SUMMARY
Malaria is a major health problem for people who live on the border between Brazil and French Guiana. Here we discuss Plasmodium vivax distribution pattern in the town of Oiapoque, Amapá State using the circumsporozoite (CS) gene as a marker. Ninety-one peripheral blood samples from P. vivax patients have been studied. Of these, 64 individuals were from the municipality of Oiapoque (Amapá State, Brazil) and 27 patients from French Guiana (August to December 2011). DNA extraction was performed, and a fragment of the P. vivax CS gene was subsequently analyzed using PCR/RFLP. The VK210 genotype was the most common in both countries (48.36% in Brazil and 14.28% in French Guiana), followed by the P. vivax-like (1.10% in both Brazil and French Guiana) and VK247 (1.10% only in Brazil) in single infections. We were able to detect all three CS genotypes simultaneously in mixed infections. There were no statistically significant differences either regarding infection site or parasitaemia among individuals with different genotypes. These results suggest that the same genotypes circulating in French Guiana are found in the municipality of Oiapoque in Brazil. These findings suggest that there may be a dispersion of parasitic populations occurring between the two countries. Most likely, this distribution is associated with prolonged and/or more complex transmission patterns of these genotypes in Brazil, bordering French Guiana.
KEYWORDS:
Plasmodium vivax; Circumsporozoite protein; Brazil-French Guiana border; Genetic marker
Despite continuous efforts to control malaria, outside Africa, more malaria cases are caused by Plasmodium vivax, resulting in a daunting morbidity and economic burden for many countries across Asia and the Americas, especially in rural areas11. World Health Organization. World malaria report 2011. [cited 2012 July 24]. Available from: Available from: http://www.who.int/malaria/world_malaria_report_2011/en
http://www.who.int/malaria/world_malaria...
. Nowadays, P. vivax malaria transmission has decreased during the last decade, although its distribution persists and is heterogeneous in different areas22. González-Cerón L, Rodriguez MH, Nettel JC, Villarreal C, Kain KC, Hernandez JE. Differential susceptibilities of Anopheles albimanus and Anopheles pseudopunctipennis to infections with coindigenous Plasmodium vivax variants VK210 and VK247 in southern Mexico. Infect Immun. 1999;67:410-2..
Among Amazon countries, Brazil has the highest proportion of malaria cases (56%)33. Stefani A, Dusfour I, Corrêa AP, Cruz MC, Dessay N, Galardo AK, et al. Land cover, land use and malaria in the Amazon: a systematic literature review of studies using remotely sensed data. Malar J. 2013;12:192.. In Brazil, gradual decreases from 2006 until 2012 have been observed. Nevertheless, P. vivax now accounts for more than 84% of clinical malaria cases annually reported in the Amazon region44. Brasil. Ministério da Saúde. Sistema de Informação de Vigilância Epidemiológica: notificação de casos. Malária. [cited 2013 Dec]. Available from: Available from: http://portalweb04.saude.gov.br/sivep_malaria/default.asp
http://portalweb04.saude.gov.br/sivep_ma...
. The circumsporozoite protein (CS) of the infective sporozoite is considered to be a major target for the development of recombinant malaria vaccine55. Herrera S, Bonelo A, Perlaza BL, Fernández OL, Victoria L, Lenis AM, et al. Safety and elicitation of humoral and cellular responses in colombian malaria-naive volunteers by a Plasmodium vivax circumsporozoite protein-derived synthetic vaccine. Am J Trop Med Hyg. 2005;73 (Suppl 5):3-9. and, this protein can be evidenced in the process of sporozoite maturation and salivary invasion in the vector as well as in human liver cells66. Coppi A, Natarajan R, Pradeln G, Bennett BL, James ER, Roggero MA, et al. The malaria circumsporozoite protein has two functional domains, each with distinct roles as sporozoites journey from mosquito to mammalian host. J Exp Med. 2011;208:341-56.,77. Suphavilai C., Looareesuwan S., Good MF. Analysis of circunsporozoite protein-specific immune responses following recent infection with Plasmodium vivax Am J Trop Med Hyg . 2004;71:29-39.. By serological and/or molecular approaches, different authors have evaluated the occurrence of P. vivax variants (VK210, VK247 and P. vivax-like) in endemic areas of the Amazon region88. Arruda ME, Zimmerman RH, Souza RM, Oliveira-Ferreira J. Prevalence and level of antibodies to the circumsporozoite protein of human malaria parasites in five states of the Amazon region of Brazil. Mem Inst Oswaldo Cruz. 2007;102:367-71.
9. Bonilla JA, Validum L, Cummings R, Palmer CJ. Genetic diversity of Plasmodium vivax PvCSP and PvMSP1 in Guyana, South America. Am J Trop Med Hyg . 2006;75:830-5.
10. Machado RL, Póvoa MM. Distribution of Plasmodium vivax variants (VK210, VK247 and P. vivax-like) in three endemic areas of the Amazon region of Brazil and their correlation with chloroquine treatment. Trans R Soc Trop Med Hyg. 2000;94:377-81.-1111. Storti-Melo LM, de Souza-Neiras WC, Cassiano GC, Joazeiro AC, Fontes CJ, Bonini-Domingos CR, et al. Plasmodium vivax circumsporozoite variants and Duffy blood group genotypes in the Brazilian Amazon region. Trans R Soc Trop Med Hyg . 2009;103:672-8.. Furthermore, the VK210 and VK247 were detected in An. aquasalis and An. darlingi in endemic areas of Pará State, Brazil12.
Border areas between countries are often characterized by intense cross-border population flows1313. Peiter PC, Machado LO, Iñiguez Rojas L. Doenças transmissíveis na faixa de fronteira Amazônica: o caso da malária. In: Miranda AC, Barcellos C, Moreira JC, Monken M, organizadores. Território, ambiente e saúde. Rio de Janeiro: Editora Fiocruz; 2008. p.257-72.. The city of Oiapoque lies on the border between French Guiana and the Brazilian State of Amapá, where there is a well-documented and intense population flux with the French municipality of Saint Georges1414. Nacher M, Stefani A, Basurko C, Lemonnier D, Djossou F, Demar M, et al. The burden of Plasmodium vivax relapses in an Amerindian village in French Guiana. Malar J . 2013;12:367.. In French Guiana malaria is endemic and distributed along the Maroni and Oiapoque rivers, whereas the coastal area bordering the Atlantic Ocean has almost no malaria transmission1414. Nacher M, Stefani A, Basurko C, Lemonnier D, Djossou F, Demar M, et al. The burden of Plasmodium vivax relapses in an Amerindian village in French Guiana. Malar J . 2013;12:367.. A serological study in seven locations of the French Guiana (Cayenne, Camopi, Maripa Soula, Saint Laurent du Maroni, Gran Santi and Sinnamary) suggested that the three variant forms of P. vivax are circulating in these areas1515. Volney B, Pouliquen JF, De Thoisy B, Fandeur T. A sero-epidemiological study of malaria in human and monkey populations in French Guiana. Acta Trop. 2002;82:11-23..
P. vivax CS genotypes VK210 and VK247 have a worldwide distribution and have been identified in several studies. However, the P. vivax-like genotype has been only detected in Papua New Guinea, Brazil, Indonesia and Madagascar1616. Parobek CM, Bailey JA, Hathaway NJ, Socheat D, Rogers WO, Juliano JJ. Differing patterns of selection and geospatial genetic diversity within two leading Plasmodium vivax candidate vaccine antigens. PLoS Negl Trop Dis. 2014;8:e2796.,1717. Qari SH, Shi YP, Póvoa MM, Alpers MP, Deloron P, Murphy GS, et al. Global occurrence of Plasmodium vivax-like human malaria parasite. J Infect Dis. 1993;168:1485-9.. In Brazil, the presence of three variant genotypes was detected in samples obtained from indigenous populations and other communities of the Amazon region. Furthermore, there is evidence from Mexico22. González-Cerón L, Rodriguez MH, Nettel JC, Villarreal C, Kain KC, Hernandez JE. Differential susceptibilities of Anopheles albimanus and Anopheles pseudopunctipennis to infections with coindigenous Plasmodium vivax variants VK210 and VK247 in southern Mexico. Infect Immun. 1999;67:410-2. and Brazil1212. da Silva AN, Santos CC, Lacerda RN, Machado RL, Póvoa MM. Susceptibility of Anopheles aquasalis and An. darlingi to Plasmodium vivax VK210 and VK247. Mem Inst Oswaldo Cruz . 2006;101:547-50. that VK210 and VK247 have differential infectivity rates in local vectors. Here we discuss P. vivax distribution pattern in the town of Oiapoque, Amapá State using the CS gene as a marker.
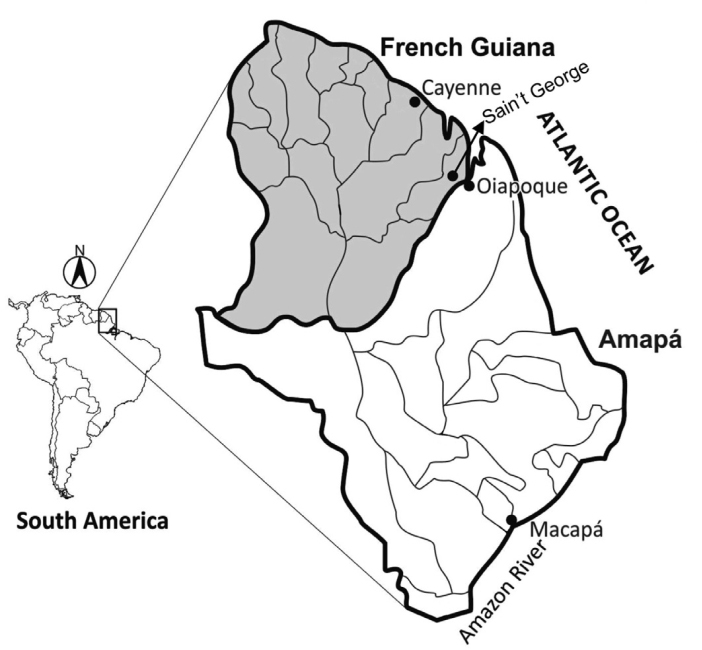
A subset of 91 patients was analyzed out of 103 individuals previously evaluated by Gomes et al.1818. Gomes MS, Vieira JL, Machado RL, Nacher M, Stefani A, Musset L, et al. Efficacy in the treatment of malaria by Plasmodium vivax in Oiapoque, Brazil, on the border with French Guiana: the importance of control over external factors. Malar J . 2015;14:402.. The peripheral blood samples, which had been kept at -70 °C, were from P. vivax carriers who lived in Oiapoque, Amapá State, a Brazilian malaria-endemic area (Fig. 1). The study took place from August to December of 2011 and was conducted by the staff of the Central Public Health Laboratory (LACEN) of the Amapá State. The patients who were enrolled in this study signed the written informed consent and fulfilled the following criteria: both genders, aged 16-60 years, they sought medical assistance due to clinical malaria symptoms, and had a positive malaria diagnosis by thick blood film or molecular techniques. Of the evaluated patients, for 64 individuals (70.33%) the infection site was the municipality of Oiapoque and for 27 patients (29.67%) it was French Guiana.
Map of the Brazilian Amazon region, showing the locations of the two populations from which isolates of Plasmodium vivax studied here were obtained: French Guiana and Oiapoque, Brazil.
The DNA was extracted from frozen pellets of infected erythrocytes using the Easy-DNA(tm) Kit (Invitrogen, Carlsbad, CA, USA), according to the manufacturer recommendations. The CS P. vivax genotypes were assessed using PCR/RFLP as previously described by Cassiano et al.1919. Cassiano GC, Storti-Melo LM, Póvoa MM, Galardo AK, Rossit AR, Machado RL. Development of PCR-RFLP assay for the discrimination of Plasmodium species and variants of P. vivax (VK210, VK247 and P. vivax-like) in Anopheles mosquitoes. Acta Trop . 2011;118:118-22.. Briefly, a reaction mix with a final volume of 25 µL containing P. vivax DNA (1.5 µL), 1 X PCR buffer (20 mM Tris-HCl pH 8.4, 50 mM KCl), 1.6 mM MgCl, 0.2 mM of each dNTP, 0.2 µM of each primer (5'-AGGCAGAGGACTTGGTGAGA-3' and 5'-CCACAGGTTACACTGCATGG-3') and 1 U of Taq Platinum. The reaction was performed in a thermocycler (DNA MasterCycler, Eppendorf, Madison, WI, USA) as follows: an initial cycle of 94 °C for 15 min, followed by 30 cycles of 94 °C for 1 min, 58 °C for 1 min and 72 °C for 1 min, with a final extension at 72 °C for 10 min. As a positive control, three plasmids were used, containing a gene insert of the repeated portion of the CSP amplification product from VK210, VK247 and P. vivax-like variants (BlueScript, Stratagene, La Jolla, USA). As the negative control of the reaction, sterile water was used. The RFLP (Restriction Fragment Length Polymorphism) reaction was performed in a final volume of 20 µL: 10 U of the restriction enzyme AluI (Invitrogen, USA), 2 µL of the reaction buffer, 10 µL of the PCR product and 7 µL of sterile DNAse-free water. Reactions were performed in a water bath at 37 °C overnight.
Analyses were performed using the R statistical software, version 2.4.1 (The R Foundation for Statistical Computing, Vienna, Austria [http://www.r-project.org]). The distribution of P. vivax CSP variants between the two studied areas was evaluated by the Chi-square test or the Fisher exact test, and the level of significance was set at p < 0.05.
The distribution of P. vivax CS genotypes and the genotypic frequencies of the 91 blood samples obtained from malaria patients are summarized in Table 1. The VK210 genotype was the commonest (62.64%), followed by P. vivax-like (2.20%) and VK247 (1.10%) in single infections. We were unable to detect all three CS genotypes simultaneously in mixed infections. However, double detections of VK210 plus VK247 (26.37%) and VK210 plus P. vivax-like (7.69%) were recorded. There were no statistically significant differences for P. vivax CS genotypes and the site of infection (Chi- square, p value = 0.1963). The parasitaemia on the thick blood films ranged from 200 to 36,000 parasites/mm3 (geometric mean + SD: 1,167.86 ± 3.32 parasites/mm3). The geometric means of parasitaemias were 1,323 parasites/mm3 (1,001- 1,750) for VK210; 1,205 parasites/mm3 (744- 1,951) for the VK210 + VK247 and 750 parasites/mm3 (295- 1,890) for VK210 + P. vivax-like infections. There were no statistically significant differences of the geometric mean of parasite density among the different genotypes detected (Table 1).
Epidemiological and genetic studies performed in malaria endemic areas of frontier malaria in Brazil could provide valuable information on parasite transmission and dispersion. The genetic diversity of the CS gene has been useful in molecular epidemiological studies, understanding transmission, dynamics and evolutionary relationships2020. Souza-Neiras WC, Storti-Melo LM, Cassiano GC, Couto VS, Couto AA, Soares IS, et al. Plasmodium vivax circumsporozoite genotypes: a limited variation or new subspecies with major biological consequences? Malar J . 2010;9:178.. Moreover, biological and genetic characteristics of the parasite, the host immunity and local vectors may influence the different patterns of demographic expansion, which are also modulated by eco-epidemiological conditions. However, the human effect can become a factor to reduce the risk of malaria, without necessarily modifying the environment33. Stefani A, Dusfour I, Corrêa AP, Cruz MC, Dessay N, Galardo AK, et al. Land cover, land use and malaria in the Amazon: a systematic literature review of studies using remotely sensed data. Malar J. 2013;12:192.,2121. González-Cerón L, Martinez-Barnetche J, Montero-Solís C, Santillán F, Soto AM, Rodríguez MH, et al. Molecular epidemiology of Plasmodium vivax in Latin America: polymorphism and evolutionary relationships of the circumsporozoite gene. Malar J . 2013;12:243..
Parasitic sampling in Oiapoque was performed, and followed a similar gradual pattern of P. vivax CS genotypes observed in others area of the Brazilian Amazon region. The three genotypes were found as single and mixed double infections, with VK210 and VK247 more frequently detected in mixed infections1010. Machado RL, Póvoa MM. Distribution of Plasmodium vivax variants (VK210, VK247 and P. vivax-like) in three endemic areas of the Amazon region of Brazil and their correlation with chloroquine treatment. Trans R Soc Trop Med Hyg. 2000;94:377-81.. Interestingly, in French Guiana Volney et al.1515. Volney B, Pouliquen JF, De Thoisy B, Fandeur T. A sero-epidemiological study of malaria in human and monkey populations in French Guiana. Acta Trop. 2002;82:11-23.performed a seroepidemiological study on malaria and found positive reactions with all P. vivax CS peptides in malaria patients from the Maroni, Oiapoque and coastal areas. Antibodies against VK210, VK247 and P. vivax- like were used. Our results suggest that this may be a common feature of frontier malaria. Thus, the same genotypes circulating in French Guiana were also found in the municipality of Oiapoque in Brazil. Moreover, the detection of VK210 subtypes circulating in the municipality of Oiapoque needs to be considered. In our research, some samples diagnosed as VK210 showed a nonspecific fragment in the RFLP gel. Recently, six different VK210 subtypes (VK210a, VK210b, VK210c, VK210d, VK210e the VK210f) were identified in isolates of P. vivax from Nicaragua and Mexico. The VK210a subtype was detected in both countries and was the most frequent. VK210f, was the less frequent, and has only been found in Nicaragua2121. González-Cerón L, Martinez-Barnetche J, Montero-Solís C, Santillán F, Soto AM, Rodríguez MH, et al. Molecular epidemiology of Plasmodium vivax in Latin America: polymorphism and evolutionary relationships of the circumsporozoite gene. Malar J . 2013;12:243.. Unfortunately, the sequencing of these samples was not performed, which is a limitation that we acknowledge. Molecular investigations are currently being conducted to understand the role of VK210 subtypes in malaria epidemiology in bordering French Guiana.
This pattern of CS genotypes distribution at the border between Brazil and French Guiana, can bring serious implications to the strategies for effective malaria control. First, approximately 40% of P. vivax malaria cases detected in Oiapoque are from French Guiana44. Brasil. Ministério da Saúde. Sistema de Informação de Vigilância Epidemiológica: notificação de casos. Malária. [cited 2013 Dec]. Available from: Available from: http://portalweb04.saude.gov.br/sivep_malaria/default.asp
http://portalweb04.saude.gov.br/sivep_ma...
. From another perspective, the P. vivax treatment prescription used in Brazil is not the same applied in French Guiana. In Brazil the standard treatment recommended by Brazilian Ministry Health are chloroquine 25 mg/kg for three days (10 mg/kg on day 1 and 7.5 mg/kg on days 2 and 3), plus primaquine 0.50 mg/kg for 7 days. In French Guiana, when the diagnosis is confirmed, the patient has already received an unsupervised three-day treatment with chloroquine (25 mg/Kg). To receive a primaquine prescription, the individual requires a G6PD-deficiency test performed in Cayenne and a nominative temporary use authorization from the State drug authority. Thus, this procedure causes delay in the administration of primaquine. Consequently, approximately half of the population was presented P. vivax relapses1414. Nacher M, Stefani A, Basurko C, Lemonnier D, Djossou F, Demar M, et al. The burden of Plasmodium vivax relapses in an Amerindian village in French Guiana. Malar J . 2013;12:367.. Moreover, the periodicity of vivax malaria relapses may be explained by the activation of latent hypnozoites by subsequent infections with P. falciparum. Evidence from a simultaneous typhoid and malaria epidemic suggest that typhoid fever might activate P. vivax hypnozoites2222. Shanks GD, White NJ. The activation of vivax malaria hypnozoites by infectious diseases. Lancet Infect Dis. 2013;13:900-6.,2323. White NJ. Determinants of relapse periodicity in Plasmodium vivax malaria. Malar J . 2011;10:297.. However, previous reports suggest that in mixed infections, one Plasmodium species may suppress the blood-stage density of another species2424. Lorenzetti A, Fornazari PA, Bonini-Domingos AC, Penhalbel RS, Fugikaha E, Bonini-Domingos CR, et al. Mixed Plasmodium falciparum infections and its clinical implications in four areas of the Brazilian Amazon region. Acta Trop . 2008;107:8-12.,2525. McKenzie FE, Bossert WH. Multi-species Plasmodium infections of humans. J Parasitol. 1999;85:12-8.. Thus, species interactions may cause alterations in the typical clinical manifestation of infections caused by only one species, therefore additional investigations are necessary in order to clarify the role of mixed-infections on disease severity and also in the parasite transmission dynamics, considering that events taking place in French Guiana can influence the transmission and spread of the parasite on the Brazilian side.
In conclusion, a large proportion of all malaria cases in South America occur in the Brazilian Amazon region. The emergence of epidemic and endemic foci is affected by colonization of different areas by different human groups. However, it remains unclear whether parasites are commonly spread from one area to another by migrants or whether they emerge mainly from local endemic populations2626. Machado RL, Póvoa MM, Calvosa VS, Ferreira MU, Rossit AR, dos Santos EJ, et al. Genetic structure of Plasmodium falciparum populations in the Brazilian Amazon region. J Infect Dis . 2004;190:1547-55.. The same CS genotypes circulating in French Guiana are found in the municipality of Oiapoque in Brazil. These findings suggest that it may occur via the dispersion of parasite populations between the two countries. Most likely, this distribution is associated with prolonged and/or more complex transmission patterns of these genotypes in Brazil, bordering French Guiana. Furthermore, studies based in CSP should consider these effects to obtain a protective vaccine.
ACKNOWLEDGMENTS
We thank all the subjects who participated in the study, as well as the health teams and coordination group from the Oiapoque District. We thank the following people for assistance in obtaining samples: Manoel do Carmo Barbosa da Cruz and to Valmir Corrêa and Corrêa. We thank Valéria Fraga and Luciana Moran for their technical support.
REFERENCES
-
1World Health Organization. World malaria report 2011. [cited 2012 July 24]. Available from: Available from: http://www.who.int/malaria/world_malaria_report_2011/en
» http://www.who.int/malaria/world_malaria_report_2011/en -
2González-Cerón L, Rodriguez MH, Nettel JC, Villarreal C, Kain KC, Hernandez JE. Differential susceptibilities of Anopheles albimanus and Anopheles pseudopunctipennis to infections with coindigenous Plasmodium vivax variants VK210 and VK247 in southern Mexico. Infect Immun. 1999;67:410-2.
-
3Stefani A, Dusfour I, Corrêa AP, Cruz MC, Dessay N, Galardo AK, et al. Land cover, land use and malaria in the Amazon: a systematic literature review of studies using remotely sensed data. Malar J. 2013;12:192.
-
4Brasil. Ministério da Saúde. Sistema de Informação de Vigilância Epidemiológica: notificação de casos. Malária. [cited 2013 Dec]. Available from: Available from: http://portalweb04.saude.gov.br/sivep_malaria/default.asp
» http://portalweb04.saude.gov.br/sivep_malaria/default.asp -
5Herrera S, Bonelo A, Perlaza BL, Fernández OL, Victoria L, Lenis AM, et al. Safety and elicitation of humoral and cellular responses in colombian malaria-naive volunteers by a Plasmodium vivax circumsporozoite protein-derived synthetic vaccine. Am J Trop Med Hyg. 2005;73 (Suppl 5):3-9.
-
6Coppi A, Natarajan R, Pradeln G, Bennett BL, James ER, Roggero MA, et al. The malaria circumsporozoite protein has two functional domains, each with distinct roles as sporozoites journey from mosquito to mammalian host. J Exp Med. 2011;208:341-56.
-
7Suphavilai C., Looareesuwan S., Good MF. Analysis of circunsporozoite protein-specific immune responses following recent infection with Plasmodium vivax Am J Trop Med Hyg . 2004;71:29-39.
-
8Arruda ME, Zimmerman RH, Souza RM, Oliveira-Ferreira J. Prevalence and level of antibodies to the circumsporozoite protein of human malaria parasites in five states of the Amazon region of Brazil. Mem Inst Oswaldo Cruz. 2007;102:367-71.
-
9Bonilla JA, Validum L, Cummings R, Palmer CJ. Genetic diversity of Plasmodium vivax PvCSP and PvMSP1 in Guyana, South America. Am J Trop Med Hyg . 2006;75:830-5.
-
10Machado RL, Póvoa MM. Distribution of Plasmodium vivax variants (VK210, VK247 and P. vivax-like) in three endemic areas of the Amazon region of Brazil and their correlation with chloroquine treatment. Trans R Soc Trop Med Hyg. 2000;94:377-81.
-
11Storti-Melo LM, de Souza-Neiras WC, Cassiano GC, Joazeiro AC, Fontes CJ, Bonini-Domingos CR, et al. Plasmodium vivax circumsporozoite variants and Duffy blood group genotypes in the Brazilian Amazon region. Trans R Soc Trop Med Hyg . 2009;103:672-8.
-
12da Silva AN, Santos CC, Lacerda RN, Machado RL, Póvoa MM. Susceptibility of Anopheles aquasalis and An. darlingi to Plasmodium vivax VK210 and VK247. Mem Inst Oswaldo Cruz . 2006;101:547-50.
-
13Peiter PC, Machado LO, Iñiguez Rojas L. Doenças transmissíveis na faixa de fronteira Amazônica: o caso da malária. In: Miranda AC, Barcellos C, Moreira JC, Monken M, organizadores. Território, ambiente e saúde. Rio de Janeiro: Editora Fiocruz; 2008. p.257-72.
-
14Nacher M, Stefani A, Basurko C, Lemonnier D, Djossou F, Demar M, et al. The burden of Plasmodium vivax relapses in an Amerindian village in French Guiana. Malar J . 2013;12:367.
-
15Volney B, Pouliquen JF, De Thoisy B, Fandeur T. A sero-epidemiological study of malaria in human and monkey populations in French Guiana. Acta Trop. 2002;82:11-23.
-
16Parobek CM, Bailey JA, Hathaway NJ, Socheat D, Rogers WO, Juliano JJ. Differing patterns of selection and geospatial genetic diversity within two leading Plasmodium vivax candidate vaccine antigens. PLoS Negl Trop Dis. 2014;8:e2796.
-
17Qari SH, Shi YP, Póvoa MM, Alpers MP, Deloron P, Murphy GS, et al. Global occurrence of Plasmodium vivax-like human malaria parasite. J Infect Dis. 1993;168:1485-9.
-
18Gomes MS, Vieira JL, Machado RL, Nacher M, Stefani A, Musset L, et al. Efficacy in the treatment of malaria by Plasmodium vivax in Oiapoque, Brazil, on the border with French Guiana: the importance of control over external factors. Malar J . 2015;14:402.
-
19Cassiano GC, Storti-Melo LM, Póvoa MM, Galardo AK, Rossit AR, Machado RL. Development of PCR-RFLP assay for the discrimination of Plasmodium species and variants of P. vivax (VK210, VK247 and P. vivax-like) in Anopheles mosquitoes. Acta Trop . 2011;118:118-22.
-
20Souza-Neiras WC, Storti-Melo LM, Cassiano GC, Couto VS, Couto AA, Soares IS, et al. Plasmodium vivax circumsporozoite genotypes: a limited variation or new subspecies with major biological consequences? Malar J . 2010;9:178.
-
21González-Cerón L, Martinez-Barnetche J, Montero-Solís C, Santillán F, Soto AM, Rodríguez MH, et al. Molecular epidemiology of Plasmodium vivax in Latin America: polymorphism and evolutionary relationships of the circumsporozoite gene. Malar J . 2013;12:243.
-
22Shanks GD, White NJ. The activation of vivax malaria hypnozoites by infectious diseases. Lancet Infect Dis. 2013;13:900-6.
-
23White NJ. Determinants of relapse periodicity in Plasmodium vivax malaria. Malar J . 2011;10:297.
-
24Lorenzetti A, Fornazari PA, Bonini-Domingos AC, Penhalbel RS, Fugikaha E, Bonini-Domingos CR, et al. Mixed Plasmodium falciparum infections and its clinical implications in four areas of the Brazilian Amazon region. Acta Trop . 2008;107:8-12.
-
25McKenzie FE, Bossert WH. Multi-species Plasmodium infections of humans. J Parasitol. 1999;85:12-8.
-
26Machado RL, Póvoa MM, Calvosa VS, Ferreira MU, Rossit AR, dos Santos EJ, et al. Genetic structure of Plasmodium falciparum populations in the Brazilian Amazon region. J Infect Dis . 2004;190:1547-55.
-
FINANCIAL SUPPORT The work reported in this manuscript was funded by the National Scientific and Technological Development in the Research Program for Health System Unique (CNPq PPSUS Public Notice No. 00077/2008/MCT/CNPq/SETEC, Process No. 35.000.162/2009) , and Government of the State of Amapá, and also, Central Public Health Laboratory (LACEN) of Amapá.
Publication Dates
-
Publication in this collection
2016
History
-
Received
27 Mar 2015 -
Accepted
21 Mar 2016