Resumos
Uma análise cladística do relacionamento entre as espécies sul-americanas do grupo Sitalces (Sitalces Stål, 1878, Eusitalces Bruner, 1911, Parasitalces Bruner, 1911, Psilocirtus Bruner, 1911, Liebermannacris Costa & Carvalho, 2006, Robustusacris Costa & Carvalho, 2006, Arimacris Costa & Carvalho, 2006, Salvadoracris Costa & Carvalho, 2006 e Caruaruacris Costa & Carvalho, 2006) é apresentada. A análise incluiu 14 espécies com três grupos-externos e 34 caracteres morfológicos. A monofilia do grupo é suportada por três sinapomorfias com índices de consistência e retenção de 100%: prozona maior que a metazona, borda posterior do pronoto não angulada e placa supranal mais curta em relação aos cercos. A análise resultou em um único cladograma: (P. olivaceus (E. vittatus (L. dorsualis; L. punctifrons) (R. balzapambae (A. trinitatis (S. nigritus; C. bivittatus) (S. volxemi (P. vulneratus; P. sexnotata)))))). Comentários biogeográficos são apresentados e relacionados com padrões pré-estabelecidos.
Cladística; gafanhotos; Ommatolampinae; grupo Sitalces; Neotropical
A cladistic analysis of the relationships among the species of the South American genera of the group Sitalces (Sitalces Stål, 1878, Eusitalces Bruner, 1911, Parasitalces Bruner, 1911, Psilocirtus Bruner, 1911, Liebermannacris Costa & Carvalho, 2006, Robustusacris Costa & Carvalho, 2006, Arimacris Costa & Carvalho, 2006, Salvadoracris Costa & Carvalho, 2006 and Caruaruacris Costa & Carvalho, 2006) is presented. The analysis included 14 species with three outgroups and 34 morphological characters. The monophyly of the group is supported by three sinapomorphies with 100% consistency and retention indices: prozona larger than the metazona, posterior border of the pronotum not angulated and supranal plate shorter than cerci. The analysis resulted in a single cladogram: (P. olivaceus (E. vittatus (L. dorsualis; L. punctifrons) (R. balzapambae (A. trinitatis (S. nigritus; C. bivittatus) (S. volxemi (P. vulneratus; P. sexnotata)))))). Biogeographic comments are presented and related with pre-established standards.
Cladistics; grasshoppers; Ommatolampinae; Sitalces group; Neotropical
Análise cladística e biogeográfica dos gêneros do grupo Sitalces (Orthoptera, Acrididae, Abracrini)
Cladistic and biogeographic analysis of the genera of the group Sitalces (Orthoptera, Acrididae, Abracrini)
Maria Kátia Matiotti da Costa; Augusto Ferrari; Gervásio Silva Carvalho
Laboratório de Entomologia, Faculdade de Biociências, PUCRS, Av. Ipiranga, 6681, Prédio 12D, sala 45, Caixa Postal 1429, 90619-900 Porto Alegre, RS, Brasil. (katiamatiotti@yahoo.com.br)
RESUMO
Uma análise cladística do relacionamento entre as espécies sul-americanas do grupo Sitalces (Sitalces Stål, 1878, Eusitalces Bruner, 1911, Parasitalces Bruner, 1911, Psilocirtus Bruner, 1911, Liebermannacris Costa & Carvalho, 2006, Robustusacris Costa & Carvalho, 2006, Arimacris Costa & Carvalho, 2006, Salvadoracris Costa & Carvalho, 2006 e Caruaruacris Costa & Carvalho, 2006) é apresentada. A análise incluiu 14 espécies com três grupos-externos e 34 caracteres morfológicos. A monofilia do grupo é suportada por três sinapomorfias com índices de consistência e retenção de 100%: prozona maior que a metazona, borda posterior do pronoto não angulada e placa supranal mais curta em relação aos cercos. A análise resultou em um único cladograma: (P. olivaceus (E. vittatus (L. dorsualis; L. punctifrons) (R. balzapambae (A. trinitatis (S. nigritus; C. bivittatus) (S. volxemi (P. vulneratus; P. sexnotata)))))). Comentários biogeográficos são apresentados e relacionados com padrões pré-estabelecidos.
Palavras-chave: Cladística, gafanhotos, Ommatolampinae, grupo Sitalces, Neotropical.
ABSTRACT
A cladistic analysis of the relationships among the species of the South American genera of the group Sitalces (Sitalces Stål, 1878, Eusitalces Bruner, 1911, Parasitalces Bruner, 1911, Psilocirtus Bruner, 1911, Liebermannacris Costa & Carvalho, 2006, Robustusacris Costa & Carvalho, 2006, Arimacris Costa & Carvalho, 2006, Salvadoracris Costa & Carvalho, 2006 and Caruaruacris Costa & Carvalho, 2006) is presented. The analysis included 14 species with three outgroups and 34 morphological characters. The monophyly of the group is supported by three sinapomorphies with 100% consistency and retention indices: prozona larger than the metazona, posterior border of the pronotum not angulated and supranal plate shorter than cerci. The analysis resulted in a single cladogram: (P. olivaceus (E. vittatus (L. dorsualis; L. punctifrons) (R. balzapambae (A. trinitatis (S. nigritus; C. bivittatus) (S. volxemi (P. vulneratus; P. sexnotata)))))). Biogeographic comments are presented and related with pre-established standards.
Keywords: Cladistics, grasshoppers, Ommatolampinae, Sitalces group, Neotropical.
AMÉDÉGNATO (1974) propôs uma nova classificação para os acridóideos neotropicais. Nessa classificação, várias subfamílias novas foram acrescentadas em Acrididae, entre as quais Ommatolampinae, onde Abracrini foi incluída.
Ommatolampinae constitui um grupo grande e diversificado, incluindo aproximadamente 100 gêneros e 280 espécies (CIGLIANO & LANGE, 1988). Seus membros diferenciam-se dos demais pelo mesonoto não visível, ou visível somente na região posterior. Abracrini (AMÉDÉGNATO, 1974) inclui 16 gêneros e aproximadamente 90 espécies representados na região Neotropical. Os gêneros pertencentes a esta tribo podem ser diferenciados dos demais basicamente pelos caracteres da genitália do macho, onde a apófise interna é ausente ou vestigial.
As relações evolutivas nos grupos de Ommatolampinae e Abracrini ainda não foram estudadas com a utilização da metodologia da sistemática filogenética. AMÉDÉGNATO & DESCAMPS (1979) e PEREZ et al. (1995) reuniram alguns gêneros em uma classificação com base em semelhanças e diferenças de caracteres morfológicos externos e afinidades empíricas. Esses grupos, entre os quais o grupo Sitalces, apesar de não refletirem necessariamente a filogenia, são frutos de uma análise morfológica rigorosa, podendo constituir em ponto de partida para estudos cladísticos.
O grupo Sitalces, estabelecido nesse estudo, é formado pelos gêneros Sitalces Stål, 1878, Eusitalces Bruner, 1911, Parasitalces Bruner, 1911, Psiloscirtus Bruner, 1911, LiebermannacrisCosta & Carvalho, 2006, Robustusacris Costa & Carvalho, 2006, Arimacris Costa & Carvalho, 2006, Salvadoracris Costa & Carvalho, 2006, Caruaruacris Costa & Carvalho, 2006. Apresentam como características principais a micropteria ou a ausência de asas e a apófise interna na genitália vestigial. Todas as espécies do grupo em estudo são exclusivamente neotropicais.
Considerando a falta de estudos cladísticos em Abracrini, objetivou-se testar a monofilia do grupo Sitalces, apresentar uma hipótese das relações de parentesco entre suas espécies, fornecer subsídios para futuros estudos em Ommatolampinae e apresentar comentários biogeográficos do grupo em estudo.
MATERIAL E MÉTODOS
A elaboração de hipótese das relações de parentesco foi feita com base na metodologia cladística (HENNIG, 1966; ELDREDGE & CRACRAFT, 1980; WILEY, 1981; AX, 1987).
A análise e seleção dos caracteres foram baseadas em um estudo detalhado da morfologia externa e da genitália dos espécimes disponíveis dos representantes dos gêneros e dos grupos externos pertencentes a Abracrini.
O grupo interno foi representado por material-tipo ou um espécime comparado com o tipo, como a seguir: Sitalces volxemi Stål, 1878; Liebermannacris dorsualis (Giglio-Tos, 1898); Liebermannacris punctifrons (Stål, 1878); Robustusacris balzapambae (Rehn, 1913); Arimacris trinitatis (Bruner, 1906); Eusitalces vittatus Bruner, 1911; Parasitalces sexnotata Bruner, 1911; Parasitalces vulneratus (Bruner, 1919); Salvadoracris nigritus Costa & Carvalho, 2006; Caruaruacris bivittatus Costa & Carvalho, 2006 e Psiloscirtus olivaceus Bruner, 1911.
Os espécimes examinados estão depositados nas seguintes Instituições: Academy of Natural Sciences of Philadelphia, USA (ANSP); Facultad de Ciencias, Universidad de la Republica, Montevideo, Uruguay (FCUR); Instituto Nacional de Pesquisas da Amazônia, Manaus, Brasil (INPA); Museo de La Plata, Division de Entomología, Universidad Nacional de La Plata, La Plata, Argentina (MLPA); Museu de Zoologia da Universidade de São Paulo, São Paulo, Brasil (MZUSP); Museo e Instituto di Zoología e Anatomia Comparata, Universitá di Torino, Torino, Itália (MIZT); Museo Nacional de Ciencias Naturales, Madrid, Espanha (MNMS); Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brasil (MNRJ); Naturhistoriska Riksmuseet, Estocolmo, Suécia (NHRS).
Foram observados 34 caracteres, 19 da morfologia externa (caracteres 1-19), 12 do complexo fálico (caracteres 20-31) e três da genitália externa da fêmea (caracteres 32-34). Alguns foram interpretados e modificados a partir da literatura geral sobre Acrididae [RONDEROS & CIGLIANO, 1991; CIGLIANO et al. (1996); CIGLIANO (1997)], outros propostos e discutidos pela primeira vez neste trabalho. A nomenclatura utilizada para os caracteres seguiu a bibliografia citada.
A polaridade dos caracteres foi baseada no método de comparação com o grupo-externo (NIXON & CARPENTER, 1993). O grupo-externo é composto por espécies de três gêneros de Abracrini: Jodacris chapadensis Bruner, 1911; Orthoscapheus coryaceus (Giglio-Tos, 1894) e Omalotettix obliquus (Thunberg, 1824). Os critérios utilizados para escolher estes taxóns devem-se a uma análise prévia dos caracteres morfológicos. Foram examinados os espécimens comparados com o tipo.
O grupo-externo e o grupo de estudo são listados na matriz (Tab. I), indicando o estado de cada caráter: plesiomórfico (0), apomórfico (1, 2) e caráter ausente, não comparável ou desconhecido (?). Caracteres multiestados foram tratados como não-aditivos. A notação "grupo +" (AMORIM, 1982) foi usada nos resultados e discussão.
A matriz de dados foi processada pelo programa HENNIG 86 versão 1.5 (FARRIS, 1988), usando-se a opção ie (enumeração implícita), a qual garante encontrar todas as árvores mais parcimoniosas, além do cálculo dos índices de consistência (KLUGE & FARRIS, 1969) e retenção (FARRIS, 1989). Este programa foi utilizado devido à sua eficiência e velocidade em trabalhar com os dados (PLATNICK, 1989).
A matriz de dados foi elaborada com base no exame dos espécimens que estão depositados nas Instituições e de fotografias dos tipos cedidas gentilmente pelo Prof. Carlos Salvador Carbonell, e complementada com as descrições de StÅl, 1878; BRUNNER VON WATTENWYL, 1893; GIGLIO-TOS, 1898; BRUNER, 1908, 1911, 1919; REHN, 1917 e GUNTHER, 1940.
A terminologia adotada foi a comumente utilizada em Acrididae e que, de uma maneira geral, foi estabelecida ao longo dos anos pelos trabalhos de BRUNER (1911), AMÉDÉGNATO (1974), AMÉDÉGNATO & DESCAMPS (1979). As mensurações foram realizadas utilizando um paquímetro digital e, quando necessário, microscópio estereoscópico com ocular micrométrica.
No tocante à biogeografia, utilizaram-se hipóteses prévias da literatura para comparações e discussões.
RESULTADOS E DISCUSSÃO
Análise cladística. A análise resultou em um único cladograma com 64 passos, com índices de consistência 0,57 e retenção de 0,72 (Fig. 1).
A hipótese de monofilia do grupo Sitalces é corroborada por três sinapomorfias com 100% de consistência: prozona maior que a metazona (caráter 8), borda posterior do pronoto não angulada (9) e placa supranal mais curta em relação aos cercos (16).
A análise mostrou que alguns caracteres da genitália interna do macho e boa parte dos da morfologia externa mostraram-se úteis no reconhecimento de surgimentos únicos, quais sejam: forma da placa supranal (14), placa supranal em relação aos cercos (16), protuberância mediana do epífalo (21), comprimento das âncoras em relação as projeções anteriores (25), direção das âncoras (26), comprimento das valvas do edeago (28) e fastígio (4), costa frontal sobre o ocelo mediano (7), prozona maior que a metazona (8), borda posterior do pronoto não-angulada (9), número de espinhos da tíbia posterior (13). Estes resultados representam um avanço na sistemática do grupo, representando uma primeira hipótese sobre as relações filogenéticas de um grupo de Abracrini, na proposição de caracteres genéricos diagnósticos, servindo ainda para a realização de futuros trabalhos dentro de Ommatolampinae.
Abaixo segue a descrição dos 34 caracteres analisados, seus estados e índices de consistência (ic) e retenção (ir). A sequência dos caracteres corresponde à matriz de dados (Tab. I).
1. Relação cabeça/pronoto: (0) tão larga quanto o pronoto; (1) mais larga do que o pronoto. (ic= 33; ir = 60).
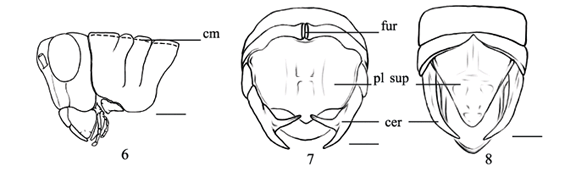
AMÉDÉGNATO & DESCAMPS (1979) comentam que a cabeça dos integrantes de Abracrini pode apresentar-se de duas formas: tão larga quanto o pronoto (Fig. 2) ou mais larga do que o pronoto (Fig. 3), sendo esta uma sinapomorfia para o grupo Sitalces com reversões em R. balzapambae e P. vulneratus + P. sexnotata.

2. Espaço entre olho e a margem lateral da cabeça: (0) presente; (1) ausente (ic = 100; ir = 100).
Ausência de espaço entre o olho e a margem lateral da cabeça é uma sinapomorfia do grupo Sitalces e ocorre, paralelamente, em O. obliquus.
3. Espaço interocular em relação ao primeiro artículo antenal: (0) menor; (1) maior (ic = 25; ir = 25).
A condição espaço interocular maior que o primeiro artículo antenal, apomórfica, é uma homoplasia para J. chapadensis, R. balzabampae, C. bivittatus e P. vulneratus+.
4. Fastígio: (0) mais largo do que longo; (1) mais longo do que largo (ic =100; ir = 100).
O fastígio localiza-se na porção dorsal da cabeça e auxilia na identificação dos acridídeos. S. nigritus + C. bivittatus compartilham a condição de fastígio mais longo do que largo (Fig. 4), representando uma sinapomorfia para o clado. O fastígio mais largo do que longo (Fig. 5) é comum entre os Abracrini e está presente em quase todos os táxons do grupo interno.
5. Número de artículos antenais: (0) 15-16; (1) 17-18; (2) 19 (ic = 66; ir = 75).
O número de artículos antenais apresenta uma grande variabilidade dentro dos táxons de Abracrini (COSTA & CARVALHO, 2006a, b). Presença de 17-18 artículos é uma condição sinapomórfica do grupo Sitalces, com reversão em C. bivittatus. Para o clado P. vulneratus + P. sexnotata, 19 artículos é uma sinapomorfia.
6. Costa frontal: (0) sulcada; (1) plana (ic = 50; ir = 83).
A costa frontal é uma estrutura que se localiza frontalmente na região mediana da cabeça dos gafanhotos. O estado apomórfico do caráter, costa frontal plana, é homoplásico para os grupos L. dorsualis + L. punctifrons e S. nigritus + C. bivittatus+. Os demais táxons apresentam a costa frontal bastante sulcada na região mediana.
7. Costa frontal sobre o ocelo mediano: (0) proeminente; (1) não-proeminente (ic = 100; ir = 100).
Costa frontal não proeminente sobre o ocelo mediano corrobora o parentesco do clado que reúne os grupos L. dorsualis + L. punctifrons e R. balzapambae+.
8. Comprimento da prozona em relação ao tamanho da metazona: (0) prozona igual à metazona; (1) prozona maior que a metazona (ic = 100; ir = 100).
A relação do comprimento entre a prozona e a metazona varia muito dentro de Abracrini, podendo ser a prozona do mesmo tamanho ou maior que a metazona (Fig. 5). O estado apomórfico é uma sinapomorfia para o grupo Sitalces. As variações do comprimento da prozona em relação à metazona no pronoto foram usadas por CIGLIANO (1997) em seus estudos para os gêneros Eurotettix Bruner, 1906, Leiotettix Bruner, 1906 e Scotussa Giglio-Tos, 1894, todos pertencentes a Melanoplinae, auxiliando na sustentação dos clados.
9. Borda posterior do pronoto: (0) angulada; (1) não-angulada (ic = 100; ir = 100).
A presença de borda posterior do pronoto não-angulada (Fig. 4) corrobora a monofilia do grupo Sitalces. Dentro de Abracrini existe uma variação em relação à borda do pronoto, apresentando-se angulada ou não. Esse caráter é usado para a sustentação de outras subfamílias de Acrididae, como Melanoplinae (CIGLIANO, 1997; CIGLIANO et al. 1996) e de famílias de Acridoidea, como Tristiridae (CIGLIANO, 1989).
10. Carena mediana do pronoto: (0) proeminente; (1) não-proeminente (ic = 50; ir = 66).
No grupo Sitalces, a carena mediana do pronoto apresenta-se não-proeminente (Fig. 6), condição sinapomórfica (COSTA & CARVALHO, 2006a, b), com reversão em R. balzapambae.
11. Desenvolvimento da tégmina: (0) macróptera; (1) micróptera; (2) áptera (ic = 66; ir = 66).
Asas micrópteras são uma sinapomorfia do grupo Sitalces. O estado 2, de ocorrência freqüente nos ortopteróides (por exemplo, Phasmatodea, Blattodea), surge independentemente em Psilocirtus olivaceus e Parasitalces sexnotata.
12. Fêmur posterior em relação ao abdome: (0) alcança a extremidade do abdome; (1) ultrapassa a extremidade do abdome (ic = 33; ir = 33).
Fêmur posterior ultrapassando a extremidade do abdome sustenta a monofilia do clado E. vittatus+, com reversões em L. punctifrons e no clado S. nigritus+ + C. bivittatus + S. volxemi+.
13. Números de espinhos na tíbia posterior: (0) 6-7; (1) 7-8 (ic = 100; ir = 100).
Nos integrantes de Ommatolampinae o número de espinhos na tíbia posterior é muito variável (AMÉDÉGNATO & DESCAMPS, 1979). Nos táxons do grupo de estudo, a presença de 7-8 espinhos na tíbia posterior é uma condição sinapomórfica do clado R. balzapambae+.
14. Forma da placa supranal: (0) subtriangular; (1) sub-retangular (ic = 100; ir = 100).
A monofilia de S. volxemi+é corroborada com base no compartilhamento do estado do caráter placa supranal sub-retangular (Fig. 7). Ocorre uma variabilidade nos integrantes de Abracrini quanto à forma da placa supranal, a qual apresenta-se subtriangular ou sub-retangular. A maioria dos táxons do grupo Sitalces apresenta a placa supranal triangular com exceção de S. volxemi+. Para os estudos da tribo Dichroplini de Melanoplinae (ROONDEROS & CIGLIANO, 1991), a forma da placa supranal ajuda na sustentação de clados.
15. Medidas da placa supranal: (0) mais longa do que larga; (1) mais larga do que longa (ic=50; ir=80).
A placa supranal do macho, mais larga do que longa, é uma condição sinapomórfica para R. balzapambae+, com reversão em C. bivittatus. Este caráter é utilizado para a identificação das espécies pertencentes ao grupo de estudo e para a sustentação de Ronderosia (Cigliano, 1997), de vários gêneros pertencentes a Dichroplini, Melanoplinae (CIGLIANO et al., 1996) e também dentro de Acridoidea, na família Tristiridae (CIGLIANO, 1989).
16. Placa supranal em relação aos cercos: (0) mais longa; (1) mais curta (ic=100; ir=100).
A grande maioria dos táxons pertecentes a Abracrini apresenta uma variação com relação ao tamanho entre a placa e os cercos (COSTA & CARVALHO, 2006a). Nos táxons do grupo interno, a placa supranal apresenta-se mais curta em relação aos cercos (Fig. 8), sendo esta uma condição sinapomórfica para o grupo Sitalces.
17. Protuberância dorsal na região mediana da placa supranal: (0) ausente; (1) presente (ic=50; ir=50).
A protuberância dorsal mediana existente na placa supranal (COSTA & CARVALHO, 2006a,b), encontrada em alguns táxons do grupo interno, representa uma sinapomorfia para o clado A. trinitatis +, sofrendo uma reversão em S. volxemi +.
18. Cercos: (0) bifurcados; (1) não-bifurcados (ic=50; ir=83).
Dentre os Abracrini os cercos apresentam-se bifurcados (Fig. 7) ou não-bifurcados (Fig. 8), sendo esta uma condição sinapomórfica do clado L. dorsualis + + R. balzapambae,+ com reversão em P. vulneratus+ P. sexnotata. Em Dichroplini (RONDEROS & CIGLIANO, 1991; CIGLIANO, 1997) a bifurcação, forma e posição dos cercos são caracteres utilizados para sustentação de clados.
19. Fúrcula: (0) ausente; (1) presente (ic = 50; ir = 83).
A fúrcula é uma estrutura encontrada dorsalmente no último tergito do abdome. A condição de fúrcula presente (Fig. 7) no grupo de estudo surge no clado A. trinitatis + e ocorre também em P. olivaceus. Em alguns táxons de Abracrini encontra-se ausente (Fig. 8), sendo esse caráter usado também nos estudos de Melanoplinae, Dichroplini (RONDEROS & CIGLIANO, 1991).
20. Forma da ponte do epífalo em vista dorsal: (0) plana; (1) convexa; (2) côncava (ic = 66; ir = 0).
A ponte do epífalo plana (Fig. 9) é a condição encontrada na maioria das espécies do grupo de estudo, porém, E. vittatus e L. dorsualis apresentam a condição convexa (Fig. 10). Em S. nigritus, a ponte do epífalo é côncava (Fig. 11), constituindo uma autapomorfia para esta espécie. O epífalo é uma estrutura encontrada na genitália interna do macho, bastante utilizada para identificação em nível de espécie nos estudos de CIGLIANO (1989, 1997) e para Scotussa Giglio-Tos, 1894 e Leiotettix Bruner, 1906 (CIGLIANO et al., 1996), mostrando-se útil para a sustentação dos clados de outras subfamílias de Acrididae e famílias de Acridoidea.
21. Protuberância mediana do epífalo: (0) presente; (1) ausente (ic=100; ir=100).
Na maioria dos táxons do grupo Sitalces, o epífalo dos machos apresenta uma protuberância mediana (Fig. 9). A ausência desta em S. nigritus + C. bivittatus (Fig. 11), constitui uma condição sinapomórfica para este clado.
22. Extremidade do lóbulo do epífalo: (0) não-curvada; (1) curvada (ic=50; ir=75).
A extremidade do lóbulo do epífalo curvada (Fig. 10), é uma sinapomorfia de L. dorsualis+ + R. balzapambae+, com reversão em S. volxemi +.
23. Projeções anteriores do epífalo em relação às âncoras: (0) desenvolvidas; (1) muito desenvolvidas (ic=50; ir=0).
Projeções anteriores do epífalo muito desenvolvidas (Fig. 10) surgem paralelamente em E. vittatus e no clado L. dorsualis + L. punctifrons. Este caráter foi utilizado pela primeira vez na descrição do grupo Sitalces por COSTA & CARVALHO (2006b).
24. Esclerotizações nos lóbulos do epífalo: (0) ausentes; (1) presentes (ic=50; ir=0).
Esclerotizações presentes dorsalmente nos lóbulos do epífalo (Fig. 11) é condição encontrada homoplasicamente em A. trinitatis e S. nigritus. Encontra-se ausente no restante dos táxons do grupo Sitalces e também em gêneros de Abracrini.
25. Comprimento das âncoras em relação às projeções anteriores: (0) mais curtas; (1) tão longas quanto (ic=100; ir=100).
Âncoras tão longas quanto as projeções anteriores é uma sinapomorfia do clado R. balzapambae+. As âncoras e projeções anteriores são estruturas do epífalo importantes para identificação das espécies e este caráter está sendo testado pela primeira vez neste trabalho.
26. Direção das âncoras do epífalo: (0) convergentes; (1) divergentes (ic=100; ir=100).
O epífalo apresenta a direção das âncoras de duas maneiras: convergente, encontrada na maioria dos táxons de Abracrini (COSTA & CARVALHO, 2006a,b); e divergente, sinapomorfia de S. nigritus + C. bivittatus.
27. Escleritos ovais do epífalo: (0) presentes; (1) ausentes (ic=33; ir=50).
Os escleritos ovais são estruturas encontradas lateralmente no epífalo e fazem parte do complexo fálico da genitália interna do macho (Amédégnato, 1976); a ausência de escleritos sustenta o clado R. balzapambae+, com reversões em C. bivittatus e S. volxemi.
28. Comprimento das valvas do edeago em relação ao arco do cíngulo: (0) curtas; (1) longas (ic=100; ir=100).
O comprimento das valvas do edeago pode variar em relação ao arco do cíngulo, estrutura encontrada no complexo fálico da genitália interna dos machos (AMÉDÉGNATO, 1976). As valvas longas do edeago em relação ao arco do cíngulo (Fig. 13) são uma condição sinapomórfica para A. trinitatis +.
29. Ápices das valvas do edeago: (0) divididos; (1) não-divididos (ic=50; ir=0).
Ápices das valvas do edeago não-divididos ocorrem homoplasicamente em C. bivittatus e P. vulneratus. Tanto em Abracrini (AMÉDÉGNATO & DESCAMPS, 1979) como em grande parte dos táxons do grupo interno, as valvas têm ápices divididos.
30. Bainha ectofálica do edeago: (0) ampla, englobando mais da metade do esclerito inferior do edeago; (1) estreita, englobando menos da metade do esclerito inferior do edeago (ic = 50; ir = 0).
O edeago apresenta a bainha ectofálica em duas formas: ampla (Fig.12) ou estreita (Fig.13) (COSTA & CARVALHO, 2006a,b). Bainha ectofálica estreita surge independentemente em E. vittatus e S. nigritus. Esse caráter auxilia também na sustentação de clados de Melanoplinae (CIGLIANO, 1997; CIGLIANO et al., 1996).
31. Forma do processo do gonóporo: (0) retangular; (1) digitiforme (ic = 50; ir = 0).
Na grande maioria dos gêneros do grupo Sitalces, a forma do processo do gonóporo é retangular (COSTA & CARVALHO, 2006a,b). A condição digitiforme ocorre homoplasicamente em R. balzapambae e C. bivittatus.
32. Valvas do ovipositor em vista lateral: (0) curtas; (1) longas (ic =50; ir = 66).
Valvas longas é uma sinapomorfia para todo o grupo Sitalces (COSTA & CARVALHO, 2006a,b), com reversão apenas em A. trinitatis.
33. Margem superior da valva dorsal do ovipositor: (0) mais de seis dentes; (1) menos de seis dentes (ic =50; ir = 66).
Margem superior da valva dorsal do ovipositor com menos de seis dentes (Fig. 14) surge independentemente em O. coryaceus e S. volxemi e a presença de mais de seis dentes na margem superior ocorre no restante dos táxons do grupo de estudo.
34. Dente apical externo da valva ventral do ovipositor: (0) proeminente; (1) não-proeminente (ic=50; ir=0).
A valva ventral do ovipositor pode apresentar um dente apical externo proeminente no (Fig. 15) ou não (Fig. 16). Dente não proeminente é uma sinapomorfia de S. nigritus + C. bivittatus + com reversão em P. vulneratus + P. sexnotata. CIGLIANO et al. (1996), nos seus estudos sobre os relacionamentos filogenéticos dos táxons Scotussa Giglio-Tos, 1894 e Leiotettix Bruner, 1906 (Melanoplinae) utilizaram esse caráter para auxiliar na sustentação dos clados.
Análise biogeográfica. As espécies do grupo Sitalces apresentam uma distribuição restrita à América do Sul (Fig. 17).
A região Neotropical foi dividida por Morrone (2001, 2004) em quatro sub-regiões biogeográficas (Caribenha, Amazônica, Chaquenha e Paranaense) e essas sub-regiões em províncias. As 11 espécies pertencentes ao grupo Sitalces distribuem-se nas sub-regiões Caribenha, Amazônica e Paranaense (Fig. 17).
Eusitalces vittatus distribui-se na sub-região Caribenha, no domínio do Noroeste da América do Sul, dentro da província de Llanos Venezolanos. Já na sub-região Amazônica a espécie distribui-se por seis províncias: Guyana, Guyana Húmeda, Imeri, Napo, Ucayali Roraima (Fig. 17).
Psiloscirtus olivaceus distribui-se na sub-região Amazônica, ocorrendo na província de Madeira situada no noroeste do Brasil, limitando-se com os rios Amazonas, Madeira, Beni e Xingu, e com a cordilheira oriental da Bolívia. Na província de Tapajós-Xingu, que compreende apenas o noroeste do Brasil, registra-se também esta espécie (Fig. 17).
O clado L. dorsualis + L. punctifrons ocorre nas sub-regiões biogeográficas Caribenhas e Amazônicas. Liebermannacris punctifrons ocorre nas províncias do domínio do Noroeste da América do Sul (Costa Venezolana e Llanos Venezolanos) alcançando a Província Guyana. Por outro lado, L. dorsualis apresenta uma ampla distribuição, ocorrendo no Brasil, Argentina, Equador, Peru e Bolívia (que abrangem parcialmente as sub-regiões Amazônica, Paranaense e Chaquenha) (Fig. 17).
Robustusacris balzapambae distribui-se na sub-região Caribenha, ocorrendo nas províncias de Cauca e Oeste do Equador, ambas pertencentes ao domínio do Noroeste da América do Sul. Entretanto, existe também o registro para a província de Páramo Norandino, que faz parte da zona de Transição Sul-americana.
Arimacris trinitatis ocorre exclusivamente na sub-região Caribenha, Província de Trinidad e Tobago, dentro do domínio do Noroeste da América do Sul.
O clado S. nigritus + C. bivittatus ocorre em duas sub-regiões Neotropicais. Salvadoracris nigritus ocorre na sub-região Amazônica, nas províncias de Madeira e Ucayali. Caruaruacris bivittatus distribui-se também na sub-região Paranaense, na província do Bosque Atlântico Brasileiro.
Sitalces volxemi apresenta distribuição restrita à sub-região Paranaense, ocorrendo na província do Bosque Atlântico Brasileiro.
Parasitalces vulneratus + P. sexnotata apresenta distribuição restrita à sub-região Amazônica (Fig. 17). Parasitalces vulneratus distribui-se tanto na sub-região Amazônica, província de Yungas, quanto na Zona de Transição Sul-americana (Puna). Parasitalces sexnotata ocorre exclusivamente na sub-região Amazônica, na província do Pantanal (centro sul do Brasil, noroeste da Bolívia e norte do Paraguai).
O cladograma de área do grupo Sitalces foi comparado com o modelo geral proposto por AMORIM & PIRES (1996). O evento vicariante proposto pela divergência entre os dois componentes (do noroeste e sudeste da América do Sul) durante o Cretáceo, com a formação de um lago ao longo dos rios Amazonas, Madeira e Mamoré, corrobora o encontrado no clado A. Com exceção de Caruaruacris bivittatus, o restante desse clado encontra-se no componente sul.
Os nós biogeográficos são de grande importância no contexto da história da diversificação dos táxons, pois estes podem estar relacionados a eventos vicariantes de biotas ancestrais que atualmente encontram-se em contato (CRISCI et al., 2003). MORRONE (2003), trabalhando com base em dados de Trichodactylidae (Crustacea), identificou quatro traços generalizados e três nós biogeográficos da região Neotropical. A distribuição de P. vulneratus + P. sexnotata corrobora o modelo de MORRONE (2003), estando ambos conectados pelo traço generalizado chaquenho. Este traço tem uma ligação histórica com o traço paranaense, hipótese sustentada pela relação de S. volxemi com P. vulneratus+, evidenciada pelo nó biogeográfico do oeste de Santa Catarina. A relação entre as biotas em que P. vulneratus + P. sexnotata se distribuem também foi evidenciada por NIHEI & CARVALHO (2005).
Apesar de um grande número de hipóteses a respeito da diversidade de táxons na Amazônia (HAFFER, 1997) e de vários trabalhos que identificam áreas de endemismo (CRACRAFT, 1985; SILVA & OREN, 1996; MORRONE, 2003), poucos inferem relações históricas entre estas áreas. Hipóteses primárias das relações das áreas utilizando o método panbiogeográfico aumentaram substancialmente nos últimos anos (MORRONE, 2003; NIHEI & CARVALHO, 2004), mas os testes destas hipóteses carecem utilização de método de biogeografia cladística para busca de homologias secundárias de áreas (MORRONE, 2004), tendo assim modelos mais consistentes das relações históricas das áreas.
Os estudos adicionais sobre as relações intercontinentais em Acrididae são fundamentais para melhor avaliar essas hipóteses e ampliar o conhecimento sobre a evolução e diversificação deste grupo de Orthoptera.
Agradecimentos. Ao Prof. Carlos Salvador Carbonell (Universidad de la Republica, Montevideo), pelo empréstimo do material. A Andressa Paladini pelas sugestões ao manuscrito. Ao CNPq pela concessão da bolsa de estudo ao primeiro autor que permitiu a realização desta pesquisa (Proc. 140687/2001-7).
Recebido em novembro de 2006. Aceito em abril de 2007.
- AMÉDÉGNATO, C. 1974. Les genres d'acridiens néotropicaux, leur classification par familles, sous-familles et tribus. Acrida 3:193-204.
- ___. 1976. Structure et evolution des genitalia chez les Acrididae et familles apparentées. Acrida 5:1-15.
- AMÉDÉGNATO, C. & DESCAMPS, M. 1979. Diagnoses génériques et affinités phylétiques d'Acridoidea néotropicaux récoltés par le Dr. Campos Seabra et M. Descamps (Orthoptera). Annales de la Société Entomologique de France (NS) 15(3):423-487.
- AMORIM, D. S. 1982. Classificação por seqüenciação: uma proposta para a denominação dos ramos retardados. Revista Brasileira de Zoologia 1(1):1-9.
- AMORIM D. L. & Pires, M. R. 1996. Neotropical Biogeography and a method for maximum biodiversity estimation In: BICUDO, C. E. M. & MENEZES, N. A. eds. Biodiversity in Brazil, a first approach. São Paulo, CNPq. p.183-219.
- AX, P. 1987. The phylogenetic system: the systematization of organisms on the basis of their phylogenesis Chichester, John Wiley. 340p.
- BRUNER, L. 1908. Acrididae. Biologia Centrali Americana 2:249-342.
- ___. 1911. I. South American Acridoidea. Annals of the Carnegie Museum 8(1):5-147.
- ___. 1919. I. Saltatorial Orthoptera from South America and the Isle of Pines. Annals of the Carnegie Museum 13(1-2):5-91.
- BRUNNER VON WATTENWYL, K. 1893. Révision du système des Orthoptères et description des espèces rapportés par M. Leonardo Fea de Birmanie. Annali del Museo Civico di Storia Naturale di Genova, ser. 2, 13:5-230.
- CIGLIANO, M. M. 1989. A cladistic analysis of the family Tristiridae (Orthoptera, Acridoidea). Cladistics 5(4):379-393.
- ___. 1997. Ronderosia, a new genus of South American Melanoplinae (Orthoptera, Acrididae). Journal of Orthoptera Research 6(1):1-19.
- CIGLIANO, M. M. & LANGE, C. E. 1988. Orthoptera. In: MORRONE, J. J. & COSCARÓN, S. Biodiversidade de Artrópodos Argentinos 1 ed. La Plata, SUR. 599p.
- CIGLIANO, M. M.; RONDEROS, R. & KEMP, W. P. 1996. Phylogenetic relationships of Scotussa and Leiotettix (Orthoptera, Acrididae). Cladistics 12(2):125-138.
- COSTA, M. K. M. & CARVALHO, G. S. 2006a. Salvadoracris e Caruaruacris, novos gêneros de gafanhotos sul-americanos (Orthoptera, Acrididae). Revista Brasileira de Entomologia 50(1):49-52.
- ___. 2006b. Revisão dos gêneros Sitalces, Eusitalces e Parasitalces (Orthoptera, Acrididae, Abracrini) e descrição de três novos gêneros. Revista Brasileira de Entomologia 50(2):137-152.
- CRACRAFT, J. 1985. Historical biogeography and patterns of differentiation within the South American avifauna: areas of endemism. In: BURCKLEY, P. A.; FOSTER, M. S; MORTON, E. S.; RIDGELY, R. S. & BURCKLEY, F. G. eds. Neotropical Ornithological Monographs 36, Washington, The American Ornithologists' Union. p.49-84.
- CRISCI, J. V.; KATINAS, L. & POSADAS, P. 2003. Historical biogeography: An Introduction Cambridge, Havard University. 250p.
- ELDREDGE, N. & CRACRAFT, J. 1980. Phylogenetic patterns and the evolutionary process New York, Columbia University. 349p.
- FARRIS, J. S. 1988. Hennig 86 reference Documentation for version 1.5. New York, Jefferson. 22p.
- ____. 1989. The retention index and rescaled consistency index. Cladistics 5(4):417-419.
- GIGLIO-TOS, E. 1898. Viaggio del Dr. Enrico Festa nella Republica dell' Ecuador e regioni vicine. VI, Ortotteri. Bollettino dei Musei di Zoologia ed Anatomia Comparata della R. Universitá di Torino 13(311):1-108.
- GUNTHER, K. 1940. Ueber die Verbreitung eininger Insekten im Gebiete des Amazonenstromes und die Frage eines columbischen Faunendistriktes in der brasilianischen Subregion. Archiv für Naturgeschichte (NF) 9(4):450-500.
- HAFFER, J. 1997. Alternative models of vertebrate speciation in Amazonia: an overview. Biodiversity and Conservation 6(3):451-476.
- HENNIG, W. 1966. Phylogenetic systematics Urbana, University of Illinois. 263p.
- KLUGE, A. G. & FARRIS, K. J. S. 1969. Quantitative phyletics and the evolution of anurans. Systematic Zoology 18(1):1-32.
- MORRONE, J. J. 2001. Homology, biogeography and areas of endemism. Diversity and Distributions 7(6):297-300.
- ___. 2003. Homología biogeográfica primaria de la familia Trichodactylidae (Crustacea: Decapoda). In: MORRONE, J. J. & LORENTE, J. eds. Una perspectiva latinoamericana de la Biogeografía. Mexico, Las Prensas de Ciencias, Facultad de Ciencias, UNAM. p.241-245.
- ___. 2004. Panbiogeografia, componentes bióticos y zonas de transición. Revista Brasileira de Entomologia 48(2):149-162.
- NIHEI, S. S. & CARVALHO, J. B. 2004. Taxonomy, cladistics and biogeography of Coenosopsia Malloch (Diptera, Anthomyiidae) and its significance to the evolution of anthomyiids in the Neotropics. Systematic Entomology 29(2):260-275.
- ___. 2005. Distributional patterns of Neotropical fly genus Polietina Schnabl & Dziedzichi (Diptera, Muscidae): a phylogeny-supported analysis using panbiogeographic tools. Papéis Avulsos de Zoologia 45(25):313-326.
- NIXON, K. C. & CARPENTER, J. M. 1993. On outgroups. Cladistics 9(4):413-426.
- PEREZ, D. G.; DOMINICI, O.; HIERRO, B. & OTTE, D. 1995. New grasshopper genera and species from the Dominican Republic (Hispaniola) (Acridoidea, Acrididae). Transactions of the American Entomological Society 121(3):153-171.
- PLATNICK, N. I. 1989. An empirical comparison of microcomputer parsimony programs II. Cladistics 5(2):145-161.
- REHN, A. G. 1917. On Orthoptera from the vicinity of Rio de Janeiro, Brazil. Transactions of the American Entomological Society 43(768):335-363.
- RONDEROS, R. A. & CIGLIANO, M. M. 1991. The Andean Dichroplini: cladistic analysis with description of Keyacris n. gen. and Ponderacris n. gen. (Orthoptera; Acrididae; Melanoplinae). Transactions of the American Entomological Society 11(3-4):167-191.
- SILVA, J. M. C. & OREN, D. C. 1996. Application of parsimony analysis of endemicity in Amazonian biogeography: an example with primates. Biological Journal of the Linnean Society 59(3):427-437.
- STÅL, C. 1878. Systema acridiodeorum. Essai d'une systematisation des acridoidées. Bihang till Kongliga Svenska Vetenskaps-akademiens Handlingar 5(4):1-100.
- WILEY, E. O. 1981. Phylogenetics. The theory and pratice of Phylogenetic Systematics New York, John Wiley and Sons. 439p.
Datas de Publicação
-
Publicação nesta coleção
30 Jan 2008 -
Data do Fascículo
Dez 2007
Histórico
-
Aceito
Abr 2007 -
Recebido
Nov 2006