ABSTRACT
Rapid assessments are the most common information source on biodiversity in the northwestern Amazon in Colombia due to limited resources and logistic constraints. These assessments are essential for decision-making on environmental policies in this region, that has been strongly impacted by the transformation of its natural ecosystems. Several local camera-trapping rapid assessments of medium and large-sized mammals (MLM) have been conducted in the Colombian Amazon, but they are difficult to compare. We analyzed information of 16 of these rapid assessments of MLM to provide the first list of MLM in the northwestern Amazon in Colombia. We also evaluated the accuracy of four estimators (ICE, Chao-2, Jackknife-1, and Jackknife-2), and the minimum sampling effort for the estimation of MLM richness in local surveys in the region. We report 26 species of MLM for the Colombian Amazon (between five and 13 species per locality), which is an underestimation of MLM richness in the region. The best estimator of MLM richness was the Jackknife-1, due to its precision and the lower influence of singletons. We recommend a minimum sampling effort of 350 camera trap-days. Although rapid assessments do not allow a robust estimation of MLM richness, they record the most common species (or core species) per locality and their abundance variation. The evaluation of the effect of habitat transformation on MLM and the estimation of population parameters of rare species require more intensive studies.
KEYWORDS:
camera trapping; neotropics; rainforest; sampling effort; satellite species; species richness
RESUMEN
Las evaluaciones rápidas de biodiversidad son la fuente de información más común sobre biodiversidad en el noroccidente amazónico en Colombia debido a que los recursos y la logística son limitados. Estas evaluaciones son esenciales para tomar decisiones ambientales en esta área que ha sido impactada por la transformación de sus ecosistemas naturales. Diferentes evaluaciones locales rápidas con cámaras trampa de mamíferos medianos y grandes (MMG) se han realizado en el noroccidente Amazónico en Colombia, pero estas son difícilmente comparables. Nosotros recolectamos la información de 16 evaluaciones rápidas de MMG para proporcionar la primera lista de MMG del noroccidente amazónico en Colombia. Adicionalmente, evaluamos la precisión de cuatro estimadores (ICE, Chao 2, Jackknife 1 y Jackknife 2) y el esfuerzo mínimo de muestreo para la estimación de la riqueza de MMG en estudios locales en esa región. Reportamos 26 especies de MMG para la Amazonía colombiana (entre cinco y 13 especies por localidad), lo que es uns subestimación de la riqueza de MMG en la región. El mejor estimador para este tipo de estudios fue Jackknife 1, debido a su precisión y la baja influencia de singletons. Recomendamos un esfuerzo mínimo de muestreo de 350 cámaras trampa-día. Aunque las evaluaciones rápidas no proveen una estimación robusta de la riqueza de MMG, estas evaluaciones registran las especies más comunes (especies core) y la variación en su abundancia por localidad. Es necesario un mayor esfuerzo de muestreo para evaluar el efecto de la transformación del hábitat sobre los MMG, en especial sobre los parámetros poblacionales de las especies raras.
PALABRAS CLAVE:
esfuerzo de muestreo; especies satélite; neotrópico; selva húmeda tropical; trampas cámara; riqueza de especies
INTRODUCTION
The Colombian Amazon, comprising the northwestern area of the Amazonian biome, is environmentally heterogeneous and recognized for its exceptional biodiversity of mammals (Rodríguez-Mahecha et al. 2006Rodríguez-Mahecha, J.V.; Alberico, M.; Trujillo, F.; Jorgenson, J. 2006. Libro Rojo de los Mamíferos de Colombia. Serie Libros Rojos de Especies Amenazadas de Colombia. Conservación Internacional Colombia, Ministerio de Medio Ambiente, Vivienda y Desarrollo Territorial. Bogotá, 433p.). Ironically, this biodiversity has been poorly studied, and the region remains vastly scientifically unexplored (Stevenson et al. 2004Stevenson, P.R.; Suescún, M.; Quiñonez, M.J. 2004. Characterization of forest types at the CIEM, Tinigua Park, Colombia. Field Studies of Fauna and Flora La Macarena Colombia, 14: 1-20. ; Montenegro 2017Montenegro, O.L. 2007. Mamíferos terrestres del sur de la Amazonia colombiana. In: Ruiz, S.L.; Sánchez, E.; Tabares, E.; Prieto, A.; Arias, J.C.; Gómez, R.; Castellanos, D.; García, P.; Rodríguez, L. (Ed.). Diversidad Biológica y Cultural del Sur de la Amazonia Colombiana. Corpoamazonia, Instituto Humboldt, Instituto Sinchi, UAESPNN, Bogotá, p.134-141.; Trujillo et al. 2019Trujillo, F.; Mosquera-Guerra, F.; Diaz-Pulido, A.; Carvajal-Castro, J.D.; Mantilla-Meluk, H. 2019. Mamíferos del Escudo Guayanés de Colombia. In: Lasso, C.A.; Señaris, J.C. (Ed.). Fauna Silvestre del Escudo Guayanés (Colombia-Venezuela). Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá, p.441-380.), while at the same time being submitted to intense and accelerated ecosystem transformation that involves the highest fire frequencies (Otavo and Murcia 2018Otavo, S.; Murcia, U. 2018. La Amazonía se quema: detección de áreas con mayor ocurrencia de incendios de vegetación como estrategia para la prevención y control. Colombia Amazónica, 11: 59-72. ) and up to 76% of the total deforestation (IDEAM 2020IDEAM. 2020. Instituto de Hidrología, Meteorología y Estudios Ambientales. Boletín de detección temprana de deforestación, # 25. ( (http://smbyc.ideam.gov.co/MonitoreoBC-WEB/pub/alertasDeforestacion.jsp?0.49368119561984636 ). Accessed on 20 Mar 2020.
http://smbyc.ideam.gov.co/MonitoreoBC-WE...
) in Colombia. After the Colombian peace process in 2016, deforestation rates increased between 46% and 346% within protected areas of the Amazon region (Clerici et al. 2020Clerici, N.; Armenteras, D.; Kareiva, P.; Botero, R.; Ramírez-Delgado, J.P.; Forero-Medina, G.; et al. 2020. Deforestation in Colombian protected areas increased during post-conflict periods. Scientific Reports, 10: 4971. ). In this scenario of few studies and high impact, it is urgent to increase the knowledge on biodiversity in the Colombian Amazon, to better understand how different taxonomic groups respond to human pressure, and to provide information for sound environmental and conservation decisions (Boron et al. 2019Boron, V.; Deere, N.J.; Xofis, P.; Link, A.; Quiñones-Guerrero, A.; Payan, E.; Tzanopoulos, J. 2019. Richness, diversity, and factors influencing occupancy of mammal communities across human-modified landscapes in Colombia. Biological Conservation, 232: 108-116. ).
Medium and large mammals are key species in neotropical ecosystems because they control plant and animal populations, acting as predators and herbivores, long-distance seed dispersers (Arévalo-Sandi et al. 2018Arévalo-Sandi, A.; Bobrowiec, P.E.D.; Rodríguez, V.J.U.; Norris, D. 2018. Diversity of terrestrial mammal seed dispersers along a lowland Amazon forest regrowth gradient. PLoS ONE, 13: e0193752. ) and pathogen reservoirs (Mollentze and Streicker 2020Mollentze, N.; Streicker, D.G. 2020. Viral zoonotic risk is homogenous among taxonomic orders of mammalian and avian reservoir hosts. Proceedings of the National Academy of Sciences, 117: 9423-9430. ), among other functions. These mammals also contribute significantly to food security in several indigenous communities (Osorno et al. 2014Osorno, M.; Atuesta-Dimian, N.; Jaramillo, L.F.; Sua, S.; Barona, A.; Roncancio, N. 2014. La Despensa del Tiquié: Diagnóstico y Manejo Comunitario de La Fauna de Consumo en la Guayana Colombiana. Instituto Amazónico de Investigaciones Científicas “SINCHI”, Bogotá, 162p. ) but are sensitive to habitat changes and overexploitation (Romero-Muñoz et al. 2020Romero-Muñoz, A.; Benítez-López, A.; Zurell, D.; Baumann, M.; Camino, M.; Decarre, J.; et al. 2020. Increasing synergistic effects of habitat destruction and hunting on mammals over three decades in the Gran Chaco. Ecography, 43: 954-966. ) and populations of many species are in decline (Benítez-López et al. 2019Benítez-López, A.; Santini, L.; Schipper, A.M.; Busana, M.; Huijbregts, M.A.J. 2019. Intact but empty forests? Patterns of hunting-induced mammal defaunation in the tropics. PLoS Biology, 17: e3000247. ).
To assess the diversity of Colombian Amazon fauna and its use by the native communities, the Instituto Amazónico de Investigaciones Científicas SINCHI has been doing local camera-trap rapid assessments throughout the Colombian Amazon since 2014. Due to economic and logistic constraints, most of these assessments are short-term and differ substantially in sampling effort, making them difficult to compare. Despite this handicap, these inventories accumulate valuable information on medium and large mammal richness in the region. Species richness is the most frequent parameter used to describe communities, being vital for the analysis of species distributions, assessment of human perturbation, and conservation planning (Pineda-López 2019Pineda-López, R. 2019. Estimadores de la riqueza de especies. In: Moreno, C.E. (Ed.). La Biodiversidad en Un Mundo Cambiante: Fundamentos Teóricos y Metodológicos para Su Estudio. Universidad Autónoma del Estado de Hidalgo, Libermex, Ciudad de México, p.159-174. ). Albeit providing only crude estimates, these rapid assessments give a general picture of local mammal assemblage composition, mostly on core species - locally abundant and regionally common species - and some satellite species - species that are sparse and occur at only a few sites with low abundance (Hanski 1982Hanski, I. 1982. Dynamics of regional distribution: the core and satellite species hypothesis. Oikos, 38: 210-221.).
Here we aimed to evaluate data on several rapid camera-trap surveys to provide the first comprehensive assessment of the diversity of medium and large mammals in the northwestern Amazon in Colombia, and to assess the effectiveness of these surveys to record core and satellite species. We also used the data to determine which of several estimators provides the most precise estimation of species richness and recommend a minimum sampling-effort to estimate species richness of medium and large mammals for future studies in the region.
MATERIAL AND METHODS
Study area
The Colombian Amazon comprises the northwestern extreme of the Amazon biome and includes an area of approximately 477,274 km2, representing 42% of the Colombian continental area. This region extends from the eastern slopes of the Andes to the border with Venezuela, Brazil, Peru and Ecuador, and to the forest line north of the Guaviare River (Murcia et al. 2007Murcia, U.G.; Rodríguez, C.H.; Castro, W. 2007. Zonificación ambiental (ecológica y económica) en la Amazonía colombiana. Revista Colombia Amazónica, 1: 119-134. ; Figure 1).

Geographic location of the 16 study areas where camera-trap rapid assessments of medium and large sized mammals were carried out in the Amazon region of Colombia (light green shade). This figure is in color in the electronic version.
The northwestern Amazon is a heterogenic region mainly covered by tropical rainforests. It includes two biogeographic provinces [Amazonas and Guyana (sensuHernández-Camacho et al. 1992Hernández-Camacho, J.I.; Guerra, A.H.; Quijano, R.O.; Walschburger, T. 1992. Unidades biogeográficas de Colombia. In: Halffter, G (Ed.). La Diversidad Biológica de Iberoamérica I. Acta Zoologica Mexicana, Instituto de Ecología, Xalapa, p.105-153.) or Imerí and Napo (sensuMorrone 2014Morrone, J.J. 2014. Biogeographical regionalization of the Neotropical region. Zootaxa, 3782: 1-110. )], and at least eleven biogeographic districts (sensu Hernández-Camacho et al. 1992). The mean annual temperature is cooler near the Andes (24.3-25.4 ºC) and increases towards the northeastern edge of the region (26.2-27.9 ºC) (Rudas 2009Rudas, A. 2009. Unidades ecogeográficas y su relación con la diversidad vegetal de la Amazonia colombiana. Doctoral thesis, Universidad Nacional de Colombia, Colombia. 147p. (https://repositorio.unal.edu.co/handle/unal/11104).
https://repositorio.unal.edu.co/handle/u...
). The precipitation is higher in the Andean foothills (5,400 mm annual rainfall), and decreasing eastwards, with the lowest record in the northeastern portion (2,200 mm), and the highest lowland precipitation records in the south between 2°N and 4°S (3,300-3,900 mm) (Rudas 2009).
Camera-trap sampling data
We used local rapid assessment information on medium and large mammals (MLM) from 16 localities obtained by the SINCHI Institute (Figure 1) in five departments (Caquetá, Guaviare, Guainía, Vaupés, and Vichada), between the Guaviare-River forest (transitional jungles between the Amazonas and the Orinoco Llanos regions) to the north, and the Caquetá River to the south. The distance between localities varies from 20 km to 928 km. The overall survey area comprises ten ecosystems and several vegetation types such as the cloud forest of the Andean foothills, lowland floodplain and terra-firme forest, and natural Amazonian savannas.
The sampling effort of the rapid assessments varied from 60 to 241 camera trap-days (the sum of days during which each camera remained active at a locality) (Table 1). The samplings did not have a specific design, but they had similar parameters: 1) between 7 and 20 cameras were installed at 0.5 to 1 km distance among them, and therefore did not cover areas larger than 15 km2; 2) cameras were set at 40 cm above the ground and programmed to take three pictures per trigger with intervals of one second between photos; 3) cameras ran 24 hours per day, recording the date and time of each photograph; 4) no bait was used to attract mammals; 5) cameras were installed close to track signals, salt licks, and feeding sites to increase capture probability; and 6) in nine surveys, the cameras were placed in the same geographic location through the complete sampling window, while in seven surveys some or all cameras were placed in one locations for a few days and then were changed to different locations. Each geographic location was considered as a different station.
Metadata of camera-trap rapid assessments in 16 localities in the Colombian Amazon. N cameras = number of cameras used in the survey; N stations = number of different sites where cameras were placed during the sampling window; Sampling window = total number of sampling days of the survey; N camera trap-days = total sampling effort defined as the sum of the days that each camera trap kept active in the sampling window.
Our analyses focused on terrestrial mammals with a weight greater than 2 kg, thus we excluded primates and other mainly arboreal mammals seldom captured on camera traps. Rats and mice that may exceed 2 kg (e.g., Proechymis) were also excluded because they cannot be identified without a voucher specimen with a clean skull (Voss and Emmons 1996Voss, R.S.; Emmons, L.H. 1996. Mammalian diversity in Neotropical Lowland Rainforest: A preliminary assessment. Bulletin of the American Museum of Natural History, 230: 1-115.). We based the taxonomic treatment of medium and large mammals on Ramírez-Chaves et al. (2019Ramírez-Chaves, H.E.; Suárez-Castro, A.F.; Sociedad Colombiana de Mastozoología. 2019. Mamíferos de Colombia. ( (https://www.gbif.org/es/dataset/e8b9ed9b-f715-4eac-ae24-772fbf40d7ae ). Accessed on 30 Aug 2019.
https://www.gbif.org/es/dataset/e8b9ed9b...
).
Records of a species at the same site were considered as independent events when the photographs were separated by at least one hour, to avoid pseudoreplication in the species counts (O’Brien et al. 2003O’Brien, T.G.; Kinnaird, M.F.; Wibisono, H.T. 2003. Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Animal Conservation, 6: 131-139. ; Srbek-Araujo and Chiarello 2013Srbek-Araujo, A.C.; Chiarello, A.G. 2013. Influence of camera-trap sampling design on mammal species capture rates and community structures in southeastern Brazil. Biota Neotropica, 13: 51-62. ). We used the detection frequency (or catch per unit effort) to calculate a relative abundance index. The index was calculated as the number of independent photographs per species/camera trap-days * 100. Measuring abundance in this way allowed us to evaluate the commonness or rarity of each species. Given the difficulty in identifying individuals on camera-trap images, this measure does not represent the abundance or density sensu stricto (Magurran and Henderson 2011Magurran, A.; Henderson, P.A. 2011. Commonness and rarity. In: Magurran, A.E.; McGill, B. (Ed.). Biological Diversity: Frontiers in Measurement and Assessment. Oxford University Press, New York, p.97-103.; O’Brien 2011O’Brien, T.G. 2011. Abundance, density and relative abundance: A conceptual framework. In: O’Connell, A.F.; Nichols, J.D.; Karanth, K.U. (Ed.). Camera Traps in Animal Ecology. Springer, Tokyo, p.71-96. ). We classified the recorded species in a dichotomous category as core and satellite species sensuHanski (1982Hanski, I. 1982. Dynamics of regional distribution: the core and satellite species hypothesis. Oikos, 38: 210-221.). Core species include the most abundant species that were present in most of the rapid inventories (~70% of all inventories). All species that occurred at only a few sites and in low abundance) were classified as satellite species.
Sampling completeness and species richness estimation
Sampling completeness was measured at each locality based on the number of independent events using species-accumulation curves and the evaluation of four non-parametric estimators for species richness: ICE, Chao 2, Jackknife 1 (Jack 1), Jackknife 2 (Jack 2) using Estimates 9.0 (Colwell 2013). The completeness of each survey was calculated based on the ratio between observed richness and the total estimated richness. To estimate the species richness and the sampling completeness of each surveyed locality, we used each independent event as a sampling unit (Willott 2001Willott, S.J. 2001. Species accumulation curves and the measure of sampling effort. Journal of Applied Ecology, 38: 484-486. ). We based our analyses on the incidence data and randomized the shuffling between samples with 1000 iterations to avoid the order of appearance in the estimation (Soberón and Llorente 1993Soberón, J.; Llorente, J. 1993. The use of species accumulation functions for the prediction of species richness. Conservation Biology, 7: 480-488. ).
To determine the least biased method to estimate species richness, we compared the four richness estimators by looking at: 1) the accuracy of the richness estimation; 2) the best fit between observed and estimated richness; and 3) which estimator is least affected by the influence of singletons (species with only one independent record in the sample). We assessed the accuracy of the estimators using the data for the locality of Tiquié (Figure 1). For this locality, we had long-term data for a whole-year survey (2111 camera trap-days) which we assumed to be a representative inventory of MLM as defined in this study. We compared the species richness resulting from the long-term data with the richness obtained with each estimator for the first sampling occasion at Tiquié (175 camera trap-days; Table 1). We used Pearson correlation to test the best fit of estimated and observed richness in all rapid assessments. To determine the influence of singletons, we performed a Pearson correlation between the estimated richness and the singletons. The significance level was p < 0.05.
Minimum sampling effort
We assessed the minimum sampling effort to reach a representative species richness for MLM in camera-trap assessments in the Colombian Amazon. We employed general linear models (GLM) using Poisson distribution (counts) to assess the effect of the sampling effort on the richness estimators by considering (a) the number of camera traps used in each survey; (b) the number of sampling stations (i.e., the number of different camera placements used during each survey); (c) the total sampling effort (the number of camera trap-days) at each locality; and (d) a null model. We determined the most parsimonious model based on Akaike’s information criterion, corrected for small sample size (AICc; Burnham et al. 2011). We selected the most plausible model using a Δ AICc < 2 and the higher relative contribution according to the Akaike Weight (ωi). To assess the explanatory power of the model and corroborate that a model was better than the null model, we used the p-values of chi-squared tests considering the model’s deviance and residual degrees of freedom in the former, and the mathematical difference between deviances of both models and one degree of freedom for the latter. Finally, we calculated the variability explained by the best model as follows: 100 x [(null deviance - residual deviance) / null deviance]. We used the R package Glmulti (Calcagno and de Mazancourt 2010Calcagno, V.; de Mazancourt, C. 2010. Glmulti: An R package for easy automated model selection with (generalized) linear models. Journal of Statistical Software, 34: 1-29. ) for GLM calculations. We used the predict function in R software (R Core Team 2020R Core Team. 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing. ( (https://www.R-project.org/ ). Accessed on 20 Mar 2020.
https://www.R-project.org/...
) to calculate the minimum sampling effort necessary to reach the species richness of the long-term data at Tiquié (which was assumed to be a more complete survey of MLM by camera-trapping) according to the predicted species richness by the best estimator.
RESULTS
Camera-trap sampling data
Overall, 26 species of MLM were recorded in the 16 rapid assessments and the long-term survey at Tiquié, distributed in seven orders, 12 families, and 22 genera (Supplementary Material, Table S1). Five species were recorded most frequently and were present at most localities, being classified as core species: Dasypus novemcinctus Linnaeus, 1758 (23.1% of all independent records), Cuniculus paca (Linnaeus, 1766) (17,2%), Tapirus terrestris (Linnaeus, 1758) (15.2%), Dasyprocta fuliginosa Wagler, 1832 (14.2%), and Pecari tajacu (Linnaeus, 1758) (7.0%) (Figure 2). The remaining species were recorded in few localities and in low abundance and were classified as satellite species. The rarest satellite species, with only one record in one locality, were Atelocynus microtis (Sclater, 1883) and Hydrochoerus hydrochaeris (Linnaeus, 1766) in lowland localities, and Panthera onca and Nasuella olivacea (Gray, 1865) in Amazon foothill localities.

Relative abundance of the species of medium and large mammals recorded in camera-trap rapid assessments in 16 localities in the Colombian Amazon. N species = number of species recorded in each locality (upper horizontal line). Colors indicate the most abundant species in all localities. This figure is in color in the electronic version.
Sampling completeness and species richness estimation
The observed species richness per locality varied from five to 13 species. The highest richness was observed in Matavén (13 species), followed by Río Yarí and Lindosa (12 species each). Lowest richness was observed in Buenos Aires and Carrizal (five species each) (Table 2).
Recorded (N species) and estimated species richness according to four non-parametric estimators [ICE, Chao 2, Jackknife 1 (Jack 1), and Jackknife 2 (Jack 2)] for camera-trap rapid assessments at 16 localities in the Colombian Amazon. The percentage of completeness for all the estimators is also indicated.
The species accumulation curves showed an increasing, non-asymptotical trend, indicating that a greater number of species was expected in all localities (Figure 3a). Sampling completeness varied among sites (ICE = 44 - 95%; Chao 2 = 50 - 100%; Jackknife 1 = 67 - 88%, Jackknife 2 = 50 - 91%) (Table 3). Estimated richness also varied among estimators (ICE = 5 - 22 species; Chao 2 = 5 - 20 species; Jackknife 1 = 6 - 17 species; Jackknife 2 = 7 - 19 species) (Table 2).

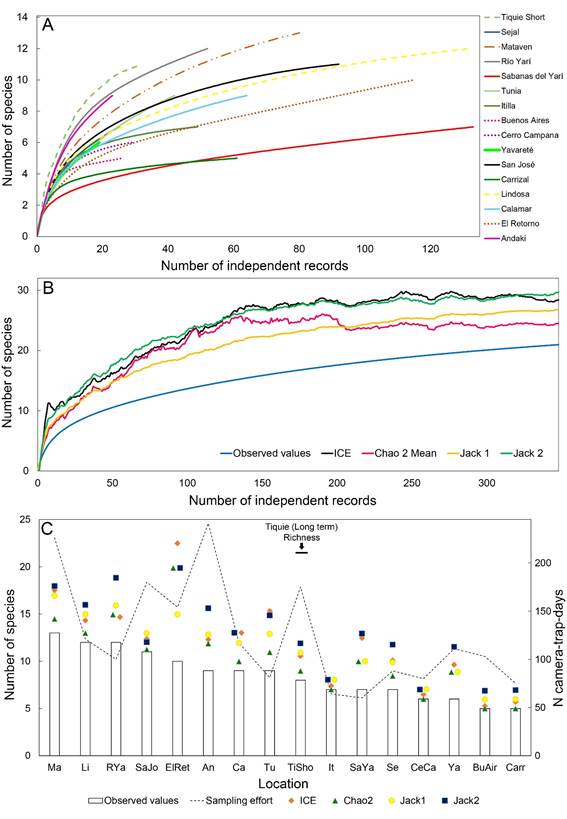
Assessment of species richness of medium and large mammals through camera-trap surveys in 16 localities in the Colombian Amazon. A - Species accumulation curves of observed independent records per locality; B - Species accumulation curves of observed and estimated values for a long-term one-year survey in Tiquié using four non-parametric estimators (ICE, Chao 2, Jackknife 1, and Jackknife 2); C - Observed and estimated species richness, and sampling effort per locality. Ma: Matavén, Li; Lindosa, RYa: Río Yarí, SaJo: San José, ElRet: El Retorno, An: Andakí, Ca: Calamar, Tu: Tunia, TiSho: Tiquié short, It: Itilla, SaYa: Sabanas del Yarí, Se: Sejal, CeCa: Cerro Campana, Ya: Yavaraté, BuAir: Buenos Aires, Carr: Carrizal.
Correlation between observed and estimated species richness, and between singletons and species richness estimates, for each of four non-parametric estimators using data of camera-trap rapid assessments in 16 localities in the Colombian Amazon. All correlations were statistically significant (p<0.05).
The long-term one-year survey at Tiquié recorded 21 species of MLM (Table 2; Supplementary Material, Table S1). Despite the longer sampling effort, all estimators for this dataset showed a similar, non-asymptotic trend in the accumulation curve, predicting that the actual species richness at Tiquié should be of 27 to 30 species (Figure 3b). This difference between estimated and observed richness may be owed to the presence of species with only one record in the survey. The comparison between the long-term and short-term survey data at Tiquié (Tables 1 and 2) showed that all estimators underestimated the species richness of the long-term survey (Figure 3c). The richness estimation at Tiquié with ICE, Jackknife 1, and Jackknife 2 was higher than with Chao 2 and, therefore, could better predict the species richness based on the short-term data (Figure 3c).
The values for species richness using the four estimators varied among the rapid assessments. Chao 2 generally had the lowest values among the four estimators. Jackknife 1 values were intermediate and showed the least variation among localities, while ICE values fluctuated the most, and Jackknife 2 had the highest estimated richness values in most cases, showing a higher influence of singletons (Figure 3c). In addition to resulting in the more stable estimates, Jackknife 1 had the highest correlation between estimated and observed richness (r = 0.96), and the lowest correlation with singletons (r = 0.82) (Table 4), thus was the estimator less influenced by singletons. ICE had the lowest correlation between estimated and observed richness (r = 0.77) and the highest correlation with singletons (r = 0.89). Chao 2 and Jackknife 2 has similar correlation values for observed and estimated richness (r = 0.81 and 0.83, respectively), both lower than the values for Jackknife1, and higher correlation with singletons than Jackknife 1 (r = 0.86 and 0.93, respectively), showing a higher bias due to the presence of singletons (Table 3).
Comparison of all GLM models using sampling variables to explain species richness of medium and large mammals in 16 localities of the Colombian Amazon using the Jackknife 1 estimator as dependent variable. Explanatory variables: N cameras = number of cameras; N stations = number of sites where cameras were placed during the sampling window; N camera trap-days = total sampling effort (sum of the days that each camera was active during the sampling window). nPars = number of parameters; AICc = Akaike information Criterion corrected for small samples; ΔAICc = Delta Akaike Information Criterion corrected for small samples; ωiAIC = Weight of Akaike Information Criterion for each model.
Minimum sampling effort
The GLM model that best explained the variation in species richness was based on total sampling effort (number of camera trap-days; AICc = 83.82, ΔAICc to next model = 1.28, ωi = 0.49) (Table 4). The model explained 39% of the data variance in the data, had a strong explanatory power (p-value = 0.6098) and differed significantly form the null model (p-value = 0.0247). The model showed a positive effect (the number of recorded species increased with the number of camera trap-days). The second-best model had ΔAICc < 2 (Table 4), and was discarded as it included the number of stations and the number camera trap-days, but only the latter had a significant effect in the model.
According to the predict function of the model and the species-richness estimates by the Jacknife 1 estimator, a minimum sampling effort of 350 camera trap-days would be necessary to record the number of species of the long-term survey at Tiquié (Figure 4).

Predictive model of minimum sampling effort to estimate a representative species richness of medium- and large-sized mammals in the Colombian Amazon based on camera-trap rapid assessment data for 16 localities. The model shows that at least 350 camera trap-days are necessary to record 21 species (the long-term record at Tiquié).
DISCUSSION
Our results showed that the local species richness of medium and large terrestrial mammals in the northwestern Amazon in Colombia is still highly underestimated. We used an extensive dataset of local camera-trap rapid assessments which, individually, did not have enough sampling effort for an effective estimation of the species richness per locality. Yet, although completeness varied among localities, the assessments generally recorded the core species and some satellite species in each locality. Considering its climatic and ecosystemic heterogeneity, a higher diversity of MLM is expected for the Colombian Amazon beyond that recorded in our surveys and in some published inventories of MLM for the northwestern and southernmost portions of the region (e.g., Negret et al. 2015Negret, P.J.; Garzón, O.; Stevenson, P. 2015. First preliminary inventory of non-flying mammals of the Alto Fragua Indi-Wasi National Park, Colombia. Mammalogy Notes, 2: 32-34. ; Payán and Escudero-Páez 2015Payán, E.; Escudero-Páez, S. 2015. Densidad de jaguares (Panthera onca) y abundancia de grandes mamíferos terrestres en un área no protegida del Amazonas colombiano. In: Payán, E.; Lasso, C.A.; Castaño, C. (Ed.). Conservación de Grandes Vertebrados en Areas no Protegidas de Colombia, Venezuela y Brasil. Serie Editorial Fauna Silvestre Neotropical. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH), Bogotá, p.225-242. ; Niño-Reyes and Velásquez-Valencia 2016Niño-Reyes, A.; Velásquez-Valencia, A. 2016. Diversidad y estado de conservación de la mastofauna terrestre del municipio de San Vicente del Caguán, Caquetá, Colombia. Biodiversidad Neotropical, 6: 152-161. ; Lizcano et al. 2019Lizcano, D.J.; Niño-Reyes, A.; Parra, J.P.; Bonell, W.; Garay, M.; Muñoz-Hernández, A.; Rodríguez-Álvarez, N. 2019. Mamíferos medianos y grandes/Large and medium-sized mammals. In: Pitman, N.; Salazar-Molano, A.; Samper-Samper, S.; Vriesendorp, C.; Vásquez-Cerón, A.; del Campo, Á.; et al. (Ed.). Colombia: Bajo Caguán-Caquetá. Rapid Biological and Social Inventories Report 30. Chicago Field Museum, Chicago, p.140-151. ; Atuesta-Dimian and Ganeden 2019Atuesta-Dimian, N.; Ganeden, J. 2019. Mamíferos medianos y grandes de las cuencas altas de los ríos Tunia, Itilla y Yarí. In: Fundación para la Conservación y el Desarrollo Sostenible FCDS (Ed.). Expediciones Científicas en Las Nuevas Areas del PNN Serranía de Chiribiquete. Fundación para la Conservación y el Desarrollo Sostenible. Bogotá, p.77- 94.; Atuesta-Dimian et al. 2020Atuesta-Dimian, N.; García-Villalba, J.; Morales-Martínez, D.M. 2020. Mamíferos medianos y grandes de la transición Andino-Amazónica del Parque Andakí, municipio de Belén de los Andaquíes, Caquetá, Colombia. Revista Colombia Amazónica, 12: 111-123.; Mena et al. 2020Mena, J.L.; Yagui, H.; Tejeda, V.; Cabrera, J.; Pacheco-Esquivel, J.; Rivero, J.; Pastor, P. 2020. Abundance of jaguars and occupancy of medium- and large-sized vertebrates in a transboundary conservation landscape in the northwestern Amazon. Global Ecology and Conservation, 23: e01079.). Some of the latter studies were also based on a small sampling effort of less than 200 camera trap-days (e.g., Negret et al. 2015, Atuesta-Dimian and Ganeden 2019; Atuesta-Dimian et al. 2020).
Singletons influenced the estimators of species richness, including the long-term one-year dataset of Tiquié. The frequency occurrence of medium and large-sized mammals depends on the biological and behavioral characteristics of the species, which are related to demographic stochasticity, vagility, habitat preference, and the spatial and temporal variation in risk perception related to human activity (Henski 1982; Magurran and Henderson 2003). Apex predators, for example, have large home ranges, small population sizes, and their presence can be affected by human activity making them difficult to record with a small sampling effort (Maffei et al. 2011Maffei, L.; Noss, A.J.; Silver, S.C.; Kelly, M.J. 2011. Abundance/density case study: Jaguars in the Americas. In: O’Connell, A.F.; Nichol, J.D.; Karanth, K.U. (Ed.). Camera Traps in Animal Ecology: Methods and Analyses. Springer, New York, p.119-144.; Figueroa 2013Figueroa, A. 2013. The ecology and conservation of jaguars (Panthera onca) in Central Belize: conservation status, diet, movement patterns, and habitat use. Doctoral thesis, University of Florida, USA. 163p. (https://ufdc.ufl.edu/UFE0045191/00001)
https://ufdc.ufl.edu/UFE0045191/00001...
). Therefore, rapid assessments might not be enough to detect these less abundant or rare species.
We recommend the use of the Jackknife 1 estimator in rapid assessments in the Colombian Amazon, based on its higher precision in the estimation of species richness of medium and large mammals in our samples, and the lower influence of singletons compared to the other tested estimators. Similarly, the Jackknife 1 has also been suggested as the best estimator in other rainforest surveys of MLM (Tobler et al. 2008Tobler, M.W.; Carrillo-Percastegui, S.E.; Leite, R.; Mares, R.; Powell, G. 2008. An evaluation of camera traps for inventorying large- and medium-sized terrestrial rainforest mammals. Animal Conservation, 11: 169-178. ). However, taking sampling bias issues and the stochasticity of data acquisition into account, it should be advisable to always test survey data with multiple estimators. The non-parametric estimators used in this study had different performance according to the nature of the samples, but empiric species richness assessments seldom take this into account (González-Oreja et al. 2010González-Oreja, J.A.; De la Fuente-Díaz-Ordaz, A.A.; Hernández-Santín, L.; Buzo-Franco, D.; Bonache-Regidor, C. 2010. Evaluación de estimadores no paramétricos de la riqueza de especies. Un ejemplo con aves en áreas verdes de la ciudad de Puebla, México. Animal Biodiversity and Conservation, 33: 31-45. ). Therefore, biological richness studies should evaluate the behavior of estimators in order to choose the most appropriate depending on the taxonomic group and ecosystem studied. Our results also corroborate Colwell and Coddington (1994Colwell, R.K.; Coddington, J.A. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society (Series B), 345: 101-118. ) in that rapid-assessment data should be calibrated with long-term data from comparable reference sites that use the same standardized sampling methods, if available. Multi-year long-term studies increase the accuracy of diversity estimates as they increase the probability of recording rare species.
The relative abundance of recorded species was heterogenous among our sampling localities. D. fuliginosa, C. paca, D. novemcinctus, P. tajacu, and T. terrestris were locally abundant in most localities, showing their ample distribution in the wider region and their status as core mammal species in Colombian Amazon forests, as is the case in other parts of the Amazon (e.g., Tobler et al. 2008Tobler, M.W.; Carrillo-Percastegui, S.E.; Leite, R.; Mares, R.; Powell, G. 2008. An evaluation of camera traps for inventorying large- and medium-sized terrestrial rainforest mammals. Animal Conservation, 11: 169-178. ; Blake and Mosquera 2014Blake, J.; Mosquera, D. 2014. Camera trapping on and off trails in lowland forest of eastern Ecuador: does location matter? Mastozoología Neotropical, 21: 17-26.; Scullion et al. 2021Scullion, J.J.; Fahrenholz, J.; Huaytalla, V.; Rengifo, E.M.; Lang, E. 2021. Mammal conservation in Amazonia’s protected areas: A case study of Peru’s Ichigkat Muja - Cordillera del Cóndor National Park. Global Ecology and Conservation, 26: e01451.). Our results showed that the rapid assessments were suitable to document the presence of core species and give some insight into their local abundance, and also the presence of some satellite species. Due to their high abundance and biomass, in general, the most common species have an important role in the functionality and stability of ecosystems (Smith and Knapp 2003Smith, M.D.; Knapp, A.K. 2003. Dominant species maintain ecosystem function with non-random species loss. Ecology Letters, 6: 509-517. ), and their abundance provides background information on the conservation status of the habitats in a locality. On the other hand, rare species, which are differentially underrepresented in rapid assessments, are most vulnerable to extinction (Purvis et al. 2000Purvis, A.; Agapow, P.M.; Gittleman, J.C.; Mace, G.M. 2000. Non-random extinction and the loss of evolutionary history. Science, 288: 328-330. ).
Colombian environmental policies, the limited funding of research institutions and environmental agencies, and the current need for rapid biodiversity assessments to take environmental decisions such as environmental licensing and the creation or enlargement of protected areas, require efficient and cost-effective studies that minimize the underestimation of biological diversity estimates (e.g., Gómez-Sandoval et al. 2017Gómez-Sandoval, P.A.; Mojica-Corzo, J.I.; Mejía-Egas, O. 2017. Trazabilidad de los registros de mamíferos en el marco del otorgamiento de licencias ambientales para proyectos de hidrocarburos en Colombia. Revista de la Academia Colombiana de Ciencias Exactas Físicas y Naturales, 41: 51-58. ). In most cases, short-term surveys are the only source of information that the environmental authorities have for decision making, therefore, they are crucial for the conservation and management of Colombian biodiversity, with the assessment of new conservation areas in the Orinoco Llanos (Mora-Fernández and Rodríguez-Posada 2017Mora-Fernández, C.; Rodríguez-Posada, M.E. 2017. Fauna y Flora del Casanare de la Montaña a la Sabana: Una Visión Integral del Territorio. Gobernación de Casanare, Yopal, 304p. ) and the extension of the Chiribiquete National Park in the Colombian Amazon (Atuesta-Dimian and Ganeden 2019Atuesta-Dimian, N.; Ganeden, J. 2019. Mamíferos medianos y grandes de las cuencas altas de los ríos Tunia, Itilla y Yarí. In: Fundación para la Conservación y el Desarrollo Sostenible FCDS (Ed.). Expediciones Científicas en Las Nuevas Areas del PNN Serranía de Chiribiquete. Fundación para la Conservación y el Desarrollo Sostenible. Bogotá, p.77- 94.) as recent examples. To strengthen the results of rapid assessments, we recommend a minimum sampling effort of 350 camera trap-days to reach a representative sample of the most common medium and large terrestrial mammals in Amazon-forest sites. We strongly suggest maximizing the sampling time because it increases the probability of capturing a higher number of rare species, allowing the calculation of detection probabilities and a more robust estimation of species richness (Kays et al. 2020Kays, R.; Arbogast, B.S.; Baker-Whatton, M.; Beirne, C.; Boone, H.M.; Bowler, M.; et al. 2020. An empirical evaluation of camera trap study design: How many, how long and when?. Methods in Ecology and Evolution, 11: 700-713. ).
It should be stressed that the results of short-term surveys do not measure the actual species richness of a locality. The precise and accurate estimation of ecological parameters such as species richness and community structure, as well as population abundance and density, and species occupancy require long-term and large-scale sampling efforts (e.g., Tobler et al. 2008Tobler, M.W.; Carrillo-Percastegui, S.E.; Leite, R.; Mares, R.; Powell, G. 2008. An evaluation of camera traps for inventorying large- and medium-sized terrestrial rainforest mammals. Animal Conservation, 11: 169-178. , 2013Tobler, M.W.; Carrillo-Percastegui, S.E.; Zuñiga, A., Powell, G. 2013. High jaguar densities and large population sizes in the core habitat of the southwestern Amazon. Biological Conservation, 159: 375-381.; Kays et al. 2020Kays, R.; Arbogast, B.S.; Baker-Whatton, M.; Beirne, C.; Boone, H.M.; Bowler, M.; et al. 2020. An empirical evaluation of camera trap study design: How many, how long and when?. Methods in Ecology and Evolution, 11: 700-713. ; Mena et al. 2020Mena, J.L.; Yagui, H.; Tejeda, V.; Cabrera, J.; Pacheco-Esquivel, J.; Rivero, J.; Pastor, P. 2020. Abundance of jaguars and occupancy of medium- and large-sized vertebrates in a transboundary conservation landscape in the northwestern Amazon. Global Ecology and Conservation, 23: e01079.). A more accurate estimation of species richness and occupancy should be based on samplings longer than three weeks with more than 25 camera sites for common species, and more than 150 camera sites for rare species (Kays et al. 2020). Short-term surveys are necessary for environmental management in the Colombian Amazon and produce valuable information about biodiversity, but these data must be understood in the light of the representativeness of the sampling. More accurate estimates can be obtained using complementary methods to camera trapping, such as interviews with local inhabitants (Voss and Emmons 1996Voss, R.S.; Emmons, L.H. 1996. Mammalian diversity in Neotropical Lowland Rainforest: A preliminary assessment. Bulletin of the American Museum of Natural History, 230: 1-115.; López-Arévalo et al. 2021López-Arévalo, H.F.; Morales-Martínez, D.M.; Mora-Beltrán, C.; Calderón-Capote, M. C.; Cárdenas-González, C.; Atuesta-Dimian, N.; Melo, M. J.; Ramírez, W. 2021. Mamíferos (Mammalia) de San José del Guaviare, Colombia. Biota Colombiana, 22: 127-146.), surveys of animal tracks or signs (Fragoso et al. 2019Fragoso, J.; Gonçalves, F.; Oliveira, L.; Overman, H.; Levi, T.; Silvius, K. M. 2019. Visual encounters on line transect surveys under-detect carnivore species: Implications for assessing distribution and conservation status. PloS ONE, 14: e0223922. ), analysis of hunting records (Voss and Emmons 1996), and the prospection of remains of consumed animals that are usually available in indigenous and settler communities (Osorno et al. 2014Osorno, M.; Atuesta-Dimian, N.; Jaramillo, L.F.; Sua, S.; Barona, A.; Roncancio, N. 2014. La Despensa del Tiquié: Diagnóstico y Manejo Comunitario de La Fauna de Consumo en la Guayana Colombiana. Instituto Amazónico de Investigaciones Científicas “SINCHI”, Bogotá, 162p. ). It is also important to stress that, although useful, rapid assessments are not enough to assess long-term effects of environmental transformation and tendencies of biodiversity loss. Therefore, besides determining minimum sampling standards that maximize the effectiveness of short-term assessments, it is crucial to also invest in standardized long-term surveys that allow evaluating processes at broader scales and making environmental decisions with solid scientific bases.
CONCLUSIONS
The local species richness of medium and large terrestrial mammals measured in 16 rapid assessments in the northwestern Amazon in Colombia was highly underestimated. Notwithstanding, rapid assessments provide richness information of good representativeness on core species and some satellite species in each locality. Our results indicate that the Jackknife-1 non-parametric estimator is the most accurate to estimate species richness in these kind of surveys in this region, due to its accurate estimation of richness and the low influence of singletons. We recommend a minimum sampling effort of 350 camera trap-days to achieve a representative estimate of species richness of medium and large terrestrial mammals in local surveys in the region, however precise ecological and populational assessments require larger sampling efforts.
ACKNOWLEDGMENTS
We are grateful mainly to the local inhabitants, researchers, and owners of the land where each survey was conducted. DMM thanks Grupo de Fauna of Programa de Ecosistemas y Recursos Naturales of Instituto Amazónico de Investigaciones Científicas SINCHI, for allowing us to use the data of rapid assessments in our analyses. Data collection of these rapid assesment was financed by SINCHI-BPIN projects “Apoyo para el Fortalecimiento de la Capacidad de Investigación del Instituto Amazónico de Investigaciones Científica Sinchi 2015”, “Conservación y aprovechamiento sostenible de la diversidad biológica, socioeconómica y cultural de la Amazonia colombiana 2016-2017”, and “Investigación en conservación y aprovechamiento sostenible de la diversidad biológica, socioeconómica y cultural de la Amazonia colombiana 2018-2021”, as well as SINCHI-Colciencias Project “Diagnóstico, evaluación y manejo comunitario de la fauna silvestre en la zona del río Tiquié, departamento del Vaupés, amazonia colombiana”, Colombia-Bio Projects “Expedición Colombia-Bio a la biodiversidad en la transición andino-amazónica del departamento del Caquetá” and “Expedición Colombia BIO Río Apaporis 2018“, SINCHI-FCDS project “Caracterización Biológica para la Ampliación del PNN Serranía de Chiribiquete”, and SINCHI-SGR Project “Investigación en relictos de bosque como estrategia para generar bienes y servicios ambientales en el Departamento del Guaviare”. Finally, we would like to thank Catalina Mora Fernandez for mammal illustrations in Figure 2, and Sofia Rojas for the comments to an early version of the manuscript. We thank the anonymous reviewers and the editor for their careful reading of our manuscript and their insightful comments and suggestions to improve the final version.
REFERENCES
- Arévalo-Sandi, A.; Bobrowiec, P.E.D.; Rodríguez, V.J.U.; Norris, D. 2018. Diversity of terrestrial mammal seed dispersers along a lowland Amazon forest regrowth gradient. PLoS ONE, 13: e0193752.
- Armenteras, D.; Rudas, G.; Rodriguez, N.; Sua, S.; Romero, M. 2006. Patterns and causes of deforestation in the Colombian Amazon. Ecological Indicators, 6: 353-368.
- Atuesta-Dimian, N.; Ganeden, J. 2019. Mamíferos medianos y grandes de las cuencas altas de los ríos Tunia, Itilla y Yarí. In: Fundación para la Conservación y el Desarrollo Sostenible FCDS (Ed.). Expediciones Científicas en Las Nuevas Areas del PNN Serranía de Chiribiquete Fundación para la Conservación y el Desarrollo Sostenible. Bogotá, p.77- 94.
- Atuesta-Dimian, N.; García-Villalba, J.; Morales-Martínez, D.M. 2020. Mamíferos medianos y grandes de la transición Andino-Amazónica del Parque Andakí, municipio de Belén de los Andaquíes, Caquetá, Colombia. Revista Colombia Amazónica, 12: 111-123.
- Benítez-López, A.; Santini, L.; Schipper, A.M.; Busana, M.; Huijbregts, M.A.J. 2019. Intact but empty forests? Patterns of hunting-induced mammal defaunation in the tropics. PLoS Biology, 17: e3000247.
- Blake, J.; Mosquera, D. 2014. Camera trapping on and off trails in lowland forest of eastern Ecuador: does location matter? Mastozoología Neotropical, 21: 17-26.
- Boron, V.; Deere, N.J.; Xofis, P.; Link, A.; Quiñones-Guerrero, A.; Payan, E.; Tzanopoulos, J. 2019. Richness, diversity, and factors influencing occupancy of mammal communities across human-modified landscapes in Colombia. Biological Conservation, 232: 108-116.
- Calcagno, V.; de Mazancourt, C. 2010. Glmulti: An R package for easy automated model selection with (generalized) linear models. Journal of Statistical Software, 34: 1-29.
- Clerici, N.; Armenteras, D.; Kareiva, P.; Botero, R.; Ramírez-Delgado, J.P.; Forero-Medina, G.; et al 2020. Deforestation in Colombian protected areas increased during post-conflict periods. Scientific Reports, 10: 4971.
- Colwell, R.K.; Coddington, J.A. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society (Series B), 345: 101-118.
- Figueroa, A. 2013. The ecology and conservation of jaguars (Panthera onca) in Central Belize: conservation status, diet, movement patterns, and habitat use. Doctoral thesis, University of Florida, USA. 163p. (https://ufdc.ufl.edu/UFE0045191/00001)
» https://ufdc.ufl.edu/UFE0045191/00001 - Fragoso, J.; Gonçalves, F.; Oliveira, L.; Overman, H.; Levi, T.; Silvius, K. M. 2019. Visual encounters on line transect surveys under-detect carnivore species: Implications for assessing distribution and conservation status. PloS ONE, 14: e0223922.
- Gómez-Sandoval, P.A.; Mojica-Corzo, J.I.; Mejía-Egas, O. 2017. Trazabilidad de los registros de mamíferos en el marco del otorgamiento de licencias ambientales para proyectos de hidrocarburos en Colombia. Revista de la Academia Colombiana de Ciencias Exactas Físicas y Naturales, 41: 51-58.
- González-Oreja, J.A.; De la Fuente-Díaz-Ordaz, A.A.; Hernández-Santín, L.; Buzo-Franco, D.; Bonache-Regidor, C. 2010. Evaluación de estimadores no paramétricos de la riqueza de especies. Un ejemplo con aves en áreas verdes de la ciudad de Puebla, México. Animal Biodiversity and Conservation, 33: 31-45.
- Hanski, I. 1982. Dynamics of regional distribution: the core and satellite species hypothesis. Oikos, 38: 210-221.
- Hernández-Camacho, J.I.; Guerra, A.H.; Quijano, R.O.; Walschburger, T. 1992. Unidades biogeográficas de Colombia. In: Halffter, G (Ed.). La Diversidad Biológica de Iberoamérica I Acta Zoologica Mexicana, Instituto de Ecología, Xalapa, p.105-153.
- IDEAM. 2020. Instituto de Hidrología, Meteorología y Estudios Ambientales. Boletín de detección temprana de deforestación, # 25. ( (http://smbyc.ideam.gov.co/MonitoreoBC-WEB/pub/alertasDeforestacion.jsp?0.49368119561984636 ). Accessed on 20 Mar 2020.
» http://smbyc.ideam.gov.co/MonitoreoBC-WEB/pub/alertasDeforestacion.jsp?0.49368119561984636 - Kays, R.; Arbogast, B.S.; Baker-Whatton, M.; Beirne, C.; Boone, H.M.; Bowler, M.; et al 2020. An empirical evaluation of camera trap study design: How many, how long and when?. Methods in Ecology and Evolution, 11: 700-713.
- Lizcano, D.J.; Niño-Reyes, A.; Parra, J.P.; Bonell, W.; Garay, M.; Muñoz-Hernández, A.; Rodríguez-Álvarez, N. 2019. Mamíferos medianos y grandes/Large and medium-sized mammals. In: Pitman, N.; Salazar-Molano, A.; Samper-Samper, S.; Vriesendorp, C.; Vásquez-Cerón, A.; del Campo, Á.; et al (Ed.). Colombia: Bajo Caguán-Caquetá. Rapid Biological and Social Inventories Report 30 Chicago Field Museum, Chicago, p.140-151.
- López-Arévalo, H.F.; Morales-Martínez, D.M.; Mora-Beltrán, C.; Calderón-Capote, M. C.; Cárdenas-González, C.; Atuesta-Dimian, N.; Melo, M. J.; Ramírez, W. 2021. Mamíferos (Mammalia) de San José del Guaviare, Colombia. Biota Colombiana, 22: 127-146.
- Maffei, L.; Noss, A.J.; Silver, S.C.; Kelly, M.J. 2011. Abundance/density case study: Jaguars in the Americas. In: O’Connell, A.F.; Nichol, J.D.; Karanth, K.U. (Ed.). Camera Traps in Animal Ecology: Methods and Analyses Springer, New York, p.119-144.
- Magurran, A.; Henderson, P.A. 2011. Commonness and rarity. In: Magurran, A.E.; McGill, B. (Ed.). Biological Diversity: Frontiers in Measurement and Assessment Oxford University Press, New York, p.97-103.
- Mena, J.L.; Yagui, H.; Tejeda, V.; Cabrera, J.; Pacheco-Esquivel, J.; Rivero, J.; Pastor, P. 2020. Abundance of jaguars and occupancy of medium- and large-sized vertebrates in a transboundary conservation landscape in the northwestern Amazon. Global Ecology and Conservation, 23: e01079.
- Mollentze, N.; Streicker, D.G. 2020. Viral zoonotic risk is homogenous among taxonomic orders of mammalian and avian reservoir hosts. Proceedings of the National Academy of Sciences, 117: 9423-9430.
- Montenegro, O.L. 2007. Mamíferos terrestres del sur de la Amazonia colombiana. In: Ruiz, S.L.; Sánchez, E.; Tabares, E.; Prieto, A.; Arias, J.C.; Gómez, R.; Castellanos, D.; García, P.; Rodríguez, L. (Ed.). Diversidad Biológica y Cultural del Sur de la Amazonia Colombiana Corpoamazonia, Instituto Humboldt, Instituto Sinchi, UAESPNN, Bogotá, p.134-141.
- Mora-Fernández, C.; Rodríguez-Posada, M.E. 2017. Fauna y Flora del Casanare de la Montaña a la Sabana: Una Visión Integral del Territorio Gobernación de Casanare, Yopal, 304p.
- Morrone, J.J. 2014. Biogeographical regionalization of the Neotropical region. Zootaxa, 3782: 1-110.
- Murcia, U.G.; Rodríguez, C.H.; Castro, W. 2007. Zonificación ambiental (ecológica y económica) en la Amazonía colombiana. Revista Colombia Amazónica, 1: 119-134.
- Negret, P.J.; Garzón, O.; Stevenson, P. 2015. First preliminary inventory of non-flying mammals of the Alto Fragua Indi-Wasi National Park, Colombia. Mammalogy Notes, 2: 32-34.
- Niño-Reyes, A.; Velásquez-Valencia, A. 2016. Diversidad y estado de conservación de la mastofauna terrestre del municipio de San Vicente del Caguán, Caquetá, Colombia. Biodiversidad Neotropical, 6: 152-161.
- O’Brien, T.G.; Kinnaird, M.F.; Wibisono, H.T. 2003. Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Animal Conservation, 6: 131-139.
- O’Brien, T.G. 2011. Abundance, density and relative abundance: A conceptual framework. In: O’Connell, A.F.; Nichols, J.D.; Karanth, K.U. (Ed.). Camera Traps in Animal Ecology Springer, Tokyo, p.71-96.
- Osorno, M.; Atuesta-Dimian, N.; Jaramillo, L.F.; Sua, S.; Barona, A.; Roncancio, N. 2014. La Despensa del Tiquié: Diagnóstico y Manejo Comunitario de La Fauna de Consumo en la Guayana Colombiana Instituto Amazónico de Investigaciones Científicas “SINCHI”, Bogotá, 162p.
- Otavo, S.; Murcia, U. 2018. La Amazonía se quema: detección de áreas con mayor ocurrencia de incendios de vegetación como estrategia para la prevención y control. Colombia Amazónica, 11: 59-72.
- Payán, E.; Escudero-Páez, S. 2015. Densidad de jaguares (Panthera onca) y abundancia de grandes mamíferos terrestres en un área no protegida del Amazonas colombiano. In: Payán, E.; Lasso, C.A.; Castaño, C. (Ed.). Conservación de Grandes Vertebrados en Areas no Protegidas de Colombia, Venezuela y Brasil Serie Editorial Fauna Silvestre Neotropical. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH), Bogotá, p.225-242.
- Pineda-López, R. 2019. Estimadores de la riqueza de especies. In: Moreno, C.E. (Ed.). La Biodiversidad en Un Mundo Cambiante: Fundamentos Teóricos y Metodológicos para Su Estudio Universidad Autónoma del Estado de Hidalgo, Libermex, Ciudad de México, p.159-174.
- Purvis, A.; Agapow, P.M.; Gittleman, J.C.; Mace, G.M. 2000. Non-random extinction and the loss of evolutionary history. Science, 288: 328-330.
- R Core Team. 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing. ( (https://www.R-project.org/ ). Accessed on 20 Mar 2020.
» https://www.R-project.org/ - Ramírez-Chaves, H.E.; Suárez-Castro, A.F.; Sociedad Colombiana de Mastozoología. 2019. Mamíferos de Colombia. ( (https://www.gbif.org/es/dataset/e8b9ed9b-f715-4eac-ae24-772fbf40d7ae ). Accessed on 30 Aug 2019.
» https://www.gbif.org/es/dataset/e8b9ed9b-f715-4eac-ae24-772fbf40d7ae - Renó, V.; Novo, E.; Escada, M. 2016. Forest fragmentation in the lower Amazon Floodplain: implications for biodiversity and ecosystem service provision to riverine populations. Remote Sensing, 8: 1-26.
- Rodríguez-Mahecha, J.V.; Alberico, M.; Trujillo, F.; Jorgenson, J. 2006. Libro Rojo de los Mamíferos de Colombia. Serie Libros Rojos de Especies Amenazadas de Colombia Conservación Internacional Colombia, Ministerio de Medio Ambiente, Vivienda y Desarrollo Territorial. Bogotá, 433p.
- Romero-Muñoz, A.; Benítez-López, A.; Zurell, D.; Baumann, M.; Camino, M.; Decarre, J.; et al 2020. Increasing synergistic effects of habitat destruction and hunting on mammals over three decades in the Gran Chaco. Ecography, 43: 954-966.
- Rudas, A. 2009. Unidades ecogeográficas y su relación con la diversidad vegetal de la Amazonia colombiana. Doctoral thesis, Universidad Nacional de Colombia, Colombia. 147p. (https://repositorio.unal.edu.co/handle/unal/11104).
» https://repositorio.unal.edu.co/handle/unal/11104 - Schöngart, J. 2008. Growth-Oriented Logging (GOL): A new concept towards sustainable forest management in Central Amazonian várzea floodplains. Forest Ecology and Management, 256: 46-58.
- Scullion, J.J.; Fahrenholz, J.; Huaytalla, V.; Rengifo, E.M.; Lang, E. 2021. Mammal conservation in Amazonia’s protected areas: A case study of Peru’s Ichigkat Muja - Cordillera del Cóndor National Park. Global Ecology and Conservation, 26: e01451.
- Smith, M.D.; Knapp, A.K. 2003. Dominant species maintain ecosystem function with non-random species loss. Ecology Letters, 6: 509-517.
- Soberón, J.; Llorente, J. 1993. The use of species accumulation functions for the prediction of species richness. Conservation Biology, 7: 480-488.
- Srbek-Araujo, A.C.; Chiarello, A.G. 2013. Influence of camera-trap sampling design on mammal species capture rates and community structures in southeastern Brazil. Biota Neotropica, 13: 51-62.
- Stevenson, P.R.; Suescún, M.; Quiñonez, M.J. 2004. Characterization of forest types at the CIEM, Tinigua Park, Colombia. Field Studies of Fauna and Flora La Macarena Colombia, 14: 1-20.
- Teixeira-Santos, J.; Ribeiro, A.C.; Wiig, Ø.; Pinto, N.S.; Cantanhêde, L.G.; Sena, L.; Mendes-Oliveira, A.C. 2020. Environmental factors influencing the abundance of four species of threatened mammals in degraded habitats in the eastern Brazilian Amazon. PLoS ONE, 15: e0229459.
- Trujillo, F.; Mosquera-Guerra, F.; Diaz-Pulido, A.; Carvajal-Castro, J.D.; Mantilla-Meluk, H. 2019. Mamíferos del Escudo Guayanés de Colombia. In: Lasso, C.A.; Señaris, J.C. (Ed.). Fauna Silvestre del Escudo Guayanés (Colombia-Venezuela) Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá, p.441-380.
- Tobler, M.W.; Carrillo-Percastegui, S.E.; Leite, R.; Mares, R.; Powell, G. 2008. An evaluation of camera traps for inventorying large- and medium-sized terrestrial rainforest mammals. Animal Conservation, 11: 169-178.
- Tobler, M.W.; Carrillo-Percastegui, S.E.; Zuñiga, A., Powell, G. 2013. High jaguar densities and large population sizes in the core habitat of the southwestern Amazon. Biological Conservation, 159: 375-381.
- Willott, S.J. 2001. Species accumulation curves and the measure of sampling effort. Journal of Applied Ecology, 38: 484-486.
- Whitmore, T.C. 1997. Tropical forest disturbance, disappearance, and species loss. In: Laurance, W.F.; Bierregaard, R.O. (Ed.). Tropical Forest Remnans: Ecology. Management and Conservation of Fragmented Communities The University of Chicago Press, Chicago, p.3-13.
- Whitmore, T.C.; Sayer, J.A. 1992. Tropical Deforestation and Species Extinction Chapman and Hall, London, 292p.
- Voss, R.S.; Emmons, L.H. 1996. Mammalian diversity in Neotropical Lowland Rainforest: A preliminary assessment. Bulletin of the American Museum of Natural History, 230: 1-115.
-
CITE AS:
Morales-Martínez, D.M.; Atuesta-Dimian, N.; Martínez-Medina, D.; Gutiérrez-Sanabria, D.R.; Rodríguez-Posada, M.E. 2021. Completeness of rapid assessments of medium and large mammal diversity in the northwestern Amazon in Colombia. Acta Amazonica 51: 224-233.
SUPPLEMENTARY MATERIAL
(only available in the electronic version)
Morales-Martínez et al. Completeness of rapid assessments of medium and large mammal diversity in the northwestern Amazon in Colombia
Relative abundance index for 26 medium and large terrestrial mammals with average body weight > 2 kg (excluding primarily arboreal species) recorded at 16 localities in the northwestern Amazon in Colombia through rapid camera-trapping assessments. The abundance index was calculated as the number of independent records/number of camera trap-days* 100. N species = number of species per locality. Tiquié (long term) refers to a long-term dataset (a year-long sampling) (see Table 1 and Material and Methods).
Edited by
ASSOCIATE EDITOR:
Publication Dates
-
Publication in this collection
20 Sept 2021 -
Date of issue
Jul-Sep 2021
History
-
Received
01 Apr 2021 -
Accepted
13 July 2021