Abstracts
Despite a natural history interest in the early 1900s, relatively little ecological research has been carried out in the Rio Purús basin of central Amazonia, Brazil. Here we describe a new study area in the region of Lago Uauaçú with an emphasis on the climate, forest structure and composition, and soil characteristics between adjacent unflooded (terra firme) and seasonally inundated forests; situated within both the white-water (várzea) and black-water (igapó) drainage systems that dominate the landscape. The climate was found to be typical of that of the central Amazon. Várzea forest soils had high concentrations of nutrients, while terra firme and igapó soils were comparatively nutrient-poor. Terra firme forests were the most floristically diverse forest type, whereas várzea was intermediate, and igapó the most species-poor. The Lecythidaceae was the most important family in terra firme while the Euphorbiaceae was the most important in both várzea and igapó. There were significant differences between forest types in terms of number of saplings, canopy cover and understorey density. In contrasting our results with other published information, we conclude that the Lago Uauaçú region consists of a typical central Amazonian forest macro-mosaic, but is a unique area with high conservation value due to the intimate juxtaposition of terra firme, várzea and igapó forests.
Amazonia; floodplain forest; floristic composition; forest structure; soil nutrients
Apesar de um interesse na história natural no início de 1900, relativamente pouca pesquisa ecológica foi realizada na região do Rio Purús, Amazônia central, Brasil. Nesse estudo nós descrevemos uma nova área de estudo na região do Lago Uauaçú com ênfase no clima, na estrutura e composição da floresta, e nas características do solo entre florestas adjacentes de terra firme e inundadas (várzea e igapó) que dominam a paisagem. O clima foi caracterizado como típico daquele da Amazônia central. Os solos da floresta de várzea tiveram concentrações elevadas de nutrientes, enquanto os solos de florestas de terra firme e de igapó foram comparativamente pobres em nutrientes. As florestas de terra firme foram floristicamente mais diversas, enquanto várzea foi intermediária, e igapó foi a mais pobre em espécies. Lecythidaceae foi a família mais importante na terra firme enquanto Euphorbiaceae foi mais importante em ambos, várzea e igapó. Foram encontradas diferenças significativas entre os tipos de floresta e o número de subadultos, cobertura de dossel e densidade do subdossel. Contrastando nossos resultados com outras informações publicadas, nós concluímos que a região do Lago Uauaçú consiste em um macro-mosaico típico de floresta da Amazônia central, mas uma área única, com elevado valor de conservação devido à íntima justaposição de florestas de terra firme, várzea e igapó.
Amazônia; floresta inundada; composição florística; estrutura da floresta; nutrientes do solo
BOTÂNICA
Floristic, edaphic and structural characteristics of flooded and unflooded forests in the lower Rio Purús region of central Amazonia, Brazil
Características florísticas, edáficas e estruturais das florestas inundadas e de terra firme na região do baixo Rio Purús, Amazônia central, Brasil
Torbjørn Haugaasen1 1 Corresponding author ; Carlos Augusto Peres
Centre for Ecology, Evolution and Conservation, School of Environmental Sciences, University of East Anglia, Norwich NR4 7TJ, UK, Phone: (+44) 1603 591299, Fax: (+44) 1603 591327, EMail address: T.Haugaasen@uea.ac.uk; C.Peres@uea.ac.uk
ABSTRACT
Despite a natural history interest in the early 1900s, relatively little ecological research has been carried out in the Rio Purús basin of central Amazonia, Brazil. Here we describe a new study area in the region of Lago Uauaçú with an emphasis on the climate, forest structure and composition, and soil characteristics between adjacent unflooded (terra firme) and seasonally inundated forests; situated within both the white-water (várzea) and black-water (igapó) drainage systems that dominate the landscape. The climate was found to be typical of that of the central Amazon. Várzea forest soils had high concentrations of nutrients, while terra firme and igapó soils were comparatively nutrient-poor. Terra firme forests were the most floristically diverse forest type, whereas várzea was intermediate, and igapó the most species-poor. The Lecythidaceae was the most important family in terra firme while the Euphorbiaceae was the most important in both várzea and igapó. There were significant differences between forest types in terms of number of saplings, canopy cover and understorey density. In contrasting our results with other published information, we conclude that the Lago Uauaçú region consists of a typical central Amazonian forest macro-mosaic, but is a unique area with high conservation value due to the intimate juxtaposition of terra firme, várzea and igapó forests.
KEY WORDS: Amazonia, floodplain forest, floristic composition, forest structure, soil nutrients
RESUMO
Apesar de um interesse na história natural no início de 1900, relativamente pouca pesquisa ecológica foi realizada na região do Rio Purús, Amazônia central, Brasil. Nesse estudo nós descrevemos uma nova área de estudo na região do Lago Uauaçú com ênfase no clima, na estrutura e composição da floresta, e nas características do solo entre florestas adjacentes de terra firme e inundadas (várzea e igapó) que dominam a paisagem. O clima foi caracterizado como típico daquele da Amazônia central. Os solos da floresta de várzea tiveram concentrações elevadas de nutrientes, enquanto os solos de florestas de terra firme e de igapó foram comparativamente pobres em nutrientes. As florestas de terra firme foram floristicamente mais diversas, enquanto várzea foi intermediária, e igapó foi a mais pobre em espécies. Lecythidaceae foi a família mais importante na terra firme enquanto Euphorbiaceae foi mais importante em ambos, várzea e igapó. Foram encontradas diferenças significativas entre os tipos de floresta e o número de subadultos, cobertura de dossel e densidade do subdossel. Contrastando nossos resultados com outras informações publicadas, nós concluímos que a região do Lago Uauaçú consiste em um macro-mosaico típico de floresta da Amazônia central, mas uma área única, com elevado valor de conservação devido à íntima justaposição de florestas de terra firme, várzea e igapó.
PALAVRAS CHAVE: Amazônia, floresta inundada, composição florística, estrutura da floresta, nutrientes do solo
INTRODUCTION
Rio Purús is a white-water tributary joining the Rio Solimões from the south, with its headwaters arising in the pre-Andean slopes of Peru. The Purús catchment covers approximately 375,000 km2, and is the sixth largest tributary drainage area in the Amazon basin (Goulding et al. 2003). Due to its meandering nature, the river is extremely long (approximately 3,200 km) and sustains approximately 40,000 km2 of floodplain forest. This is more than any other tributary of the Amazon River (Goulding et al. 2003). Erosion and sedimentation processes resulting from the kinetic energy of water discharge serve to create numerous channels, lakes and forest levees. The combination of lakes, unflooded and flooded forests and floating-meadows makes this a highly productive and diverse region. For example, the Rio Purús is the most important supplier of fish landings for the Manaus population (Bayley and Petrere, 1989) which is over 1.5 million strong. However, apart from William Chandless' seminal geographical expedition in the 1860s (Chandless, 1866) and Huber's pioneering naturalistic exploration and description of the tree communities in the Purús river region (Huber, 1906) little biological and conservation research has been carried out since (cf. Goulding et al. 2003). The purpose of this paper is to introduce an entirely new study site in the lower Purús region of central Brazilian Amazonia and describe and contrast the forest structure, floristic and soil characteristics of the unflooded and flooded forest landscape which dominates the area.
Study area
This study was conducted from August 2000 to November 2003 at Lago Uauaçú which is located in the lower Rio Purús region (04º20' S, 62º28' W) of central-western Brazilian Amazonia, about 350 km southwest of Manaus (Figure 1). Lago Uauaçú recently became the north-western boundary of the newly decreed Piagaçú-Purús Sustainable Development Reserve (see below) and parts of the study area lie within the reserve boundary (de Deus et al. 2003). Despite its proximity to the Rio Purús/Solimões juncture, this region remains relatively undisturbed and incorporates a large interdigitated mosaic of unflooded (terra firme) and flooded forests inundated by white-water (várzea) and black-water (igapó) on a seasonal basis for as long as 6 months of the year.
Lago Uauaçú itself is a pristine, 32 km long crescent-shaped black-water lake fed entirely by rainfall collected in an internal catchment consisting primarily of terra firme forest, with seasonally flooded igapó forest occurring along the lake margins and along the banks of perennial streams. The study area also incorporates a large confluence of floodplain forests under the influence of both the Rio Solimões and Rio Purús (Figure 1). The hydrology of the landscape defines a geochemical mosaic across the study area, and is the primary mechanism to which this large-scale natural forest mosaic can be attributed. The area around Lago Uauaçú contains high levels of plant and animal species richness reflecting its unique geographical context where várzea, igapó and terra firme forests come together. For example, due to the matrix of different adjacent habitat types the area harbours at least 12 sympatric primate species (Haugaasen and Peres, 2005a, b).
The study area is inhabited by 30 extractive caboclo (non-tribal) households whose village lies next to the lake entrance to the east. The villagers rely primarily on the collection of Brazil nuts from natural stands of Brazil nut trees (Bertholletia excelsa Humboldt & Bonpland Lecythidaceae) in addition to small-scale fishing and hunting for both subsistence and income. However, subsistence hunters in the study area are highly selective and are not targeting primates. Land cultivation in the study area is restricted to small slash-and-burn patches along the lake edge.
Population pressure from surrounding areas is low as the study area is 12 hours by motorised canoe from Codajás, the nearest town (Figure 1), and at least a 3 day boat-ride from Manaus depending on season. During the dry season, fluvial passage to Codajás along the Rio Solimões is completely cut off, restricting access to the lake from the Rio Purús only. Commercial fishing vessels and sport fishing visitors were regularly encountered at the lake throughout the study period. Sport hunting does not take place anywhere in the region, although commercial hunting pressure by outsiders has become an increasing problem. Another key resource in the area is the açaí palm, Euterpe precatoria, the fruits of which are processed into a drink and ice-cream. The height of the Euterpe fruiting season sees a great influx of outsiders to the area, especially from Codajás.
The Piagaçú-Purús Sustainable Development Reserve
Parts of our study area and Lago Uauaçú were recently included in the Piagaçú-Purús Sustainable Development Reserve (de Deus et al. 2003). This newly decreed (September 2003) reserve covers 1,008,167 ha and lies within the lower Rio Purús region (4º05' 5º35' S and 61º73' 63º35' W). The most obvious borders of the reserve are the Rio Purús in the east and the Abufari Biological Reserve in the south. The reserve incorporates parts of four municipal counties which together have a population of c. 114,000 people, 52,000 of which live in rural dwellings (de Deus et al. 2003).
The reserve was established to control fisheries due to the importance of the region to the Manaus fish market, but also due to its relatively pristine status, encompassing key breeding sites for endangered species such as Podocnemis turtles and manatees. Sustainable development reserves are a new model of conservation areas pioneered by the State of Amazonas in Brazil, and they focus on combining sustainable use of natural resources with a relatively intact forest cover. This model of reserve design has been built on the apparent success of the Mamirauá and Amanã reserves.
Habitat types
Upland terra firme forests are those that are above the maximum flood level of Amazonian rivers and perennial streams. Consequently, these unflooded forests lie on well drained terrain that tends to be heavily leached and nutrient-poor because they have long been deprived of alluvial sediments. Terra firme forest represents the main forest type across the Amazonian forest landscape, accounting for ~95% of the Amazon basin. However, a recent study suggests that this total should be revised to 83% (Hess et al. 2003). Several distinct plant communities can be found embedded within the closed-canopy forest, as indicated by analysis of satellite images (Tuomisto and Ruokolainen, 1994; Tuomisto et al. 1995) and floristic compositional variation (Terborgh and Andresen, 1998; ter Steege et al. 2000). These results are consistent with further studies suggesting that the high levels of diversity in terra firme forest is a consequence of high habitat heterogeneity derived from different edaphic and ecological conditions (Terborgh 1985; Salo et al. 1986; Tuomisto et al. 1995), itself a function of a range of historical, geographical and ecological factors (Liu and Colinvaux, 1985; Räsänen et al. 1987; Ayres and Clutton-Brock, 1992; Peres et al. 1996; Voss and Emmons, 1996; Peres and Janson, 1999). In the present study, however, we consider terra firme forests as a single forest type regardless of the different microhabitats it may include.
Seasonally inundated forests comprise the second major vegetation type in the Amazon. The low-lying topography of the basin and seasonality of rainfall inundate these floodplains for up to six months of the year, and the annual water level fluctuation of the Amazon river and its tributaries can reach 14 m in amplitude (Ferreira, 1997). Although seasonally flooded forests are inundated on a regular basis each year, different flooded forest types can be distinguished on the basis of both hydrochemical (Sioli, 1968) and floristic differences (Prance, 1979).
Floodplain forests along white-water rivers are known as várzea forests and account for the most extensive type of flooded forest in South America, covering approximately 180,000 km2 of the Amazon basin (Bayley and Petrere, 1989). White-water rivers originate in the geologically young Andes or the pre-Andean regions which are defined by easily erodable landscapes (Räsänen et al. 1987), contributing large amounts of nutrient-rich suspended sediments to the rivers. Due to the seasonal influx of nutrients, várzea forests are eutrophic and remain exceptionally productive (Junk and Piedade, 1993). However, despite this elevated productivity, several studies have shown that the floristic and faunal diversity of várzea forest is consistently lower than those of terra firme forest (e.g. Balslev et al. 1987; Peres, 1997; Patton et al. 2000; Haugaasen and Peres, 2005a, b).
Igapó forests are inundated on a seasonal basis by black- or clear-water that originate in the Amazonian tertiary lowlands and often drain sandy soils with a characteristically low nutrient content. Although less extensive, igapó forests comprise a widespread forest type in Amazonia and may cover an area similar to that of várzea (Kricher, 1997). In this study, igapó forests are defined as those inundated by black-water. Igapó drainages are more nutrient poor than várzea, carry less suspended inorganic elements and contain elevated concentrations of dissolved organic material such as humic and fulvic acids. Igapó forests are therefore oligotrophic and their plant communities consequently tend to support lower levels of species diversity and animal biomass than both terra firme and várzea (Ferreira, 1997).
METHODS
Data collection
Climate Water level fluctuations were recorded using a 5 m long pole divided into 5 cm sections which was penetrated vertically into the lake bed near the floating research station. Precipitation was recorded using a rain gauge attached to the station and all readings were carried out on a daily or weekly basis.
Forest structure Canopy openness was quantified using a convex spherical densitometer every 50 m, with four readings taken per point (see Lemmon, 1957), along 19,300 m of trails in terra firme, 23,600 m in várzea and 5,000 m in igapó forest. The openness of the understorey was measured using a 2.5 m graduated pole of 4 cm in diameter held vertically 15 m to the side of the transect, which was examined with a pair of binoculars by an observer standing on the transect. Understorey density readings were recorded every 50 m along 14,400 m in terra firme, 14,500 m in várzea and 5,000 m in igapó and correspond to the number of 10-cm pole sections (range: 0 - 25) that were clearly visible. All saplings taller than 30 cm were measured within 50m2 (1 x 50 m) plots, and were categorised into ten different size classes of 1 cm (up to 10 cm dbh). A total of 6 plots in terra firme, 6 in várzea and 3 in igapó forest were sampled. Forest basal area was calculated as:

where D = diameter at breast height.
Forest biomass estimates were calculated using an allometric model for aboveground dry biomass (AGDB):
AGDB = (exp 3.323 + (2.546 x (ln (D/100)))) x 600
where D = diameter at breast height, and the parameters of the model were derived from 319 destructively sampled trees ranging from 5 to 120 cm dbh in an Amazonian terra firme forest located north of Manaus (Santos, 1996).
Floristics A quantitative ecological inventory of all trees and lianas > 10 cm dbh (diameter at breast height ~1.3 m above ground) was carried out across nine hectares, comprising three 1-ha plots in each forest type. The plot shape was 10 x 1000 m in terra firme and várzea and 20 x 500 m in igapó forest because of the spatial configuration and extent of the latter forest type. Identifications were carried out in the field by a highly experienced herbarium technician from Instituto Nacional de Pesquisas da Amazônia, Manaus (INPA). Due to the use of morphospecies in the identification process, the number of species reported here should be interpreted as conservative estimates of species richness. Peripheral trees straddling the plot boundary were only included in the inventory if the mid-point of their trunks fell within the plot. The dbh of buttressed trees were measured immediately above the buttress. When direct measurement with a dbh-tape was impossible (e.g. sulcate trunks, stranglers or buttresses were too high), the diameter was estimated to the nearest 5 cm. Mean tree density, forest basal area and AGDB reported here are based on results from these plots. We calculated the Family Importance Value (FIV) for each family, which is the sum of the relative density, dominance and diversity of any tree family (Mori et al. 1983).
Soil sampling and analyses Soil samples were collected every 50 m on a single randomly chosen transect in terra firme, várzea and igapó forest. Each sample was collected >1 m from the side of the path. Samples were extracted from two depth classes: 0-20 cm and 20-40 cm. In total, 25 samples were collected from each depth class in each forest type, and used to form 5 well-mixed compositional 500 g samples used in the laboratory analyses. All analyses were carried out at the INPA soil laboratory and follow the guidelines provided by EMBRAPA (1999).
Data analyses The difference in mean tree densities between the three forest types was examined using x2 test. Differences in mean forest basal area, AGDB and tree dbh were tested using Kruskall-Wallis tests. One-way ANOVAs with Tukey's post-hoc test were used to test for differences in other habitat data between the three forest types. Two-way ANOVAs with Tukey's post-hoc tests were used to test for differences in soil granulometry and nutrient concentrations between different forest types and soil depth classes. Rarefaction curves were produced using the computer program EcoSim (Gotelli and Entsminger, 2003), on the basis of 1000 random iterations. These curves used data pooled from all plots in the same forest type, and calculated the expected number of species added per 100 individual trees sampled.
RESULTS
Climate
At Lago Uauaçú, the total annual precipitation for February 2002 January 2003 was 2664.8 mm. The monthly rainfall showed a pronounced seasonal variation, with July through October being the driest months, and February through June the wettest (Figure 2). These trends were similar in the following year (Figure 2). The wettest and driest months during the period of climatic measurements (Feb. 2002 Sep. 2003) were March 2002 with 555 mm, and July 2003 with 27 mm respectively (Figure 2).
The seasonal variation in precipitation produced marked fluctuations in the lake water level and surrounding rivers and streams. In 2002, Lago Uauaçú experienced a peak in water level in late June. By early November, the receding water had reached its lowest level, resulting in a water fluctuation exceeding 10.5 m in this particular year. The water level rose more gradually than it receded (Figure 2), and during AugustOctober 2002, the lake water level could recede by as much as 1 m or more per week.
Soil analyses
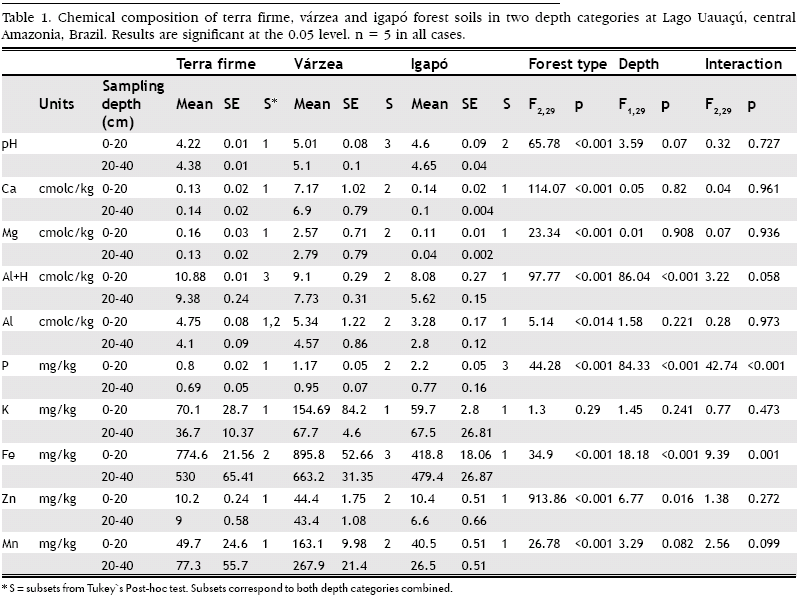
Two-way ANOVAs on soil granulometry and chemical composition showed that there was a significant difference between forest types in all cases except for potassium (K) (F2,29 = 1.3, p = 0.29). Depth was an important factor for coarse and fine sand, Al+H, P, Fe and Zn (Coarse sand, F2,29 = 52.55, p = <0.001; Fine sand, F2,29 = 15.01, p = 0.001; Al+H, F2,29 = 86.04, p = <0.001; P, F2,29 = 84.33, p = <0.001; Fe, F2,29 = 18.18, p = <0.001). Only coarse sand, P and Fe had a significant interaction between forest type and depth (Coarse sand, F2,29 = 29.64, p = <0.001; P, F2,29 = 42.74, p = <0.001; Fe, F2,29 = 9.39, p = 0.001). Tukey's post-hoc tests showed significant differences in soil texture between forest types. However, there was no significant difference in the percentage of silt between terra firme and igapó, whereas there was a significant difference between forest types in all other comparisons (Figure 3). The soils were fairly acidic in all forest types, but less so in várzea (Table 1). Várzea contained the highest concentrations of 7 (Ca, Mg, Al, K, Fe, Zn and Mn) out of 9 elements analysed. Similar levels of Ca, Mg, Al, Zn and Mn were found in terra firme and igapó as shown by Tukey's post-hoc tests (Table 1). The highest phosphorus concentrations were found in igapó forest.
Forest structure
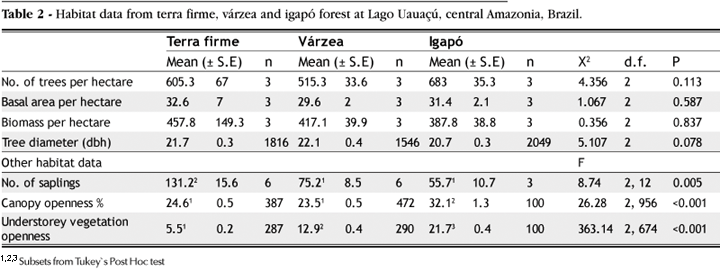
A total of 5,411 trees > 10 cm dbh occurred in the nine hectares sampled across all three forest types, yielding a mean density of 601.2 ± 34.0 trees ha-1 across the landscape. There was no significant difference between the three forest types in terms of mean number of trees, basal area and AGDB per hectare (Table 2). Similarly, the mean dbh of trees found in the three forest types did not differ significantly (Table 2).
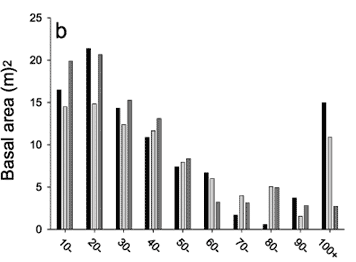
The overall abundance of trees was strongly size-dependent and the distributions of dbh size classes in the three forest types showed an inverse J-shaped curve for the tree assemblage as a whole (Figure 4a). Smaller trees between 10 and 30 cm dbh dominated the three forest types, accounting for 83.6%, 81.3%, 84.3% of all trees sampled in terra firme, várzea and igapó, respectively. Only 23 (0.43%) emergent trees in the entire sample reached diameters > 100 cm, including 11 in terra firme, nine in várzea and three in igapó (Table 3). Seven of these 11 emergents in terra firme were Brazil nut trees (Bertholletia excelsa, Lecythidaceae) (Table 3). Due to the large number of stems in the smaller dbh size classes, most of the contribution to the total tree basal area in all forest types is derived from these categories. However, due to their large sizes, trees > 100 cm dbh account for a large proportion of basal area despite their low numerical abundance (Figure 4b). This is even more pronounced when the total biomass is classed by tree dbh (Figure 4c). These trees make up 27.5%, 21.1% and 5.2% of the biomass in terra firme, várzea and igapó forest, respectively.

A total of 959 spherical densitometer readings were recorded in the three forest types. Tukey's post-hoc tests showed that there was no significant difference in canopy openness between terra firme and várzea forest, whereas the igapó forest canopy was significantly more open (Table 2).
A total of 677 understorey density readings were recorded in the three forest types. Significant differences were found between the forest types (p < 0.001), and no habitat belonged to the same subset as distinguished by Tukey's post-hoc comparisons (Table 2). The terra firme understorey was much denser than that of várzea forest, whereas the igapó understorey was the most open of the three forest types, despite a more open canopy. This largely reflects the number of established tree saplings found in each habitat as terra firme forest contained significantly greater densities of saplings than either várzea or igapó forest (p = 0.005; Table 2). Várzea and igapó were not significantly different from each other.
Floristic diversity
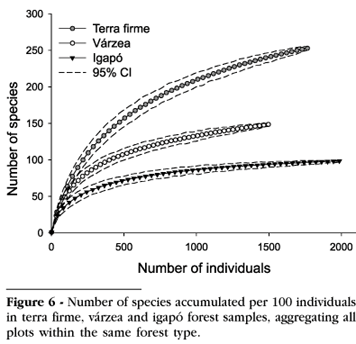
Tree assemblages in terra firme forest were the most species-rich, whereas those in várzea were intermediate with igapó being the most species-poor (Figure 5). The slope of the rarefaction curves in all forest types typically declined as sample sizes increased, but did not approach an asymptote (Figure 6). It is clear, however, that very few species were added to the igapó sample beyond 1,000 individuals inventoried, and after sampling three hectares (2,049 trees) the curve for this forest type appeared to approach an asymptote. New species were still being added to the terra firme and várzea samples after the survey of three hectares (Figure 6). These curves also clearly indicate that terra firme forest was the most diverse, whereas várzea forest had intermediate levels of tree species diversity. Igapó was the most species-poor of all forest types.
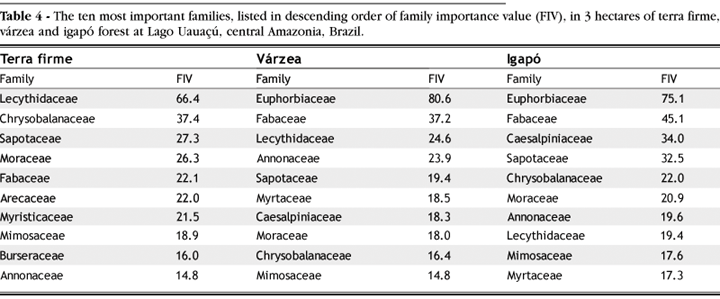
In terms of the family importance value (FIV), the Lecythidaceae was the most dominant tree family in terra firme forest, whereas the Euphorbiaceae was the most important family in várzea and igapó forests (Table 4). The dominance of Lecythidaceae in terra firme is a reflection of the large sizes attained by some species in this family, such as Bertholletia excelsa and Cariniana cf. micrantha (Table 3). Although size is also important in várzea, the dominance of Euphorbiaceae in várzea and igapó forest primarily reflects the large number of individuals and species belonging to this family.
DISCUSSION
Climate
The pattern of precipitation at Lago Uauaçú is similar to that of other parts of central Amazonia with 3-4 months of the year receiving less than 100 mm of rain and 3-4 months experiencing heavy rainfall. However, hydrological deficit in the plant community rarely takes place even in terra firme forest as the cumulative monthly rainfall fell below 50 mm only once in 20 months. The total annual precipitation is very similar to the annual mean recorded at Reserva Ducke, 30 km north of Manaus (2401 ± 328 mm; Junk and Krambeck, 2000). However, water levels along the Rio Solimões can rise up to 14 m in comparison to the 10.5 m water level fluctuation at Lago Uauaçú in 2002 (Ferreira, 1997). In 2002, the water level did not recede as much as in previous years and larger fluctuations could be expected in subsequent years.
Soils
It is a general agreement that várzea forest soils contain much higher concentrations of soil macronutrients important to plants (N, P, K, Na, Ca and Mg) than those of igapó and terra firme forest (Irion, 1978; Sombroek, 1984; Furch and Klinge, 1989; Furch, 1997, 2000). This conclusion is entirely consistent with findings reported in this study. The nutrient-rich alluvial sediments carried by white-water rivers resulting from the Andean outwash and deposited annually onto várzea forests are solely responsible for this sustained fertility. The transport of high sediment loads in white-water rivers also explains the high percentage of silt and clay in the várzea soil samples. In contrast, terra firme and igapó forest remain nutrient poor due to the lack of such seasonal influx. In a recent study, Furch (2000) also found some similarities between terra firme and igapó, but that terra firme had lower concentrations of most elements. The igapó and adjacent terra firme forest had elevated amounts of sand in soil samples due to the lack of sediment influxes. However, the high percentage of silt in the igapó sample suggests rare surface runoff of alluvial deposits across plateau areas which may have been historically determined by extreme events of white-water inundation. The likelihood of white-water inundation events remains a strong possibility due to the location of the study site, situated between the junctures of large tracts of várzea forests along the Rio Solimões and Rio Purús.
Forest structure
The inverse J-shape curve of the dbh distribution in the three forest types is typical of other Neotropical forest sites (Ayres, 1986; Ferreira and Prance, 1998a; Campbell, 1989; Amaral, 1996; Lima Filho et al. 2001). Similarly, the high proportion of trees <30 cm dbh is similar to observations made from elsewhere in the Amazon (Boom, 1986; Milliken 1998). The mean basal area of terra firme forest (32.6 m2) is almost identical to samples from elsewhere in central Amazonia (Peres, 1991; Milliken, 1998), and identical to one hectare of várzea forest sampled on the lower Japurá river (Ayres, 1986). The mean várzea forest basal area observed in this study was therefore also comparable to that of Ayres (1986) for the Mamirauá Reserve. Likewise, basal area estimates obtained in igapó forest at Lago Uauaçú were comparable to other floristic inventories in black-water floodplains in the Rio Negro basin (Jaú National Park: Ferreira, 1997).
The canopy was more open in igapó than in terra firme forest. Light conditions have been shown to have positive effect on forest regeneration (Haugaasen et al. 2003), indicating that light availability is not a limiting factor for tree regeneration in igapó. Canopy openness was similar between terra firme and várzea, whereas understorey density and sapling density was significantly lower in várzea, suggesting that light limitation is an unlikely cause of this contrast. The lack of support for light as a limiting factor suggests that an alternative mechanism is responsible for the open understorey and smaller number of saplings in both igapó and várzea. Indeed, flooding seems to be the most limiting factor influencing sapling species distribution and establishment (Klinge et al. 1995; Wittmann and Junk, 2003), which is strongly related to the height and the duration of the annual inundation event (Wittmann and Junk, 2003). This may be a result of physiological stress due to highly anoxic conditions as well as physical flood disturbance. The annual flooding event may impose temporary oxygen stress on roots, and is likely to hamper photosynthesis, particularly in smaller plants (Parolin, 2001). Saplings and seedlings may therefore experience high levels of mortality during inundation events.
Floristics
The terra firme forest matrix at Lago Uauaçú had a much higher diversity of tree morphospecies than those of várzea and igapó. This result is in agreement with patterns found in other studies where both várzea and igapó forest exhibited relatively impoverished floras compared to that of terra firme (Ayres, 1986; Black et al. 1950; Campbell et al. 1986; Ferreira, 1997; Ferreira and Prance 1998b; Prance, 1979, 1987; ter Steege et al. 2000).
The floristic composition at the family level in the three forest types also resembles that found elsewhere in Amazonia. The Lecythidaceae is typically the most important family in central Amazonian terra firme forests (Peres, 1991; Amaral, 1996; Lima Filho et al. 2001; Prance et al. 1976) and Euphorbiaceae and Leguminosae sensu lato (Caesalpiniaceae, Fabaceae and Mimosaceae) are frequently found to be dominant in várzea and igapó (Ayres 1986, 1993; Keel and Prance, 1979; Ferreira, 1997). Additionally, inspection of rarefaction curves suggest that the total area of 3 ha sampled in each forest type was probably insufficient to estimate the true number of species for terra firme and várzea forest at our study site. However, the rate of species accumulation in the igapó sample was considerably lower than in the other forest types and appears to approach an asymptote suggesting that three hectares may be an adequate sample size for sampling tree species richness in igapó forest.
CONCLUSION
In conclusion, the Lago Uauaçú region portrays a natural forest mosaic typical of central Amazonia in terms of climatic and edaphic constraints, forest structure and floristic characteristics. However, the combination of the three adjacent forest types greatly contributes to the landscape heterogeneity and high conservation value of this area. Our results indicate that the Lago Uauaçú region represents a unique system in which to study landscape-scale processes and interactions which have been largely overlooked by ecological research and conservation planning in tropical forests.
ACKNOWLEDGEMENTS
This study was supported by small grants from the Wildlife Conservation Society, WWF-US and the Amazon Conservation Team (ACT). Logistical help was provided by the Amazon Association for the Preservation of Areas of High Biodiversity (AAP). Special thanks to Joanne M. Tuck, Marilene, Edivar, Zé, and Evineu for their help in the field, to Sr. José Lima for carrying out the tree identifications and to Dr Newton Falcão for performing the soil analysis. P. Judge prepared Figure 1.
LITERATURE CITED
Recebido em 03/02/2005
Aceito em 02/01/2006
- Amaral, I.L. 1996. Diversidade florística em floresta terra firme, na região do Rio Urucu - AM Master's thesis, Instituto Nacional de Pesquisas da Amazônia, Universidade do Amazonas, Manaus. 104p.
- Ayres, J.M. 1986. Uakaris and Amazonian flooded forest PhD thesis, University of Cambridge, Cambridge. 338p.
- Ayres, J.M. 1993. As matas de várzea do Mamirauá Sociedade Civil Mamirauá, Belém, 96p.
- Ayres, J.M.; Clutton-Brock, T.H. 1992. River boundaries and species range size in Amazonian primates. American Naturalist, 140(3):531-537.
- Balslev, H.; Luteyn, J.; Øllgard, B.; Holm-Nielsen, L.B. 1987. Composition and structure of adjacent unflooded and floodplain forest in Amazonian Ecuador. Opera Botanica, 92:37-57.
- Bayley, P.N.; Petrere, M. 1989. Amazon fisheries: assessment methods, current status and management options. Special publications of the Canadian Journal of Fisheries and Aquatic Sciences, 106:385-398.
- Black, G.A.; Dobzhansky, T.H.; Pavan, C. 1950. Some attempts to estimate species diversity and population density of trees in Amazonian forests. Botanical Gazette, 111:413-425.
- Boom, B.M. 1986. A forest inventory in Amazonian Bolivia. Biotropica, 18(4):287-294.
- Campbell, D.G. 1989. Brazilian Amazon. In: Campbell, D.G.; Hammond, H.D. (Eds). Floristic Inventory of Tropical Countries New York Botanical Garden, New York. p. 523-533.
- Campbell, D.G.; Daly, D.C.; Prance, G.T.; Maciel, U.N. 1986. Quantitative ecological inventory of terra-firme and várzea tropical forest on the Rio-Xingú, Brazilian Amazon. Brittonia, 38(4):369-393.
- Chandless, W. 1866. Ascent of the River Purús. Journal of the Royal Geographical Society. 36:86-118.
- de Deus, C.P.; da Silveira, R.; Py-Daniel, L.H.R. 2003. Piagacu-Purús: Bases científicas para criação de uma reserva de desenvolvimento sustentável Instituto de Desenvolvimento Sustentável Mamirauá, Manaus. 83p.
- EMBRAPA. 1999. Manual de análises químicas de solos, plantas e fertilizantes EMBRAPA, Brasília. 370p.
- Ferreira, L.V. 1997. Effects of the duration of flooding on species richness and floristic composition in three hectares in the Jau National Park in floodplain forests in central Amazonia. Biodiversity and Conservation, 6(10):1353-1363.
- Ferreira, L.V.; Prance, G.T. 1998a. Species richness and floristic composition in four hectares in the Jaú National Park in upland forests in Central Amazonia. Biodiversity and Conservation, 7(10):1349-1364.
- Ferreira, L.V.; Prance, G.T. 1998b. Structure and species richness of low-diversity floodplain forest on the Rio Tapajós, Eastern Amazonia, Brazil. Biodiversity and Conservation, 7(5):585-596.
- Furch, K. 1997. Chemistry of várzea and igapó soils and nutrient inventory of their floodplain forests. In: Junk, W.J. (Ed). The Central Amazon Floodplain: Ecology of a pulsing system Springer-Verlag, Berlin. p. 47-67.
- Furch, K. 2000. Chemistry and bioelement inventory of contrasting Amazonian forest soils. In: Junk, W.J.; Ohly, J.J.; Piedade, M.T.F.; Soares, M.G.M. (Eds). The Central Amazonian Floodplain: Actual use and options for a sustainable management Backhuys Publishers, Leiden. p. 109-128.
- Furch, K.; Klinge, H. 1989. Chemical relationship between vegetation, soil and water in contrasting inundation areas of Amazonia. In: Proctor, J. (Ed). Mineral nutrients in tropical forest and savanna ecosystems Blackwell, Oxford. p. 189-204.
- Gotelli, N.J.; Entsminger, G.L. 2003. EcoSim: Null models software for ecology Version 7. Burlington, VT 05465: Acquired Intelligence Inc. & Kesey-Bear.
- Goulding, M.; Barthem, R.; Ferreira, E.J.G. 2003. The Smithsonian Atlas of the Amazon Smithsonian Institution Press, Washington D.C. 256p.
- Haugaasen, T.; Barlow, J.; Peres, C.A. 2003. Surface wildfires in central Amazonia: Short-term impact on forest structure and carbon loss. Forest Ecology and Management, 179(1-3):321-331.
- Haugaasen, T.; Peres, C.A. 2005a. Mammal assemblage structure in Amazonian flooded and unflooded forests. Journal of Tropical Ecology 21(2):133-145.
- Haugaasen, T.; Peres, C.A. 2005b. Primate assemblage structure in Amazonian flooded and unflooded forests. American Journal of Primatology 67:243-258.
- Hess, L.L.; Melack, J.M.; Novo, E.; Barbosa, C.C.F.; Gastil, M. 2003. Dual-season mapping of wetland inundation and vegetation for the central Amazon basin. Remote Sensing of Environment 87:404-428.
- Huber, J. 1906. La vegetation de la valee Rio Purus. Bulletin de L'Herbier Boissier, 4:249-289.
- Irion, G. 1978. Soil infertility in the Amazonian rain forest. Naturwissenschaften, 65:515-519.
- Junk, W.J.; Krambeck, H.-J. 2000. Climate and hydrology. In: Junk, W.J.; Ohly, J.J.; Piedade, M.T.F.; Soares, M.G.M. (Eds). The Central Amazonian Floodplain: Actual use and options for a sustainable management Backhuys Publishers, Leiden. p. 95-108.
- Junk, W.J.; Piedade, M.T.F. 1993. Herbaceous plants of the flood plain near Manaus: species diversity and adaptations to the flood pulse. Amazoniana, 12:467-484.
- Keel, S.H.; Prance, G.T. 1979. Studies of the vegetation of a white-sand black-water igapó (Rio Negro, Brazil). Acta Amazonica, 9:645-655.
- Klinge, H.; Adis, J.; Worbes, M. 1995. The vegetation of a seasonal várzea in the lower Solimões river, Brazilian Amazon. Acta Amazonica, 25:201-220.
- Kricher, J. 1997. A Neotropical companion: an introduction to the animals, plants and ecosystems of the New World tropics Princeton University Press, Princeton. 504p.
- Lemmon, P.E. 1957. A new instrument for measuring forest overstory density. Journal of Forestry, 55(9):667-668.
- Lima Filho, D.A.; Matos, F.D.A.; Amaral, L.; Revilla, J.D.; Coelho, L.S.; Ramos, J.F.; Santos, J.L. 2001. Inventário florístico de floresta ombrófila densa de terra firme, na região do Rio Urucu - Amazonas, Brasil. Acta Amazonica, 31(4):565-579.
- Liu, K.B.; Colinvaux, P.A. 1985. Forest changes in the Amazon Basin during the last glacial maximum. Nature, 318(6046):556-557.
- Milliken, W. 1998. Structure and composition of one hectare of central Amazonian terra firme forest. Biotropica, 30(4):530-537.
- Mori, S.A.; Boom, B.M.; Carvalho, A.M.; Santos, T.S. 1983. Ecological importance of Myrtaceae in Eastern Brazilian wet forest. Biotropica, 15:68-70.
- Parolin, P. 2001. Morphological and physiological adjustments to waterlogging and drought in seedlings of Amazonian floodplain trees. Oecologia, 128(3):326-335.
- Patton, J.L.; Da Silva, M.N.F.; Malcolm, J.R. 2000. Mammals of the Rio Jurua and the evolutionary and ecological diversification of Amazonia. Bulletin of the American Museum of Natural History, 244:1-306.
- Peres, C.A. 1991. Ecology of mixed-species groups of tamarins in Amazonian terra firme forests PhD, Cambridge, Cambridge. 242p.
- Peres, C.A. 1997. Primate community structure at twenty western Amazonian flooded and unflooded forests. Journal of Tropical Ecology, 13:381-405.
- Peres, C.A.; Janson, C.H. 1999. Species coexistence, distribution and environmental determinants of neotropical primate richness: A community-level zoogeographic analysis. In: Fleagle, J.G.; Janson, C.H.; Reed, K.E. (Eds). Primate Communities Cambridge University Press, Cambridge. p. 55-74.
- Peres, C.A.; Patton, J.L.; daSilva, M.N.F. 1996. Riverine barriers and gene flow in Amazonian saddle-back tamarins. Folia Primatologica, 67(3):113-124.
- Prance, G.T. 1979. Notes on the vegetation of Amazonia III. The terminology of Amazonian forest types subject to inundation. Brittonia, 31(1):26-38.
- Prance, G.T. 1987. Vegetation. In: Whitmore, T.C.; Prance, G.T. (Eds). Biogeography and quartenary History in Tropical America Clarendon Press, Oxford. p. 28-44.
- Prance, G.T.; Rodrigues, W.A.; da Silva, M.F. 1976. Inventário florestal de um hectare de mata de terra firme km 30 da Estrada Manaus - Itacoatiara. Acta Amazonica, 6(1):9-35.
- Räsänen, M.E.; Salo, J.S.; Kalliola, R.J. 1987. Fluvial perturbance in the Western Amazon Basin - regulation by long-term sub-Andean tectonics. Science, 238(4832):1398-1401.
- Salo, J.; Kalliola, R.; Hakkinen, I.; Makinen, Y.; Niemela, P.; Puhakka, M.; Coley, P.D. 1986. River dynamics and the diversity of Amazon lowland forest. Nature, 322(6076):254-258.
- Santos, J. 1996. Análise de modelos de regressão para estimar a fitomassa de floresta tropical úmida de terra-firme da Amazonia Central. Tese de doutorado, Universidade Federal de Viçosa, Minas Gerais.
- Sioli, H. 1968. Hydrochemistry and geology in the Brazilian Amazon region. Amazoniana, 1:267-277.
- Sombroek, W.G. 1984. Soils of the Amazon region. In: Sioli, H. (Ed). The Amazon - Limnology and landscape ecology of a mighty tropical river and its basin Junk Publishers, Dordrecht. p. 521-535.
- ter Steege, H.; Sabatier, D.; Castellanos, H.; Van Andel, T.; Duivenvoorden, J.; De Oliveira, A.A.; Ek, R.; Lilwah, R.; Maas, P.; Mori, S. 2000. An analysis of the floristic composition and diversity of Amazonian forests including those of the Guiana Shield. Journal of Tropical Ecology, 16:801-828.
- Terborgh, J. 1985. Habitat selection in Amazonian birds. In: Cody, M.L. (Ed). Habitat selection in birds Academic Press Inc., New York. p. 311-338.
- Terborgh, J.; Andresen, E. 1998. The composition of Amazonian forests: patterns at local and regional scales. Journal of Tropical Ecology, 14:645-664.
- Tuomisto, H.; Ruokolainen, K. 1994. Distribution of Pteridophyta and Melastomataceae along an edaphic gradient in an Amazonian rain-forest. Journal of Vegetation Science, 5(1):25-34.
- Tuomisto, H.; Ruokolainen, K.; Kalliola, R.; Linna, A.; Danjoy, W.; Rodriguez, Z. 1995. Dissecting Amazonian Biodiversity. Science, 269(5220):63-66.
- Voss, R.S.; Emmons, L.H. 1996. Mammalian diversity in Neotropical lowland rainforests: A preliminary assessment. Bulletin of the American Museum of Natural History,(230):3-115.
- Wittmann, F.; Junk, W.J. 2003. Sapling communities in Amazonian white-water forests. Journal of Biogeography, 30(10):1533-1544.
Publication Dates
-
Publication in this collection
13 Apr 2006 -
Date of issue
Mar 2006
History
-
Received
03 Feb 2005 -
Accepted
02 Jan 2006