Resumos
Este estudo explora a maturação de gametas e biometria de Phragmatopoma caudata Krøyer in Mörch, 1863 para endossar uma metodologia e oferecer uma técnica adequada para estudos que objetivam avaliar a ecologia populacional. A análise de correlação de Pearson confirmou a relação positiva (r = 0,90, P <0,0001) entre o comprimento do corpo e o comprimento da coroa opercular. Indivíduos com opercular crown < 0,9 mm podem ser considerados como juvenil devido à ausência de gametas. Portanto, utilizando-se o método aprovado para separar as classes de tamanho, a população dos recifes de P. caudata no Parque Estadual Xixová-Japuí (PEJX) na Baía de Santos, Estado de São Paulo, foi examinada durante dois anos, com o objetivo de analisar a densidade populacional e o padrão sazonal da classe juvenil. Em período de elevadas taxas de juvenis, a densidade populacional atingiu 128.115 ind./m², porém, a média foi 65.090±22.033 ind./m². As análises estatísticas (Kruskal-Wallis H = 18,475, p < 0,01) revelaram existir variação significativa na composição juvenil entre as estações chuvosa e seca. Apesar da presença de juvenis em meses de seca, as estações chuvosas contemplaram 92,1% dos juvenis amostrados. O padrão de juvenis observado pode estar relacionado com fatores biológicos (e.g. gametogênese e ciclo de vida) e abióticos (e.g. suprimento alimentar e correntes marinhas). Estes resultados destacam a necessidade de programas de monitoramento de longo prazo que integrem elementos ecológicos e abióticos, a fim de obter uma compreensão mais completa da ecologia desse poliqueta e ajudar a gerenciar a biodiversidade marinha do PEJX.
Morfometria; dinâmica de população; costão rochoso; bentos marinho
This study explores the achievement of gametes maturity and biometric analysis of Phragmatopoma caudata Krøyer in Mörch, 1863 to endorse a methodology and provide a proper technique for studies aiming to assess the population ecology. Analysis via Pearson's test confirmed a significant evidence of positive correlation (r = 0.90, P <0.0001) between body length and length of the opercular crown. Individuals with opercular crown < 0.9 mm can be regarded as juvenile due to the absence of gametes. Therefore, through the endorsed method to separate the size classes aimed to analyze the population density and seasonal pattern of the juvenile class. The reef population of P. caudata in the Xixová-Japuí State Park (PEJX) at the Santos Bay, state of São Paulo, was examined for two years. In the highest rate of juveniles, population density reached 128,115 ind./m², however, the average was 65,090 ind./m². Statistical analyzes (Kruskal-Wallis H = 1.475, p < 0.01) showed that significant temporal variation occurs in the juvenile composition between rainy and dry seasons. Despite the presence of representatives in the population during the dry months, rainy seasons encompassed 92.1% of juveniles sampled. The observed pattern of juveniles may be associated to biological factors (e.g. gametogenesis and life cycle) and abiotic (e.g. food supply and ocean currents). These results highlight the need for long-term monitoring programs that integrate ecological and abiotic elements to obtain a more complete understanding of the ecology of these worms and providing feedback for a better conservation of the marine biodiversity in the PEJX.
Morphometric analysis; population dynamics; rocky shore; benthic invertebrates
Larisse Faroni-PerezI, II
IPrograma de Pós-Graduação em Zoologia, Departamento de Zoologia, Instituto de Biociências, Universidade Estadual Paulista "Júlio de Mesquita Filho" - UNESP, Av. 24-A, 1515, 13506-900, Rio Claro, SP, Brasil
IIPrograma de Pós-Graduação em Ecologia - PPGECO, Departamento de Ecologia e Zoologia, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina, Campus Universitário, s/n, CCB, Córrego Grande, Florianópolis, SC, 88010-970, Brasil. (faroni.perez@gmail.com)
RESUMO
Este estudo explora a maturação de gametas e biometria de Phragmatopoma caudata Krøyer in Mörch, 1863 para endossar uma metodologia e oferecer uma técnica adequada para estudos que objetivam avaliar a ecologia populacional. A análise de correlação de Pearson confirmou a relação positiva (r = 0,90, P <0,0001) entre o comprimento do corpo e o comprimento da coroa opercular. Indivíduos com opercular crown < 0,9 mm podem ser considerados como juvenil devido à ausência de gametas. Portanto, utilizando-se o método aprovado para separar as classes de tamanho, a população dos recifes de P. caudata no Parque Estadual Xixová-Japuí (PEJX) na Baía de Santos, Estado de São Paulo, foi examinada durante dois anos, com o objetivo de analisar a densidade populacional e o padrão sazonal da classe juvenil. Em período de elevadas taxas de juvenis, a densidade populacional atingiu 128.115 ind./m2, porém, a média foi 65.090±22.033 ind./m2. As análises estatísticas (Kruskal-Wallis H = 18,475, p < 0,01) revelaram existir variação significativa na composição juvenil entre as estações chuvosa e seca. Apesar da presença de juvenis em meses de seca, as estações chuvosas contemplaram 92,1% dos juvenis amostrados. O padrão de juvenis observado pode estar relacionado com fatores biológicos (e.g. gametogênese e ciclo de vida) e abióticos (e.g. suprimento alimentar e correntes marinhas). Estes resultados destacam a necessidade de programas de monitoramento de longo prazo que integrem elementos ecológicos e abióticos, a fim de obter uma compreensão mais completa da ecologia desse poliqueta e ajudar a gerenciar a biodiversidade marinha do PEJX.
Palavras-chave: Morfometria, dinâmica de população, costão rochoso, bentos marinho.
ABSTRACT
This study explores the achievement of gametes maturity and biometric analysis of Phragmatopoma caudata Krøyer in Mörch, 1863 to endorse a methodology and provide a proper technique for studies aiming to assess the population ecology. Analysis via Pearson's test confirmed a significant evidence of positive correlation (r = 0.90, P <0.0001) between body length and length of the opercular crown. Individuals with opercular crown < 0.9 mm can be regarded as juvenile due to the absence of gametes. Therefore, through the endorsed method to separate the size classes aimed to analyze the population density and seasonal pattern of the juvenile class. The reef population of P. caudata in the Xixová-Japuí State Park (PEJX) at the Santos Bay, state of São Paulo, was examined for two years. In the highest rate of juveniles, population density reached 128,115 ind./m2, however, the average was 65,090 ind./m2. Statistical analyzes (Kruskal-Wallis H = 1.475, p < 0.01) showed that significant temporal variation occurs in the juvenile composition between rainy and dry seasons. Despite the presence of representatives in the population during the dry months, rainy seasons encompassed 92.1% of juveniles sampled. The observed pattern of juveniles may be associated to biological factors (e.g. gametogenesis and life cycle) and abiotic (e.g. food supply and ocean currents). These results highlight the need for long-term monitoring programs that integrate ecological and abiotic elements to obtain a more complete understanding of the ecology of these worms and providing feedback for a better conservation of the marine biodiversity in the PEJX.
Keywords: Morphometric analysis, population dynamics, rocky shore, benthic invertebrates.
Poliquetas representantes de Sabellariidae são tubícolas e algumas espécies constroem substratos biogênicos no entremarés (KIRTLEY, 1994). O local de assentamento das larvas fisiologicamente competentes é induzido pelo muco secretado por organismos pré-estabelecidos (JENSEN & MORSE, 1984; PAWLIK & FAULKNER, 1986). Este muco tem propriedades de importância medicinal (SHAO et al., 2009; WINSLOW et al., 2010), mas principalmente, promove o assentamento larval gregário dos sabelarídeos. Quando a densidade de tubos se torna elevada, estes constituem os substratos biogênicos (CALINE et al., 1992), conhecidos como recifes de areia. Em virtude desta peculiaridade de criar substratos tridimensionais, estes poliquetas são considerados como engenheiros de ecossistemas (JONES et al., 1994; COLEMAN & WILLIAMS, 2002). Os substratos biogênicos dos sabelarídeos aumentam a biodiversidade local (DUBOIS et al., 2002), servem de filtro biológico devido às altas densidades populacionais (CALINE et al., 1992) e reduzem os processos de erosão costeira por reterem grande quantidade de sedimentos (GRAM, 1968; KIRTLEY & TANNER, 1968; MAIN & NELSON, 1988).
Na costa brasileira, Phragmatopoma caudata Krøyer in Mörch, 1863 foi registrada desde as baixas latitudes (3°S) no estado do Ceará até as altas latitudes (32°S), no estado do Rio Grande do Sul (KIRTLEY, 1994; LANA & BREMEC, 1994). Embora a ocorrência geográfica seja ampla, pouco se conhece sobre a dinâmica populacional da espécie na costa brasileira. É plausível que o grau de dificuldade em obter os exemplares íntegros após a retirada do tubo, assim como a falta de metodologia específica, sejam fatores que limitaram o desenvolvimento de estudos de zoologia aplicada desses organismos.
Perante os fatos, o presente estudo objetivou utilizar a gametogênese e a biometria de Phragmatopoma caudata para aferir uma técnica metodológica com categorização de classes etárias. Em seguida, objetivou examinar a população da espécie no Parque Estadual Xixová-Japuí (PEXJ) (Baía de Santos, Estado de São Paulo, Brasil) para verificar valores de densidade populacional e o padrão temporal de abundância de juvenis. Este estudo tem como hipótese que o recrutamento de juvenis na população é temporalmente homogêneo, tendo-se em vista a premissa de que adultos exibem gametas durante todos os meses ao longo do ano (ECKELBARGER, 1976). Dada a falta de informações na literatura para P. caudata na costa do Brasil, a densidade e a sazonalidade no pico de juvenis registrados são comparados a outros sabelarídeos e discutidos no âmbito dos processos ecológicos dirigentes e ciclo de vida.
MATERIAL E MÉTODOS
Biometria e validação da metodologia para identificação das classes etárias. Os recifes de areia do entremarés rochoso de duas localidades (23°58'51.57"S, 46°22'02.64"O; 23°59'47.14"S, 46°23'22.97"O) na baía de Santos foram amostrados aleatoriamente. Após a desfragmentação manual dos tubos, obteve-se 237 indivíduos inteiros que foram utilizados nas análises. A fim de relacionar as medidas biométricas e a classe etária (juvenil/adulto) dos organismos, os indivíduos foram observados quanto à maturidade pela presença ou ausência de gametas. A presença de gametas pode ser visualmente identificada pela coloração dos indivíduos. Quando há gametas estocados no celoma, a coloração dos segmentos abdominais muda devido à transparência da parede do corpo (ECKELBARGER, 1976; KIRTLEY, 1994). Em cada exemplar de P. caudata foi mensurado o comprimento da coroa opercular (C.O.) e o comprimento do corpo dos indivíduos (C.C.), sendo o apêndice caudal excluído devido ao comprometimento da estrutura pela fragilidade, e logo, fácil ruptura ao manusear os indivíduos. A medida da C.O. foi realizada na posição ventro-dorsal e por vista superior tomada com microscópio estereoscópico com régua acoplada à lente ocular (0,1 mm). A medida do C.C. foi obtida com paquímetro digital. A fim de verificar a possível correlação morfométrica, foi empregado o teste de correlação de Pearson. A regressão linear foi utilizada para verificar relação entre o comprimento da coroa opercular (C.O.) e comprimento do corpo dos indivíduos (C.C.) (SOKAL & ROHLF, 1995). Foi utilizada a função y = bx + a, em que y = comprimento da coroa opercular; x = comprimento dos indivíduos; a = intercepção do logaritmo da regressão linear sobre "y"; b = coeficiente de crescimento relativo.
Variação interanual da população. Por dois anos, entre fevereiro de 2007 e janeiro de 2009, a população de P. caudata nos costões rochosos do Parque Estadual Xixová-Japuí, na praia de Itaquitanduva (23°59'41.33"S e 46°23'11.72"O até 23°59'58.71"S e 46°23'29.91"O) foi amostrada mensalmente. A coleta de material ocorreu durante as marés de sizígia e de modo aleatório por toda a extensão do substrato biogênico. A linha de costa amostrada foi de aproximadamente 632 m, sendo esta extensão descontínua devido à faixa de areia entre os matacões rochosos. O espaço amostral foi delimitado verticalmente pela amplitude de ocorrência dos recifes de areia, na zona do mediolitoral no entremarés. Cada estação considerada neste estudo compreende três meses consecutivos. Em cada mês foram obtidas 20 unidades amostrais com a utilização de um core de 3,45 cm2, inserido até 15 cm de profundidade com auxílio de martelo de borracha (51,9 cm3/core). O volume total amostrado por estação foi de 3.114 cm3. Após a coleta os fragmentos do recife de areia foram congelados até o momento da triagem. A triagem de cada unidade amostral consistiu na desfragmentação dos tubos manual e remoção de todos os exemplares de P. caudata, que foram posteriormente conservados em álcool 70-90%. Todos os indivíduos encontrados foram contabilizados para análises de densidade e tiveram o comprimento da C.O. mensurados. Posteriormente, a classe etária (juvenil/adulto) de cada indivíduo foi determinada conforme as medidas biométricas.
Com o objetivo de examinar a variação sazonal da população juvenil, os dados foram agrupados a cada três meses de modo a cobrir as duas estações chuvosa e seca (verão e inverno, respectivamente). Na área deste estudo, a sazonalidade de variáveis abióticas é conhecida (GIANNINI & PAIVA FILHO 1990; HARARI et al., 2008). Desta maneira, o intervalo temporal e as respectivas siglas utilizadas neste estudo são: Cf1 (Final da Chuvosa, ano I - fevereiro a abril de 2007); Si1 (Início da Seca, ano I - maio a julho de 2007); Sf1 (Final da Seca, ano I - agosto a outubro de 2007); Ci1 (Início da Chuvosa, ano I - novembro, dezembro de 2007 e janeiro de 2008); Cf2 (Final da Chuvosa, ano II - fevereiro a abril de 2008); Si2 (Início da Seca, ano II - maio a julho de 2008); Sf2 (Final da Seca, ano II - agosto a outubro de 2008); Ci2 (Início da Chuvosa, ano II - novembro, dezembro de 2008 e janeiro de 2009). Foi utilizado o teste não-paramétrico de Kruskal-Wallis (α = 0,01) para verificar diferenças significativas entre os grupos temporais. O teste de Dunn (α = 0,01) foi utilizado complementarmente para as comparações múltiplas.
RESULTADOS
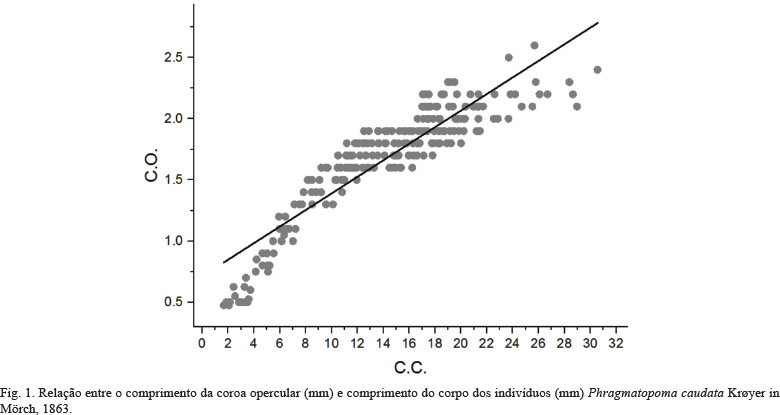
Validação da metodologia proposta. Em Phragmatopoma caudata a correlação de Pearson apresentou resultados estatisticamente significativos entre C.O. e C.C. (r = 0,90; p < 0,0001) (Fig. 1). A análise de regressão linear entre C.C. e C.O. indicou que o comprimento opercular cresce significativamente com o aumento do comprimento do corpo (R2 = 0,81; F1,235 = 1011; p < 0,0001) conforme a equação C.O. = 0,06744*C.C + 0,7175. Da mesma maneira, a morfometria do C.O. demonstrou ser um indicador válido para a avaliação da estrutura etária da população. Todos os espécimes com C.O. até 0,9 mm foram considerados juvenis, visto que não apresentaram produção evidente de gametas maduros. A partir da classe de tamanho C.O. = 1,0 mm foram observados gametas em alguns representantes. Deste modo, os subadultos e adultos foram agrupados na classe C.O. ≥ 1,0 mm.
Número de animais capturados e densidade. Durante os dois anos de amostragem de Phragmatopoma caudata foram examinados 10.779 indivíduos, 1.086 deles juvenis (10,08%). A estação Cf2 apresentou os meses com os picos máximos de juvenis na população estudada, compreendendo 51,01% do número total de juvenis observados por este estudo em ambos os anos de amostragem (Fig. 2). Portanto, o ano II abrangeu 75,23% do total de juvenis examinado, e evidenciou a disparidade na proporção interanual (Fig. 2). Ainda, o ano II contemplou 55,3% dos adultos observados.
A densidade populacional apresentou padrão sazonal distinto e entre os anos, conforme a variação observada durante o período de estudo (Fig. 3). A menor e a maior densidade populacional ocorreram na estação Cf. Ainda, a menor foi no ano I (março de 2007) com 39.420 ind./m2, enquanto a maior densidade ocorreu no ano II (março de 2008) quando atingiu 128.115 ind./m2. No total, a densidade média e o desvio padrão foram 65.090±22.033 ind./m2. Analisando-se a flutuação temporal da população, foi observado que valores elevados nas taxas de juvenis são concomitantes às elevadas taxas na densidade (Figs 3, 4). A figura 4 demonstra frequência média e o erro padrão de juvenis para cada estação. Ressalta-se que o maior número de juvenis ocorreu na estação chuvosa final do ano II (N=554), mais precisamente no mês de fevereiro. Ainda, a ocorrência de juvenis também foi alta no mês seguinte (março de 2008). Por outro lado, o total de juvenis foi extremamente baixo nas estações Sf1 e Si2 (N=12).
Estrutura da população segundo as classes etárias. Ao analisar a proporção interanual de juvenis, observou-se que houve alteração entre as estações (Fig. 2). A análise de variância não-paramétrica de Kruskal-Wallis (α = 0,01) demonstrou haver diferença temporal significativa no número de juvenis (H = 18,475, g.l. = 7, p < 0,01), evidenciando a ausência de similaridade sazonal para o padrão de juvenis na população. Aplicando-se o teste Dunn de comparações múltiplas (α = 0,01), ficaram evidentes as diferenças significativas entre os grupos Sf1 versus Cf2 e Cf2 versus Si2.
Considerando-se a frequência relativa de juvenis na população estudada, foi observado que as taxas estiveram sempre mais elevadas nas estações chuvosas (Ci e Cf) (Fig. 2). Neste sentido, estas estações chuvosas contemplam 92,1% dos juvenis amostrados em todo o estudo. No ano I, a estação Ci representou grande importância no recrutamento juvenil, contemplando 77.2% dos juvenis amostrados nas estações chuvosas e mais da metade (63%) dos exemplares obtidos (Fig. 2). Por outro lado, observou-se que as Cf (1 e 2) contemplam 55.6% dos juvenis registrados para as estações chuvosas. Analisando-se a flutuação interanual na proporção dos juvenis nas estações chuvosas, o ano II contempla 78% dos juvenis amostrados. Deste modo, ao avaliar a variabilidade entre os anos I e II, constatou-se que o número de juvenis capturados somente na estação Ci2 foi superior ao somatório das estações chuvosas no ano I e que o número de juvenis na estação Cf2 foi superior ao total de juvenis durante todas as estações do ano I (Fig. 2). Portanto, observou-se que, para o padrão de juvenis amostrado, houve variabilidade na flutuação intra- e interanual das estações chuvosas.
Dentre os 24 meses estudados, as maiores classes do comprimento opercular (C.O. = 2,9 - 3,0 mm) representando os adultos foram registradas somente em seis meses, sendo, à exceção do mês de outubro no ano I (Sf1), esta classe etária foi observada nas estações chuvosas (Ci1, Cf2, Cf2). Por outro lado, dentre os juvenis, das maiores classes de comprimento opercular (C.O. = 0,8 - 0,9 mm), as estações de seca representam apenas 8,2% do total observado para a classe etária, evidenciando baixas taxas de juvenis nos meses referentes a estas estações (Fig. 5). Constatou-se que as menores classes de comprimento opercular (C.O. = 0,3 - 0,5 mm), representando os juvenis recém-recrutados e após metamorfose, ocorreram em todas as estações no ano I e II. No entanto, sazonalmente esta classe etária não foi registrada na mesma proporção (Fig. 5). As estações de seca (Si e Sf) representam somente 7,5% do total observado para os juvenis recém-recrutados. Embora ocorrentes em todas as estações no ano I e II, as estações Si1 e Sf2 apresentaram baixas taxas de juvenis recém-recrutados (Fig. 5) e somente um representante dos juvenis recém-recrutados foi observado em cada uma das estações de seca Sf1 e Si2, evidenciando ausência de recém-recrutados em alguns meses referentes a estas estações (Fig. 5).
DISCUSSÃO
Metodologia proposta. O modelo biométrico proposto neste estudo para definir classes etárias de P. caudata apresenta correlação alta, e pode-se afirmar que 81% da variabilidade do comprimento opercular em função do comprimento do corpo é explicada mediante a reta de regressão correspondente. Deste modo, a metodologia pode ser utilizada por pesquisas no âmbito da biologia populacional. Durante o período do estudo foi possível mensurar todos os indivíduos, confirmando a eficácia da técnica biométrica proposta para estudos de dinâmica populacional e, principalmente, por minimizar a perda de dados biológicos durante a dissecação dos tubos. A C.O. é formada por rígidas cerdas modificadas (páleas) na região anterior do corpo, sendo assim a parte corpórea do animal mais resistente à manipulação e dissecção dos tubos.
Um estudo realizado com Sabellaria alveolata Linnaeus, 1767, com similar ocorrência na região entremarés e também construtora de recifes biogênicos, utilizou a biometria dos indivíduos em testes de correlação estatística (GRUET, 1984). No entanto, o autor utilizou medidas do diâmetro anterior do animal, dentre outras. Os gêneros Phragmatopoma e Sabellaria diferem-se no pedúnculo e na coroa opercular, sendo completamente fusionados apenas no primeiro caso (KIRTLEY, 1994). Neste sentido, para S. alveolata o autor selecionou o diâmetro do pedúnculo, parte abaixo da coroa opercular, e discutiu sobre esta ser uma parte do corpo resistente, cuja perda é menor no processo de dissecção dos tubos (GRUET, 1984). Ao dissecar os tubos de sabelarídeos, os exemplares podem ser facilmente rompidos, o que impossibilita medidas morfométricas em setígeros abdominais, um método comumente aplicado em outros poliquetas (PAGLIOSA & LANA, 2000; MACCORD & AMARAL, 2005; PARDO et al., 2010; OTEGUI et al., 2012). Deste modo, para estudos populacionais de Phragmatopoma, este estudo viabilizou uma técnica de morfometria viável e com acuidade estatística. É plausível ainda que a técnica metodológica apresentada por este estudo possa ser também aplicada em estudos populacionais de Neosabellaria e Gunnarea, pois, em ambos gêneros a coroa opercular também é fusionada.
Os resultados mostraram que, devido à ausência de gametas, P. caudata com C.O. < 0,9 mm podem ser considerados como juvenis. Contudo, é possível que dentre os juvenis, alguns animais das maiores classes de comprimento opercular (C.O. = 0,8 - 0,9 mm) poderiam ter fisiologicamente iniciado o processo de gametogênese. Embora seja desconhecido o ciclo reprodutivo e o crescimento da espécie para a localidade do presente estudo, na costa da Flórida ECKELBARGER (1976) estimou que P. lapidosa (sinônima de P. caudata) inicia a produção de gametas entre 6 a 8 semanas após o assentamento, mesmo que a sinonímia entre as populações do hemisfério norte e sul pode ser prematura (CAPA et al., 2012).
Densidade. No arquipélago Las Perlas, Panamá, o valor da densidade média do sabelarídeo Idanthyrsus cretus foi de 72.200 ind/m2 (BARRIOS et al., 2009). Na Baía de Mont Saint-Michel, França, a densidade de S. alveolata variou entre 15.000 até 60.000 ind/m2 (CALINE et al., 1992; DUBOIS et al., 2002). Em diferentes localidades na costa da Flórida, a densidade média de P. lapidosa variou entre 44.000 e 84.000 tubos/m2 (Daniel McCarthy, com. pess.) Por outro lado, FANTA (1968) estudou a espécie em três diferentes localidades no Estado de São Paulo e o número de tubos de P. lapidosa variou entre 500-2.700/m2. Ao contrastar a densidade da população estudada com outros sabelarídeos construtores de recifes, P. caudata apresentou valor médio inferior ao de I. cretus mas superior aos valores mínimo e máximo descritos para S. alveolata. Por outro lado, a densidade média observada esteve entre os valores encontrados para representativos do gênero na costa da Flórida, e superior aos anteriormente reportados para a costa de São Paulo. Ressalta-se ainda que, em estações de elevadas taxas de densidade foram observados 60-90 organismos por unidade amostral. Sabe-se que sabelarídeos são filtradores, cada indivíduo precisa da abertura do tubo em contato com a coluna dʼágua para obtenção de suprimento alimentar e sedimentar e que os tubos são individuais. Neste sentido, demonstra-se a importância de análises tridimensionais dos recifes para descrever como se dá o processo de construção de tubos individuais e a sobrevivência dos indivíduos nestes casos de elevadas taxas de densidade.
Estrutura da população. Este é o primeiro trabalho a utilizar biometria para averiguar as classes etárias e analisar a dinâmica populacional de P. caudata na costa brasileira. A população estudada mostrou variação interanual para a frequência dos juvenis nos recifes biogênicos. Deste modo, o padrão de juvenis na população não apresentou homogeneidade sazonal, sendo ainda evidente inconsistência interanual na abundância do recrutamento juvenil. Diante dos resultados obtidos, três possíveis cenários são plausíveis de advirem: (1) reprodução não contínua, (2) reprodução contínua com influência nos estágios pré-assentamento ou (3) reprodução contínua com influência nos estágios pós-assentamento, os quais serão discutidos posteriormente.
Nos dois anos de estudo, juvenis ocorreram em baixas taxas nas estações secas, representando apenas 7,8% do total observado. Em alguns meses das estações de seca somente as maiores classes de tamanho dos juvenis foram amostradas, o que sugere o recrutamento em baixa escala nos meses anteriores. Por outro lado, os resultados evidenciaram que as estações chuvosas (verão) representam grande importância para o recrutamento juvenil, visto como, representou o recrutamento massivo. Deste modo, presume-se a importância destas estações para a presença larvas no plâncton adjacente aos recifes. Ainda, os fatores ambientais comprimento do dia e temperatura da água do mar são importantes na sazonalidade da gametogênese da espécie (MCCARTHY et al., 2003). Sabe-se que o Sistema Estuarino de Santos contribui para a zona costeira adjacente com o aporte de nutrientes, matéria suspensa e clorofila a, sendo maior durante as estações chuvosas (MOSER et al., 2005), cujas temperaturas são significativamente maiores (HARARI et al., 2008). A biomassa fitoplanctônica na região é maior nos meses mais quentes (GIANESELLA-GALVÃO, 1982); sendo assim, é plausível que nas estações chuvosas, quando existe abundância de suprimento alimentar, haja um esforço reprodutivo maior - tanto para os adultos investirem na gametogênese como para o desenvolvimento das larvas planctônicas. Embora elevadas taxas de juvenis tenham sido observadas nos meses das estações chuvosas, estas não foram proporcionalmente similares nos dois anos. Este resultado mostra grandes variações no esforço reprodutivo de ano para ano, como reportado para S. alveolata (DUBOIS et al., 2007). Portanto, o padrão obtido indica existência de ciclos interanuais em maior escala de tempo, refutando-se alusões de recrutamento intranual e interanual homogêneo nos substratos biogênicos de sabelarídeos.
(1) Reprodução não contínua. Na população de P. caudata, valores elevados de densidade juvenil ocorreram sazonalmente nos 24 meses estudados. A sazonalidade observada entre as densidades de juvenis pode não refletir um mecanismo, em nível populacional, de reprodução contínua. No hemisfério norte, a variação temporal no assentamento larval de sabelarídeos foi também registrada, mostrando concordância com os dados aqui obtidos. Em estudo de 28 meses na costa da Flórida, apenas quatro eventos de assentamento ocorreram para P. lapidosa (sinônima de P. caudata) (ECKELBARGER, 1976). No Oceano Pacífico, Phragmatopoma californica Fewkes, 1889 também apresentou denso recrutamento sazonal (TAYLOR & LITTLER, 1982). Ainda que P. lapidosa tenha habilidade para produzir gametas por todo o ano (ECKELBARGER, 1976), a presença de gametas não necessariamente implica que os indivíduos se reproduzem. Em S. alveolata, WILSON (1968) estimulou a liberação de gametas e observou o progresso da fertilização pelos diferentes estágios de maturação do ovócito, concluindo que apenas óvulos maduros apresentam requisitos necessários para completar o desenvolvimento larval. Em vista da semelhança na ovogênese em sabelarídeos (ECKELBARGER, 1988), é provável que o mesmo ocorra em outros taxa. Neste sentido, os resultados mostram evidência para refutar a teoria de reprodução contínua. Picos sazonais de juvenis na população podem evidenciar liberação não contínua de gametas, como reportado para S. alveolata (DUBOIS et al., 2007). Levando-se em conta o resultado do presente estudo, um possível cenário de sincronismo para a liberação de gametas pela população poderia ser considerado tendo-se em vista que as estações chuvosas incorporaram mais de 90% do total de juvenis. Contudo, deve ser salientado que essa é apenas uma hipótese plausível de ser futuramente testada. Não foram encontrados estudos que abordem o ritmo de emissão de gametas e o tempo de desenvolvimento no plâncton de P. caudata na costa do Brasil. Desta maneira, apesar de detectado picos de recrutas nas estações chuvosas e devido à limitação dos dados deste estudo, é difícil inferir sobre quais são os processos biológicos que possam estar associados ao padrão temporal de juvenis observado.
(2) Reprodução contínua com influência nos estágios pré-assentamento. Muitos invertebrados marinhos bentônicos são considerados como 'populações abertas' devido aos recrutas serem muitas vezes efetivos imigrantes de outras fontes (GAINES & ROUGHGARDEN, 1985; UNDERWOOD & FAIRWEATHER, 1989). A dinâmica de populações destes organismos envolve mecanismos de transporte e aptidão larval que asseguram a conectividade entre as diferentes subunidades (BHAUD, 2000; PINEDA, et al., 2007). A dispersão e migração dos primeiros estágios de vida, o comportamento e transporte larval estão associados aos processos pré-assentamento e são dirigentes importantes para a dinâmica populacional (PINEDA et al., 2007; SHANKS, 2009). O potencial de dispersão das larvas em invertebrados marinhos pode ser bem variável, desde unidades centimétricas, centenas, ou até milhares de metros (ROUGHGARDEN et al., 1988; MORGAN & SHEPHERD, 2006; SHANKS, 2009). Algumas explicações traçam relações ciclo de vida e distribuição vertical das larvas na coluna d'água (BHAUD, 2000; SHANKS, 2009). Por outro lado, alguns estudos salientaram que os processos oceanográficos são significativos no processo de transporte ou na retenção das larvas (DUBOIS et al., 2007; PINEDA et al., 2007; AYATA et al., 2009). Os sabelarídeos têm potencial de serem classificados como 'populações abertas' por apresentarem ciclo de vida com estágios ontogenéticos planctônicos onde as larvas podem permanecer no plâncton durante 2 a 10 semanas (ECKELBARGER, 1976; DUBOIS et al., 2007), ter períodos de inanição (starvation) (WILSON, 1968), e definir o local de assentamento em resposta a indutores químicos de coespecíficos (JENSEN & MORSE, 1984; PAWLIK & FAULKNER, 1986). Sabe-se que as larvas de sabelarídeos podem utilizar as cerdas provisórias como mecanismos de defesa à predação (PENNINGTON & CHIA, 1984).
Com base nesse modelo, teríamos então de esperar que populações de sabelarídeos sejam do tipo abertas. Entretanto, em S. alveolata um padrão incomum contrasta com o que seria esperado para as larvas planctônicas de longo período de desenvolvimento. Na Baía de Saint Michel, as larvas permaneceram perto das áreas de assentamento, evitando assim a dispersão offshore (DUBOIS et al., 2007). Os autores evidenciaram a importância das correntes de marés residuais para a retenção das larvas. Posteriormente, na mesma localidade, a distribuição espacial das larvas foi modelada sob condições hidroclimáticas realistas e os resultados evidenciaram que além do elevado auto-recutamento, o hidrodinamismo da baía pode favorecer a migração larval entre recifes biogênicos separados por 15 km (AYATA et al., 2009). A baixa abundância larval observada no estudo nas estações de seca (inverno) poderia ser resultante dos processos oceanográficos, já que, nestes meses ocorrem períodos de instabilidade atmosférica com ventos fortes do sul, sobretudo entre agosto e outubro (HARARI et al., 2008), denominados estação Sf neste estudo. Ressalta-se que foi totalizado para as estações Sf (1 e 2) sete juvenis da classe etária recém-recrutados. Portanto, correntes marinhas resultantes dos fortes ventos comuns do sul podem ter favorecido a migração e dispersado as larvas sentido norte, corroborando assim a baixa densidade larval analisada. Deste modo, é plausível que estes eventos abióticos extremos sejam pulsos para o transporte e a dispersão larval de P. caudata, e, portanto, proporcionam entre populações a conectividade populacional e o fluxo gênico.
Atualmente, para a maioria dos ecologistas a definição de uma população refere-se a "um grupo de indivíduos coespecíficos que habitam uma dada área" (CAMUS & LIMA, 2002). No entanto, uma vez que o intervalo de dispersão larval é desconhecido para P. caudata e a conectividade de subpopulações é claramente um elemento essencial da dinâmica de metapopulações, permanece a questão de como é que diferentes cenários de dispersão larval podem influenciar o tamanho espacial da população. Embora o principal problema na aplicação do conceito de sistemas marinhos seja delinear para as espécies, as unidades populacionais e sua escala espacial (locais e regionais), para aplicação do conceito de conservação marinha e ecologia de metapopulação a estrutura espacial precisa ser claramente definida (CAMUS & LIMA, 2002). A conectividade das populações de P. caudata pode ser regulada por fatores biológico (e.g. ciclo de vida) e ambiental (e.g. ventos, correntes, e geomorfologia da linha de costa). A dispersão ou a retenção das larvas dependerá dos processos de pré-assentamento mas não há estudos in situ que considere a distribuição vertical dos estágios larvais e os processos oceanográficos físicos em condições extremas. Esta integração de conhecimento é um processo contínuo e crescentes esforços são necessários para compreender o movimento entre populações de P. caudata.
(3) reprodução contínua com influência nos estágios pós-assentamento. Os resultados deste estudo ainda revelaram que após elevadas taxas na densidade populacional de adultos e juvenis existe declínio, porém, os dados não permitem inferir sobre existência de controle populacional pela densidade, predação ou qualquer outro fator ambiental (e.g. meteorológico, poluição química). Processos pós-assentamento são importantes na estrutura de populações de invertebrados sésseis, que podem sofrer eventos de flutuações importantes no espaço e no tempo (BARROS, 2005; HELMUTH et al., 2006). Uma das causas mais comuns dos eventos de flutuações temporais são os gargalos populacionais por predação. Na costa do Chile foi constatado que Phragmatopoma virgini Kinberg, 1867 teve diminuição da abundância juvenil produzida pela mortalidade pós-assentamento, a qual foi induzida por consumidores móveis (ZAMORANO et al., 1995). É desconhecida a tolerância e os níveis de estresse em P. caudata para as condições ambientais na baía de Santos e os dados deste estudo permitem apenas sugestivas relações à dinâmica populacional observada. Atividades antropogênicas têm uma série de efeitos sobre a densidade e diversidade de organismos nos costões rochosos (CROWE et al., 2000). Sabe-se que P. lapidosa pode tolerar curtos períodos de assoreamento, turbidez e aumento dos níveis de sulfeto (MAIN & NELSON 1988; SLOAN & IRLANDI, 2008). O Sistema Estuarino de Santos é uma região poluída com passivos ambientais, visto que a água, os sedimentos e os organismos habitantes apresentam algum tipo de contaminação (ABESSA et al., 1998; BRAGA et al., 2003; SOUSA et al., 2007). Deste modo, relações interespecíficas e condições ambientais inadequadas podem ser co-reguladores da dinâmica populacional dos construtores de recifes de areia na baía de Santos.
A análise do padrão de juvenis na população de P. caudata do Parque Estadual Xixová-Japuí demonstrou picos sazonais, registrados nas estações chuvosas. Os dados evidenciaram a ocorrência de importante evento de assentamento larval com êxito nos primeiros estágios do recrutamento, visto que uma única estação chuvosa respondeu por mais da metade dos juvenis analisados. Sendo assim, é plausível que este tipo de evento seja responsável pela conservação da população no PEXJ, a qual é registrada pela ciência desde os anos 1960 (KIRTLEY, 1994). Estudos suplementares são necessários para melhor compreensão da ecologia dos recifes de sabelarídeos na área visando demonstrar os benefícios fornecidos à biota local pela presença dos engenheiros de ecossistema e consequente formação de substrato biogênico. Ainda, é imprescindível estimar a importância da população do PEXJ como fonte de recrutas para populações de outras áreas. Deste modo, para que haja adequado gerenciamento dos substratos biogênicos e consequente biodiversidade marinha do PEXJ, este estudo sugere que sejam realizadas novas pesquisas de ecologia experimental para averiguar se a fauna se favoreceria pela existência dos recifes, assim como aprofundar sobre as taxas de produção de gametas e os processos ecológicos de pré-assentamento e pós-assentamento de P. caudata. Os resultados obtidos poderão ser utilizados como instrumento de manejo e auxiliar no gerenciamento da biodiversidade marinha do PEJX.
Agradecimentos. A Fundação de Amparo à Pesquisa do Estado de São Paulo pela bolsa de mestrado (FAPESP/Proc. 07/56340-3). Ao Centro de Biologia Marinha da Universidade de São Paulo (CEBIMar/Proc. 2008/04) e a National Geographic Society (Proc. 8447-08) pelo apoio. Ao Dr. Daniel McCarthy, que gentilmente permitiu citar seus dados em preparação.
Recebido em 13 de maio de 2013
Aceito em 27 de fevereiro de 2014
- ABESSA, D. M. S; SOUSA, E. C. P. M.; RACHID, B. R. F. & MASTROTI, R. R. 1998. Use of the burrowing amphipod Tiburonella viscana as a tool in marine sediments contamination assessment. Arquivos de Biologia e Tecnologia 41:225-230.
- AYATA, S. D.; ELLIEN, C.; DUMAS, F.; DUBOIS, S. & THIÉBAUT, E. 2009. Modelling larval dispersal e settlement of the reef-building polychaete Sabellaria alveolata: role of hydroclimatic processes on the sustainability of biogenic reefs. Continental Shelf Research 29:1605-1623.
- BARRIOS, L. M.; CHAMBERS, S. J.; ISMAIL, N.; GUZMAN, H. M & MAIR, J. M. 2009. Distribution of Idanthyrsus cretus (Polychaeta: Sabellariidae) in the Tropical Estern Pacific and application of PCR-RAPD for population analysis. Zoosymposia 2:487-503.
- BARROS, F. 2005. Evaluating the importance of predation on subtidal benthic assemblages in sandy habitats around rocky reefs. Acta Oecologica 27:211-223.
- BHAUD, D. 2009 Two contradictory elements that determine invertebrate recruitment: dispersion of larvae e spatial restrictions on adults. Oceanologica Acta 23:409-422.
- BRAGA, E. S.; FONSECA, A. L. O.; BOSQUILHA, G. E.; DUCATTI, G. M. F.; AGUIAR, V. M. C.; LIMA, C. A. C. & ARASAKI, E. 2003. Eutrophication and bacterial pollution assessment risks on the Santos Bay sandy beaches (Brazil): influence of seasonal conditions. Journal of Coastal Research 35:516-524.
- CALINE, B.; GRUET, Y.; LEGENDRE, C.; LE RHUN, J.; L'HOMER, A.; MATHIEU, R. & ZBINDEN, R. 1992. The Sabellariid reefs in the bay of Mont Saint-Michel, France. Ecology, Geomorphology, Sedimentology, and Geologic Implications Stuart, Florida Oceanographic Society, Contributions to Marine Science.156p.
- CAMUS, P. A. & LIMA, M. 2002. Populations, metapopulations, and the open-closed dilemma: the conflict between operational and natural population concepts. Oikos 97:433-438
- CAPA, M.; HUTCHINGS, P. & PEART, R. 2012. Systematic revision of Sabellariidae (Polychaeta) and their relationships with other polychaetes using morphological and DNA sequence data. Zoological Journal of the Linnean Society 164:245-284.
- COLEMAN, F. C. & WILLIAMS, S. L. 2002. Overexploiting marine ecosystem engineers: potential consequences for biodiversity. Trends in Ecology & Evolution 17:40-44.
- CROWE, T. P.; THOMPSON, R. C.; BRAY, S. & HAWKINS, S. J. 2000. Impacts of anthropogenic stress on rocky intertidal communities. Journal of Aquatic Ecosystem Stress and Recovery 7:273-297.
- DUBOIS. S.; COMTET, T,; RETIÈRE, C. & THIÉBAUT, E. 2007. Distribution e retention of Sabellaria alveolata larvae (Polychaeta: Sabellariidae) in the Bay of Mont Saint Michel, France. Marine Ecology Progress Series 346:243-254.
- DUBOIS, S.; RETIERE, C. & OLIVIER, F. 2002. Biodiversity associated with Sabellaria alveolata (Polychaeta: Sabellariidae) reefs: effects of human disturbances. Journal of the Marine Biological Association of the United Kingdom 82:817-826.
- ECKELBARGER, K. J. 1976. Larval development and population aspects of reef-building polychaete Phragmatopoma lapidosa from east coast of Florida. Bulletin of Marine Science 26:117-132.
- _____. 1988. Oogenesis and female gametes. In: WESTHEIDE, W. & HERMANS, C. O. eds. The Ultrastructure of Polychaeta Stuttgart, New York, Microfauna Marina 4. p.281-307.
- FANTA, E. S. 1968. Sôbre a biologia e ecologia de Phragmatopoma lapidosa (Sabellariidae, Polychaeta). Ciência e Cultura 20:327-328.
- GAINES, S. & ROUGHGARDEN, J. 1985. Larval settlement rate: a leading determinant of structure in an ecological community of the marine intertidal zone. Proceedings of the National Academy of Sciences 82:3707-3711.
- GIANESELLA-GALVÃO, S. M. F. 1982. Standing-stock and potential of phytoplankton production in the Bay of Santos, Brazil. Boletim do Instituto Oceanográfico 31:85-94.
- GIANNINI, R. & PAIVA FILHO, A. M. 1990 Os Sciaenidae (Teleostei: Perciformes) da Baía de Santos (SP), Brasil. Boletim do Instituto Oceanográfico 38:69-86.
- GRAM, R. 1968. A Florida Sabellariidae reef and its effect on sedment distribution. Journal of Sedimentary Petrology 38:863-868.
- GRUET, Y. 1984. Granulometric evolution of the sand tube in relation to growth of the polychaete annelid Sabellaria alveolata (Linne) (Sabellariidae). Ophelia 23:181-193.
- HARARI, J., FRANÇA, C. A. S. & CAMARGO, R. 2008. Climatology and hydrography of Santos Estuary. In: RAMIRO, N. R.; BARETTA, J. & MATEUS, M. eds. Perspectives on Integrated Coastal Zone Management in South America Lisboa, IST Press. p.147-160.
- HELMUTH, B.; BROITMAN, B. R.; BLANCHETTE, C. A.; GILMAN, S.; HALPIN, P.; HARLEY, C. D. G.; O'DONNELL, M. J.; HOFMANN, G. E.; MENGE, B. & STRICKLAND, D. 2006. Mosaic patterns of thermal stress in the rocky intertidal zone: Implications for climate change. Ecological Monographs 76:461-479.
- JENSEN, R. A. & MORSE, D. E. 1984. Intraspecific facilitation of larval recruitment - gregarious settlement of the polychaete Phragmatopoma californica (Fewkes). Journal of Experimental Marine Biology and Ecology 83:107-126.
- JONES, C. G.; LAWTON, J. H. & SHACHAK, M. 1994. Organisms as ecosystem engineers. Oikos 69:373-386.
- KIRTLEY, D. W. 1994. A review and taxonomic revision of the Family Sabellariidae Johnston, 1865 (Annelida: Polychaeta). Vero Beach, Sabecon Press. 223p.
- KIRTLEY, D. W. & TANNER, W. F. 1968. Sabellariid worms - builders of a major reef type. Journal of Sedimentary Petrology 38:73-78.
- LANA, P. C. & BREMEC, C. S. 1994. Sabellariidae (Annelida, Polychaeta) from South America. Mémoires du Muséum national d'Histoire naturelle. Série A, Zoologie 162:211-222.
- MACCORD, F. S. & AMARAL, A. C. Z. 2005. Morphometric analyses of two species of Scolelepis (Polychaeta: Spionidae). Journal of Experimental Marine Biology and Ecology 85:829-834.
- MAIN, M. B. & NELSON, W. G. 1988. Sedimentary characteristics of sabellariid worm reefs (Phragmatopoma lapidosa Kinberg). Estuarine Coastal and Shelf Science 26:105-109.
- MCCARTHY, D. A.; YOUNG, C. M. & EMSON, R. H. 2003. Influence of wave-induced disturbance on seasonal spawning patterns in the sabellariid polychaete Phragmatopoma lapidosa Marine Ecology Progress Series 256:123-133.
- MORGAN, L. E. & SHEPHERD, S. A. 2006. Population and spatial structure of two common temperate reef herbivores: abalone and sea urchins In: KRITZER, J. P. & SALE, P. F. eds. Marine metapopulations. Amsterdam, Elsevier. p.205-246
- MOSER, G. A. O.; GIANESELLA, S. M.; ALBA, J. J.; BERGAMO, A. L.; SALDANHA-CORRÊA, F. M. P.; MIRANDA, L. B. & HARARI, J. 2005 Instantaneous transport of salt, nutrients, suspended matter and chlorophyll-a in the tropical estuarine system of Santos. Brazilian Journal of Oceanography 53:115-127.
- OTEGUI, M. P.; BLANKENSTEYN, A. & PAGLIOSA, P. 2012. Population structure, growth and production of Thoracophelia furcifera (Polychaeta: Opheliidae) on a sandy beach in Southern Brazil. Helgoland Marine Research 66:479-488.
- PAGLIOSA, P. R. & LANA, P. C. 2000. Population dynamics and secondary production of Nereis oligohalina (Nereididae: Polychaeta) from a subtropical marsh in southeast Brazil. Bulletin of Marine Science 67:259-268.
- PARDO, E. V.; TEIXEIRA, L. L. S. & AMARAL, A. C. Z. 2010. Morphometric analysis of Capitella capitata (Polychaeta, Capitellidae). Iheringia, Série Zoologia 100:13-18.
- PAWLIK, J. R. & FAULKNER, D. J. 1986. Specific free fatty acids induce larval settlement and metamorphosis of the reef-building tube worm Phragmatopoma califomica (Fewkes). Journal of Experimental Marine Biology and Ecology 102:301-310.
- PENNINGTON, J. T. & CHIA, F.-S. 1984. Morphological and behavioral defenses of trochophore larvae of Sabellaria cementarium (Polychaeta) against four planktonic predators. Biological Bulletin 167:168-175.
- PINEDA, J.; HARE, J. A. & SPONAUGLE, S. 2007. Larval transport and dispersal in the coastal ocean and consequences for population connectivity. Oceanography 20:22-39.
- ROUGHGARDEN, J.; GAINES, S. & POSSINGHAM, H. 1988. Recruitment dynamics in complex life cycles. Science 241:1460-1466.
- SHANKS, A. L. 2009. Pelagic larval duration and dispersal distance revised. The Biological Bulletin 216:373-385.
- SHAO, H.; BACHUS, K. N. & STEWART, R. J. 2009. A water-borne adhesive modeled after the sandcastle glue of P. californica Macromolecular Bioscience 9:464-471.
- SOKAL, R. R. & ROHLF, F. J. 1995. Biometry: the principles and practice of statistics in biological research 3 ed. New York, W. H. Freeman and Company. p.451-544.
- SOUSA, E. C. P. M.; ABESSA. D. M. S.; RACHID, B. R. F.; GASPARRO, M. R. & ZARONI, L. P. 2007. Ecotoxicological assessment of sediments from the port Santos and the disposal sites of dredged material. Brazilian Journal of Oceanography 55:75-81.
- TAYLOR, P. R. & LITTLER, M. M. 1982. The roles of compensatory mortality, physical disturbance, and substrate retention in the development and organization of a sand-influenced, rocky-intertidal community. Ecology 63:135-146.
- UNDERWOOD, A. J. & FAIRWEATHER, P. G. 1989. Supply-side ecology e benthic marine assemblages. Trends in Ecology and Evolution 4:16-20.
- WILSON, D. P. 1968. Some aspects of the development of eggs and larvae of Sabellaria alveolata (L.). Journal of the Marine Biological Association of the United Kingdom 48:367-386.
- WINSLOW, B. D.; SHAO, H.; STEWART, R. J. & TRESCO, P. A. 2010. Biocompatibility of adhesive complex coacervates modeled after the sandcastle glue of Phragmatopoma californica for craniofacial reconstruction. Biomaterials 31:9373-9381.
- ZAMORANO, J. H.; MORENO, C. A. & DUARTE, W. E. 1995. Post-settlement mortality in Phragmatopoma virgini (Polychaeta, Sabellariidae) at the Mehuin Marine Reserve, Chile. Marine Ecology-Progress Series 127:149-155.
Variação sazonal no recrutamento de Phragmatopoma caudata (Polychaeta, Sabellariidae) na costa sudeste do Brasil: biometria e validação de metodologia para categorização de classes etárias
Datas de Publicação
-
Publicação nesta coleção
27 Jun 2014 -
Data do Fascículo
Mar 2014
Histórico
-
Recebido
13 Maio 2013 -
Aceito
27 Fev 2014