Resúmenes
En el presente trabajo se describen las características morfohistológicas del testículo de individuos adultos (n=5) de Ceratophrys ornata (Bell, 1843) provenientes de humedales del centro de Argentina. Los mismos se procesaron mediante técnicas histológicas de rutina, se cortaron a 8 µm y las láminas obtenidas se tiñeron con hematoxilina-eosina y tricrómico de Masson. Las gónadas son órganos pares, amarillentos, alargados y contorneados de 18,58 ± 0,23 mm de largo por 1,51 ± 0,13 mm de ancho. Histológicamente se observa una delgada túnica albugínea (6,29 ± 0,83 µm) rodeando a los testículos. En su interior se hallan lóculos seminíferos que miden 240,64 ± 38,52 µm de diámetro, en ellos se distinguen cistos con células espermatogénicas en distintas etapas de desarrollo. El tejido intersticial es escaso y en él se destacan las células de Leydig y vasos sanguíneos. Las espermatogonias I son las células más grandes de la serie germinal (20,03 ± 2,27 µm); poseen la cromatina granular y de aspecto multilobular, hallándose comúnmente una por cisto, estas originan a las espermatogonias II, más pequeñas (12,06 ± 1,14 µm). Los espermatocitos I presentan la cromatina levemente condensada y son un poco más chicos que sus precedentes (11,64 ± 0,36 µm). Los espermatocitos II miden 8,85 ± 0,54 µm. Las espermátidas I son esféricas, miden 5,95 ± 0,42 µm y se agrupan en cistos redondeados. Las espermátidas II, en cambio son alargadas y no se hallan dentro de cistos, pero siguen organizadas en paquetes asociadas a células de Sertoli. Los espermatozoides son células libres hacia el centro del lóculo, alargadas, flageladas y con una notable compactación nuclear. La morfohistología de los testículos analizados muestran características macroscópicas e histológicas similares a las observadas en otras especies de anfibios anuros neotropicales, presentando todas las células del linaje espermatogénico en un mismo lóculo, lo que indicaría que presentan ciclos espermatogénicos continuos.
Células espermatogénicas; morfometría; histología
This paper describes the morphohistological features of the testes of adult Ceratophrys ornata (Bell, 1843) (n = 5) from a wetland agroecosystem of central Argentina. The testes were processed by routine histological techniques, sectioned at 8 µm. The sheets obtained were stained with hematoxylin-eosin and Masson trichrome. The gonads are paired, yellowish, elongated contoured organs, 18.58 ± 0.23 mm long and 1.51 ± 0.13 mm wide. Histologically, there is a thin tunica albuginea (6.29 ± 0.83 µm) surrounding the testes. Inside them, there are seminiferous locules measuring 240.64 ± 38.52 µm in diameter, distinguished in cysts with spermatogenic cells at different stages of development. The interstitial tissue is scarce and the Leydig cells and blood vessels stand out. Spermatogonia I are the largest cells of the germinal series (20.03 ± 2.27 µm), have granular chromatin and multilobular aspect, commonly found in each cyst, and they originate spermatogonia II, which are smaller (12.06 ± 1.14 µm). Spermatocytes I have slightly condensed chromatin and are also slightly smaller than its predecessors (11.64 ± 0.36 µm). Spermatocytes II measure 8.85 ± 0.54 µm. Spermatids I are spherical, measuring 5.95 ± 0.42 µm and are grouped in rounded cysts. In contrast, the spermatids II are elongate and not within the cysts, but they are organized in packets associated with Sertoli cells. Sperms are free, elongated cells oriented towards the center of the loculus, flagellated, and with a remarkable nuclear compaction. The morphohistology of testes analyzed shows macroscopic and histologic features similar to those observed in other species of neotropical anuran amphibians, presenting all espermatogenic lineage cells in a single, indicating that they have continuous spermatogenic cycles.
Spermatogenic cells; morphometry; histology
Morfohistología testicular de Ceratophrys ornata (Bell) (Amphibia, Anura, Ceratophryidae)
Testicular morphohistology of Ceratophrys ornata (Bell) (Amphibia, Anura, Ceratophryidae)
Fernando CarezzanoI,II; Sergio UrquizaII; Karina DorflingerI; Matías AlonsoII
ICátedra de Morfología Animal, Departamento de Diversidad Biológica y Ecología, Escuela de Biología, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Avda. Vélez Sarsfield 299, Córdoba (X5000JJC), Argentina
IICátedra de Anatomía Comparada, Departamento de Diversidad Biológica y Ecología, Escuela de Biología, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Avda. Vélez Sarsfield 299, Córdoba (X5000JJC), Argentina. (fercarezza@yahoo.com)
RESUMEN
En el presente trabajo se describen las características morfohistológicas del testículo de individuos adultos (n=5) de Ceratophrys ornata (Bell, 1843) provenientes de humedales del centro de Argentina. Los mismos se procesaron mediante técnicas histológicas de rutina, se cortaron a 8 µm y las láminas obtenidas se tiñeron con hematoxilina-eosina y tricrómico de Masson. Las gónadas son órganos pares, amarillentos, alargados y contorneados de 18,58 ± 0,23 mm de largo por 1,51 ± 0,13 mm de ancho. Histológicamente se observa una delgada túnica albugínea (6,29 ± 0,83 µm) rodeando a los testículos. En su interior se hallan lóculos seminíferos que miden 240,64 ± 38,52 µm de diámetro, en ellos se distinguen cistos con células espermatogénicas en distintas etapas de desarrollo. El tejido intersticial es escaso y en él se destacan las células de Leydig y vasos sanguíneos. Las espermatogonias I son las células más grandes de la serie germinal (20,03 ± 2,27 µm); poseen la cromatina granular y de aspecto multilobular, hallándose comúnmente una por cisto, estas originan a las espermatogonias II, más pequeñas (12,06 ± 1,14 µm). Los espermatocitos I presentan la cromatina levemente condensada y son un poco más chicos que sus precedentes (11,64 ± 0,36 µm). Los espermatocitos II miden 8,85 ± 0,54 µm. Las espermátidas I son esféricas, miden 5,95 ± 0,42 µm y se agrupan en cistos redondeados. Las espermátidas II, en cambio son alargadas y no se hallan dentro de cistos, pero siguen organizadas en paquetes asociadas a células de Sertoli. Los espermatozoides son células libres hacia el centro del lóculo, alargadas, flageladas y con una notable compactación nuclear. La morfohistología de los testículos analizados muestran características macroscópicas e histológicas similares a las observadas en otras especies de anfibios anuros neotropicales, presentando todas las células del linaje espermatogénico en un mismo lóculo, lo que indicaría que presentan ciclos espermatogénicos continuos.
Palabras-clave: Células espermatogénicas, morfometría, histología.
ABSTRACT
This paper describes the morphohistological features of the testes of adult Ceratophrys ornata (Bell, 1843) (n = 5) from a wetland agroecosystem of central Argentina. The testes were processed by routine histological techniques, sectioned at 8 µm. The sheets obtained were stained with hematoxylin-eosin and Masson trichrome. The gonads are paired, yellowish, elongated contoured organs, 18.58 ± 0.23 mm long and 1.51 ± 0.13 mm wide. Histologically, there is a thin tunica albuginea (6.29 ± 0.83 µm) surrounding the testes. Inside them, there are seminiferous locules measuring 240.64 ± 38.52 µm in diameter, distinguished in cysts with spermatogenic cells at different stages of development. The interstitial tissue is scarce and the Leydig cells and blood vessels stand out. Spermatogonia I are the largest cells of the germinal series (20.03 ± 2.27 µm), have granular chromatin and multilobular aspect, commonly found in each cyst, and they originate spermatogonia II, which are smaller (12.06 ± 1.14 µm). Spermatocytes I have slightly condensed chromatin and are also slightly smaller than its predecessors (11.64 ± 0.36 µm). Spermatocytes II measure 8.85 ± 0.54 µm. Spermatids I are spherical, measuring 5.95 ± 0.42 µm and are grouped in rounded cysts. In contrast, the spermatids II are elongate and not within the cysts, but they are organized in packets associated with Sertoli cells. Sperms are free, elongated cells oriented towards the center of the loculus, flagellated, and with a remarkable nuclear compaction. The morphohistology of testes analyzed shows macroscopic and histologic features similar to those observed in other species of neotropical anuran amphibians, presenting all espermatogenic lineage cells in a single, indicating that they have continuous spermatogenic cycles.
Keywords: Spermatogenic cells, morphometry, histology.
Desde hace años los anfibios se hallan en franco retroceso numérico a nivel mundial (Pounds & Crump, 1994; Houlahan et al., 2000; Lavilla, 2001; Kiesecker et al., 2001; Blaustein et al., 1994; 2003; Carey & Alexander, 2003; Daszak et al., 2003; Stuart et al., 2004; Beebee & Griffiths, 2005; McCallum, 2007; Roelants et al., 2007; Aguilar-Miguel et al., 2009). Entre las causas de esta disminución se cuentan la contaminación del agua y el suelo por agroquímicos, la destrucción y fragmentación del hábitat, la introducción de especies exóticas depredadoras, la radiación ultravioleta, la lluvia ácida, etc. (Lips et al., 1999). De continuar este fenómeno se convertiría en la sexta extinción masiva, abarcando a todo un grupo (Wake & Vredenburg, 2008), es por ello que conocer aspectos sobre la biología reproductiva de estos organismos en muy importante puesto que permiten diseñar e implementar estrategias y planes de acción para su protección y conservación.
La biología reproductiva de una especie es la combinación de características morfológicas, fisiológicas y conductuales que permiten obtener un número de descendientes óptimos y aptos bajo ciertas condiciones ambientales (Duellman & Trueb, 1994). Entre los ítems que se analizan se halla la morfohistología gonadal, ya que además de aportar información básica sobre la estructura y composición tisular de estos órganos, permite conocer las etapas del ciclo reproductivo de las especies (Oliveira et al., 2007).
Los ciclos reproductivos en los anuros se relacionan estrechamente con las condiciones ambientales tales como la temperatura, el fotoperíodo y las lluvias. Tales factores afectan la gametogénesis. Se han descrito tres ciclos espermatogénicos en los anfibios anuros en relación a factores ambientales y hormonales, el ciclo continuo, el potencialmente continuo y el discontinuo. En el primero la producción de espermatozoides se realiza ininterrumpídamente durante todo el año pudiendo presentar ciertos grados de variación estacional, histológicamente se observan en los lóculos seminíferos todo el linaje de las células espermatogénicas. Los ciclos potencialmente continuos se caracterizan por ser estacionales, presentando una moderada supresión de la espermatogénesis en los meses fríos, pero pueden ser estimulados intrínseca y extrínsecamente. Los ciclos discontinuos se ven interrumpidos durante parte del año, caracterizándose los testículos por presentar quistes germinales con células en degeneración durante el período de éstasis, no pudiéndose ser reactivados ni endógena ni exógenamente (Rastogi, 1976; Duellman & Trueb, 1994; Oliveira et al., 2002; Asenjo et al., 2011).
Estudios sobre la morfohistología de los testículos de anfibios neotropicales son escasos. En ellos se los describe como órganos pares, generalmente ovoides, localizados en la región abdominal y a ventral de los riñones. Están revestidos por la túnica albugínea de tejido conectivo, la cual puede o no presentar células pigmentarias. En su interior se observan lóculos seminíferos con células espermáticas en diferentes estadios de desarrollo, agrupadas en cistos y asociadas a células de Sertoli. Rodeando los lóculos se halla la túnica propia formada por células planas y tejido conectivo laxo. En el tejido intersticial se destacan las células de Leydig, secretoras de hormonas (Echeverría & Maggese, 1987; Rastogi et al., 1988; Báo et al., 1991; Oliveira et al., 2002; 2007; Oliveira & Zieri, 2005; Ferreira et al., 2008; 2009; Carezzano & Cabrera, 2010; Asenjo et al., 2011).
El objetivo del presente estudio es aportar datos básicos sobre la morfohistología del testículo de individuos de C. ornata capturados en plena época reproductiva en el centro de Argentina y así conocer parte de su ciclo reproductivo.
MATERIAL Y MÉTODOS
El presente estudio se realizó en humedales próximos a la localidad de Ucacha (33º01'S; 63º30'W), provincia de Córdoba, República Argentina, en la frontera de las ecorregiones del Espinal y la Pampa. Esta región presenta lagunas de origen pluvial y fluvial. El clima es estacional, presentando inviernos fríos y secos y veranos cálidos y lluviosos. La temperatura media anual es de 16ºC, variando de 9ºC en julio a 24ºC en enero. La precipitación media anual es de 850 mm y está concentrada entre octubre y marzo (Gorgas & Tassile, 2003; Aimar et al., 2010).
Se capturaron a mano cinco machos adultos de C. ornata entre el 20 y 22 de diciembre de 2011, en plena época reproductiva (Langone, 1994), localizándolos por sus vocalizaciones. Los animales colectados se acondicionaron en recipientes de plástico con un poco de agua para evitar la desecación y se transportaron al laboratorio. Al día siguiente se anestesiaron y eutanasiaron mediante vapores de cloroformo. Posteriormente se les efectuó una incisión en la pared abdominal y se extrajeron los órganos reproductores. Los mismos se fijaron en solución de Bouin durante 12 horas, conservándolos luego en alcohol etílico al 70%. Los especímenes se hallan depositados en la colección herpetológica de la Cátedra de Anatomía Comparada de la Facultad de Ciencias Exactas, Físicas y Naturales de la Universidad Nacional de Córdoba bajo la numeración AC-680 al 684.
Es de hacer notar que esta especie es considerada como no amenazada por Lavilla & Heatwole (2010), por lo que este estudio no implica un impacto negativo sobre las poblaciones de este anfibio.
Las gónadas se midieron en ambos ejes con un calibre digital de precisión 0,01 mm. Posteriormente se deshidrataron en alcoholes de graduación creciente, se diafanizaron en tres baños de xilol y finalmente se las incluyó en parafina. Se realizaron cortes seriados a 8 µm de espesor, los que se tiñeron con hematoxilina de Harris-eosina y tricrómico de Masson a los fines de identificar los distintos componentes tisulares.
Para el estudio morfohistológico se analizaron veinte cortes de cada espécimen a intervalos regulares, de modo de cubrir toda la longitud de la gónada, y con la ayuda de una cámara digital Zeiss AxioCam ERc 5s adosada a un microscopio trinocular Zeiss Primo Star se midieron las diferentes células utilizando el programa AxioVision 4.0 V 4.8.2.0. También se midió la túnica albugínea y el diámetro de los lóculos seminíferos. Para la identificación de los distintos tipos de células espermatogénicas se siguieron los trabajos realizados en anfibios de Oliveira et al. (2002), Jamieson (2003), Ferreira et al. (2008) y Carezzano & Cabrera (2010).
Las fotografías de las preparaciones histológicas se tomaron con una cámara Leica DFC 300 Fx, adosada a un microscopio Leica DM 4000 B.
RESULTADOS Y DISCUSIÓN
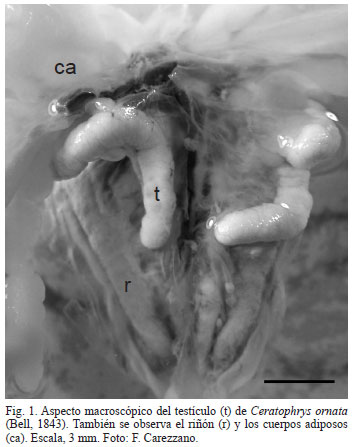
Los testículos de C. ornata son órganos pares, amarillentos, alargados y contorneados de 18,58 ± 0,23 mm de largo por 1,51 ± 0,13 mm de ancho (Fig. 1).
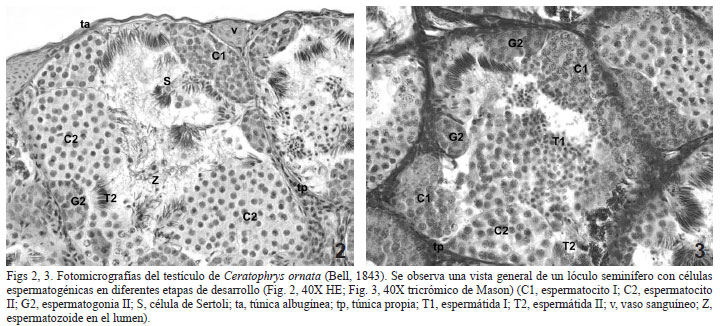
Histológicamente se observan rodeados por una delgada túnica albugínea (6,29 ± 0,83 µm) formada por tejido conectivo denso, en el que predominan fibras de colágeno. También se observan células de músculo liso y vasos sanguíneos. En su interior se hallan numerosos lóculos seminíferos que miden 240,64 ± 38,52 µm de diámetro, en ellos se distinguen cistos con células espermáticas en distintas etapas de desarrollo (Figs 2, 3). Rodeando los lóculos se halla la túnica propia, formada por tejido conectivo laxo y células planas, las células peritubulares. El tejido intersticial es escaso y en él se destacan células poliédricas con un núcleo central esférico, las células de Leydig y vasos sanguíneos (Figs 2, 4). No se observaron células pigmentarias.
En el interior de los lóculos se identifican las células espermatogénicas, en distintas etapas de desarrollo y asociadas a las células de Sertoli (Fig. 2).
Espermatogonias. Los cistos con espermatogonias diploides están próximos a la pared de los lóculos. Se pueden identificar espermatogonias I y espermatogonias II. Las primeras son las células más grandes (20,03 ± 2,27 µm); poseen escasa cromatina granular y son de aspecto multilobular, hallándose comúnmente una por cisto. Estas células por mitosis originan las espermatogonias II (Duellman & Trueb, 1994), más pequeñas (12,06 ± 1,14 µm) y con la cromatina más condensada ya que se preparan para la primer división meiótica (Rastogi et al., 1988; Jamieson, 2003) (Figs. 2, 3 y 4).
Espermatocitos. Células originadas por diferenciación de las espermatogonias II (Rastogi et al., 1988; Jamieson, 2003). Presentan la cromatina levemente condensada debido a que se hallan en las distintas fases de la primer división meiótica. Son un poco más pequeñas que sus precedentes (11,64 ± 0,36 µm). Los espermatocitos II originados por meiosis de los espermatocitos I (Rastogi et al., 1988; Jamieson, 2003), son aún más pequeños (8,85 ± 0,54 µm) y se observan más teñidos (Figs 2, 3).
Espermátidas. Células con una variedad de formas, desde esféricas a alargadas. Los espermatocitos II completan la meiosis II (Rastogi et al., 1988) y se transforman en espermátidas I, células esféricas agrupadas en cistos redondeados (5,95 ± 0,42 µm), las que se diferenciarán en las espermátidas II, células alargadas que no se hallan dentro de cistos, se organizan en paquetes asociadas a células de Sertoli (Figs 2, 3, 4).
Espermatozoides. Las espermátidas II sufren el proceso de espermiogénesis y originan a los espermatozoides, células alargadas, con una notable compactación nuclear y en donde se observa un flagelo. Se hallan libres en el lumen del lóculo (Fig. 2).
El análisis de los testículos observados indica que los cinco ejemplares estaban en la misma etapa gametogénica y muestran características macroscópicas e histológicas similares a las observadas en otras especies de anfibios anuros neotropicales (Oliveira et al., 2002; Jamieson, 2003; Oliveira & Zieri, 2005; Zieri et al., 2008; Carezzano & Cabrera, 2010). El hecho de exhibir lóculos con todas las células del linaje espermatogénico lo que indicaría que presentan ciclos continuos.
Estudios histológicos del ciclo reproductivo completo, no solo de esta especie, sino de otras simpátricas, y no solo analizando la espermatogénesis, sino también la oogénesis son necesarios para poder diseñar estrategias de manejo y conservación de las mismas, ya que los humedales en donde desarrollan sus actividades están siendo alterados por el hombre a causa de la expansión de la frontera agrícola. Esto esta provocando desecación extensiva de lagunas, simplificación de habitats y contaminación por agroquímicos, lo que indefectiblemente afectará las comunidades de estos anuros. Además permitirá extrapolar estos datos a especies en peligro actual, y sobre las que no se puede operar debido al riesgo de sacrificar sus efectivos.
Agradecimientos. A la Dra. Claudia Bregonzio por facilitar el microscopio Leica para obtener las fotografías de los cortes histológicos. Al Biólogo Cristian Lagger por su predisposición que permitió lograr la fotografía del aspecto macroscópico de las gónadas. A la Técnica de Laboratorio Ana Chalabe por realizar los preparaciones histológicas. A dos revisores anónimos por los comentarios y sugerencias realizadas.
Recibido 13 de abril de 2013.
Aceptado 27 de junio de 2013.
- Aguilar-Miguel, X.; Legorreta, G. & Casas-Andreu, G. 2009. Reproducción ex situ en Ambystoma granulosum y Ambystoma lermaense (Amphibia: Ambystomatidae). Acta Zoológica Mexicana 25(3):443-454.
- Aimar, L.; Bustamante, A.; Molina, C; Giovanola, C. & Menghi, M. 2010. Relación entre la limnología de lagunas pampeanas (SE Córdoba) y la cobertura y uso del suelo. RASADEP 1 - Número Especial. Cambios de uso de la tierra. Causas, consecuencias y mitigación. p.3-14.
- Asenjo, A.; Ting, K. S. & Pino, J. 2011. Morfometría de testículos y células germinales de Allobates femoralis (Boulenger 1883) (Dendrobatidae: Anura: Amphibia). The Biologist (Lima) 9(2):167-175.
- Báo, S. N.; Dalton, G. C. & de Oliveira, S. F. 1991. Spermiogenesis in Odontophrynus cultripes(Amphibia, Anura, Leptodactylidae): Ultrastructural and cytochemical studies of proteins using E-PTA. Journal of Morphology 207:303-314.
- Beebee, T. J. C. & Griffiths, R. A. 2005. The amphibian decline crisis: a watershed for conservation biology? Biological Conservation 125:271-285.
- Blaustein, A. R.; Romansic, J. M.; Kiesecker, J. M. & Hatch, A. C. 2003. Ultraviolet radiation, toxic chemicals and amphibian population declines. Diversity and Distributions 9:123-140.
- Blaustein, A.; Wake, D. & Sousa, W. 1994. Amphibian declines: judging stability, persistence, and susceptibility of populations to local and global extinctions. Conservation Biology 8:60-71.
- Carey, C. & Alexander, M. A. 2003. Climate change and amphibian declines: is there a link? Diversity and Distributions 9:111-121.
- Carezzano, F. J. & Cabrera. M. R. 2010. Morfohistología del testículo de Physalaemus biligonigerus (Amphibia, Leiuperidae) en un ecosistema agrícola. Boletín de la Asociación Herpetológica 21:39-42.
- Daszak, P.; Cunninngham A. A. & Hyatt, A. D. 2003. Infectious disease and amphibian population declines. Diversity and Distributions 9:141-150.
- Duellman, W. E. & Trueb, L. 1994. Biology of Amphibians London and Baltimore, The Johns Hopkins University Press. 670p.
- Echeverría, D. D. & Maggese, M. C. 1987. Desarrollo del testículo en Bufo arenerum (Anura, Bufonidae). Ciclos espermatogénicos y variaciones estacionales en los sapos juveniles. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia 14(9):125-138.
- Ferreira, A.; Mehanna, M. & Prados, C. P. A. 2008. Morphologic and morphometric analysis of testis of Pseudis limellum (Cope, 1862) (Anura, Hylidae) during the reproductive cycle in the Pantanal, Brazil. Biocell 32:185-194.
- Ferreira, A.; dos Santos Rosa, A. & Mehanna, M. 2009. Organização celular dos testículos em Hylidae e Leptodactylidae, no Pantanal (Estado do Mato Grosso do Sul, Brasil). Acta Scientiarum, Biological Sciencies 31(4):447-452.
- Gorgas, J. & Tassile, J. 2003. Recursos Naturales de la Provincia de Córdoba: Los Suelos, nivel de Reconocimiento 1:500.000. Agencia Córdoba Ambiente S. E., Instituto Nacional de Tecnología Agropecuaria EEA Manfredi.
- Houlahan, J. E.; Findlay, C. S.; Schmidt, B. R.; Meyer, A. H. & Kuzmin, S. L. 2000. Quantitative evidence for global amphibian population declines. Nature 404:752-755.
- Jamieson, B. G. M. 2003. Reproductive Biology and Phylogeny of Anura Plymouth, Science Publishers. 462p.
- Kiesecker, J. M.; Blaustein, A. R. & Belden, L. K. 2001. Complex causes of amphibian decline. Nature 410:681-684.
- Langone, J. A. 1994. Ranas y sapos del Uruguay. Reconocimiento y aspectos biológicos. Montevideo, Museo Dámaso Antonio Larrañaga, Intendencia Municipal de Montevideo (Serie Divulgación 5). 127p.
- Lavilla, E. O. 2001. Amenazas, declinaciones poblacionales y extinciones en anfibios argentinos. Cuadernos de Herpetología 15(1):59-82.
- Lavilla, E. O. & Heatwole, H. 2010. Status of amphibian conservation and decline in Argentina. In: Heatwole, H. ed. Amphibian biology. Status of decline of Amphibians: western hemisphere v. 9. Chipping Norton, Surrey Beatty & Sons. P. 30-78.
- Lips, K.; Reaser, J. K. & Young, B. E. 1999. El monitoreo de Anfibios en América Latina. Un manual para coordinar esfuerzos The Nature Conservancy. 42p.
- McCallum, M. 2007. Amphibian decline or extinction? Current declines dwarf background extinction rate. Journal of Herpetology 41:483-491.
- Oliveira, C.; Zanetoni, C. & Zieri, R. 2002. Morphological observations on the testes of Physalaemus cuvieri (Amphibia, Anura). Revista Chilena de Anatomía 20:263-268.
- Oliveira, C. & Zieri, R. 2005. Pigmentação testicular em Physalaemus nattereri (Steindachner) (Amphibia, Anura) com observações anatômicas sobre o sistema pigmentar extracutâneo. Revista Brasileira de Zoologia 22:454-460.
- Oliveira, E. F.; Feio, R. N. & Matta, S. L. P. 2007. Aspectos reprodutivos de Dendropsophus minutus (Peters, 1872) no município de Viçosa, Minas Gerais. Revista Ceres 54(313):230-238.
- Pounds, J. A. & Crump, M. L. 1994. Amphibian declines and climate disturbance: the case of the golden toad and the harlequin frog. Conservation Biology 8:72-85.
- Rastogi, R. K. 1976. Seasonal cycle in anuran (Amphibia) testis: the endocrine and environmental controls. Bolletino di Zoologia 43:151-172.
- Rastogi, R. K.; Bagnara, J. T.; Iela, L. & Krasovich, M. A. 1988. Reproduction in the mexican leaf frog, Pachymedusa dacnicolor. IV. Spermatogenesis: A light and ultrasonic study. Journal of Morphology 197:277-302.
- Roelants, K.; Gower, D. J.; Wilkinson, M.; Loader, S.; Biju, S. D.; Guillaume, K.; Moriau, L. & Bossuyt, F. 2007. Global patterns of diversification in the history of modern amphibians. Proceedings of the National Academy of Sciences 104:887-892.
- Stuart, S.; Chanson, J. S.; Cox, N. A.; Young, B. E.; Rodrigues, A. S. L.; Fishman, D. L. & Waller, R. W. 2004. Status and trends of amphibian declines and extinctions worldwide. Science 306:1783-1786.
- Wake, D. & Vredenburg, V. T. 2008. Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proceedings of the National Academy of Sciences of the United States of America 105(1):11466-11473.
- Zieri, R.; Taboga, S. R. & de Oliveira, C. 2008. Espermiogênese em Eupemphix nattereri (Anura, Leiuperidae): aspectos ultra-estruturais Iheringia,Série Zoologia 98(2):193-199.
Fechas de Publicación
-
Publicación en esta colección
08 Ago 2013 -
Fecha del número
Jun 2013
Histórico
-
Recibido
13 Abr 2013 -
Acepto
27 Jun 2013