botany, 57:3553-3562.
- Bilger W, Schreiber U & Bock M (1995) Determination of the quantum efficiency of photosystem II and of non-photochemical quenching of chlorophyll fluorescence in the field. Oecologia, 102:425-432.
- Cardona T, Shao S & Nixon PJ (2018) Enhancing photosynthesis in plants: the light reactions. Essays in biochemistry, 62:85-94.
- Chammoun N, Geller DP & Das KC (2013) Fuel properties, performance testing and economic feasibility of Raphanus sativus (Oilseed radish) biodiesel. Industrial Crops and Products, 45:155-159.
- Chen WL, Yang WJ, Lo HF & Yeh DM (2014) Physiology, anatomy, and cell membrane thermostability selection of leafy radish (Raphanus sativus var. oleiformis Pers.) with different tolerance under heat stress. Scientia Horticulturae, 179:367-375.
- Christopher JT & Holtum JA (2000) Dicotyledons lacking the multisubunit form of the herbicide-target enzyme acetyl coenzyme A carboxilase may be restricted to the family Geraniaceae. Functional Plant Biology, 27:845-850.
- Concenço G, Ferreira EA, Marques RF, Nunes TC, Santos AS, Palharini WG, Marschall IR, Alves MES & Mendonça CG (2014) Características fisiológicas de Crambe abyssinica sob aplicação de herbicidas. Revista de Ciências Agrárias, 37:361-369.
- Costa AC, Rosa M, Megguer CA, Silva FG, Pereira FD & Otoni WC (2014) A reliable methodology for assessing the in vitro photosynthetic competence of two Brazilian savanna species: Hyptis marrubioides and Hancornia speciosa Plant Cell, Tissue and Organ Culture (PCTOC), 117:443-454.
- Crusciol CAC, Cottica RL, Lima EDV, Andreotti M, Moro E & Marcon E (2005) Persistência de palhada e liberação de nutrientes do nabo forrageiro no plantio direto. Pesquisa Agropecuária Brasileira, 40:161-168.
- Dayan FE & Watson SB (2011) Plant cell membrane as a marker for light-dependent and light-independent herbicide mechanisms of action. Pesticide Biochemistry and Physiology , 101:182-190.
- Dayan FE & Zaccaro MLM (2012) Chlorophyll fluorescence as a marker for herbicide mechanisms of action. Pesticide Biochemistry and Physiology , 102:189-197.
- de Souza ADV, Fávaro SP, Ítavo LCV & Roscoe R (2010) Caracterização química de sementes e tortas de pinhão-manso, nabo forrageiro e crambe. Pesquisa Agropecuária Brasileira , 44:1328-1335.
- de Souza AP, Ferreira FA, da Silva AA, Cardoso AA & Ruiz HA (2000) Uso da equação logística no estudo de dose-resposta de glyphosate e imazapyr por bioensaios. Planta Daninha, 18:17-28.
- Délye C (2005) Weed resistance to acetyl coenzyme A carboxylase inhibitors: an update. Weed Science, 53:728-746.
- Duke SO & Kenyon WH (1993) Peroxidizing activity determined by cellular leakage. In: Böger P & Sandmann G (Ed.) Target assays for modern herbicides and related phytotoxic compouds. Boca Raton, CRC Press. p.61-66.
- Fayez KA, Radwan DEM, Mohamed AK & Abdelrahman AM (2014) Fusilade herbicide causes alterations in chloroplast ultrastructure, pigment content and physiological activities of peanut leaves. Photosynthetica, 52:548-554.
- Genty B, Briantais JM & Baker NR (1989) The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimicaet Biophysica Acta, 990:87-92.
- Heinz R, Garbiate MV, Viegas Neto AL, Mota LHDS, Correia AMP & Vitorino ACT (2011) Decomposition and nutrient release of crambe and fodder radish residues. Ciência Rural, 41:1549-1555.
- Kaundun SS (2014) Resistance to acetyl-CoA carboxylase-inhibiting herbicides. Pest Management Science, 70:1405-1417.
- Kromdijk J, Głowacka K, Leonelli L, Gabilly ST, Iwai M, Niyogi KK & Long SP (2016) Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science, 354:857-861.
- Laisk A & Loreto F (1996) Determining photosynthetic parameters from leaf CO2 exchange and chlorophyll fluorescence (ribulose-1, 5-bisphosphate carboxylase/oxygenase) specificity factor, dark respiration in the light, excitation distribution between photosystems, alternative electron transport rate, and mesophyll diffusion resistance. Plant Physiology, 110:903-912.
- Lima DA, Müller C, Costa AC, Batista PF, Dalvi VC & Domingos M (2017) Morphoanatomical and physiological changes in Bauhinia variegata L. as indicators of herbicide diuron action. Ecotoxicology and environmental safety, 141:242-250.
- Mathur S, Jain L & Jajoo A (2018) Photosynthetic efficiency in sun and shade plants. Photosynthetica , 56:354-365.
- Maxwell K & Johnson GN (2000) Chlorophyll Fluorescence: A Practical Guide. Journal Experimental Botany, 51:659-668.
- Moura LMF, Costa AC, Müller C, Silva-Filho RO, Almeida GM, Vital RG, Castro JN & Teixeira MB (2018) Drought tolerance in potential oilseed plants for biofuel production. Australian Journal of Crop Science, 12:289-298.
- Rigoli RP, Agostinetto D, Schaedler CE, Dal Magro T & Tironi S (2008) Relative competitive ability of wheat (Triticum aestivum) intercropped with Italian ryegrass (Lolium multiflorum) or wild radish (Raphanus raphanistrum). Planta Daninha , 26:93-100.
- Rodrigues BN & Almeida FS (2011) Guia de herbicidas. 6ª ed. Londrina, Edição dos autores. 697p.
- Ruban AV (2016) Nonphotochemical chlorophyll fluorescence quenching: mechanism and effectiveness in protecting plants from photodamage. Plant Physiology , 170:1903-1916.
- Sasaki Y & Nagano Y (2004) Plant acetyl-CoA carboxylase: structure, biosynthesis, regulation, and gene manipulation for plant breeding. Bioscience, Biotechnology, and Biochemistry, 68:1175-1175.
- Silva FB, Costa AC, Alves RRP & Megguer CA (2014) Chlorophyll fluorescence as an indicator of cellular damage by glyphosate herbicide in Raphanus sativus L. plants. American Journal of Plant Sciences, 05:2509-2519.
- Vidal RA, Kruse ND, Fleck NG & Merotto Jr A (2000) Selectivity of the herbicide fluazifop-p-buthyl for cucurbitaceae. Planta Daninha , 18:413-417.
- Wellburn AR (1994) The Spectral Determination of Chlorophylls a and b, As Well As Total Carotenoids, UsingVarious Solvents with Spectrophotometers of Different Resolution. Journal Plant Physiology , 144:307-313.
- Xia XJ, Huang YY, Wang L, Huang LF, Yu YL, Zhou YH & Yu JQ (2006) Pesticides-induced depression of photosynthesis was alleviated by 24-epibrassinolide pretreatment in Cucumis sativus L. Pesticide Biochemistry and Physiology , 86:42-48
-
1
This work is part of the project "Study of the potential of crambe (Crambe abyssinica) and forage turnip (Raphanus sativus) for biofuels: phytotechnical, physiological characterization and quality of seeds and oil". PNPD Project N°. 02737/09 - 4 - MEC / CAPES line.
Publication Dates
-
Publication in this collection
02 Mar 2020 -
Date of issue
Jan-Feb 2020
History
-
Received
28 Feb 2019 -
Accepted
30 Nov 2019