Resumos
A apelina é um peptídeo recentemente descoberto e identificado como o ligando endógeno do receptor APJ. A apelina e o receptor APJ são expressos numa grande variedade de tecidos, tais como coração, cérebro, rins e pulmões, onde a sua interação pode ter importantes efeitos fisiopatológicos. Com efeito, a última década foi fértil no esclarecimento de possíveis papéis desempenhados pela apelina na fisiologia humana, nomeadamente como peptídeo regulador dos sistemas cardiovascular, hipotálamo-hipófisário, gastrointestinal e imunitário. Um possível envolvimento da apelina na patogênese de doenças com elevada prevalência e co-morbilidades, como a hipertensão arterial, a insuficiência cardíaca e o diabete melito tipo 2, perspectivam-na como um possível alvo terapêutico a explorar no futuro. Este trabalho fornece uma visão geral dos efeitos fisiológicos da apelina e apresenta o possível papel desse peptídeo na patogênese de várias doenças, associado a implicações terapêuticas que poderão vir a ser, assim, exploradas.
Peptídeos; peptídeos; apelina; hipertensão; baixo débito cardíaco; diabetes mellitus
Apelin is a recently discovered peptide, identified as an endogenous ligand of receptor APJ. Apelin and receptor APJ are expressed in a wide variety of tissues including heart, brain, kidneys and lungs. Their interaction may have relevant pathophysiologic effects in those tissues. In fact, the last decade has been rich in illustrating the possible roles played by apelin in human physiology, namely as a regulating peptide of cardiovascular, hypothalamus-hypophysis, gastrointestinal, and immune systems. The possible involvement of apelin in the pathogenesis of high prevalence conditions and comorbidities - such as hypertension, heart failure, and Diabetes Mellitus Type 2 (T2DM) - rank it as a likely therapeutic target to be investigated in the future. The present paper is an overview of apelin physiologic effects and presents the possible role played by this peptide in the pathogenesis of a number of conditions as well as the therapeutic implications that might, therefore, be investigated.
Peptides; peptides; apeline; hypertension; cardiac output, low; diabetes mellitus
ARTIGO DE REVISÃO
O sistema apelinérgico: papel na fisiologia e patologia humanas e potenciais aplicações terapêuticas
Ricardo Ladeiras-Lopes; João Ferreira-Martins; Adelino F. Leite-Moreira
Serviço de Fisiologia, Faculdade de Medicina da Universidade do Porto, Porto - Portugal
Correspondência Correspondência: Adelino F. Leite-Moreira Alameda Prof. Hernâni Monteiro 4200-319 Porto - Portugal E-mail: amoreira@med.up.pt
RESUMO
A apelina é um peptídeo recentemente descoberto e identificado como o ligando endógeno do receptor APJ. A apelina e o receptor APJ são expressos numa grande variedade de tecidos, tais como coração, cérebro, rins e pulmões, onde a sua interação pode ter importantes efeitos fisiopatológicos. Com efeito, a última década foi fértil no esclarecimento de possíveis papéis desempenhados pela apelina na fisiologia humana, nomeadamente como peptídeo regulador dos sistemas cardiovascular, hipotálamo-hipófisário, gastrointestinal e imunitário. Um possível envolvimento da apelina na patogênese de doenças com elevada prevalência e co-morbilidades, como a hipertensão arterial, a insuficiência cardíaca e o diabete melito tipo 2, perspectivam-na como um possível alvo terapêutico a explorar no futuro. Este trabalho fornece uma visão geral dos efeitos fisiológicos da apelina e apresenta o possível papel desse peptídeo na patogênese de várias doenças, associado a implicações terapêuticas que poderão vir a ser, assim, exploradas.
Palavras-chave: Peptídeos/fisiologia, peptídeos/efeitos de drogas, apelina, hipertensão, baixo débito cardíaco, diabetes mellitus.
Introdução
Em 1993, O'Dowd e cols.1 identificaram um gene que apresentava uma grande homologia com o gene que codifica o receptor de tipo 1 da angiotensina II (AT-1). Denominado APJ, esse receptor permaneceu "órfão" até 1998, quando Tatemoto e cols.2 descobriram um ligando endógeno seletivo para o mesmo, o qual foi denominado apelina ("apelin") para "APJ endogenous ligand". Os primeiros estudos demonstraram que a apelina e o seu receptor são expressos de uma forma praticamente ubiquitária, adquirindo especial destaque em órgãos como coração, cérebro, rins e pulmões3-6.
A distribuição do sistema apelinérgico por essa grande variedade de tecidos sugeriu que esse poderia desempenhar papéis importantes na fisiologia humana. Com efeito, a apelina está envolvida na regulação das funções cardiovascular, gastrointestinal e imunitária, na fisiologia óssea, na homeostasia de fluidos e no desenvolvimento embrionário do sistema cardiovascular7-9. Recentemente, foi igualmente demonstrado o envolvimento do sistema apelinérgico na patogênese de diversas doenças com uma elevada prevalência, tais como a hipertensão arterial, a insuficiência cardíaca (IC), a obesidade, a intolerância a glicose e o diabete melito tipo 2, bem como na infecção pelo VIH (vírus da imunodeficiência humana), no diabete insípido, na úlcera gástrica e na osteoporose7-9. Assim, é cada vez mais consensual que a modulação do binômio apelina/APJ poderá ser um possível alvo terapêutico a explorar no futuro10,11.
Nesse sentido, o objetivo deste trabalho é fazer uma revisão do sistema apelinérgico e proporcionar uma visão geral dos processos fisiológicos e patológicos em que esse está envolvido, para que uma melhor compreensão do binômio apelina/APJ permita a idealização de novas investigações a desenvolver e de possíveis aplicações clínicas no futuro.
O receptor APJ
Em 1993, O'Dowd e cols.1 caracterizaram um fragmento de 700 pares de bases, usando a tecnologia de reação em cadeia da polimerase (PCR), cuja análise detalhada revelou várias semelhanças com as seqüências gênicas dos domínios transmembranares dos receptores acoplados a proteínas G (GPCR). A proteína codificada possuía 380 aminoácidos e foi denominada APJ.
Cedo se verificou uma grande homologia entre o receptor APJ e o receptor AT-1: 115 aminoácidos (a.a.) (30%) no total e 86 a.a. (54%) nas regiões transmembranares1, bem como uma grande semelhança na expressão tecidual dos dois receptores3. Porém, a apelina, ligando endógeno do receptor APJ, não se liga ao receptor AT-112, nem a angiotensina II (AngII) ao APJ1,2.
Apesar de predominantemente localizado na membrana, o receptor APJ foi também encontrado no núcleo de células do cerebelo e do hipotálamo humanos, tendo sido identificado na terceira alça intracelular um sinal de localização nuclear13. Esses dados sugerem que esse receptor poderá ter um possível papel na regulação da transcrição de genes, tal como foi anteriormente demonstrado para a AngII10, aspecto que deverá ser alvo de investigação no futuro.
Após o estudo da distribuição do receptor APJ em ratos, camundongos e humanos, concluiu-se que esse é abundantemente expresso no sistema nervoso central (SNC) e em vários tecidos periféricos, merecendo especial destaque os pulmões e o coração1,3-5,14. No sistema cardiovascular, os diversos estudos já realizados demonstram que o receptor APJ está presente nas células endoteliais de pequenos vasos intramiocárdicos, renais, pulmonares e supra-renais, artérias coronárias, células endoteliais endocárdicas e células musculares lisas vasculares5. É interessante notar que a imuno-reatividade do APJ está disposta nos cardiomiócitos segundo um padrão estriado transversal, indicando a co-localização do receptor com os túbulos-T5.
Os peptídeos apelinérgicos
O receptor APJ permaneceu órfão até 1998, quando Tatemoto e cols.2 isolaram um peptídeo com 36 a.a. a partir de extratos de estômago de bovino, que denominaram apelina, o qual induzia a acidificação extracelular e a inibição da formação de AMPc numa linha celular de ovários de hamster chinês que expressava o receptor APJ. O gene que codifica a pré-proapelina, um pré-propeptídeo com 77 a.a., está localizado, no humano, no cromossoma Xq25-26.1, e possui 1.726 pares de bases com 3 exons. Curiosamente, demonstrou-se uma grande homologia entre diferentes espécies na seqüência protéica da pré-proapelina, nomeadamente nos 23 a.a. do terminal C12. Com efeito, o fragmento terminal 65-77 C dos peptídeos apelinérgicos é essencial para a ligação ao receptor APJ e para a sua atividade biológica, enquanto o terminal N desempenha um papel fundamental na modulação da interação ligando-receptor15.
Nos primeiros trabalhos sobre a apelina foram identificadas diferentes isoformas que se pensa poderem existir in vivo: apelina-36 (apelina42-77), apelina-17 (apelina61-77), apelina-13 (apelina65-77) e apelina-13 na forma piroglutaminada [(Pyr1)apelina-13], ou seja, com um resíduo glutamato no terminal N2. Enquanto as isoformas mais curtas parecem ser mais potentes na ação cardiovascular, a apelina-36 é mais eficiente no bloqueio da infecção pelo HIV em células APJ-positivas. Atualmente pensa-se que a apelina-36 funciona como um precursor com uma atividade biológica limitada até sofrer proteólise e modificações pós-translacionais para dar origem a peptídeos biologicamente mais ativos, predominantemente a (Pyr1)apelina-13, cuja piroglutaminação preserva a função biológica e impede a degradação enzimática8. Foi já descrito por Lee e cols.16 um possível antagonista do receptor APJ: por substituição da fenilalanina do terminal C da apelina-13 por um resíduo alanina cria-se um peptídeo que anula, de uma forma dependente da dose, a ação hipotensora da injeção intravenosa de apelina-13 em ratos.
Os primeiros estudos que caracterizaram a distribuição tecidual da apelina foram realizados em tecidos de ratos e evidenciaram a presença do seu pré-propeptídeo numa grande diversidade de tecidos3,4,6,12,17. Em humanos, o RNAm da pré-proapelina existe abundantemente no SNC e na placenta, bem como em quantidades mais moderadas no rim, no coração, nos pulmões e na glândula mamária. É de salientar o fato de a apelina biologicamente ativa se encontrar no miocárdio, no endotélio cardíaco e no endotélio dos grandes vasos e de pequenas veias e artérias7. Nas células endoteliais, a apelina não se localiza nos corpos de Weibel-Palade, vesículas resultantes da secreção peptídica endotelial induzida, sugerindo uma liberação constitutiva da apelina5.
A concentração de apelina circulante em humanos foi estimada entre 3 e 4 ng/mL através de um ensaio imunoenzimático18. Essa é uma concentração baixa quando comparada com aquela de outros hormônios circulantes, sendo, porém, comparável à de mediadores vasoativos derivados do endotélio, o que sugere uma liberação endotelial da apelina.
Atualmente, atribui-se à carboxipeptídase ECA2 (enzima de conversão da angiotensina de tipo 2) o papel de degradação da apelina, ao clivar a fenilalanina do terminal C dos peptídeos apelinérgicos com alta potência enzimática e eficácia19. Essa relação sugere, assim, uma grande importância da apelina na regulação cardiovascular, uma vez que a ECA2 participa no eixo renina-angiotensina-aldosterona8. De fato, a ECA2 também participa na degradação da AngII a angiotensina 1-7, a qual, ao contrário da AngII, parece desempenhar importantes funções de proteção cardíaca e renal20. Assim, a indução da ação dessa enzima poderá, no futuro, ser um alvo terapêutico nas principais doenças cardiovasculares.
Interação apelina/APJ
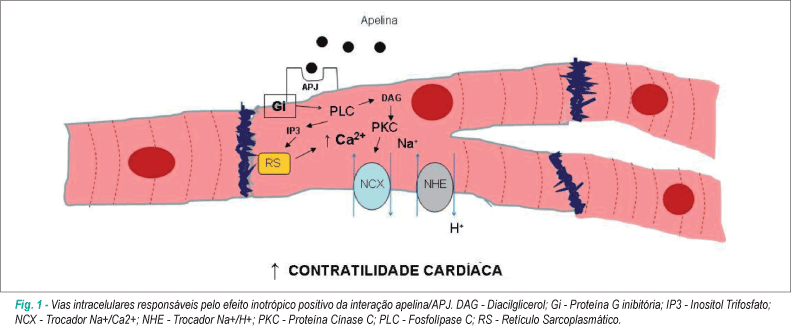
Uma das principais vias de transdução de sinal da apelina depende da interação com uma proteína Gi acoplada ao receptor APJ, independentemente da proteína Ras, sendo, contudo, dependente da proteína cínase C (PKC)7. Com efeito, se inibirmos a PKC, a fosfolípase C (PLC), o trocador Na+-H+ (NHE) e o trocador Na+-Ca2+ (NCX), os efeitos da apelina são significativamente reduzidos, nomeadamente no que diz respeito ao seu efeito inotrópico positivo21. É de salientar que a ativação da PKC resulta na fosforilação do NHE, o qual promove a alcalinização do interior da célula e a sensibilização dos miofilamentos ao Ca2+, bem como numa ativação do NCX, que passa a funcionar em modo reverso e promove, assim, a entrada de Ca2+ para a célula (fig. 1). Hashimoto e cols.22 demonstraram recentemente que a apelina induz a fosforilação da cadeia leve de miosina em células musculares lisas vasculares, a qual é de máxima importância na iniciação da contração do músculo liso, e a inibição da PKC, do NHE e do NCX se traduz numa redução significativa desse efeito.
Para além da via de inibição da adenilcíclase, a apelina ativa a via das ERKs (cínases reguladas extracelularmente) através de uma proteína G sensível à PTX (toxina pertussis), num processo dependente da PKC23. A via de controle da proliferação das células endoteliais PI3K-p70S6K é também ativada pela apelina mediante dois mecanismos, um ERK-dependente e outro PI3K-dependente24. É importante referir que a via PI3K/Akt parece ser a responsável pela fosforilação e conseqüente ativação da síntase do óxido nítrico endotelial (eNOS), a qual é imprescindível para a maioria dos efeitos vasoativos da apelina25.
Apelina/APJ: papel na fisiologia e patologia humanas e potenciais aplicações terapêuticas
Desenvolvimento e função cardiovasculares
A apelina e o seu receptor são expressos em tecidos embrionários e adultos. Dessa forma, desempenham um amplo espectro de ações, com início em fases muito precoces do desenvolvimento cardiovascular e que se continua até a idade adulta, sendo cada vez mais evidente a sua contribuição em vários processos fisiopatológicos.
Assim, foi já demonstrada a elevada expressão do receptor APJ no endotélio dos vasos embrionários14 e na túnica íntima dos vasos da retina, bem como o aumento e a diminuição da expressão desse receptor durante a formação e a estabilização dos vasos da retina, respectivamente26.
Esses dados fortalecem a idéia de que a apelina desempenha um papel importante na proliferação das células endoteliais, quer no desenvolvimento embrionário e estados fisiológicos quer em estados patológicos (exemplo, neoplasias malignas e retinopatia diabética). Desse modo, poderão ser imaginadas possíveis aplicações terapêuticas futuras, tais como o uso de agonistas para a promoção da angiogênese terapêutica (exemplo, situações isquêmicas), e de antagonistas angiogênicos, como estratégia para controlar o crescimento tumoral e a retinopatia diabética.
Os primeiros estudos sobre a distribuição tecidual da apelina e do receptor APJ apontaram para uma alta expressão dos respectivos RNAm no coração e em vasos sangüíneos de ratos e humanos4,6,14,17,18, sendo a expressão do APJ especialmente elevada no coração de rato15 e a apelina abundantemente expressa nas células endoteliais dos grandes vasos de condução humanos3. Mais recentemente, foi detectado RNAm do receptor APJ na túnica média dos grandes vasos27 e imuno-reatividade à apelina nas células endoteliais de pequenos vasos de resistência28.
Em 2000, Lee e cols.12 demonstraram, pela primeira vez, que após a infusão intravenosa de apelina-13 em ratos anestesiados, as pressões arteriais sistólica e diastólica sofriam uma queda temporária (de cerca de 10 mmHg). No seguimento desse trabalho, outro grupo de investigadores demonstrou que a eficácia do efeito hipotensor estava inversamente correlacionada com o peso molecular da isoforma da apelina, sendo esse efeito abolido após a administração de um inibidor da síntase de NO (NOS)25, o que sugere que a apelina exerce os seus efeitos vasoativos mediante um mecanismo dependente do NO. Com efeito, a apelina causa uma vasodilatação dependente do endotélio mediante a ativação da Akt que fosforila a eNOS e promove a liberação de NO e o aumento dos níveis de GMPc25. No entanto, pode desencadear uma resposta vasoconstritora na presença de um endotélio disfuncionante, atuando, nesse caso, sobre as células musculares lisas vasculares, que também expressam o receptor APJ27 (fig. 2).
Relativamente aos efeitos cardíacos, Szokodi e cols.21 demonstraram que, em preparações de coração intacto de rato, a apelina-16 tinha um efeito inotrópico positivo, aumentando a contractilidade com um EC50 de, aproximadamente, 30 pM, e um efeito máximo de cerca de 70% da resposta à isoprenalina (agonista beta-adrenérgico). Experiências in vivo confirmaram esses resultados mediante um aumento da pressão máxima desenvolvida (Pmáx) e da velocidade de elevação da pressão (dP/dtmáx) intraventriculares, tanto em ratos normais como em ratos com IC29. Estudos hemodinâmicos em camundongos demonstraram que a administração de apelina diminui a pré e a pós-carga do ventrículo esquerdo, aumentando cronicamente o débito cardíaco sem evidência de hipertrofia30.
Um estudo do nosso grupo, para além de confirmar o efeito inotrópico positivo in vivo, demonstrou um efeito inotrópico negativo da apelina no músculo cardíaco isolado, sugerindo que poderão ser necessárias outras células, para além das miocárdicas, para que o efeito inotrópico positivo se manifeste; demonstramos, também, que a apelina não exerce nenhuma função evidente sobre as propriedades diastólicas do miocárdio7.
O par apelina/APJ está intrinsecamente relacionado com o par AngII/AT-1 na regulação da fisiopatologia da função cardiovascular. Camundongos knock-out para o receptor APJ apresentam uma resposta vasopressora aumentada à Ang II; porém, apresentam valores normais de tensão arterial31. Num importante estudo de Iwanaga e cols.32, em ratos com IC (que apresentavam uma expressão reduzida da apelina e do receptor APJ), o tratamento com telmisartan, um antagonista dos receptores AT-1, fez que os níveis de apelina/APJ voltassem ao normal, sugerindo que a eficácia da inibição dos receptores AT1 na IC pode também dever-se à restituição dos níveis normais de apelina, um potente agente inotrópico positivo endógeno. Assim, é sugerida uma regulação direta do par apelina/APJ pelo par AngII/AT1.
A demonstração de que o exercício físico promove o aumento da expressão de apelina e do receptor APJ no miocárdio de ratos hipertensos e a transformação de uma hipertrofia patológica, de etiologia hipertensiva, numa hipertrofia fisiológica, em razão do exercício33, leva-nos a pensar que o efeito benéfico da atividade física na redução dos valores de tensão arterial em hipertensos poderá, em parte, ficar a dever-se ao aumento da expressão do sistema apelina/APJ em nível cardiovascular.
Para além dos seus efeitos fisiológicos, a apelina foi também implicada na patogênese da IC. Nos primeiros estudos, verificou-se que os níveis circulantes de apelina estavam aumentados em doentes com IC de início recente, diminuindo progressivamente com o agravamento da doença34. Um estudo populacional de grandes dimensões verificou que a concentração sérica de apelina estava diminuída em doentes com IC sistólica por disfunção ventricular esquerda35. Essa diminuição pode resultar da disfunção endotelial que acompanha a IC (afetando a produção de apelina)34 ou da exaustão de um possível mecanismo compensatório com vista à manutenção de um débito cardíaco adequado levado a cabo pelo sistema apelinérgico em doentes com IC. Em 2006, Dai e cols.36 demonstraram que a apelina exerce uma ação inotrópica positiva significativa no miocárdio insuficiente em conseqüência de um aumento no transiente de Ca2+.
Estudos recentes revelam que: 1) a terapia com apelina em ratos com lesões miocárdicas induzidas pela isoprenalina promove a recuperação da função cardíaca, tendo também um efeito cardioprotetor, por exemplo, contra a peroxidação lipídica37; 2) os níveis de apelina estão reduzidos em pacientes com doença coronária sujeitos a hemodiálise, sendo especulado que a apelina possa estar envolvida na fisiopatologia da doença cardiovascular secundária à insuficiência renal crônica, e que poderá ser utilizada no futuro como forma de tratamento da cardiomiopatia urêmica38; 3) a terapia de ressincronização ventricular promove, em longo prazo, um aumento dos níveis plasmáticos de apelina39, o que poderá estar associado à melhoria da função cardíaca; 4) apesar de ter sido considerada um possível marcador de hipóxia cardíaca (aguda e crônica)40,41 os níveis circulantes de apelina não parecem ser úteis na avaliação clínica e no prognóstico de doentes com IC aguda ou crônica35,42,43; 5) a apelina poderá ser utilizada como um marcador para avaliar o risco de desenvolver fibrilação auricular isolada44 e como marcador diagnóstico para distinguir a dispnéia de causa pulmonar daquela de origem cardiovascular45.
Em relação a este último aspecto, foi demonstrado por Ellinor e cols.44 que em pacientes com fibrilação auricular isolada os níveis de apelina estão reduzidos, o que sugere que a concentração sérica desse peptídeo poderá vir a ser utilizada como índice de risco para o aparecimento desse tipo de arritmia em sujeitos que ainda não a manifestaram. É interessante notar que os níveis plasmáticos de apelina-36 estão diminuídos em pacientes com doença crônica do parênquima pulmonar e função cardíaca preservada, e a determinação dos níveis de apelina-36 e proBNP (precursor do peptídeo natriurético do tipo B) poderá constituir, dessa forma, um novo meio diagnóstico que permita distinguir a dispnéia de causa pulmonar daquela de origem cardiovascular45.
Eixo hipotálamo-hipofisário e homeostasia de fluidos
Tal como já descrito, o receptor APJ e a apelina são expressos em várias áreas do SNC e, de forma particularmente intensa. nos núcleos supra-óptico (SON) e paraventricular (PVN) do hipotálamo46,47. Dado esse padrão de expressão, cedo se sugeriu que o sistema apelinérgico pudesse estar envolvido na modulação da atividade do eixo hipotálamo-hipofisário, assim como na homeostasia de fluidos.
Com efeito, foi já demonstrado que a administração de apelina aumenta os níveis séricos de ACTH (hormônio adrenocorticotrófico) e de corticosterona, diminuindo os de prolactina, FSH (hormônio folículo-estimulante) e LH (hormônio luteinizante), nos 30 minutos seguintes à infusão. Um estudo recente48 demonstrou que a apelina se localiza sobretudo nas células corticotrópicas, promovendo a liberação de ACTH de uma forma autócrina/parácrina, o que aponta para a existência de um sistema apelinérgico na hipófise de ratos adultos.
Apesar de a intensa expressão do receptor APJ e de a apelina em neurônios vasopressinérgicos ter sugerido uma importante função dessa na homeostasia dos fluidos, existe uma grande discrepância nos resultados dos estudos relativos a esse assunto, uma vez que a administração de apelina a ratos surgiu associada à redução do consumo de água46, bem como ao seu aumento12,49. A ausência de efeito no comportamento dípsico de ratos foi também descrita50.
Uma vez desvendada a associação entre a apelina e a homeostasia hídrica corporal, novas propostas terapêuticas relacionadas com o binômio apelina/APJ poderão surgir no campo de doenças tais como a hipertensão arterial e o diabetes insípido.
Relação com a insulina, a obesidade e a ingestão de alimentos
Em 2005, Boucher e cols.51 demonstraram que a apelina era uma nova adipocina endócrina, ao ser produzida em adipócitos maduros de camundongos e humanos. Para além disso, importantes correlações com os níveis de insulina e com a obesidade foram estabelecidas, sendo evidente um grande aumento dos níveis de apelina no plasma e nos adipócitos de camundongos obesos com hiperinsulinemia, bem como uma reduzida secreção de apelina pelos adipócitos de ratos insulino-dependentes com baixos níveis de insulina e durante o jejum, sendo essa secreção rapidamente reposta após a ingestão de alimentos, permitindo concluir que a insulina provoca um aumento da expressão da apelina. Essa importante relação com a insulina ganhou um novo contorno quando se demonstrou que o receptor APJ é expresso nos ilhotas pancreáticos e que a apelina inibe a secreção de insulina dependente da glicose através de uma ação direta sobre as células-b, em camundongos52.
É interessante verificar que existe uma correlação positiva entre os níveis séricos de apelina e o Índice de Massa Corporal53, que os níveis séricos de apelina estão aumentados em doentes diabéticos ou com intolerância a glicose54 e que o fator de necrose tumoral a (TNF-a) provoca um aumento da quantidade de apelina em adipócitos humanos55.
Num dos mais recentes estudos sobre o papel fisiopatológico da apelina, verificou-se que a administração intra-peritoneal dessa, em camundongos normais e obesos durante 14 dias, diminuiu a adiposidade corporal sem afetar o consumo de alimentos, diminuiu os níveis séricos de insulina, de leptina e de triglicerídeos, aumentou os níveis de adiponectina, aumentou a expressão de proteínas desacopladoras (UCP) e diminuiu o quociente respiratório56.
A apelina pode desempenhar também um importante papel na regulação da função cardiovascular no diabetes: o tratamento de camundongos diabéticos com apelina promove um aumento da resposta vasodilatadora à acetilcolina, através da via PI3K/Akt/eNOS57.
Todos esses resultados fazem acreditar que a apelina é um importante regulador do metabolismo glicolipídico normal, desempenhando também um possível papel em situações patológicas tais como a obesidade, a resistência a insulina e o diabete melito tipo 2. Desse modo, a sua utilização poderá ser explorada como um potencial alvo terapêutico nesse conjunto de doenças.
Tal como verificado para a homeostasia de fluidos, o papel da apelina no comportamento alimentar não está bem estabelecido, uma vez que os dados disponíveis ainda são escassos e contraditórios56,58,59, pelo que mais estudos são necessários para que novas conclusões sobre o papel da apelina na ingestão de alimentos possam ser obtidas.
Sistema apelinérgico e novas implicações fisiopatológicas
As ações do sistema apelinérgico anteriormente descritas foram aquelas que até agora receberam mais atenção por parte da comunidade médica. Porém, existe atualmente uma intensa frente de investigação com o intuito de demonstrar novas implicações fisiopatológicas desse recém-descoberto sistema.
Com efeito, no que respeita à função gastrointestinal, o binômio apelina/APJ parece desempenhar importantes funções na secreção gástrica de ácido e de colecistocinina e na proliferação do epitélio da mucosa do estômago60, estando possivelmente envolvido na patogênese da úlcera péptica, colite ulcerosa61 e doença de Crohn61. A apelina estimula a proliferação dos osteoblastos62,63 e inibe a sua apoptose62,63, podendo vir a ser no futuro usada como auxílio terapêutico em algumas doenças ósseas. Por último, o receptor APJ é um co-receptor para a entrada do VIH nas células do hospedeiro18,64, verificando-se que a apelina inibe a infecção de células CD4+ e APJ+, sendo essa eficácia maior para isoformas mais pesadas65-67.
Conclusão
Os novos papéis do sistema apelinérgico na fisiologia e patologia humanas estão em constante expansão. Com efeito, a apelina está envolvida na regulação da homeostasia de importantes sistemas do nosso organismo, bem como na patogênese de inúmeras doenças, muitas das quais com um peso enorme na morbilidade e mortalidade mundiais. Porém, nem tudo está feito. São necessários novos estudos para que todos os segredos da apelina e do receptor APJ possam ser desvendados, aumentando, assim, o nosso conhecimento sobre as funções do sistema apelinérgico na fisiologia e patologia humanas, sendo possível, então, equacionar o potencial que esse sistema nos oferece como futuro alvo terapêutico.
Potencial Conflito de Interesses
Declaro não haver conflito de interesses pertinentes.
Fontes de Financiamento
O presente estudo não teve fontes de financiamento externas.
Vinculação Acadêmica
Não há vinculação deste estudo a programas de pós-graduação.
Artigo recebido em 03/08/07; revisado recebido em 17/11/07; aceito em 04/12/07.
- 1. O'Dowd B, Heiber M, Chan A, Heng H, Tsui L, Kennedy J, et al. A human gene that shows identity with the gene encoding the angiotensin receptor is located on chromosome 11. Gene. 1993; 136 (1-2): 355-60.
- 2. Tatemoto K, Hosoya M, Habata Y, Fujii R, Kakegawa T, Zou MX, et al. Isolation and characterization of a novel endogenous peptide ligand for the human APJ receptor. Biochem Biophys Res Commun. 1998; 251 (2): 471-6.
- 3. Medhurst AD, Jennings CA, Robbins MJ, Davis RP, Ellis C, Winborn KY, et al. Pharmacological and immunohistochemical characterization of the APJ receptor and its endogenous ligand apelin. J Neurochem. 2003; 84 (5): 1162-72.
- 4. O'Carroll AM, Selby TL, Palkovits M, Lolait SJ. Distribution of mRNA encoding B78/apj, the rat homologue of the human APJ receptor, and its endogenous ligand apelin in brain and peripheral tissues. Biochim Biophys Acta. 2000; 1492 (1): 72-80.
- 5. Kleinz MJ, Skepper JN, Davenport AP. Immunocytochemical localisation of the apelin receptor, APJ, to human cardiomyocytes, vascular smooth muscle and endothelial cells. Regul Pept. 2005; 126 (3): 233-40.
- 6. Kawamata Y, Habata Y, Fukusumi S, Hosoya M, Fujii R, Hinuma S, et al. Molecular properties of apelin: tissue distribution and receptor binding. Biochim Biophys Acta. 2001; 1538 (2-3): 162-71.
- 7. Falcao-Pires I, Leite-Moreira AF. Apelin: a novel neurohumoral modulator of the cardiovascular system: pathophysiologic importance and potential use as a therapeutic target. Rev Port Cardiol. 2005; 24 (10): 1263-76.
- 8. Kleinz MJ, Davenport AP. Emerging roles of apelin in biology and medicine. Pharmacol Ther. 2005; 107 (2): 198-211.
- 9. Masri B, Knibiehler B, Audigier Y. Apelin signalling: a promising pathway from cloning to pharmacology. Cell Signal. 2005; 17 (4): 415-26.
- 10. Sorli SC, van den Berghe L, Masri B, Knibiehler B, Audigier Y. Therapeutic potential of interfering with apelin signalling. Drug Discov Today. 2006; 11 (23-24): 1100-6.
- 11. Lee DK, George SR, O'Dowd BF. Unravelling the roles of the apelin system: prospective therapeutic applications in heart failure and obesity. Trends Pharmacol Sci. 2006; 27 (4): 190-4.
- 12. Lee DK, Cheng R, Nguyen T, Fan T, Kariyawasam AP, Liu Y, et al. Characterization of apelin, the ligand for the APJ receptor. J Neurochem. 2000; 74 (1): 34-41.
- 13. Lee DK, Lanca AJ, Cheng R, Nguyen T, Ji XD, Gobeil F Jr., et al. Agonist-independent nuclear localization of the Apelin, angiotensin AT1, and bradykinin B2 receptors. J Biol Chem. 2004; 279 (9): 7901-8.
- 14. Devic E, Rizzoti K, Bodin S, Knibiehler B, Audigier Y. Amino acid sequence and embryonic expression of msr/apj, the mouse homolog of Xenopus X-msr and human APJ. Mech Dev. 1999; 84 (1-2): 199-203.
- 15. Hosoya M, Kawamata Y, Fukusumi S, Fujii R, Habata Y, Hinuma S, et al. Molecular and functional characteristics of APJ: tissue distribution of mRNA and interaction with the endogenous ligand apelin. J Biol Chem. 2000; 275 (28): 21061-7.
- 16. Lee DK, Saldivia VR, Nguyen T, Cheng R, George SR, O'Dowd BF. Modification of the terminal residue of apelin-13 antagonizes its hypotensive action. Endocrinology. 2005; 146 (1): 231-6.
- 17. Habata Y, Fujii R, Hosoya M, Fukusumi S, Kawamata Y, Hinuma S, et al. Apelin, the natural ligand of the orphan receptor APJ, is abundantly secreted in the colostrum. Biochim Biophys Acta. 1999; 1452 (1): 25-35.
- 18. Edinger A, Hoffman T, Sharron M, Lee B, Yi Y, Choe W, et al. An orphan seven-transmembrane domain receptor expressed widely in the brain functions as a coreceptor for human immunodeficiency virus type 1 and simian immunodeficiency virus. J Virol. 1998; 72 (10): 7934-40.
- 19. Vickers C, Hales P, Kaushik V, Dick L, Gavin J, Tang J, et al. Hydrolysis of biological peptides by human angiotensin-converting enzyme-related carboxypeptidase. J Biol Chem. 2002; 277 (17): 14838-43.
- 20. Danilczyk U, Penninger J. Angiotensin-converting enzyme II in the heart and the kidney. Circ Res. 2006; 98: 463-71.
- 21. Szokodi I, Tavi P, Foldes G, Voutilainen-Myllyla S, Ilves M, Tokola H, et al. Apelin, the novel endogenous ligand of the orphan receptor APJ, regulates cardiac contractility. Circ Res. 2002; 91 (5): 434-40.
- 22. Hashimoto T, Kihara M, Ishida J, Imai N, Yoshida S, Toya Y, et al. Apelin stimulates myosin light chain phosphorylation in vascular smooth muscle cells. Arterioscler Thromb Vasc Biol. 2006; 26 (6): 1267-72.
- 23. Masri B, Lahlou H, Mazarguil H, Knibiehler B, Audigier Y. Apelin (65-77) activates extracellular signal-regulated kinases via a PTX-sensitive G protein. Biochem Biophys Res Commun. 2002; 290 (1): 539-45.
- 24. Masri B, Morin N, Cornu M, Knibiehler B, Audigier Y. Apelin (65-77) activates p70 S6 kinase and is mitogenic for umbilical endothelial cells. Faseb J. 2004; 18 (15): 1909-11.
- 25. Tatemoto K, Takayama K, Zou MX, Kumaki I, Zhang W, Kumano K, et al. The novel peptide apelin lowers blood pressure via a nitric oxide-dependent mechanism. Regul Pept. 2001; 99 (2-3): 87-92.
- 26. Saint-Geniez M, Argence CB, Knibiehler B, Audigier Y. The msr/apj gene encoding the apelin receptor is an early and specific marker of the venous phenotype in the retinal vasculature. Gene Expr Patterns. 2003; 3 (4): 467-72.
- 27. Katugampola SD, Maguire JJ, Matthewson SR, Davenport AP. [(125)I]-(Pyr(1))Apelin-13 is a novel radioligand for localizing the APJ orphan receptor in human and rat tissues with evidence for a vasoconstrictor role in man. Br J Pharmacol. 2001; 132 (6): 1255-60.
- 28. Kleinz MJ, Davenport AP. Immunocytochemical localization of the endogenous vasoactive peptide apelin to human vascular and endocardial endothelial cells. Regul Pept. 2004; 118 (3): 119-25.
- 29. Berry MF, Pirolli TJ, Jayasankar V, Burdick J, Morine KJ, Gardner TJ, et al. Apelin has in vivo inotropic effects on normal and failing hearts. Circulation. 2004; 110 (11 Suppl 1): II187-93.
- 30. Ashley EA, Powers J, Chen M, Kundu R, Finsterbach T, Caffarelli A, et al. The endogenous peptide apelin potently improves cardiac contractility and reduces cardiac loading in vivo. Cardiovasc Res. 2005; 65 (1): 73-82.
- 31. Ishida J, Hashimoto T, Hashimoto Y, Nishiwaki S, Iguchi T, Harada S, et al. Regulatory roles for APJ, a seven-transmembrane receptor related to angiotensin-type 1 receptor in blood pressure in vivo. J Biol Chem. 2004; 279 (25): 26274-9.
- 32. Iwanaga Y, Kihara Y, Takenaka H, Kita T. Down-regulation of cardiac apelin system in hypertrophied and failing hearts: possible role of angiotensin II-angiotensin type 1 receptor system. J Mol Cell Cardiol. 2006; 41 (5): 798-806.
- 33. Zhang J, Ren CX, Qi YF, Lou LX, Chen L, Zhang LK, et al. Exercise training promotes expression of apelin and APJ of cardiovascular tissues in spontaneously hypertensive rats. Life Sci. 2006; 79 (12): 1153-9.
- 34. Foldes G, Horkay F, Szokodi I, Vuolteenaho O, Ilves M, Lindstedt KA, et al. Circulating and cardiac levels of apelin, the novel ligand of the orphan receptor APJ, in patients with heart failure. Biochem Biophys Res Commun. 2003; 308 (3): 480-5.
- 35. Chong KS, Gardner RS, Morton JJ, Ashley EA, McDonagh TA. Plasma concentrations of the novel peptide apelin are decreased in patients with chronic heart failure. Eur J Heart Fail. 2006; 8 (4): 355-60.
- 36. Dai T, Ramirez-Correa G, Gao WD. Apelin increases contractility in failing cardiac muscle. Eur J Pharmacol. 2006; 553 (1-3): 222-8.
- 37. Jia YX, Pan CS, Zhang J, Geng B, Zhao J, Gerns H, et al. Apelin protects myocardial injury induced by isoproterenol in rats. Regul Pept. 2006; 133 (1-3): 147-54.
- 38. Malyszko J, Malyszko JS, Kozminski P, Mysliwiec M. Apelin and cardiac function in hemodialyzed patients: possible relations? Am J Nephrol. 2006; 26 (2): 121-6.
- 39. Francia P, Salvati A, Balla C, De Paolis P, Pagannone E, Borro M, et al. Cardiac resynchronization therapy increases plasma levels of the endogenous inotrope apelin. Eur J Heart Fail. 2007; 9 (3): 306-9.
- 40. Ronkainen VP, Ronkainen JJ, Hanninen SL, Leskinen H, Ruas JL, Pereira T, et al. Hypoxia inducible factor regulates the cardiac expression and secretion of apelin. Faseb J. 2007; 21 (8): 1821-30.
- 41. Atluri P, Morine KJ, Liao GP, Panlilio CM, Berry MF, Hsu VM, et al. Ischemic heart failure enhances endogenous myocardial apelin and APJ receptor expression. Cell Mol Biol Lett. 2007; 12 (1): 127-38.
- 42. van Kimmenade RR, Januzzi JL Jr., Ellinor PT, Sharma UC, Bakker JA, Low AF, et al. Utility of amino-terminal pro-brain natriuretic peptide, galectin-3, and apelin for the evaluation of patients with acute heart failure. J Am Coll Cardiol. 2006; 48 (6): 1217-24.
- 43. Miettinen KH, Magga J, Vuolteenaho O, Vanninen EJ, Punnonen KR, Ylitalo K, et al. Utility of plasma apelin and other indices of cardiac dysfunction in the clinical assessment of patients with dilated cardiomyopathy. Regul Pept. 2007; 140 (3): 178-84.
- 44. Ellinor PT, Low AF, Macrae CA. Reduced apelin levels in lone atrial fibrillation. Eur Heart J. 2006; 27 (2): 222-6.
- 45. Goetze JP, Rehfeld JF, Carlsen J, Videbaek R, Andersen CB, Boesgaard S, et al. Apelin: a new plasma marker of cardiopulmonary disease. Regul Pept. 2006; 133 (1-3): 134-8.
- 46. Reaux A, De Mota N, Skultetyova I, Lenkei Z, El Messari S, Gallatz K, et al. Physiological role of a novel neuropeptide, apelin, and its receptor in the rat brain. J Neurochem. 2001; 77 (4): 1085-96.
- 47. Reaux A, Gallatz K, Palkovits M, Llorens-Cortes C. Distribution of apelin-synthesizing neurons in the adult rat brain. Neuroscience. 2002; 113 (3): 653-62.
- 48. Reaux-Le Goazigo A, Alvear-Perez R, Zizzari P, Epelbaum J, Bluet-Pajot MT, Llorens-Cortes C. Cellular localization of apelin and its receptor in the anterior pituitary: evidence for a direct stimulatory action of apelin on ACTH release. Am J Physiol Endocrinol Metab. 2007; 292 (1): E7-15.
- 49. Taheri S, Murphy K, Cohen M, Sujkovic E, Kennedy A, Dhillo W, et al. The effects of centrally administered apelin-13 on food intake, water intake and pituitary hormone release in rats. Biochem Biophys Res Commun. 2002; 291 (5): 1208-12.
- 50. Mitra A, Katovich MJ, Mecca A, Rowland NE. Effects of central and peripheral injections of apelin on fluid intake and cardiovascular parameters in rats. Physiol Behav. 2006; 89 (2): 221-5.
- 51. Boucher J, Masri B, Daviaud D, Gesta S, Guigne C, Mazzucotelli A, et al. Apelin, a newly identified adipokine up-regulated by insulin and obesity. Endocrinology. 2005; 146 (4): 1764-71.
- 52. Sorhede Winzell M, Magnusson C, Ahren B. The apj receptor is expressed in pancreatic islets and its ligand, apelin, inhibits insulin secretion in mice. Regul Pept. 2005; 131 (1-3): 12-7.
- 53. Heinonen MV, Purhonen AK, Miettinen P, Paakkonen M, Pirinen E, Alhava E, et al. Apelin, orexin-A and leptin plasma levels in morbid obesity and effect of gastric banding. Regul Pept. 2005; 130 (1-2): 7-13.
- 54. Li L, Yang G, Li Q, Tang Y, Yang M, Yang H, et al. Changes and relations of circulating visfatin, apelin, and resistin levels in normal, impaired glucose tolerance, and type 2 diabetic subjects. Exp Clin Endocrinol Diabetes. 2006; 114 (10): 544-8.
- 55. Daviaud D, Boucher J, Gesta S, Dray C, Guigne C, Quilliot D, et al. TNFalpha up-regulates apelin expression in human and mouse adipose tissue. Faseb J. 2006; 20 (9): 1528-30.
- 56. Higuchi K, Masaki T, Gotoh K, Chiba S, Katsuragi I, Tanaka K, et al. Apelin, an APJ receptor ligand, regulates body adiposity and favors the messenger ribonucleic acid expression of uncoupling proteins in mice. Endocrinology. 2007; 148 (6): 2690-7.
- 57. Zhong JC, Yu XY, Huang Y, Yung LM, Lau CW, Lin SG. Apelin modulates aortic vascular tone via endothelial nitric oxide synthase phosphorylation pathway in diabetic mice. Cardiovasc Res. 2007; 74 (3): 388-95.
- 58. Sunter D, Hewson AK, Dickson SL. Intracerebroventricular injection of apelin-13 reduces food intake in the rat. Neurosci Lett. 2003; 353 (1): 1-4.
- 59. O'Shea M, Hansen MJ, Tatemoto K, Morris MJ. Inhibitory effect of apelin-12 on nocturnal food intake in the rat. Nutr Neurosci. 2003; 6 (3): 163-7.
- 60. Wang G, Anini Y, Wei W, Qi X, AM OC, Mochizuki T, et al. Apelin, a new enteric peptide: localization in the gastrointestinal tract, ontogeny, and stimulation of gastric cell proliferation and of cholecystokinin secretion. Endocrinology. 2004; 145 (3): 1342-8.
- 61. Han S, Wang G, Qiu S, de la Motte C, Wang HQ, Gomez G, et al. Increased colonic apelin production in rodents with experimental colitis and in humans with IBD. Regul Pept. 2007; 142 (3): 131-7.
- 62. Tang SY, Xie H, Yuan LQ, Luo XH, Huang J, Cui RR, et al. Apelin stimulates proliferation and suppresses apoptosis of mouse osteoblastic cell line MC3T3-E1 via JNK and PI3-K/Akt signaling pathways. Peptides. 2007; 28 (3): 708-18.
- 63. Xie H, Yuan LQ, Luo XH, Huang J, Cui RR, Guo LJ, et al. Apelin suppresses apoptosis of human osteoblasts. Apoptosis. 2007; 12 (1): 247-54.
- 64. Choe H, Farzan M, Konkel M, Martin K, Sun Y, Marcon L, et al. The orphan seven-transmembrane receptor apj supports the entry of primary T-cell-line-tropic and dualtropic human immunodeficiency virus type 1. J Virol. 1998; 72 (7): 6113-8.
- 65. Puffer BA, Sharron M, Coughlan CM, Baribaud F, McManus CM, Lee B, et al. Expression and coreceptor function of APJ for primate immunodeficiency viruses. Virology. 2000; 276 (2): 435-44.
- 66. Cayabyab M, Hinuma S, Farzan M, Choe H, Fukusumi S, Kitada C, et al. Apelin, the natural ligand of the orphan seven-transmembrane receptor APJ, inhibits human immunodeficiency virus type 1 entry. J Virol. 2000; 74 (24): 11972-6.
- 67. Zou MX, Liu HY, Haraguchi Y, Soda Y, Tatemoto K, Hoshino H. Apelin peptides block the entry of human immunodeficiency virus (HIV). FEBS Lett. 2000; 473 (1): 15-8.
Datas de Publicação
-
Publicação nesta coleção
27 Maio 2008 -
Data do Fascículo
Maio 2008
Histórico
-
Aceito
04 Dez 2007 -
Revisado
17 Nov 2007 -
Recebido
03 Ago 2007