Resumos
A fixação biológica de nitrogênio que ocorre em leguminosas é realizada por um grupo de bactérias conhecidas como rizóbios. A sobrevivência destas bactérias no solo é influenciada por diversos fatores como a temperatura, umidade e fertilidade do solo. O objetivo deste trabalho foi avaliar a dinâmica da população de rizóbios em solo após o cultivo de soja, durante o período de estiagem no cerrado de Roraima. Foram amostradas três áreas: i) cerrado nativo como referência; ii) área cultivada uma vez com soja inoculada com rizóbio; iii) e área cultivada duas vezes com soja inoculada com rizóbio em anos consecutivos. O solo foi coletado na profundidade de 0-10 cm em cinco períodos a partir do inicio da estiagem no mês de setembro de 2006 coincidindo com a época de colheita da soja e prolongando-se até março de 2007 (0, 45, 90, 135 e 180 dias). A população de rizóbios no solo foi avaliada pela técnica do número mais provável (NMP) utilizando plantas de soja e de feijão-caupi como espécies isca. Foi observado que na área nativa praticamente não existiam bactérias nodulantes de soja, mas havia uma população capaz de nodular o feijão-caupi de até algumas centenas de rizóbios por gramas de solo. O cultivo da soja utilizando sementes inoculadas elevou a população de rizóbios no solo que foi constatada por ambas às espécies de plantas isca. Nas áreas cultivadas, constatou-se uma intensa redução da população de rizóbios no solo, em especial logo após a colheita da soja, continuando o decréscimo até o último período de avaliação. Conclui-se que o cultivo da soja inoculada com rizóbio eleva a densidade de rizóbios em solo do cerrado, mas durante a estiagem ocorre uma drástica redução dessa população, que pode chegar a mais de 99% considerando o início e final do período.
Glycine max; Vigna unguiculata; nodulação; número mais provável
The biological nitrogen fixation in legumes is performed by a group of bacteria known as rhizobia. The survival of these bacteria in soils is affected by several factors, such as temperature, drought and soil fertility. This study was performed to evaluate the dynamics of rhizobia in the soil after soybean cultivation and during a dry season in the cerrado of Roraima. Three areas were sampled: i) native cerrado as reference; ii) an area previously cultivated with soybean for one season; and iii) another one cultivated for two seasons also with soybean. The soil was sampled at a depth of 0-10 cm in five times (0, 45, 90, 135 and 180 days) during the dry season (September 2006 to March 2007). The rhizobial density in the soil was evaluated by the most probable number method with infection of soybean and cowpea plants. It was observed very low number of soybean nodulating bacteria in the reference area, but a high density, of up to several hundred rhizobia capable to nodulate cowpea was measured in this same area. Cropping of soybean with inoculated seeds increased rhizobial density evaluated by both trapping hosts. In cropped areas, an intense reduction of rhizobium density was observed just after soybean harvest, and this reduction continued until the end of the period of evaluation. It was concluded that soybean cultivation increases the density of rhizobial in the cerrado soil; however, this density is drastically reduced, during the dry season, by 99% at the end of the dry period.
Glycine max; Vigna unguiculata; nodulation; most probable number
AGRONOMIA
Dinâmica de rizóbios em solo do cerrado de Roraima durante o período de estiagem
Dynamic of rhizobia in the soil during the dry season in cerrado of Roraima
Jerri Édson ZilliI; Gilmara Maria Duarte PereiraII; Izaias França JúniorII; Krisle da SilvaIII; Mariangela HungriaIV; Janaina Ribeiro Costa RouwsI
IEmbrapa Agrobiologia. Rodovia BR 465, km 7, Seropédica - RJ, CEP: 23890-000. Fone: (21) 3441-1500 - Fax: (21) 2682-1230. E-mail: jerri.zilli@embrapa.br; janaina.rouws@embrapa.br
IIUniversidade Federal de Roraima - UFRR. Av. Ene Garcez, 2413, Boa Vista - RR, CEP: 69304-000. E-mail: gmdpereira@hotmail.com; izafrajunior@yahoo.com.br
IIIEmbrapa Roraima. Rodovia BR 174, km 8, Boa Vista - RR, CEP: 69301-970. E-mail: krisle.silva@embrapa.br
IVEmbrapa Soja. Rod. Carlos João Strass - Distrito de Warta, Londrina- PR, CEP 86001-970. E-mail: mariangela.hungria@embrapa.br
RESUMO
A fixação biológica de nitrogênio que ocorre em leguminosas é realizada por um grupo de bactérias conhecidas como rizóbios. A sobrevivência destas bactérias no solo é influenciada por diversos fatores como a temperatura, umidade e fertilidade do solo. O objetivo deste trabalho foi avaliar a dinâmica da população de rizóbios em solo após o cultivo de soja, durante o período de estiagem no cerrado de Roraima. Foram amostradas três áreas: i) cerrado nativo como referência; ii) área cultivada uma vez com soja inoculada com rizóbio; iii) e área cultivada duas vezes com soja inoculada com rizóbio em anos consecutivos. O solo foi coletado na profundidade de 0-10 cm em cinco períodos a partir do inicio da estiagem no mês de setembro de 2006 coincidindo com a época de colheita da soja e prolongando-se até março de 2007 (0, 45, 90, 135 e 180 dias). A população de rizóbios no solo foi avaliada pela técnica do número mais provável (NMP) utilizando plantas de soja e de feijão-caupi como espécies isca. Foi observado que na área nativa praticamente não existiam bactérias nodulantes de soja, mas havia uma população capaz de nodular o feijão-caupi de até algumas centenas de rizóbios por gramas de solo. O cultivo da soja utilizando sementes inoculadas elevou a população de rizóbios no solo que foi constatada por ambas às espécies de plantas isca. Nas áreas cultivadas, constatou-se uma intensa redução da população de rizóbios no solo, em especial logo após a colheita da soja, continuando o decréscimo até o último período de avaliação. Conclui-se que o cultivo da soja inoculada com rizóbio eleva a densidade de rizóbios em solo do cerrado, mas durante a estiagem ocorre uma drástica redução dessa população, que pode chegar a mais de 99% considerando o início e final do período.
Palavras chave:Glycine max; Vigna unguiculata; nodulação; número mais provável.
ABSTRACT
The biological nitrogen fixation in legumes is performed by a group of bacteria known as rhizobia. The survival of these bacteria in soils is affected by several factors, such as temperature, drought and soil fertility. This study was performed to evaluate the dynamics of rhizobia in the soil after soybean cultivation and during a dry season in the cerrado of Roraima. Three areas were sampled: i) native cerrado as reference; ii) an area previously cultivated with soybean for one season; and iii) another one cultivated for two seasons also with soybean. The soil was sampled at a depth of 0-10 cm in five times (0, 45, 90, 135 and 180 days) during the dry season (September 2006 to March 2007). The rhizobial density in the soil was evaluated by the most probable number method with infection of soybean and cowpea plants. It was observed very low number of soybean nodulating bacteria in the reference area, but a high density, of up to several hundred rhizobia capable to nodulate cowpea was measured in this same area. Cropping of soybean with inoculated seeds increased rhizobial density evaluated by both trapping hosts. In cropped areas, an intense reduction of rhizobium density was observed just after soybean harvest, and this reduction continued until the end of the period of evaluation. It was concluded that soybean cultivation increases the density of rhizobial in the cerrado soil; however, this density is drastically reduced, during the dry season, by 99% at the end of the dry period.
Keywords:Glycine max; Vigna unguiculata; nodulation; most probable number.
INTRODUÇÃO
A fixação biológica de nitrogênio (FBN) realizada por bactérias formadoras de nódulos nas raízes de plantas pertencentes à família das leguminosas, conhecidas coletivamente como rizóbios, é afetada por diversos fatores edafoclimáticos (Zahran 1999; Sadowsky 2005). Entre estes fatores, a umidade do solo representa uma das maiores limitações em regiões tropicais, onde a escassez de chuvas pode ocorrer associada com altas temperaturas (Hungria e Vargas 2000).
De forma geral, o solo é considerado sob déficit hídrico quando os teores de umidade estão abaixo de 50% da capacidade de campo sendo comum nessa condição o aparecimento de efeitos negativos ao processo de FBN (Chalk et al. 2010). Os longos períodos de estiagem, com potencial hídrico do solo abaixo de -0,5 MPa representam uma condição que tende a reduzir a densidade populacional de rizóbios no solo (Hungria e Vargas 2000; Sadowsky 2005); embora algumas estirpes sejam capazes de sobreviver mesmo em condições áridas (Waldon et al. 1989). A escassez de água provoca redução na abundância de microrganismos porque, além de fazer parte da estrutura celular, é indispensável à maioria das reações do solo, como a produção primária de energia, a decomposição da matéria orgânica, a liberação de nutrientes, o crescimento vegetal e a exsudação radicular (Sadowsky 2005).
Alguns estudos têm sido conduzidos para avaliar o impacto do déficit hídrico sobre a FBN, tanto em soja [Glycine max (L.) Merrill], quanto em feijão-caupi [Vigna uniguiculata (L.) Walp] (Serraj e Sinclair 1998; Figueiredo et al. 1999; Streeter 2003). Entretanto, existem poucas informações sobre a sobrevivência dos rizóbios em solos cultivados com soja em condições de estiagem prolongada, característica edafoclimática marcante no cerrado do estado de Roraima.
O cerrado de Roraima, segundo a classificação de Köppen, apresenta clima tipo Aw tropical chuvoso (Barbosa 1997), com precipitação média anual de aproximadamente 1800 mm e concentração de mais de 80% das chuvas entre os meses de abril e setembro e estiagem de até 120 dias (Mourão et al . 2006). Aliado a isto, o solo da região apresenta baixa capacidade natural de retenção de água devido aos baixos teores de matéria orgânica - frequentemente inferiores a 10 g kg-1 - e textura arenosa a franco-arenosa. Estas condições caracterizam um déficit hídrico no solo que pode reduzir drasticamente a densidade de rizóbios.
A quantificação da população de rizóbios no solo constitui uma informação útil, haja vista que define a necessidade ou não de inoculação das leguminosas, quando estas forem reintroduzidas em uma mesma área. Além disso, o conhecimento da dinâmica de rizóbio no solo pode ser utilizado estrategicamente para melhorar o desempenho de inoculantes. Por exemplo, Martins et al. (2003), verificaram aumento da resposta da inoculação do feijão-caupi após longo período de estiagem, isto porque a estiagem havia reduzido a população de bactérias estabelecida no solo. Nestas condições, portanto, o conhecimento da dinâmica da população de rizóbio no solo permitiu aumentar a competitividade da estirpe inoculante frente à população estabelecida no solo.
Neste estudo, para avaliar a dinâmica de bactérias nodulantes presentes no solo durante o período de estiagem foram realizadas amostragens em uma área de referência, uma área cultivada uma vez com soja inoculada com rizóbio e outra área cultivada duas vezes com soja inoculada com rizóbio em anos consecutivos, utilizando-se como espécies isca a soja e o feijão-caupi. Esta estratégia foi adotada, pois diferentemente da cultura da soja, o feijão-caupi possui maior capacidade de nodulação quando associado com bactérias nativas de solos brasileiros (Zilli et al. 2009) e o histórico de cultivo do solo interfere na densidade de rizóbio (Ferreira et al. 2000).
O objetivo deste trabalho foi avaliar a dinâmica da população de rizóbios presentes em solos de Cerrado de Roraima após o cultivo de soja e durante o período de estiagem.
MATERIAL E MÉTODOS
As amostragens do solo foram realizadas no Campo Experimental Água Boa da Embrapa Roraima, localizado no município de Boa Vista, Roraima (02º15'00"N, 60º39'54"W). As amostragens foram realizadas em três condições: i) uma área de cerrado nativo apenas com a presença de vegetação espontânea, como referência; ii) uma área aberta para o plantio em 2005 com correção do solo com cerca de 1500 kg ha-1 de calcário dolomítico (poder relativo de neutralização de 85%), 500 kg ha-1 de superfosfato simples e 50 kg ha-1 de adubo formulado contendo micronutrientes (FTE BR-12) e, que havia recebido um único cultivo de soja em 2006 (1º cultivo); e iii) uma área também preparada em 2005, mas com cultivo de soja nas safras de 2005 e 2006 (2º cultivo). Em todos os cultivos, as sementes de soja foram inoculadas com estirpes de Bradyrhizobium spp. indicadas para a cultura na proporção de 1,2 milhão de células por semente e durante o período de estiagem todas as áreas permaneceram apenas com vegetação espontânea. Todas as áreas apresentavam a mesma declividade e eram distanciadas umas das outras em aproximadamente 100 m. Foram realizadas amostragens de solo para análise química e física e, na sede do campo experimental, situado a cerca de 600 m de distância da área de estudo, coletou-se diariamente a precipitação pluvial com um pluviômetro de campo.
Para as amostragens, cada área foi definida com dimensões aproximadas de 60 m x 100 m e, cada uma, foi dividida em três talhões com largura aproximada de 20 m, considerando-se cada talhão um bloco. Em cada bloco coletou-se uma amostra de solo composta por dez amostras simples retiradas a cada 10 m, aproximadamente, no sentido da maior dimensão. As amostras simples foram retiradas na profundidade de 0-10 cm com o auxílio de um trado tipo holandês, o qual foi desinfestado com etanol 95% antes das tradagens em cada bloco e área. Uma vez coletadas, as amostras foram transportadas imediatamente na temperatura ambiente e abrigadas da luz para o laboratório de Microbiologia do Solo da Embrapa Roraima, quando foram processadas no máximo 48 h após a coleta. Foram realizadas cinco amostragens de solo ao longo de sete meses, iniciando-se no dia 15 de setembro de 2006 -cerca de uma semana após a colheita da soja e coincidindo com o início do período de estiagem- e repetindo-se a cada 45 dias até o mês de março de 2007. Desta forma, os períodos amostrados compreenderam 0, 45, 90, 135 e 180 dias.
A determinação da população de rizóbio no solo foi realizada pelo método do número mais provável (NMP) com infecção em plantas de soja (cv BRS Tracajá) e feijão-caupi (BRS Mazagão), as quais foram cultivadas em vasos de Leonard com substrato de areia: vermiculita (2:1) esterilizado em autoclave. Antes do plantio, as sementes foram desinfestadas pela imersão em etanol 70% por dois minutos, peróxido de hidrogênio 10% por cinco minutos e lavadas 10 vezes com água destilada esterilizada. Foram semeadas três sementes por vaso, deixando-se apenas uma planta após o desbaste realizado por ocasião da emergência. Os vasos, em triplicata, foram mantidos em câmara de crescimento com temperatura e umidade relativa do ar de aproximadamente 28 ºC e 65%, respectivamente.
O inóculo foi preparado de acordo com Andrade e Hamakawa (1994), partindo-se da retirada de fragmentos de material orgânico, seguindo peneiramento com malha de 4 mm, diluição em solução salina (85 g L-1 de NaCl em água destilada) e sucessivas diluições até a obtenção da diluição 10-7, da qual, juntamente com as diluições 10-4, 10-5 e 10-6, foram tomadas alíquotas de 1 mL que foram inoculadas nas plântulas de soja e feijão-caupi logo após o desbaste. Semanalmente cada vaso recebeu 300 mL da solução nutritiva de Norris (Andrade e Hamakawa 1994).
As coletas das plantas foram realizadas aos 25 dias após a emergência, quando avaliou-se a nodulação das raízes, atribuindo-se caráter positivo às amostras cujas plantas apresentaram ao menos um nódulo e, negativo, para aquelas que não apresentaram nódulos. Para a estimativa do número de células bacterianas nodulantes utilizou-se a tabela de número mais provável (Andrade e Hamakawa 1994) para a obtenção do fator NMP. O número final de células bacterianas nodulantes foi obtido pela média das três repetições para cada amostra de solo, realizando também a correção em relação à umidade do solo.
A comparação das densidades médias de rizóbio entre as áreas foi realizada através de análise de variância e teste t a 5% de probabilidade, em cada período de avaliação para cada cultura e, também, na média geral dos períodos. Quando necessário foram realizadas transformação dos dados (log de x+1) para o atendimento das pressuposições da análise.
Para avaliar a flutuação da densidade de rizóbio ao longo do período de estudo, foi realizada uma análise de variância a 5% de probabilidade com os dados de cada uma das áreas, totalizando assim três análises, considerando o delineamento em blocos casualizados, com três repetições, em esquema fatorial 5x2, correspondendo às cinco épocas de coleta dos dados (0, 45, 90, 135 e 180 dias) e as duas espécies isca (soja e feijão-caupi). A partir destas análises foram verificados ajustes de modelos de regressão aos dados para o fator época para cada espécie isca.
RESULTADOS E DISCUSSÃO
Constatou-se com as análises física e química do solo que todas as áreas estudadas apresentavam solo arenoso e baixo teor de matéria orgânica e de potássio (Tabela 1), que são características naturais dos solos dos cerrados de Roraima (Benedetti et al. 2011). Entretanto, em decorrência da correção e adubação durante o preparo do solo, as áreas cultivadas apresentaram teores mais elevados de Ca, Mg e P, comparativamente com a área de referência, não havendo diferenças expressivas entre elas (Tabela 1).
Durante o período deste estudo houve precipitação pluvial decrescente desde setembro de 2006 até o final de fevereiro de 2007 (Tabela 2). Além disso, foram observadas irregularidades na distribuição das chuvas dentro de cada mês e, praticamente, ausência por mais de 90 dias, a partir da última semana de novembro até o final de fevereiro de 2007. Este regime de chuvas no período de estudo foi semelhante à série histórica registrada para esta área de cerrado desde o ano de 1994, quando as medições foram iniciadas (Mourão et al. 2006).
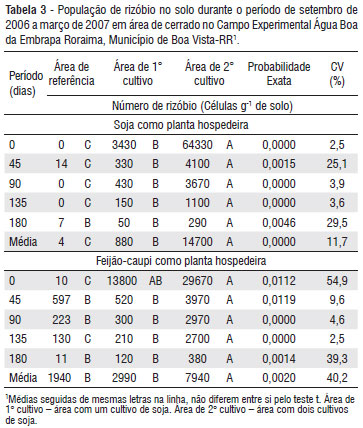
De forma geral, na área de referência foi observado baixo número de bactérias nodulantes, cerca de 10 células por grama de solo na média geral das duas espécies isca sendo inferior às médias das áreas com um e dois cultivos de soja, as quais apresentaram valores de 1,94 x 103 a 11,3 x 103 células g-1 solo, respectivamente. Além disso, na área de referência ocorreram diferenças entre as duas espécies isca, isto é, praticamente não foi observada nodulação em soja, enquanto que para o feijão-caupi detectou-se a presença de rizóbios em todas as coletas, que variou de algumas dezenas a até centenas de células por grama de solo (Tabela 3).
Estes dados corroboram informações anteriores que indicaram que a soja praticamente não estabelece nodulação com bactérias nativas de solos brasileiros e, ao contrário, o feijão-caupi, apresenta ampla capacidade de nodulação com estas bactérias (Hungria et al. 2005; Zilli et al. 2009). Esta capacidade do feijão-caupi nodular com bactérias nativas de solos brasileiros já foi demonstrada em diferentes regiões incluindo Mata Atlântica, Cerrado, Amazônia e Semiárido (Martins et al. 2003; Zilli et al. 2004; Lima et al; 2005; Chagas Júnior et al. 2009).
Considerando a soja como planta isca, observou-se que o cultivo com essa leguminosa ocasionou maior densidade de rizóbio em todos os períodos de avaliação, sendo o valor na área referência menor do que o das demais áreas e o da área de 1º cultivo maior que a área de referência, porém menor do que o da área de 2º cultivo (Tabela 3). Isto indica que o cultivo da soja a partir de sementes inoculadas promoveu, de forma geral, aumento na densidade de rizóbio no solo e, esta densidade foi elevada com o aumento do número de cultivos.
Por outro lado, considerando o feijão-caupi como espécie isca foi observado diferenças significativas entre a área de referência e a de 1º cultivo apenas na primeira e terceira avaliação, enquanto que nas duas áreas cultivadas ocorreram diferenças significativas em todas as coletas a exceção da primeira (Tabela 3). Isto mostra que mesmo em área nativa existe uma população de rizóbios estabelecida capaz de nodular o feijão-caupi e que o cultivo da soja tende a aumentar esta população, entretanto, em proporções menores do que as observadas para a soja.
O cultivo sucessivo de uma leguminosa em uma mesma área tende a elevar a densidade de bactérias nodulantes no solo, isto porque a multiplicação dos rizóbios é favorecida com a nodulação das plantas e estes nódulos representam uma proteção contra as adversidades edafoclimáticas (Ferreira et al. 2000; Hungria e Vargas 2000). Ferreira et al. (2000), em estudo conduzido no Paraná em sistema de plantio direto e convencional em rotação e sucessão de culturas, observaram que o cultivo da soja em sucessão com trigo (Triticum aestivum) ou milho (Zea mays), mesmo sem a inoculação de sementes durante um período de 15 anos, proporcionou aumento da população de bactérias nodulantes da ordem de 102 para 103, comparativamente a uma área sem o cultivo de leguminosas. Além disso, também observaram que não apenas a densidade de Bradyrhizobium, mas também a diversidade genética destas bactérias aumentou na área com cultivo de soja (Ferreira et al. 2000).
No Brasil, a introdução da soja nos solos de cerrado da região central constitui exemplo de que a presença da leguminosa proporciona alteração na estrutura da comunidade de bactérias nodulantes no solo (Mendes et al. 2004; Hungria et al. 2005). Segundo estes autores, quando da introdução da soja nas décadas de 60 e 70 do século passado, os solos desta região praticamente não possuíam bactérias nodulantes de soja e, portanto, recomendava-se o uso de algumas estirpes exóticas, incluindo a CB1809, a 532C e a SEMIA 5020. Passados 15 a 20 anos, entretanto, estes solos passaram a apresentar alta diversidade de Bradyrhizobium nodulantes de soja com estirpes com perfis de proteínas semelhantes às estirpes originais, mas apresentando diferenças genéticas e de eficiência na FBN marcantes (Galli-Terasawa et al. 2003; Mendes et al. 2004).
Com relação ao número de rizóbios ao longo do período dos 180 dias do estudo não foi observada uma tendência definida de flutuação da população na área de referência. Para a soja, como mencionado anteriormente, ocorreu nodulação apenas esporádica e, para o feijão-caupi, em termos absolutos ora observou-se população mais elevada, ora mais baixa, mas sem identificação de possíveis fatores que estivessem influenciando esta população. Provavelmente, o principal fator envolvido na manutenção desta população seja a presença de leguminosas nativas, que são abundantes nestas áreas de cerrado (Flores e Rodrigues 2010).
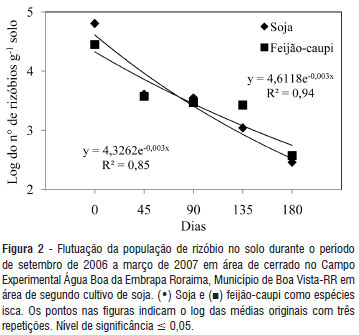
Para as áreas cultivadas, embora no primeiro período existisse uma média até acima de 60 x103 rizóbios por grama de solo no caso da soja e, que o aumento do número de cultivos de soja tenha elevado este número (Tabela 3), nos períodos subsequentes a média ficou entre uma dezena e uma centena de rizóbios por grama de solo na área de 1º cultivo (Figura 1 e 2) e entre uma centena a alguns milhares na área de 2º cultivo (Figura 1 e 2). Também observou-se que a flutuação da população de rizóbio para ambas espécies isca foi semelhante, assim como a proporção de redução em ambas as áreas cultivadas (Figura 1 e 2). Além disso, foi possível constatar que a densidade de rizóbios no solo sofreu uma intensa redução (na média mais de 90% em teremos absolutos) nos primeiros 45 dias do estudo e, considerando o inicio e final do estudo a redução desta densidade chega a superar 99% da população inicial.
No período inicial ocorreram chuvas moderadas, com boa distribuição e acúmulo de mais de 120 mm e (Tabela 2) e que, portanto, aparentemente ainda não configuraria uma escassez de água no solo que afetasse grandemente as bactérias do solo. Por outro lado, devido à primeira coleta de solo ter sido realizada logo após a colheita da soja, a ausência da cultura tornou-se um fator importante capaz de justificar a intensa redução da densidade de rizóbios no solo do primeiro para o segundo período de amostragem. Posteriormente, entretanto, no mês de novembro, as chuvas se tornaram escassas, passando a ser o fator mais marcante na redução da população de rizóbios no solo, que variou de alguns milhares a uma centena de células g-1 solo durante cada período, respectivamente na média da área de 1º cultivo e na área de 2º cultivo (Figura 1 e 2).
É sabido que tanto a presença da leguminosa quanto a disponibilidade de água afetam a sobrevivência de rizóbios no solo (Hungria e Vargas 2000) e, portanto a colheita da cultura da soja e a escassez de chuvas no período do estudo certamente influenciaram na densidade de rizóbios no solo do cerrado de Roraima. Neste caso, a retirada da cultura reduziria grandemente a população de rizóbio logo nas primeiras semanas após a colheita e, a escassez de chuvas continuaria a reduzir esta população durante os meses de estiagem subsequentes.
No caso específico da soja, observou-se que, após a estiagem, restaram no solo cerca de 50 e 200 células de rizóbio g-1 solo, respectivamente na área de 1º e 2º cultivos. Pesquisas realizadas nas décadas passadas evidenciaram que uma concentração entre 100 a 1000 células de rizóbio g-1 de solo teoricamente garantiriam uma adequada nodulação das plantas e o processo de FBN em diversas leguminosas (Herridge 2008). Entretanto, em experimentos conduzidos nas principais regiões produtoras de soja do Brasil constatou-se que em áreas com plantio de soja inoculada por várias safras, há o estabelecimento de uma população nodulante da ordem de 103 a até 106 células g-1 solo e, mesmo com esta alta população de bactérias nesses solos, foram observados incrementos de produtividade de grãos de até 15% pela re-inoculação da semente de soja anualmente (Hungria et al. 2007).
Desta forma, este trabalho demonstra que, em solos do cerrado, a estiagem provoca grande redução na densidade rizóbio e, mesmo que o cultivo da soja ao longo das safras tenda a elevar esta população, a inoculação da soja é necessária, haja vista que a população de bactérias no solo seria insuficiente para garantir uma boa nodulação e, portanto, os riscos de insucesso na lavoura seriam evidentes.
CONCLUSÕES
O número de cultivos de soja inoculada eleva a densidade de rizóbios no solo do cerrado de Roraima;
Durante o período de estiagem no cerrado de Roraima ocorre uma drástica redução na densidade de rizóbios no solo, que pode superar 99%, considerando a população no início e final do período.
BIBLIOGRAFIA CITADA
Recebido em: 24/01/2012
Aceito em:05/06/2012
- Andrade, D.S.; Hamakawa, P.J. 1994. Estimativa do número de células viáveis de rizóbio no solo e em inoculantes por infecção em plantas, p.63-94. In: Hungria, M.; Araújo, R.S. (Eds). Manual de métodos empregados em estudos de microbiologia agrícola Embrapa-SPI, Brasília, Distrito Federal.
- Barbosa, R.I. Homem, ambiente e ecologia no Estado de Roraima, p.325-335. In: Barbosa, R.I.; Ferreira, E.J.; Castellón, E.G. (Eds). Homem, ambiente e ecologia no Estado de Roraima. INPA, Manaus, Amazonas.
- Benedetti, U.G.; Vale Júnior, J. F.; Schaefer, C.E. G.R.; Melo, V.F.; Uchôa, S.C.P. 2011. Gênese, química e mineralogia de solos derivados de sedimentos pliopleistocênicos e de rochas vulcânicas básicas em Roraima, Norte Amazônico. Revista Brasileira de Ciência do Solo, 35:299-312.
- Chagas Junior, A.F.; Oliveira, L.A.; Oliveira, A.N.; Willerding, A.L. 2009. Efetividade de rizóbios e caracterização fenotípica dos isolados que nodulam feijão-caupi em solos da Amazônia Central. Acta Amazonica, 39:489-494.
- Chalk, P.M.; Alves, B.J.R.; Boddey, R.M.; Urquiaga, S. 2010. Integrated effects of abiotic stresses on inoculant performance, legume growth and symbiotic dependence estimated by 15N dilution. Plant and Soil, 328:1-16.
- Empresa Brasileira de Pesquisa Agropecuária - EMBRAPA. Centro Nacional de Pesquisa de Solos. Sistema Brasileiro de Classificação de Solos. 2 ed. Rio de Janeiro, 2006. 306p.
- Ferreira, M.C.S.; Andrade, D.O.; Chueire, L.M.; Takemura, S.M.; Hungria, M. 2000. Tillage method and crop rotation effects on the population sizes and diversity of bradyrhizobia nodulating soybean. Soil Biology and Biochemistry, 32:627-637.
- Figueiredo, M.V.B.; Vilar, J.J.; Burity, H.A.; França, F.P. 1999. Alleviation of water stress effects in cowpea by Bradyrhizobium spp. inoculation. Plant and Soil, 207:67-75.
- Flores, A. S.; Rodrigues, R. S. 2010. Diversidade de leguminosae em uma área de savana do estado de Roraima, Brasil. Acta Botanica Brasilica, 24:175-183.
- Galli-Terasawa, L.V.; Glienke-Blanco, C.; Hungria, M. 2003. Diversity of a soybean rhizobial population adapted to a Cerrados soil. World Journal of Microbiology and Biotechnology, 19:933-939.
- Herridge, D.F. 2008. Inoculation technology for legumes, p. 77-115. In: Dilworth, M.J.; James, E.K.; Sprent, J.I.; Newton, W.E. (Eds.). Nitrogen-fixing Leguminous Symbioses v. 7. Springer, Dordrecht, Netherlands.
- Hungria, M.; Campo, R.J.; Mendes, I.C. 2007. A importância do processo de fixação biológica do nitrogênio para a cultura da soja: componente essencial para a competitividade do produto brasileiro Documentos, 283. Embrapa Soja, Londrina, Paraná. 80pp.
- Hungria, M.; Franchini, J.; Campo, R.; Graham, P. 2005. The importance of nitrogen fixation to soybean cropping in South America, p. 25-42. In: Werner, D.; Newton, W. (Eds.). Nitrogen Fixation in Agriculture, Forestry, Ecology, and the Environment v.4. Springer, Dordrecht, Netherlands.
- Hungria, M.; Vargas, M.A.T. 2000. Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Research, 65:151-164.
- Lima, A.S.; Pereira, J.P.A.R.; Moreira, F.M.S. 2005. Diversidade fenotípica e eficiência simbiótica de estirpes de Bradyrhizobium spp. de solos da Amazônia. Pesquisa Agropecuária Brasileira, 40:1095-1104.
- Martins, L.M.V.; Xavier, G.R.; Rangel, F.W.; Ribeiro, J.R.A.; Neves, M.C.P.; Morgado, L.B.; Rumjanek, N.G. 2003. Contribution of biological nitrogen fixation to cowpea: a strategy for improving grain yield in the semi-arid region of Brazil. Biology and Fertility of Soils, 38:333-339.
- Mendes, I.; Hungria, M.; Vargas, M. 2004. Establishment of Bradyrhizobium japonicum and B. elkanii strains in a Brazilian Cerrado oxisol. Biology and Fertility of Soils, 40:28-35.
- Mourão Junior, M.; Moura Neto, M.A.; Bendahan, A.B.; Xaud, M.R.; Xaud, H.A.M. 2006. Probabilidade de precipitação pluvial mensal no campo experimental Água Boa, Boa Vista-Roraima (1991-2005). Boletim de Pesquisa e Desenvolvimento, 08. Embrapa Roraima, Boa Vista, Roraima. 18 pp.
- Sadowsky, M. 2005. Soil stress factors influencing symbiotic nitrogen fixation, p. 89-112. In: Werner, D.; Newton, W. (Eds.). Nitrogen Fixation in Agriculture, Forestry, Ecology, and the Environment v. 4. Springer, Dordrecht, Netherlands.
- Serraj, R.; Sinclair, T.R. 1998. Soybean cultivar variability for nodule formation and growth under drought. Plant and Soil, 202:159-166.
- Streeter, J.G. 2003. Effects of drought on nitrogen fixation in soybean root nodules. Plant, Cell and Environment, 26:1199-1204.
- Waldon, H. B.; Jenkins, M. B.; Virginia, R.A.; Harding, E.E. 1989. Characteristics of woodland rhizobial populations from surface and deep soil environment of the Sonoran Desert. Applied and Environmental Microbiology, 55:3058-3064.
- Zahran, H.H. 1999. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiology and Molecular Biology Reviews, 63:968-989.
- Zilli, J.E.; Valisheski, R.R.; Freire Filho, F.R.; Neves, M.C.P.; Rumjanek, N.G. 2004. Assessment of cowpea rhizobium diversity in Cerrado areas of northeastern Brazil. Brazilian Journal of Microbiology, 35:281-287.
- Zilli, J.E.; Vilarinho, A.A.; Alves, J.M.A. 2009. A cultura do feijão-caupi na Amazônia brasileira: Embrapa Roraima, Boa Vista, Roraima. 356 pp.
Datas de Publicação
-
Publicação nesta coleção
27 Fev 2013 -
Data do Fascículo
Jun 2013
Histórico
-
Recebido
24 Jan 2012 -
Aceito
05 Jun 2012