
ABSTRACT
The genus Bostryx Troschel, 1847 is endemic to South America, extending from Ecuador to Chile and Argentina. The southernmost Argentinian species of the genus that inhabit San Luis, San Juan and Mendoza provinces, specially the pre-Andes, Andes and Sierras Pampeanas mountain ranges, were examined. This is the first time the anatomy of Bostryx pastorei (Holmberg, 1912), Bostryx reedi (Parodiz, 1947) and Bostryx strobeli (Parodiz, 1956) has been described.Bostryx cordillerae (Strobel, 1874) is re-described regarding shell and anatomy due to new morphological data. The main differences among the species examined are based on shell characters. The distribution ofBostryx mendozanus (Strobel, 1874) and Bostryx cuyanus (Pfeiffer, 1867), other species found in this region, was also discussed.
KEYWORDS
Stylommatophora; Precordillera; Sierras Pampeanas; systematics
RESUMEN
El género Bostryx Troschel, 1847 es endémico de Sudamérica, extendiéndose desde Ecuador hasta Chile y Argentina. Las especies argentinas más australes habitan en las provincias de San Luis, San Juan y Mendoza, especialmente en la precordillera y cordillera Andina y las Sierras Pampeanas, fueron examinadas. Se describe por primera vez la anatomía de Bostryx pastorei (Holmberg, 1912), Bostryx reedi (Parodiz, 1947) y Bostryx strobeli (Parodiz, 1956). Bostryx cordillerae (Strobel, 1874) es redescripta en conchilla y anatomía con nuevos datos morfológicos. Las principales diferencias entre las especies examinadas están basadas en caracteres de conchilla. La distribución deBostryx mendozanus (Strobel, 1874) y Bostryx cuyanus (Pfeiffer, 1867), otras especies encontradas en esta región, es también discutida.
PALABRAS-CLAVE
Stylommatophora; Precordillera; Sierras Pampeanas; sistemática
The genus Bostryx Troschel, 1847 is endemic to South America, extending from Ecuador to Chile, along the Pacific coastal desert and the western Andean slopes. In contrast, in Argentina, Paraguay and Bolivia, it is mainly found east of the Andes mountain range. The Cordillera de los Andes runs along the Pacific coast of South America from Venezuela to Argentina, and it shows a relative decrease in height from North to South. The Andes have a direct mechanical influence on the climatology of South America due to that induced precipitations along the eastern flanks of the Andes and blocking western currents from the Pacific (Inselet al., 2009Insel, N.; Poulsen, C. J. & Ehlers, T. A. 2009. Influence of the Andes Mountains on South American moisture transport, convection, and precipitation. Climate Dynamics 35:1477-1492. ). This contrast determines a continuous and arid vegetation belt (around 30°S) that crosses diagonally from SE to NW, separating the region of the Chilean-Argentinian southern forests from other forests in South America, the so called Arid Diagonal of South America (Villagran & Hinojosa, 1997Villagran, C. & Hinojosa, L. F. 1997. Historia de los bosques del sur de Sudamérica, II: Análisis fitogeográfico. Revista Chilena de Historia Natural 70:241-267.; Abrahan de Vazquezet al., 2000Abraham de Vazquez, E. M.; Garleff, K.; Liebricht, H.; Regairaz, A. C.; Schäbitz, F.; Squeo, F. A.; Stingl, H.; Veit, H. & Villagrán, C. 2000. Geomorphology and paleoecology of the arid diagonal in Southern South AmericaZeitschrift für Angewandte Geologie: SonderheftSH 1:55-61.). In Argentina, Bostryx is the genus of Bulimulidae with most species (Cuezzoet al., 2013Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.). Of the 19 species found in the country, 17 are endemic whereas Bostryx stelzneri (Dohrn, 1875) is distributed throughout Bolivia and Bostryx torallyi (d'Orbigny, 1835) throughout Bolivia, Paraguay and Peru (Quintana, 1982Quintana, M. 1982. Catálogo preliminar de la malacofauna del Paraguay. Revista del Museo Argentino de Ciencias Naturales , Zoología 11(3):61-158.; Ramírez et al., 2003Ramírez, R.; Paredes, C. & Arenas, j. 2003. Moluscos del Perú. Revista de Biología Tropical 51(3):225-284.). All Argentinian species of the genus have originally been described based on shell characters and only a few descriptions include jaw and radula. Anatomical information is known only for 9 species (Hylton Scott, 1954Hylton-Scott, M. I. 1954. Una nueva especie deBostryx (Bulimulidae) (Mol.Pulm.). Physis 20(59):409-413.;Breure, 1978Breure, A. S. H. 1978. Notes on and descriptions of Bulimulidae (Mollusca, Gastropoda). Zoologische Verhandelingen 164:1-255.; Miranda & Cuezzo, 2014Miranda, M. J. & Cuezzo, M. G. 2014. Taxonomic revision of theBostryx stelzneri Species complex, with description of a new species (Gastropoda: Orthalicoidea: Bulimulidae). American Malacological Bulletin 32(1):74-93.; Miranda, 2015Miranda, M. J. 2015. Bostryx tororanus (Doering, 1879) species complex (Gastropoda: Stylommatophora: Bulimulidae), a review of taxonomy and distribution of endemic species from Argentina. Journal of Natural History 49:995-1022. ).
The provinces of Mendoza, San Juan and San Luis are located in the central west of Argentina and cover an area of approximately 315.226 km2, between 28°22'-33°17'S and 66°20'-70°36'W. This region has arid climatic conditions with low relative humidity and limited water availability. Between 29°S and 33°S latitude, the Andean Cordillera of San Juan and Mendoza provinces is characterized by parallel mountain ranges with North-South direction: The Cordillera Principal (spanning Argentina and Chile), the Cordillera Frontal, the Argentinian Precordillera and the Sierras Pampeanas (from west to east) (Siame et al., 2006Siame, L. L.; Bellier, O. & Sebrier, M. 2006. Active tectonics in the Argentine precordillera and western Sierras Pampeanas. Revista de la Asociación Geológica Argentina 61(4):604-619.). The Precordillera extends for about 450 km in the West of Argentina (28°-33°S), from La Rioja to south of the Mendoza river; peaks can reach altitudes between 3,000 m-4,900 m (Roig-Juñentet al., 2003Roig-Juñent, S.; Flores, G. E. & Mattoni, C. 2003. Consideraciones biogeográficas de la Precordillera (Argentina), con base en artrópodos epígeos. : Morrone, J. & Llorente-Bousquets, J. eds. Una perspectiva latinoamericana de la Biogeografía. México, D.F., Las Prensas de Ciencias, Facultad de Ciencias, UNAM. 307p.). The southern part of the Precordillera comprises the Sierra de Tontal, Sierra de Villavicencio and Sierra de Uspallata (Furque & Cuerda, 1979Furque, G. & Cuerda, A. J. 1979. Precordillera de La Rioja, San Juan y Mendoza. Segundo Simposio Geología Regional Argentina 1:455-522.). The eastern slopes are more humid because they receive winds from the Atlantic Ocean that provide water and even form banks of fog. The western slopes are drier because they receive dry winds that may even reduce the little water available in the environment (Roig-Juñent et al., 2003Roig-Juñent, S.; Flores, G. E. & Mattoni, C. 2003. Consideraciones biogeográficas de la Precordillera (Argentina), con base en artrópodos epígeos. : Morrone, J. & Llorente-Bousquets, J. eds. Una perspectiva latinoamericana de la Biogeografía. México, D.F., Las Prensas de Ciencias, Facultad de Ciencias, UNAM. 307p.). The Uspallata-Calingasta valley is a small area located between the Andean Cordillera (5,000-6,000 m) and the Precordillera, from northern San Juan to north of Mendoza province, with very scarce vegetation (Roig-Juñentet al., 2001Roig-Juñent, S.; Flores, G.; Claver, S.; Debandi, G. & Marvaldi, A. 2001. Monte Desert (Argentina): Insect biodiversity and natural areas. Journal of Arid Environments 47(1):77-94.). According to the classification proposed by Olson & Dinerstein (1998Olson, D. M. & Dinerstein, E. 1998. The Global 200: A Representation Approachto Conserving the Earth's Most Biologically Valuable Ecoregions. Conservation Biology 12:502-515.) and Olson et al. (2001)Olson, D. M.; Dinerstein, E.; Wikramanayake, E. D.; Burgess, N. D.; Powell, G. V. N.; Underwood, E. C.; D'Amico, J. A.; Itoua, I.; Strand, H. E.; Morrison, J. C. et al. 2001. Terrestrial Ecoregions of the world: A new map of Life on Earth. BioScience 51(11):933-938., this region is characterized by the presence of seven dry ecoregions (Southern Andean Steppe, Central Andean Puna, High and Low Monte, Dry Chaco, Patagonian Steppe and Espinal). In this region the genus Bostryx has a notably high richness, and is represented by 9 of its 19 Argentinian species (Miquel, 1993Miquel, S. E. 1993. Las especies del género BostryxTroschel, 1847 en la República Argentina (1ra. parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde 121:157-171., 1995; Cuezzo et al., 2013Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.).
The objectives of the present study are to provide information on the anatomy of four species from central west in Argentina and review the taxonomic status and distribution area of all species of this genus in this region. This is the first time Bostryx pastorei (Holmberg, 1912), B. reedi (Parodiz, 1947) andB. strobeli (Parodiz, 1956) have been described based on anatomical systems and B. cordillerae (Strobel, 1874) is newly described incorporating data from new characters.
MATERIAL AND METHODS
The specimens examined come from Argentinian malacological collections: Instituto Fundación Miguel Lillo (IFML-Moll, Tucumán), Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (MACN-In, Buenos Aires), Museo de La Plata (MLP, La Plata) and Museo de Ciencias Naturales "José Lorca" (Mendoza, MCNL). Shell measurements (mm), such as total shell height (tsh), body whorl height (bwh), spire height (sh), major diameter of the shell (masd), minor diameter (misd), shell apertural height (ah), shell apertural diameter (ad), parietal space length (psl), penultimate whorl height (pwh) and major diameter (pwd), were obtained using a calliper (Figs 1, 2). Measurements of the perimeter and area of the shell and parts of the shell (Figs 3-5) were obtained from digital images of specimens in standardised apertural view (axis of coiling and aperture parallel to the camera lens). The images were used to construct Tps files with TpsUtil version 1.58 (Rohlf, 2013aRohlf, F. J. 2013a. ´TpsUtil. version 1.58`. Available at <Available at http://life.bio.sunysb.edu/morph/
>. Accessed on January 2015.
http://life.bio.sunysb.edu/morph/...
) software. Shell outline drawings of all species were carried out with TpsDig2 version 2.17 (Rohlf, 2013b_____. 2013b. ´TpsDig2. version 2.17`. Available at <http://life.bio.sunysb.edu/morph/>. Accessed on January 2015.
http://life.bio.sunysb.edu/morph/...
) directly on the shell photograph with the Draw Curves tool, using a line of dots around the outline. This outline allows computation of the enclosed area (cm²) and the perimeter (cm) of the outline. Anatomical systems were dissected out from alcohol-fixed specimens. Anatomical description is given in full for Bostryx reedi while only the main differences are presented for the remaining species.
The geographic distribution map was produced using Diva-Gis Software v.7.5.0.0 (Hijmans et al., 2007Hijmans, R.; Guarino, L.; Jarvis, A & O'Brien, R. 2007. DIVA-GIS Version 7.3.0.1. Available at <Available athttp://www.diva-gis.org/
>. Accessed on January 2015.
http://www.diva-gis.org/...
). Ecoregional' delimitations are according to Olson & Dinerstein (1998Olson, D. M. & Dinerstein, E. 1998. The Global 200: A Representation Approachto Conserving the Earth's Most Biologically Valuable Ecoregions. Conservation Biology 12:502-515.) and Olsonet al. (2001)Olson, D. M.; Dinerstein, E.; Wikramanayake, E. D.; Burgess, N. D.; Powell, G. V. N.; Underwood, E. C.; D'Amico, J. A.; Itoua, I.; Strand, H. E.; Morrison, J. C. et al. 2001. Terrestrial Ecoregions of the world: A new map of Life on Earth. BioScience 51(11):933-938..

Measurements of the shell: 1, 2, linear measurements; 3, total shell perimeter; 4, spire and body whorl perimeter; 5, aperture perimeter.
RESULTS
Bostryx reedi (Parodiz, 1947)
Peronaeus (Lissoacme) reediParodiz, 1947_____. 1947. Contribuciones al conocimiento de los moluscos terrestres Sudamericanos, V. Comunicaciones Zoológicas del Museo de Historia Natural de Montevideo2:1-32.:10; 1957b:24; 1961:61; Fernández, 1973Fernández, D. & Castellanos, Z. A. 1973. Clave genérica de la malacofauna terrestre Argentina. Revista del Museo Argentino de Ciencias Naturales 11:263-285. :95; Tablado & Mantinian, 2004Tablado, A. & Mantinian, J. 2004. Catálogo de ejemplares tipo de la División Invertebrados del Museo Argentino de Ciencias Naturales. II. Mollusca. Revista del Museo Argentino de Ciencias Naturales 6(2):363-384. :375.
Bostryx reediBreure,1979_____. 1979. Systematics, Phylogeny and Zoogeography of Bulimulinae (Mollusca). Zoologische Verhandelingen168:1-215.:57;Miquel, 1993Miquel, S. E. 1993. Las especies del género BostryxTroschel, 1847 en la República Argentina (1ra. parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde 121:157-171.:162; Cuezzo et al., 2013Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.:143.
Distribution. Endemic to High Monte ecoregion of the precordillera (pre-Andes) of Mendoza Province (32°S, 68°-69°W) (Fig. 6). The Andean piedmont of Mendoza, located between latitudes 32°-34°S, is situated within the domain of the South America Arid Diagonal.

Geographical distribution of the southernmost Argentinian species of the genus Bostryx (Gastropoda, Bulimulidae).
Type material. Holotype, ARGENTINA, Mendoza: Cordillera de Mendoza, La Cueva, VI.1919, Reed C. col. (MACN-In 10001).
Other material examined. ARGENTINA, Mendoza: Cordillera de Mendoza, La Cueva, VI.1919, Reed C. col. (MACN-In 10001-1); Guamparito, 3,000 m, Reed C. col. (MACN-In 10001); Cerro Pelado (MACN-In 9592-1); Cerro Pelado, 3,400 m, 1954, Roig F. col. (MCNL 1116).
Shell. Small, elongate-ovate, thick (Figs 7, 8). Six convex whorls. Shell cream, first whorls darker (Fig. 7). Protoconch with thin axial shallow ribs and slightly wavy and fine spiral grooves crossing them (Figs 10, 11). Spire high conic, acute, body whorl tall in relation to the total height of the shell, 60% of the total length (Figs 7, 8). Teleoconch with oblique elevated shallow ribs, separated by regular wide spaces (Fig. 9). Suture simple, deep (Fig. 9). Aperture ovate, narrow and short, 40% of the total shell length (Figs 7, 8). Parietal space narrow, with thin parietal callus in some specimens (Figs 7, 8). Peristome simple, not expanded (Fig. 7). Umbilicus wide (Fig. 7). Holotype measurements. masd= 7.12, misd= 6.86, tsh= 13.33, bwh= 8.83, sh= 5.46, ah= 5.75, ad= 4.19. Additional measurements in Tabs I,II.

Bostryx reedi (Parodiz, 1947). 7, holotype (MACN 10001) in ventral, dorsal and laterals view; 8, size and shape variation of shells (MCNL 1116) (scale bar: 1 cm).

Bostryx reedi (Parodiz, 1947). 9, general view of the protoconch and 3 first whorls (scale bar: 200 µm); 10, 11, detail of sculpture of protoconch (scale bar: 100 and 20 µm, respectively).
Pallial system. Kidney triangular, 1/3 of pulmonary roof length. Primary ureter bordering kidney along its length and secondary ureter opening at distal portion of pulmonary roof. Ureteric interramus rectangular in shape. Rectum opening at mantle collar. Main pulmonary vein well marked, parallel to rectum.
Reproductive system. Albumen gland bean-shaped (Fig. 12). Hermaphroditic duct divided into thin proximal and distal portions and convoluted seminal vesicle (Fig. 12). Fertilization pouch-spermathecal complex finger-shaped. Bursa copulatrix sac rounded and duct long reaching middle portion of albumen gland (Fig. 12), inner wall with longitudinal zigzag folds. Vagina cylindrical, 1/3 of penis length (Fig. 12), inner wall with thick longitudinal straight folds. Penial retractor muscle inserting terminally in flagellum. Flagellum shorter than epiphallus, inner wall with diagonal folds and central longitudinal fold (Fig. 14). Epiphallus same length as penis, progressively increasing its width towards penis. Proximal epiphallus internal wall sculpture with parallel straight folds, zigzag folds in the distal portion (Fig. 14). Penis with thinner section, which divides it into swollen proximal and cylindrical distal portion (Fig. 13). Proximal portion penis inner wall with relaxed longitudinal zigzag folds, very thin, followed by smooth area and penial gland triangular (Fig. 14). Distal portion with inner relaxed longitudinal zigzag folds (Fig. 14). Penial sheath with retractor muscle on upper end overlapping distal portion of penis. Vas deferens inserting at basal penial sheath.
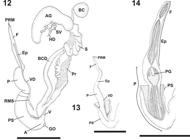
Bostryx reedi (Parodiz, 1947). 12, View of dissected reproductive system; 13, phallic complex without penial sheat; 14, phallic complex inner wall (scale bar: 5 mm) (A, atrium; AG, albumen gland; BC, bursa copulatrix; BCD, bursa copulatrix duct; Ep, epiphallus; F, flagellum; GO, genital opening; HD, hermaphroditic duct; P, penis; PG, penial gland; Pr, prostate; PS, penial sheath; PRM, retractor muscle; S, spermoviduct; SV, seminal vesicle; V, vagina; VD, vas deferens).
Remarks. According to labels included in lot MACN 9592-1, Bostryx reedi was identified by Doello Jurado as a new species, under the provisional name of Bulimulus sanzini. However, Parodiz (1947_____. 1947. Contribuciones al conocimiento de los moluscos terrestres Sudamericanos, V. Comunicaciones Zoológicas del Museo de Historia Natural de Montevideo2:1-32.) described and published this new species under the name of Peronaeus (Lissoacme) reedi. Later Breure (1979_____. 1979. Systematics, Phylogeny and Zoogeography of Bulimulinae (Mollusca). Zoologische Verhandelingen168:1-215.) classified it underBostryx, which was maintained by Miquel (1993Miquel, S. E. 1993. Las especies del género BostryxTroschel, 1847 en la República Argentina (1ra. parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde 121:157-171.) and Cuezzoet al. (2013Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.) subsequently.
Bostryx reedi is one of the smallest species of the genus. The convex whorls of the spire, presence of darker first whorls, marked parietal callus and wide umbilicus are characters that clearly distinguish B. reedifrom other species.
Shell measurements (ad, shell apertural diameter; ah, shell apertural height; bwh, body whorl height; masd, major diameter of the shell; misd, minor diameter; psl, parietal space length; pwd, penultimate whorl major diameter; pwh, penultimate whorl height; sh, spire height; tsh, shell height).
Shell measurements (n, number of measured specimens; SD, standard deviation; Min., minimum; Max., maximum; Mean, average).
Bostryx pastorei (Holmberg, 1912)
Bulimulus (Mesembrinus) pastoreiHolmberg, 1912Holmberg, E. L. 1912. Moluscos argentinos en parte nuevos coleccionados por Franco Pastore. Physis 1:19-26.:22.
Bulimulus (Scansicochlea) pastoreiParodiz, 1946Parodiz, J. J. 1946. Los géneros de los Bulimulinae Argentinos. Revista Museo de La Plata (N. S.) 4(zool. 30):303-371.:315; 1957a:134; Fernández, 1973Fernández, D. & Castellanos, Z. A. 1973. Clave genérica de la malacofauna terrestre Argentina. Revista del Museo Argentino de Ciencias Naturales 11:263-285. :85.
Peronaeus (Lissoacme) puntanusParodiz, 1947_____. 1947. Contribuciones al conocimiento de los moluscos terrestres Sudamericanos, V. Comunicaciones Zoológicas del Museo de Historia Natural de Montevideo2:1-32.:13; Miquel, 1995_____. 1995. Las especies del género BostryxTroschel, 1847 en la República Argentina (2da. y última parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde124:119-127.:121; Tablado & Mantinian, 2004Tablado, A. & Mantinian, J. 2004. Catálogo de ejemplares tipo de la División Invertebrados del Museo Argentino de Ciencias Naturales. II. Mollusca. Revista del Museo Argentino de Ciencias Naturales 6(2):363-384. :375.
Bostryx puntanusBreure, 1979_____. 1979. Systematics, Phylogeny and Zoogeography of Bulimulinae (Mollusca). Zoologische Verhandelingen168:1-215.:57.
Bostryx pastoreiMiquel, 1995_____. 1995. Las especies del género BostryxTroschel, 1847 en la República Argentina (2da. y última parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde124:119-127.:121; Cuezzo et al., 2013Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.:143.
Bulimulus pastoreiTablado & Mantinian, 2004Tablado, A. & Mantinian, J. 2004. Catálogo de ejemplares tipo de la División Invertebrados del Museo Argentino de Ciencias Naturales. II. Mollusca. Revista del Museo Argentino de Ciencias Naturales 6(2):363-384. :371.
Distribution. Endemic to San Luis province (32°-34°S, 65°-66°W), this species is the southernmost of the genus in Argentina (in Cerro Varela, Low Monte ecorregion). It is distributed between the Andean Cordillera (west) and the Sierras of San Luis (east), an orographic subsystem belonging to the Sierras Pampeanas, that belongs to the Arid Diagonal of South America. It covers the Dry Chaco, Espinal and Low Monte ecoregions (Fig. 2).
Type material. Lectotype, ARGENTINA, San Luis: La Capital, Cerro Varela, Pastore F. col. (MACN-In 1517). Paralectotypes, same data as holotype (MACN-In 1517-1).
Other material examined. ARGENTINA, San Luis: Coronel Pringles, Carolina, 1,600 m, 26.IX.1970, Viana M. col. (MACN-In 28800); La Capital, Cerro Varela, Pastore F. col. (IFML-Moll 15582); General Pedernera, Cerro del Morro, 01.I.1914, Pastore F. col. (MACN-In 1316, MACN-In 9917, MACN-In 9917-1); Cerro Sololasta, III.1919, Pastore F. col. (MACN-In 9834, MLP 9671); San Martín, Quebrada between Quines and Libertador San Martín, 32°17'19"S-65°41'21"W, 555 m, Cuezzo M. G. col. (IFML-Moll 15573).
Shell. Small to medium, fusiform, slender and thin (Figs 15-17). Six to 6 1/4 whorls flat to slightly convex whorls (Figs 15-17). Shell pale cream, with axial darker bands (Figs 15,16) or uniform coloration (Fig. 17). Protoconch with thin axial costules, parallel to each other. Thin spiral grooves crossing them. Spire high conic, body whorl tall in relation to total height of the shell, 70% of the total lenght (Figs 15-17). Teleoconch with axial oblique shallow ribs slightly pronunced, not elevated, separated by regular narrow spaces. Suture simple, slightly deep (Figs 15-17). Aperture elongated-ovate, narrow and tall, 50% of total shell length. Parietal space narrow, smooth (Figs 15-17). Peristome simple, sharp (Figs 15-17). Umbilicus rimate to narrow, overlapped by the peristome (Figs 15-17). Lectotype measurements. masd= 9.60, misd= 8.31, tsh= 23.0, bwh= 15.80, sh= 7.54, ah= 10.68, ad= 6.45. Additional measurements in Tabs I, II.

Bostryx pastorei (Holmberg, 1912). Shells in ventral, dorsal and lateral view: 15, lectotype (MACN 1517); 16, IFML 15573; 17, MACN 9917. (Scale bar: 5 mm).
Pallial system. Secondary ureter opening at proximal portion of pulmonary roof.
Reproductive system. Bursa copulatrix duct reaching distal portion of albumen gland (Fig. 18), inner wall with proximal longitudinal zigzag folds and distal straight folds. Vagina 1/4 of penis length (Fig. 18), inner wall with longitudinal thick straight folds. Epiphallus length three quarters that of penis (Fig. 18). Proximal and distal portions of inner wall of penis with longitudinal straight folds (Fig. 19).
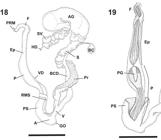
Bostryx pastorei (Holmberg, 1912). 18, View of dissected reproductive system; 19, phallic complex inner wall (scale bar: 5 mm) (A, atrium; AG, albumen gland; BC, bursa copulatrix; BCD, bursa copulatrix duct; Ep, epiphallus; F, flagellum; GO, genital opening; HD, hermaphroditic duct; P, penis; PG, penial gland; Pr, prostate; PS, penial sheath; PRM, retractor muscle; RMS, retractor muscle of penial sheath; S, spermoviduct; SV, seminal vesicle; V, vagina; VD, vas deferens).
Remarks. Bostryx pastorei was originally described asBulimulus (Mesembrinus) Albers by Holmberg (1912Holmberg, E. L. 1912. Moluscos argentinos en parte nuevos coleccionados por Franco Pastore. Physis 1:19-26.). In 1946, Parodiz classified it within the subgenus Bulimulus (Scansicochlea) Pilsbry, since it was believed that the sculpture of the protoconch fitted the description for this subgenus, characterized by a thin spiral striation, completely lacking axial costules. Afterwards, Miquel (1995_____. 1995. Las especies del género BostryxTroschel, 1847 en la República Argentina (2da. y última parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde124:119-127.) classified the species as B. pastorei in Bostryx, because he disagreed with Parodiz (1946)Parodiz, J. J. 1946. Los géneros de los Bulimulinae Argentinos. Revista Museo de La Plata (N. S.) 4(zool. 30):303-371., he observed strong axial cordons and thinner spiral cordons in the protoconch of this species. The present study coincides with the protoconch morphology for B. pastorei described by Miquel (1995)_____. 1995. Las especies del género BostryxTroschel, 1847 en la República Argentina (2da. y última parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde124:119-127.. This author considered that Peronaeus (Lissoacme) puntanus Parodiz, 1947 is a synonym of B. pastorei, given the fact that both species have similar shells. This synonymy was maintained by Cuezzo et al. (2013Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.) and on basis of the observation of type material of both species, has been reformulated in the present study.
Bostryx pastorei shows high variability in shell coloration, body whorl area, aperture shape and spire height. Specimens isolated from Cerro Varela and Quebrada between Quines and Libertador General San Martín, shared shell characters such as the presence of darker shell bands and a more slender configuration of the shell and aperture. In contrast, specimens from Cerro del Morro and Cerro Sololasta were uniformly colored and had a higher spire and a more voluminous body whorl.
Bostryx strobeli (Parodiz, 1956)
Bulimulus (Scansicochlea) strobeliParodiz, 1956_____. 1956. Cuatro nuevas especies deScansicochlea de Argentina (Mol. Pulm.). Primera parte. Neotrópica 2(8):59-64.:62; Fernández, 1973Fernández, D. & Castellanos, Z. A. 1973. Clave genérica de la malacofauna terrestre Argentina. Revista del Museo Argentino de Ciencias Naturales 11:263-285. :85; Fernández & Castellanos, 1973:279; Tablado & Mantinian, 2004Tablado, A. & Mantinian, J. 2004. Catálogo de ejemplares tipo de la División Invertebrados del Museo Argentino de Ciencias Naturales. II. Mollusca. Revista del Museo Argentino de Ciencias Naturales 6(2):363-384. :371.
Bulimulus (Scansicochlea) lolaeHylton-Scott, 1967_____. 1967. Nuevas Scansicochlea de la región central de la Argentina. Neotrópica 13:7-12. :11; Fernández & Castellanos, 1973Fernández, D. & Castellanos, Z. A. 1973. Clave genérica de la malacofauna terrestre Argentina. Revista del Museo Argentino de Ciencias Naturales 11:263-285. :278; Miquel, 1995_____. 1995. Las especies del género BostryxTroschel, 1847 en la República Argentina (2da. y última parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde124:119-127.:122; Tablado & Mantinian, 2004Tablado, A. & Mantinian, J. 2004. Catálogo de ejemplares tipo de la División Invertebrados del Museo Argentino de Ciencias Naturales. II. Mollusca. Revista del Museo Argentino de Ciencias Naturales 6(2):363-384. :371.
Bostryx lolaeBreure, 1979_____. 1979. Systematics, Phylogeny and Zoogeography of Bulimulinae (Mollusca). Zoologische Verhandelingen168:1-215.:55.
Bostryx strobeliBreure, 1979_____. 1979. Systematics, Phylogeny and Zoogeography of Bulimulinae (Mollusca). Zoologische Verhandelingen168:1-215.:58; Miquel, 1995_____. 1995. Las especies del género BostryxTroschel, 1847 en la República Argentina (2da. y última parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde124:119-127.:122; Cuezzo et al., 2013Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.:146.
Distribution. San Luis and Cordoba Provinces (32°-33°S, 64°-65°W) in the eastern Sierras Pampeanas. Dry Chaco and Espinal ecoregions (Fig. 6).
Type material. Holotype, ARGENTINA, San Luis: General Pedernera, Cerro del Morro, 1,500 m, 1913, Pastore F. col. (MACN-In 9916). Paratypes, same data as holotype (MACN-In 9916-1)
Other material examined. ARGENTINA, San Luis: General Pedernera, Cerro del Morro, 1,500 m, Parodiz J. J. col. (MLP 11018); Córdoba: Calamuchita, Santa Rosa de Calamuchita, foot of Champaquí, 1,500 m, I.1967, Cichero J. A. col. (MACN-In 27283); Villa Alpina, foot of Champaquí, riverside of Los Reartes river, 28.I.1967, Cichero J. A. col. (MLP 11009, MLP 11010).
Shell. Small to medium, elongate-ovate (Fig. 20). Five-5 1/2 slightly convex whorls. Shell yellowish brown with darker bands or uniform coloration (Figs 20, 21). Protoconch with axial shallow ribs and thin spiral grooves crossing them. Spire high conic, body whorl tall in relation to the total height of the shell, 70% of the total length (Figs 20, 21). Periostracum easily removed. Teleoconch with axial oblique shallow ribs, slightly pronunced, regularly arranged. Suture simple, slightly deep (Figs 20, 21). Aperture elongate-ovate, wide and tall, 50% the total shell length (Figs 20, 21). Parietal space narrow, smooth (Figs 20, 21). Peristome simple, not expanded (Fig. 20). Umbilicus rimate (Fig. 20), overlapped by the peristome. Holotype measurements. masd= 7.98, misd= 6.60, tsh= 16.94, bwh= 12.63, sh= 4.75, ah= 8.43, ad= 5.09. Additional measurements in Tabs I, II.

Bostryx strobeli (Parodiz, 1956). 20, Holotype (MACN 9916) in ventral, dorsal and laterals view; 21, size and shape variation of shells (MLP 11010, MACN 27283, MLP 11018) (scale bar: 1 cm).
Pallial system. Secondary ureter opening at distal portion of pulmonary roof.
Reproductive system. Bursa copulatrix duct long reaching proximal portion of albumen gland (Fig. 22), inner wall with longitudinal zigzag folds. Vagina, 1/4 of penis length (Fig. 22), inner wall with longitudinal zigzag folds. Epiphallus same length as penis (Fig. 22). Proximal and distal portions of penis inner wall with longitudinal straight folds (Fig. 23).

Bostryx strobeli (Parodiz, 1956). 22, View of reproductive system dissected out; 23, phallic complex inner wall (scale bar: 5 mm) (A, atrium; AG, albumen gland; BC, bursa copulatrix; BCD, bursa copulatrix duct; Ep, epiphallus; F, flagellum; GO, genital opening; P, penis; PG, penial gland; Pr, prostate; PS, penial sheath; PRM, retractor muscle; RMS, retractor mucle of penial sheath; S, spermoviduct; V, Vagina; VD, vas deferens).
Remarks. Bostryx strobeli was described for the first time by Parodiz (1956_____. 1956. Cuatro nuevas especies deScansicochlea de Argentina (Mol. Pulm.). Primera parte. Neotrópica 2(8):59-64.) as Bulimulus (Scansicochlea) Pilsbry, presenting a protoconch with fine spiral striation.Breure (1979_____. 1979. Systematics, Phylogeny and Zoogeography of Bulimulinae (Mollusca). Zoologische Verhandelingen168:1-215.) synonymized Scansicochlea with Bostryx thereby relocating this species as B. strobeli in Bostryx.Miquel (1995_____. 1995. Las especies del género BostryxTroschel, 1847 en la República Argentina (2da. y última parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde124:119-127.) maintained B. strobeli in Bostryx, because of the presence of axial and spiral lines in the protoconch, contrary to Parodiz (1956)_____. 1956. Cuatro nuevas especies deScansicochlea de Argentina (Mol. Pulm.). Primera parte. Neotrópica 2(8):59-64.. Based on molecular analysis performed by Breure & Romero (2012)Breure, A. S. H. & Romero, P. E. 2012. Support and surprises: molecular phylogeny of the land snail superfamily Orthalicoidea using a three-locus gene Analysis with a divergence time analysis and ancestral area reconstruction (Gastropoda: Stylommatophora). Archiv für Molluskenkunde 141(1):1-20., Breure (2012)_____. 2012. The status of the genus BostryxTroschel, 1847, with description of a new subfamily (Mollusca, Gastropoda, Bulimulidae). Zookeys 216:1-3. indicated that B. strobeli could be classified as Naesiotus Albers. However, according to the sculpture of the protoconch observed, typical of the genusBostryx, and several anatomical characters such as a penis with cup shape and internally with triangular penial gland and a portion with sculpture (consisting in folds) and other without sculpture, and an epiphallus with variable diameter and two different sculptures in the inner wall, the present study concludes that of B. strobeli must be maintained in Bostryxas proposed by Cuezzo et al. (2013Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.).
Miquel (1995_____. 1995. Las especies del género BostryxTroschel, 1847 en la República Argentina (2da. y última parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde124:119-127.) synonymized B. strobeli and Bulimulus (Scansicochlea) lolae Hylton Scott, 1967 because their shell characters were very similar. B. strobeli does differ from B. lolae by smaller dimensions of the total height and diameter of the shell, higher and more acute spire and by the fact that the teleoconch sculpture has only axial costules, whereas axial costules in B. lolae are intersected by spiral lines in the teleoconch in some specimens. However, B. lolae exhibit big intraspecific variations in their measurements and sculpture of the shells, due to that these differences are considered intraspecific variations, whereby B. strobeli and B. lolae are considered the same species.
Bostryx strobeli and B. pastorei have a similar distribution in San Luis province, but B. strobeli differs fromB. pastorei in having a shell that is more elongate-ovate with a lower spire and a yellowish brown coloration. The most noteworthy characteristic of B. strobeli is an exceptionally long bursa copulatrix duct, a character not seen in any of the other species studied.
Bostryx cordillerae (Strobel, 1874)
Bulimulus (Mesembinus) cordilleraeStrobel, 1874Strobel, P. 1874. Materiali per una malacostatica di terra e di acqua dolce dell' Argentinia meridionale. Pisa, Biblioteca Malacologica. lxxx + 142 p.:22.
Bulimus (Mesembinus) CordilleraDoering, 1875Doering, A. 1875. Apuntes sobre la fauna de Moluscos de la República Argentina, 3. Boletín de la Academia Nacional de Ciencias de la República Argentina 2:300-340.:340 [incorrect subsequent spelling].
Bulimulus (Bulimulus) cordilleraeDoering, 1879_____. 1879. Apuntes sobre la fauna de Moluscos de la República Argentina, 4. Boletín de la Academia Nacional de Ciencias de la República Argentina 3:63-84.:72.
Bulimulus (Bulimulus) monticolaDoering, 1879_____. 1879. Apuntes sobre la fauna de Moluscos de la República Argentina, 4. Boletín de la Academia Nacional de Ciencias de la República Argentina 3:63-84.:69 [nomen praeocc].
Bostryx sophieaeBreure, 1979_____. 1979. Systematics, Phylogeny and Zoogeography of Bulimulinae (Mollusca). Zoologische Verhandelingen168:1-215.:58 [new name for B. (B.) monticola]; Miquel, 1995_____. 1995. Las especies del género BostryxTroschel, 1847 en la República Argentina (2da. y última parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde124:119-127.:123.
Peronaeus (Peronaeus) cordilleraeParodiz, 1946Parodiz, J. J. 1946. Los géneros de los Bulimulinae Argentinos. Revista Museo de La Plata (N. S.) 4(zool. 30):303-371.:315; 1947:2; Fernández, 1973Fernández, D. & Castellanos, Z. A. 1973. Clave genérica de la malacofauna terrestre Argentina. Revista del Museo Argentino de Ciencias Naturales 11:263-285. :88; Fernández & Castellanos, 1973:277.
Peronaeus cordilleraeParodiz, 1957b_____. 1957b. Catalogue of the Land Mollusca of Argentina. The Nautilus 71(1):22-30.:23; 1961:60.
Peronaeus monticolaParodiz, 1957b_____. 1957b. Catalogue of the Land Mollusca of Argentina. The Nautilus 71(1):22-30.:23.
Bostryx cordilleraeBreure, 1978Breure, A. S. H. 1978. Notes on and descriptions of Bulimulidae (Mollusca, Gastropoda). Zoologische Verhandelingen 164:1-255.:66; 1979:52; Köhler, 2007Köhler, F. 2007. Annotated type catalogue of the Bulimulinae (Pulmonata: Orthalicoidea, Bulimulidae) in the Museum fur Naturkunde Berlin. Mitteilungen aus dem Museen für Naturkunde in Berlin, Zoologische Reihe 83(2):125-159.:132; Cuezzo et al.,
2013:141.
Distribution. Mendoza and San Juan provinces (31°-32°S, 68°-70°W) (Fig. 6). It occurs in the precordillera and Uspallata-Calingasta valley. Doering (1879_____. 1879. Apuntes sobre la fauna de Moluscos de la República Argentina, 4. Boletín de la Academia Nacional de Ciencias de la República Argentina 3:63-84.) cited the presence of B. cordillerae in Sierra de los Granadillos (Catamarca province) and Cuesta de Tocina in Sierra de Famatina (La Rioja province), at altitudes between 3,300 and 4,000 m. However, no malacological collection consulted contained material from the localities mentioned. Additionally, there is no other bibliographic record indicating this distribution. Therefore, the geographic distribution is considered to be restricted to San Juan and Mendoza provinces, which covers High Monte and Southern Andean Steppe ecoregions.
Type material. Lectotype, ARGENTINA, Mendoza: Casa de Piedra, Strobel col. (ZMB 24975 a). Paralectotypes, same data as lectotype (ZMB 24975 b).
Other material examined. ARGENTINA, Mendoza: Holmberg col. (MACN-In 10001-2, MACN-In 1354, MACN-In 1906); Las Heras, Cerro Pelado, Reed & Sanzin col. (MACN-In 9592); Cordillera of Uspallata, 1931, Nágera J. J. col. (MACN-In 19625); Cordillera of Uspallata, Quebrada to East of serrania of San Bartolo, 1916, Nágera J. J. col. (MACN-In 9849); Quebrada de los Césares, Cordillera of Uspallata, I.1916, Nágera J. J. col. (MACN-In 9850); near and above of Villavicencio, 1,900 m, 03.VIII.1967, Weyrauch W. col. (IFML-Moll 15580); Villavicencio, 1873, Stelzner A. col. (MACN-In 3234, MACN-In 3234-1); Villavicencio, Quebrada de las Pircas, VII.1949, Rolleri col. (MLP 8062, MACN-In 32867); Quebrada de Las Pircas, 08.VII.1949, Olivieri col. (MLP 897); Quebrada del Toro, III.1940, Birabén-Scott col. (MLP 9660, MACN-In 36944); Quebrada del Toro, Hylton Scott M. I. col. (MLP 9669); Cerro El Arco, 1942, Spinoza H. col. (MCNL 361); San Juan: Calingasta, Sierra del Tontal, 3,500 m, 22.IV.1921, Budín E. col. (MACN-In 12114).
Shell. Small, fusiform, slender and very thin (Figs 24, 25). Five to 6 whorls, flat to slightly convex (Figs 24, 25). Shell pale brown with axial oblique dark brown bands, more evident from fourth whorl. Protoconch with very thin, axial, slightly elevated costules, parallel to each other, more marked near suture and pronunced spiral grooves densely arranged crossing them (Figs 26, 27). Spire high, conic (Figs 24, 25). Body whorl tall in relation with the height of the shell, 60% of the total length (Figs 24, 25). Teleoconch with axial, straight, oblique, elevated, shallow ribs, separated by regular narrow spaces (Figs 24, 25). Suture simple, deep (Figs 24, 25). Aperture ovate, narrow and short, 40% of total shell length (Figs 24, 25). Parietal space narrow (Figs 24, 25). Peristome simple, not expanded (Figs 24, 25). Umbilicus narrow to rimate, partially overlapped by the peristome (Figs 24, 25). Lectotype measurements. masd= 10.30, misd= 2.30, tsh= 13.50, bwh= 10.0, sh= 3.3, ah= 6.0, ad= 4.40. Additional measurements in Tabs I, II.
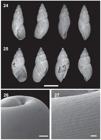
Bostryx cordillerae (Strobel, 1874). 24, 25, Size and shape variation of shells (IFML 15580, MACN 12114) (scale bar: 1 cm); 26, general view of protoconch (scale bar: 100 μm); 27, detail of protoconch's sculpture (scale bar: 20 μm).
Pallial system. Secondary ureter opening at half pulmonary roof length.
Reproductive system. Bursa copulatrix duct long, reaching distal portion of albumen gland (Fig. 28), inner longitudinal straight folds. Vagina, 1/3 of penis length (Fig. 28), inner wall with longitudinal parallel zigzag folds. Epiphallus length 2/3 of penis length (Fig. 28). Proximal penis inner wall with straight longitudinal folds and distal portion with inner tightly in zigzag folds followed by straight longitudinal folds (Fig. 29).

Bostryx cordillerae (Strobel, 1874). 28, View of reproductive system dissected out; 29, phallic complex inner wall (scale bar: 5 mm) (A, atrium; AG, albumen gland; BC, bursa copulatrix; BCD, bursa copulatrix duct; Ep, epiphallus; F, flagellum; GO, genital opening; HD, hermaphroditic duct; P, penis; PG, penial gland; Pr, prostate; PS, penial sheath; PRM, retractor muscle; RMS, retractor muscle of penial sheath; S, spermoviduct; V, vagina; VD, vas deferens).
Remarks. Based on shell characters, Bostryx cordillerae was originally described as Bulimulus (Mesembinus) Albers by Strobel (1874Strobel, P. 1874. Materiali per una malacostatica di terra e di acqua dolce dell' Argentinia meridionale. Pisa, Biblioteca Malacologica. lxxx + 142 p.). Doering (1879_____. 1879. Apuntes sobre la fauna de Moluscos de la República Argentina, 4. Boletín de la Academia Nacional de Ciencias de la República Argentina 3:63-84.) thereafter classified it in Bulimulus (Bulimulus) because of its affinity whith Bulimulus (Bulimulus) monticola Doering, 1879 (=Bostryx sophieae Breure) regarding shell and radula morphology. In his study of Argentinian Bulimulinae (=Bulimulidae), Parodiz (1946Parodiz, J. J. 1946. Los géneros de los Bulimulinae Argentinos. Revista Museo de La Plata (N. S.) 4(zool. 30):303-371.) classifiedB. cordillerae as Peronaeus(Peronaeus) Albers, because this subgenus is characterized by a smooth protoconch. According to thorough observations and in disagreement with Parodiz (1946)Parodiz, J. J. 1946. Los géneros de los Bulimulinae Argentinos. Revista Museo de La Plata (N. S.) 4(zool. 30):303-371., B. cordilleraehas sculpture in the protoconch. In 1978, BreureBreure, A. S. H. 1978. Notes on and descriptions of Bulimulidae (Mollusca, Gastropoda). Zoologische Verhandelingen 164:1-255. made the first anatomical description of B. cordillerae and classified it within the genusBostryx. Miquel (1993Miquel, S. E. 1993. Las especies del género BostryxTroschel, 1847 en la República Argentina (1ra. parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde 121:157-171., 1995) did not mention this species in his study about Argentinian Bostryxspecies. Based on the similarity in measurements and shell morphology betweenB. cordillerae and B. sophieae, the synonymy proposed by Cuezzo et al. (2013Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.) remains.
Bostryx cordillerae shows variability in spire height and body whorl area. Specimens from Villavicencio and Casa de Piedra (Mendoza province) are more slender and present a higher spire than specimens from Sierra de Tontal (San Juan province). According to Strobel (1874Strobel, P. 1874. Materiali per una malacostatica di terra e di acqua dolce dell' Argentinia meridionale. Pisa, Biblioteca Malacologica. lxxx + 142 p.),B. cordillerae is usually found in crevices of schistose rocks like B. mendozanus and B. cuyanus, but B. cordillerae clearly differs from both species. It has smaller shell dimensions, a more slender shell configuration and a higher spire than B. mendozanus. Bostryx cuyanus is the only Argentinian species of the genus with a discoidal shell, and hence the species cannot be confused.
DISCUSSION
Morphological examinations of shell and anatomical features were carried out to assess morphological differentiation among several Bostryx species distributed in the central west of Argentina. Differences among the examined species are mainly observed in shell size and shape, coloration and spire hight. In all species examined the sculpture of the protoconch corresponded to the pattern previously described for other Argentinian species of the genus. The teleoconch sculpture was similar among the species and consisted of shallow ribs. In the pallial system the position of secondary ureter opening was the only character varying between species. The differences observed in the reproductive system were few and related with length and inner wall sculpture of the organs. Comparison with other species of the genus Bostryx present in Argentina (Miranda & Cuezzo, 2014Miranda, M. J. & Cuezzo, M. G. 2014. Taxonomic revision of theBostryx stelzneri Species complex, with description of a new species (Gastropoda: Orthalicoidea: Bulimulidae). American Malacological Bulletin 32(1):74-93.; Miranda, 2015Miranda, M. J. 2015. Bostryx tororanus (Doering, 1879) species complex (Gastropoda: Stylommatophora: Bulimulidae), a review of taxonomy and distribution of endemic species from Argentina. Journal of Natural History 49:995-1022. ) allowed to establish a typical pattern in the genital system of Bostryx, penis with cup shape, inner penial gland present, inner wall of penis with folds and a portion without sculpture; epiphallus without uniform diameter and two different sculptures in the inner wall. It is remarkable that in contrast to most land snails, ArgentinianBostryx species have a homogeneous character of the reproductive system, which differs from related-genera (BulimulusLeach and Naesiotus Albers). The opposite occurs with shell characters, which vary among the species, but are similar to those of related genera. The detection of this typical pattern of the reproductive system allows maintains to B. strobeli in the genus Bostryx, unlike proposed by Breure (2012Breure, A. S. H. & Romero, P. E. 2012. Support and surprises: molecular phylogeny of the land snail superfamily Orthalicoidea using a three-locus gene Analysis with a divergence time analysis and ancestral area reconstruction (Gastropoda: Stylommatophora). Archiv für Molluskenkunde 141(1):1-20.). A cladistic analysis based on morphological characters (in preparation) will provide support to this classification.
Species of the genera Bostryx, Bulimulus,Naesiotus (Bulimulidae), EpiphragmophoraDoering (Epiphragmophoridae), Spixia Pilsbry & Vanatta andPlagiodontes Doering (Odontostomidae) can be found in San Luis province (Cuezzo, 2006Cuezzo, M. G. 2006. Systematic revision and cladistic analysis ofEpiphragmophora Doering from Argentina and Southern Bolivia (Gastropoda: Stylommatophora: Xanthonychidae). Malacologia 49(1):121-188.; Pizá & Cazzaniga, 2010Pizá, J. & Cazzaniga, N. J. 2010. Allopatry and anatomical distinctiveness of two puzzling land snails in genusPlagiodontes, from Argentina (Gastropoda: Orthalicidae, Odontostominae). Malacologia 53(1):1-24., 2012_____. 2012. Barrels and pagodas: clarifying the identity and variability of two nominal taxa of carinated land snails in the genusPlagiodontes (Gastropoda: Orthalicidae: Odontostominae). Journal of Natural History 46(7-8):383-410.; Cuezzo et al., 2013Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.). However, in San Juan and Mendoza provinces, the land snail diversity is low in comparison with other regions such as the Northwest Argentina and Orthalicoidea is almost exclusively represented by the genusBostryx (Parodiz, 1961_____. 1961. Moluscos terrestres, hasta ahora conocidos, de la provincia de Mendoza. Revista Científica Museo de Historia Natural San Rafael 1(3):59-62.;Miquel, 1993Miquel, S. E. 1993. Las especies del género BostryxTroschel, 1847 en la República Argentina (1ra. parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde 121:157-171., 1995_____. 1995. Las especies del género BostryxTroschel, 1847 en la República Argentina (2da. y última parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde124:119-127.; Cuezzo et al., 2013Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.). The low diversity recorded could be related to environmental conditions of the area, such as low precipitation, high solar radiation and evapotranspiration, little vegetation cover and immature soils with continuous cryoturbation processes, which determine the environment of the Andean desert (Roig & Martínez-Carretero, 1998Roig, F. A. & Martínez-Carretero, E. 1998. La vegetación puneña en la provincia de Mendoza, Argentina. Phytocoenologia 28(4):565-608.;Dalmasso et al., 1999Dalmasso, A.; Martínez-Carretero, E.; Videla, F.; Puig, S. & Candia, R. 1999. Reserva Natural Villavicencio (Mendoza, Argentina). Plan de manejo. Multequina 8:11-50.). Genera such as Bostryx are distributed in desert ecosystems and they are adapted to survive under harsh climatic conditions (Ramírez & Ramírez, 2013Ramírez, J. & Ramírez, R. 2013. Molecular evidence for the polyphyly of Bostryx (Gastropoda: Bulimulidae) and genetic diversity of Bostryx aguilari. Zoologia 30:80-87.). The species are mainly observed in the Precordillera and the Uspallata-Calingasta valley.
In adittion to the species examined in the present study, species likeBostryx stelzneri (San Juan province), Bostryx mendozanus (Strobel) (Mendoza province) and Bostryx cuyanus (Pfeiffer) (San Juan and Mendoza provinces) have also been recorded. Unfortunately, live specimens of B. mendozanus were not found in the distribution area and malacological collections also did not provide anatomical information about this species. The shell descriptions were provided byMiquel (1993Miquel, S. E. 1993. Las especies del género BostryxTroschel, 1847 en la República Argentina (1ra. parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde 121:157-171., 1995) and the anatomical description of B. cuyanus and B. stelzneri was performed by Hylton-Scott (1954Hylton-Scott, M. I. 1954. Una nueva especie deBostryx (Bulimulidae) (Mol.Pulm.). Physis 20(59):409-413.) and Miranda & Cuezzo (2014Miranda, M. J. & Cuezzo, M. G. 2014. Taxonomic revision of theBostryx stelzneri Species complex, with description of a new species (Gastropoda: Orthalicoidea: Bulimulidae). American Malacological Bulletin 32(1):74-93.), respectively.Bostryx mendozanus has a restricted distribution area in the Precordillera of Mendoza (between 1,500-1,800 m, Southern Andean Steppe and High Monte ecoregions of Sierra de Uspallata and Villavicencio). Bostryx cuyanus is restricted to high altitude environments in two isolated areas. One of these areas is the Precordillera (between 2,000-3,400 m, Southern Andean Steppe and High Monte) and the other one is the Principal Cordillera (3,151 m, Southern Andean Steppe). The Principal cordillera consists of great rugged mountains with steep detritus-covered slopes and deep valleys. Between 3,000-4,000 m, it has a Tundra climate while above 4,000 m the climate becomes Polar. Vegetation is typical of steppe, grassland and meadows (Méndezet al., 2006Méndez E.; Martínez-Carretero, E. & Peralta, I. 2006. La Vegetación del Parque Provincial Aconcagua (Altos Andes centrales de Mendoza, Argentina). Boletín de la Sociedad Argentina de Botánica 41(1-2):41-69.). Although since the valley that separates the precordillera of the Cordillera has a width almost 50 km and only in some places it reaches 100 km, the lack of similarity or relationships between their flora and fauna that has been demonstrated for diverses taxa (Roig & Martínez- Carretero, 1998Roig, F. A. & Martínez-Carretero, E. 1998. La vegetación puneña en la provincia de Mendoza, Argentina. Phytocoenologia 28(4):565-608.; Roig-Juñent et al., 2003Roig-Juñent, S.; Flores, G. E. & Mattoni, C. 2003. Consideraciones biogeográficas de la Precordillera (Argentina), con base en artrópodos epígeos. : Morrone, J. & Llorente-Bousquets, J. eds. Una perspectiva latinoamericana de la Biogeografía. México, D.F., Las Prensas de Ciencias, Facultad de Ciencias, UNAM. 307p.; Ochoa, 2004Ochoa, J. A. 2004. Filogenia del géneroOrobothriurus y descripción de un nuevo género de Bothriuridae (Scorpiones). Revista Ibérica de Aracnología 9:43-73.). However, Bostryx cuyanus is the only species of the genus present in both mountain ranges.
The criteria most frequently cited for establishing conservation priorities are species richness, endemism and presence of rare species (Benkendorff & Davis, 2002Benkendorff, K. & Davis, A. R. 2002. Identifying hotspots of molluscan species richness on rocky intertidal reefs. Biodiversity and Conservation 11:1959-1973.). The central west area of the country meets all three requirements: it includes 42% of the total richness of the genus in the country, with five endemic species and a restricted distribution area. Moreover, gastropods pay a high price for their occupancy of dry and cold terrestrial environments, which is reflected, for instance, in slow growth rates and high stochastic mortality (Dallinger et al., 2001Dallinger, R.; Berger, B.; Triebskorn- Köhler, R. & Köhler, H. 2001. Soil biology and ecotoxicology. : BARKER, G.M. ed. The Biology of Terrestrial Molluscs. Wallingford, Cabi Publishing. 558p.). For these reasons, this Precordillera, should be considered an area with high priority of conservation.
Acknowledgements
I thank to curators of the Malacological collections consulted: A. Tablado (MACN-In), G. Darrigran and M. Tassara (MLP) and B. Bender (MCNL) for their assistance and loan of material. I also thank A. Tablado for the photographs of B. reedi, B. strobeli and B. puntanus type material. Thanks to M. G. Cuezzo for the collected material and the reading of the manuscript. I sincerely appreciate comments provided by the anonymous reviewers, their suggestions greatly improved the quality of this paper. The author is a fellow of the Argentine National Council of Scientific Research (CONICET).
REFERENCES
- Abraham de Vazquez, E. M.; Garleff, K.; Liebricht, H.; Regairaz, A. C.; Schäbitz, F.; Squeo, F. A.; Stingl, H.; Veit, H. & Villagrán, C. 2000. Geomorphology and paleoecology of the arid diagonal in Southern South AmericaZeitschrift für Angewandte Geologie: SonderheftSH 1:55-61.
- Benkendorff, K. & Davis, A. R. 2002. Identifying hotspots of molluscan species richness on rocky intertidal reefs. Biodiversity and Conservation 11:1959-1973.
- Breure, A. S. H. 1978. Notes on and descriptions of Bulimulidae (Mollusca, Gastropoda). Zoologische Verhandelingen 164:1-255.
- _____. 1979. Systematics, Phylogeny and Zoogeography of Bulimulinae (Mollusca). Zoologische Verhandelingen168:1-215.
- _____. 2012. The status of the genus BostryxTroschel, 1847, with description of a new subfamily (Mollusca, Gastropoda, Bulimulidae). Zookeys 216:1-3.
- Breure, A. S. H. & Romero, P. E. 2012. Support and surprises: molecular phylogeny of the land snail superfamily Orthalicoidea using a three-locus gene Analysis with a divergence time analysis and ancestral area reconstruction (Gastropoda: Stylommatophora). Archiv für Molluskenkunde 141(1):1-20.
- Cuezzo, M. G. 2006. Systematic revision and cladistic analysis ofEpiphragmophora Doering from Argentina and Southern Bolivia (Gastropoda: Stylommatophora: Xanthonychidae). Malacologia 49(1):121-188.
- Cuezzo, M. G.; Miranda, M. J. & Ovando, X. M. C. 2013. Species catalogue of Orthalicoidea in Argentina (Gastropoda: Stylommatophora). Malacologia 56(1-2):135-191.
- Dallinger, R.; Berger, B.; Triebskorn- Köhler, R. & Köhler, H. 2001. Soil biology and ecotoxicology. : BARKER, G.M. ed. The Biology of Terrestrial Molluscs. Wallingford, Cabi Publishing. 558p.
- Dalmasso, A.; Martínez-Carretero, E.; Videla, F.; Puig, S. & Candia, R. 1999. Reserva Natural Villavicencio (Mendoza, Argentina). Plan de manejo. Multequina 8:11-50.
- Doering, A. 1875. Apuntes sobre la fauna de Moluscos de la República Argentina, 3. Boletín de la Academia Nacional de Ciencias de la República Argentina 2:300-340.
- _____. 1879. Apuntes sobre la fauna de Moluscos de la República Argentina, 4. Boletín de la Academia Nacional de Ciencias de la República Argentina 3:63-84.
- Fernández, D. 1973. Catálogo de la malacofauna terrestre Argentina. Comisión de Investigaciones Científicas de la Provincia de Buenos Aires 4:1-197.
- Fernández, D. & Castellanos, Z. A. 1973. Clave genérica de la malacofauna terrestre Argentina. Revista del Museo Argentino de Ciencias Naturales 11:263-285.
- Furque, G. & Cuerda, A. J. 1979. Precordillera de La Rioja, San Juan y Mendoza. Segundo Simposio Geología Regional Argentina 1:455-522.
- Hijmans, R.; Guarino, L.; Jarvis, A & O'Brien, R. 2007. DIVA-GIS Version 7.3.0.1. Available at <Available athttp://www.diva-gis.org/ >. Accessed on January 2015.
» http://www.diva-gis.org/ - Holmberg, E. L. 1912. Moluscos argentinos en parte nuevos coleccionados por Franco Pastore. Physis 1:19-26.
- Hylton-Scott, M. I. 1954. Una nueva especie deBostryx (Bulimulidae) (Mol.Pulm.). Physis 20(59):409-413.
- _____. 1967. Nuevas Scansicochlea de la región central de la Argentina. Neotrópica 13:7-12.
- Insel, N.; Poulsen, C. J. & Ehlers, T. A. 2009. Influence of the Andes Mountains on South American moisture transport, convection, and precipitation. Climate Dynamics 35:1477-1492.
- Köhler, F. 2007. Annotated type catalogue of the Bulimulinae (Pulmonata: Orthalicoidea, Bulimulidae) in the Museum fur Naturkunde Berlin. Mitteilungen aus dem Museen für Naturkunde in Berlin, Zoologische Reihe 83(2):125-159.
- Méndez E.; Martínez-Carretero, E. & Peralta, I. 2006. La Vegetación del Parque Provincial Aconcagua (Altos Andes centrales de Mendoza, Argentina). Boletín de la Sociedad Argentina de Botánica 41(1-2):41-69.
- Miquel, S. E. 1993. Las especies del género BostryxTroschel, 1847 en la República Argentina (1ra. parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde 121:157-171.
- _____. 1995. Las especies del género BostryxTroschel, 1847 en la República Argentina (2da. y última parte) (Gastropoda Stylommatophora: Bulimulidae). Archiv für Molluskenkunde124:119-127.
- Miranda, M. J. 2015. Bostryx tororanus (Doering, 1879) species complex (Gastropoda: Stylommatophora: Bulimulidae), a review of taxonomy and distribution of endemic species from Argentina. Journal of Natural History 49:995-1022.
- Miranda, M. J. & Cuezzo, M. G. 2014. Taxonomic revision of theBostryx stelzneri Species complex, with description of a new species (Gastropoda: Orthalicoidea: Bulimulidae). American Malacological Bulletin 32(1):74-93.
- Ochoa, J. A. 2004. Filogenia del géneroOrobothriurus y descripción de un nuevo género de Bothriuridae (Scorpiones). Revista Ibérica de Aracnología 9:43-73.
- Olson, D. M. & Dinerstein, E. 1998. The Global 200: A Representation Approachto Conserving the Earth's Most Biologically Valuable Ecoregions. Conservation Biology 12:502-515.
- Olson, D. M.; Dinerstein, E.; Wikramanayake, E. D.; Burgess, N. D.; Powell, G. V. N.; Underwood, E. C.; D'Amico, J. A.; Itoua, I.; Strand, H. E.; Morrison, J. C. et al. 2001. Terrestrial Ecoregions of the world: A new map of Life on Earth. BioScience 51(11):933-938.
- Parodiz, J. J. 1946. Los géneros de los Bulimulinae Argentinos. Revista Museo de La Plata (N. S.) 4(zool. 30):303-371.
- _____. 1947. Contribuciones al conocimiento de los moluscos terrestres Sudamericanos, V. Comunicaciones Zoológicas del Museo de Historia Natural de Montevideo2:1-32.
- _____. 1956. Cuatro nuevas especies deScansicochlea de Argentina (Mol. Pulm.). Primera parte. Neotrópica 2(8):59-64.
- _____. 1957a. Catalogue of the Land Mollusca of Argentina. The Nautilus 70(1):127-135.
- _____. 1957b. Catalogue of the Land Mollusca of Argentina. The Nautilus 71(1):22-30.
- _____. 1961. Moluscos terrestres, hasta ahora conocidos, de la provincia de Mendoza. Revista Científica Museo de Historia Natural San Rafael 1(3):59-62.
- Pizá, J. & Cazzaniga, N. J. 2010. Allopatry and anatomical distinctiveness of two puzzling land snails in genusPlagiodontes, from Argentina (Gastropoda: Orthalicidae, Odontostominae). Malacologia 53(1):1-24.
- _____. 2012. Barrels and pagodas: clarifying the identity and variability of two nominal taxa of carinated land snails in the genusPlagiodontes (Gastropoda: Orthalicidae: Odontostominae). Journal of Natural History 46(7-8):383-410.
- Quintana, M. 1982. Catálogo preliminar de la malacofauna del Paraguay. Revista del Museo Argentino de Ciencias Naturales , Zoología 11(3):61-158.
- Ramírez, R.; Paredes, C. & Arenas, j. 2003. Moluscos del Perú. Revista de Biología Tropical 51(3):225-284.
- Ramírez, J. & Ramírez, R. 2013. Molecular evidence for the polyphyly of Bostryx (Gastropoda: Bulimulidae) and genetic diversity of Bostryx aguilari Zoologia 30:80-87.
- Rohlf, F. J. 2013a. ´TpsUtil. version 1.58`. Available at <Available at http://life.bio.sunysb.edu/morph/ >. Accessed on January 2015.
» http://life.bio.sunysb.edu/morph/ - _____. 2013b. ´TpsDig2. version 2.17`. Available at <http://life.bio.sunysb.edu/morph/>. Accessed on January 2015.
» http://life.bio.sunysb.edu/morph/ - Roig, F. A. & Martínez-Carretero, E. 1998. La vegetación puneña en la provincia de Mendoza, Argentina. Phytocoenologia 28(4):565-608.
- Roig-Juñent, S.; Flores, G.; Claver, S.; Debandi, G. & Marvaldi, A. 2001. Monte Desert (Argentina): Insect biodiversity and natural areas. Journal of Arid Environments 47(1):77-94.
- Roig-Juñent, S.; Flores, G. E. & Mattoni, C. 2003. Consideraciones biogeográficas de la Precordillera (Argentina), con base en artrópodos epígeos. : Morrone, J. & Llorente-Bousquets, J. eds. Una perspectiva latinoamericana de la Biogeografía. México, D.F., Las Prensas de Ciencias, Facultad de Ciencias, UNAM. 307p.
- Siame, L. L.; Bellier, O. & Sebrier, M. 2006. Active tectonics in the Argentine precordillera and western Sierras Pampeanas. Revista de la Asociación Geológica Argentina 61(4):604-619.
- Strobel, P. 1874. Materiali per una malacostatica di terra e di acqua dolce dell' Argentinia meridionale. Pisa, Biblioteca Malacologica. lxxx + 142 p.
- Tablado, A. & Mantinian, J. 2004. Catálogo de ejemplares tipo de la División Invertebrados del Museo Argentino de Ciencias Naturales. II. Mollusca. Revista del Museo Argentino de Ciencias Naturales 6(2):363-384.
- Villagran, C. & Hinojosa, L. F. 1997. Historia de los bosques del sur de Sudamérica, II: Análisis fitogeográfico. Revista Chilena de Historia Natural 70:241-267.
Publication Dates
-
Publication in this collection
Oct-Dec 2015
History
-
Received
20 July 2015 -
Accepted
18 Dec 2015













