Abstract
On the southwest Pacific Coast of Colombia, a field study was initiated to determine the human-vector association between Anopheles (Kerteszia) neivai and fishermen, including their nearby houses. Mosquitoes were collected over 24-h periods from mangrove swamps, marshlands and fishing vessels in three locations, as well as in and around the houses of fishermen. A total of 6,382 mosquitoes were collected. An. neivai was most abundant in mangroves and fishing canoes (90.8%), while Anopheles albimanus was found indoors (82%) and outdoors (73%). One An. neivai and one An. albimanus collected during fishing activities in canoes were positive for Plasmodium vivax , whereas one female An. neivai collected in a mangrove was positive for P. vivax . In the mangroves and fishing canoes, An. neivai demonstrated biting activity throughout the day, peaking between 06:00 pm-07:00 pm and there were two minor peaks at dusk and dawn. These peaks coincided with fishing activities in the marshlands and mangroves, a situation that places the fishermen at risk of contracting malaria when they are performing their daily activities. It is recommended that protective measures be implemented to reduce the risk that fishermen will contract malaria.
malaria; Kerteszia; Anopheles neivai; biting activity; Colombia
Anopheles (Kerteszia) neivai has a wide distribution in the neotropics, ranging from southern Mexico to Ecuador, with some records of its appearance in Bolivia, Peru and northern Brazil ( Zavortink 1973Zavortink TJ 1973. Mosquitoes studies (Diptera: Culicidae). XXIX. A review of the subgenus Kerteszia of Anopheles . Contr Amer Ent Inst 9 : 1-59. , Marrelli et al. 2007Marrelli MT, Malafronte RS, Sallum MAM, Natal D 2007. Kerteszia subgenus of Anopheles associated with the Brazilian Atlantic rainforest: current knowledge and future challenges. Malar J 2007, 6 : 127. ). This species is mainly found on the Pacific Coast of Colombia in association with epiphytic and terrestrial bromeliads ( Montoya-Lerma et al. 2011Montoya-Lerma J, Solarte YA, Giraldo-Calderón GI, Quiñones ML, Ruiz-López F, Wilkerson RC, González R 2011. Malaria vector species in Colombia - A review. Mem Inst Oswaldo Cruz 106 (Suppl. I): 223-238. ). In this region, An. neivai is considered a vector of human malaria ( Carvajal et al. 1989Carvajal H, de Herrera MA, Quintero J, Alzate A, Herrera S 1989. Anopheles neivai : a vector of malaria in the Pacific low lands of Colombia. Trans R Soc Trop Med Hyg 83 : 609. , Olano et al. 2001Olano V, Brochero H, Sáenz R, Quiñones M, Molina J 2001. Mapas preliminares de la distribución de Anopheles vectores de malaria en Colombia. Biomédica 21 : 402-403. , Gutiérrez et al. 2008Gutiérrez LA, Naranjo N, Jaramillo LM, Muskus C, Luckhart S, Conn JE, Correa M 2008. Natural infectivity of Anopheles species from the Pacific and Atlantic Regions of Colombia. Acta Tropica 107 : 99-105 ). Of all Kerteszia species reported in Colombia, An. neivai has been the most studied, although much remains to be learned about its distribution and epidemiological importance. Previous studies have examined biting activity, oviposition sites, vectorial incrimination and epidemiological and sociological data, which have shown exophilic and exophagic preferences for biting throughout the day, sylvatic biting activity, high anthropophily, multiparity and natural infection with Plasmodium ( Astaiza et al. 1988Astaiza R, Murillo C, Fajardo P 1988. Biología de Anopheles (Kerteszia) neivai h., d. & k., 1913 (Diptera: Culicidae) en la Costa Pacífica de Colombia. Fluctuación de la población adulta. Rev Saude Publica 22 : 101-108. , Murillo et al. 1988Murillo C, Astaiza V, Fajardo P 1988. Biología de Anopheles (Kerteszia) neivai H., D. & K., 1913 (Diptera: Culicidae) en la Costa Pacífica de Colombia. I. Fluctuación de la población larval y caracteristicas de sus criaderos. Rev Saude Publica: 22 : 94-100. , Carvajal et al. 1989Carvajal H, de Herrera MA, Quintero J, Alzate A, Herrera S 1989. Anopheles neivai : a vector of malaria in the Pacific low lands of Colombia. Trans R Soc Trop Med Hyg 83 : 609. , Solarte et al. 1996Solarte Y, Hurtado C, Gonzalez R, Alexander B 1996. Man-biting Activity of Anopheles (Nyssorhynchus) albimanus and An . (Kerteszia) neivai (Diptera: Culicidae) in the Pacific lowlands of Colombia. Mem Inst Oswaldo Cruz 91 : 141-146. , Montoya-Lerma et al. 2011Montoya-Lerma J, Solarte YA, Giraldo-Calderón GI, Quiñones ML, Ruiz-López F, Wilkerson RC, González R 2011. Malaria vector species in Colombia - A review. Mem Inst Oswaldo Cruz 106 (Suppl. I): 223-238. ).
The Colombian Pacific region has optimal conditions for the survival of this species. This is likely a result of the abundance of bromeliad epiphytes, which are its oviposition sites and the fact that many settlements are located near forested areas, which facilitates mosquito-human contact. Given these biological and behavioural characteristics, it is believed that this species could be playing an important role in the transmission of extradomiciliary malaria along the Pacific Coast ( Solarte et al. 1996Solarte Y, Hurtado C, Gonzalez R, Alexander B 1996. Man-biting Activity of Anopheles (Nyssorhynchus) albimanus and An . (Kerteszia) neivai (Diptera: Culicidae) in the Pacific lowlands of Colombia. Mem Inst Oswaldo Cruz 91 : 141-146. , Montoya-Lerma et al. 2011Montoya-Lerma J, Solarte YA, Giraldo-Calderón GI, Quiñones ML, Ruiz-López F, Wilkerson RC, González R 2011. Malaria vector species in Colombia - A review. Mem Inst Oswaldo Cruz 106 (Suppl. I): 223-238. ).
For over two decades, the term “occupational malaria” has been used to indicate occupation as a risk factor for contracting malaria. This relationship has been documented in several studies. The occupation types associated with malaria are diverse; among others, health workers ( Rajasekhar & Nandakumar 2000Rajasekhar M, Nandakumar NV 2000. Occupational malaria and health risk among select occupational health care employee groups in an urban hospital at Tirupati, AP. Indian J Malariol 37 : 53-60. ), workers in the alluvial gold mining areas ( Barbieri & Sawyer 2007Barbieri AF, Sawyer DO 2007. Heterogeneidade da prevalência de malária em garimpos do norte de Mato Grosso, Brasil. Cad Saude Publica 23 : 2878-2886. ) and farmers ( Clark & Kelly 1993Clark J, Kelly K 1993. Human genetics, paleoenvironments and malaria: relationships and implications for the settlement of Oceania. Am Anthropol 95 : 612-630. , Dolo et al. 2004Dolo G, Briet OJ, Dao A, Traore SF, Boaure M, Sogoba N 2004. Malaria transmission in relation to rice cultivation in the irrigated Sahel of Mali. Acta Trop 89 : 147-159. , Sanabria et al. 2004Sanabria H, Hernández A, Villafuerte A, Erazo P 2004. Percepción y condiciones de trabajo de personas que tuvieron malaria. Rev Peru Med Exp Salud Publica 21 : 210-216. , da Silva et al. 2010da Silva NS, da Silva-Nunes M, Malafronte RS, Menezes MJ, D’Arcadia RR, Komatsu NT, Scopel KK, Braga EM, Cavasini CE, Cordeiro JA, Ferreira MU 2010. Epidemiology and control of frontier malaria in Brazil: lessons from community-based studies in rural Amazonia. Trans R Soc Trop Med Hyg 104 : 343-350. ) are at increased risk for contracting the disease. Similarly, illegal labour activities, such as agriculture and mining without control, may favour the proliferation of oviposition sites, allowing for the maintenance of high densities of mosquito vectors ( Padilla et al. 2011Padilla JC, Alvarez G, Montoya R, Chaparro P, Herrera S 2011. Epidemiology and control of malaria in Colombia. Mem Inst Oswaldo Cruz 106 (Suppl. I): 114-122. ). In addition, the anopheline habitat can be increased by certain activities, such as extensive breeding of cattle, pigs, goats and some birds ( Mboera et al. 2010Mboera LEG, Senkoro KP, Mayala BK, Rumisha SF, Rwegoshora RT, Mlozi MR, Shayo EH 2010. Spatio-temporal variation in malaria transmission intensity in five agro-ecosystems in Mvomero district, Tanzania. Geospat Health 4 : 167-178. ).
Even non-endemic areas may present occupational malaria, as reported by Jaremin et al. (1993)Jaremin B, Nahorski W, Goljan J, Felczak-Korzybska I, Górski J, Myjak P, Kotłowski A 1993. Malaria as an occupational disease. Bull Inst Marit Trop Med Gdynia 44-45 : 43-50. in Poland, a country where malaria cases were documented only in marine fishermen and missionaries who had worked or were working in endemic areas.
In Colombia, there have been few studies examining the relationship between fishing and malaria risk. Sevilla-Casas (1993)Sevilla-Casas E 1993. Human mobility and malaria risk in the Naya River basin of Colombia. Social Sci Med 37 : 1155-1167. established that, in the Naya River Basin in the Colombian Pacific, an area where malaria rates are high, economically related human migration coincides with the highest densities of mosquitoes, ensuring intense and continuous transmission of malaria. In the delta region, the beach is a high-risk zone, especially for adults, possibly as a result of high mosquito biting rates in the beach area. Other studies have shown that fishermen may be at risk of being bitten by exophagic mosquitoes such as An. neivai , principally by diurnal exposure or by entering sylvatic environments ( Valderrama & Santander 1985Valderrama FG, Santander AG 1985. Riesgo de infección extradomiciliar de la malaria en diferentes ámbitos sociológicos ocupacionales en Bajo Calima, Buenaventura, Colombia . MSc Thesis, University of Valle, Cali, 191 pp. , Murillo et al. 1988Murillo C, Astaiza V, Fajardo P 1988. Biología de Anopheles (Kerteszia) neivai H., D. & K., 1913 (Diptera: Culicidae) en la Costa Pacífica de Colombia. I. Fluctuación de la población larval y caracteristicas de sus criaderos. Rev Saude Publica: 22 : 94-100. , Carvajal et al. 1989Carvajal H, de Herrera MA, Quintero J, Alzate A, Herrera S 1989. Anopheles neivai : a vector of malaria in the Pacific low lands of Colombia. Trans R Soc Trop Med Hyg 83 : 609. , Solarte et al. 1996Solarte Y, Hurtado C, Gonzalez R, Alexander B 1996. Man-biting Activity of Anopheles (Nyssorhynchus) albimanus and An . (Kerteszia) neivai (Diptera: Culicidae) in the Pacific lowlands of Colombia. Mem Inst Oswaldo Cruz 91 : 141-146. ). The aim of this study was to establish the biting behaviour of An. neivai at locations indoors, outdoors, outside homes and in fishing canoes used by fishermen over a 24-h period and to determine if there is a spatial and temporal correlation between the peaks of An. neivai biting activity and fishing activities of inhabitants in the area.
The current study was conducted on the Pacific Coast of the Nariño department, an area endemic for malaria and where artisanal fishing is an important economic activity. The Pacific region has the highest incidence of malaria by Plasmodium falciparum infection. Furthermore, high chemoresistance of P. falciparum to antimalarial drugs has been reported for this department ( Aponte et al. 2011Aponte SL, Díaz G, Pava Z, Echeverry DF, Ibarguen D, Rios M, Murcia LM, Quelal C, Murillo C, Gil P, Björkman A, Osorio L 2011. Sentinel network for monitoring in vitro susceptibility of Plasmodium falciparum to antimalarial drugs in Colombia: a proof of concept. Mem Inst Oswaldo 106 (Suppl. I): 123-129. ). In 2010, 32 cases of malaria (87% P. falciparum and 13% Plasmodium vivax ) were reported, while in 2011, 99 cases (78% P . falciparum and 22% P. vivax ) occurred. Of these cases, 55% occurred in males (72 cases) and 45% occurred in females (59 cases). Among females, the most affected group was school age children (49.2% of cases), followed by housewives (45.8%). Among males, fishing is associated with infection by Plasmodium ; 55.6% of reported cases have been documented in active fishermen. The second group of males at risk comprises students (40.3%) (Public Health Surveillance System Colombia, Municipal Health Secretary of the Municipality of Santa Bárbara, unpublished observations).
MATERIALS AND METHODS
Site study - The field work was carried out at three localities of Santa Bárbara-Iscuandé municipality (2º24’54.93”N 77º57’38.35”O). This municipality is located in the northern part of the department of Nariño (Colombia) and covers 1,232 km 2 .
The main site used to determine the biting activity of An. neivai was the locality of Las Varas. This settlement is surrounded by mangroves and consists of approximately 80 houses and 250 inhabitants. Of the men, 92% are fishermen and 98% of women work at home, cooking and cleaning the house. The children attend school in the mornings, but they go to the mangroves to catch crabs, shells and other molluscs in their free time.
Observations of fishing activities were conducted in three localities: Las Varas, Juanchillo and La Ensenada ( Fig. 1 ). Natural infection was determined by the total mosquitoes caught in all three localities, including mangroves and canoes.
: study area. Municipality of Santa Bárbara-Iscuandé, localities of Las Varas, Juanchillo and La Ensenada.
Mosquito collection and sampling strategy - Human landing catches were constructed in mangrove swamps, marshlands, fishing canoes, outdoors and inside houses. The outdoor collections were performed at a distance of up to 50 m from dwellings. Approximately 500 m from the locality of Las Varas, three wooden platforms were constructed among the mangroves with the aim of collecting female mosquitoes without interruption from the rising tide. The platforms were placed 150 m from each other. Canoes were placed at sites within 500 m of Las Varas, at distances ranging from 0-200 m from the coast, depending on the ease with which a stable canoe position could be maintained.
The collections were made over a period of 14 days between August 2009-November 2011 and they were divided into two sessions of seven days each to obtain 144 h (6 days) of continuous sampling at every site. All the sites were sampled simultaneously with one collector per site, except in canoes, where there were two people per site (a fisherman and a collector). Collections were made using mouth aspirators and adult females were captured as they tried to land on the collectors, who were entomological technicians.
The general procedure for 144 h of sampling at each site (6 full days) was as follows: samplings were made simultaneously at every location (indoors, outdoors, in mangroves and in canoes) during consecutive 24-h periods. In total, 10 people participated in the mosquito collections. Sampling sessions were conducted for six continuous hours, followed by 6 h of rest. Five collectors began the sampling for 6 h and then were replaced by five others; thus, the sample-rest stage was repeated to achieve 24 h of sampling at each site. This procedure was repeated until six complete 24-h samplings were achieved at every site. Every time a 24-h collection was completed at all the sampling sites, there was a 12-h break to relocate the collectors and begin the sampling at a new time and place. The collectors were rotated between sites with the aim of avoiding bias from personal variations in attracting mosquitoes. The captured females were individually stored in vials and brought to the laboratory for preservation and taxonomic determination.
Mosquito identification - The mosquitoes were identified using keys for Anopheles ( González & Carrejo 2009González R, Carrejo N 2009. Introducción al estudio taxonómico de Anopheles de Colombia. Claves y notas de distribución , 1st ed., Universidad del Valle, Cali, 237 pp. ) and the Kerteszia subgenus species ( Zavortink 1973Zavortink TJ 1973. Mosquitoes studies (Diptera: Culicidae). XXIX. A review of the subgenus Kerteszia of Anopheles . Contr Amer Ent Inst 9 : 1-59. ).
Molecular confirmation of species - Genomic DNA was extracted from adult mosquito abdomens with the DNeasy Blood & Tissue Kit (QIAgen ® , Germantown, MD, USA). The rDNA internal transcribed spacer 2 (ITS2) region was amplified using the primers of Collins and Paskewitz (1996) Collins FH, Paskewitz SM 1996. A review of the use of ribosomal DNA (rDNA) to differentiate among cryptic Anopheles species. Insect Mol Biol 5 : 1-9. and polymerase chain reaction (PCR) conditions described by Linton et al . (2001)Linton YM, Harbach RE, Anthony TG, Chang MS, Asmad M 2001. Morphological and molecular identity of Anopheles (Cellia) sundaicus (Diptera: Culicidae), the nominotypical member of a malaria vector species complex in Southeast Asia. Syst Entomol 26 : 357-366. . A portion (710 bp, including primers) of the barcoding region of cytochrome c oxidase I ( COI ) (mDNA) was amplified using the primers designed by Folmer et al. (1994)Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol 3 : 294-299. and the PCR conditions described by Ruiz et al. (2010)Ruiz F, Linton Y-M, Ponsonby DJ, Conn JE, Herrera M, Quiñones ML, Vélez ID, Wilkerson RC 2010. Molecular comparison of topotypic specimens confirms Anopheles (Nyssorhynchus) dunhami Causey (Diptera: Culicidae) in the Colombian Amazon. Mem Inst Oswaldo Cruz 105: 899-903. . The products were visualised on a 1% agarose gel containing 0.5 mg/mL of ethidium bromide. The PCR product was purified using ExoSAP-IT ® (USB Corporation, Cleveland, OH, USA).
Sequencing reactions were carried out in both directions using the Big Dye Terminator Kit ® (PE Applied BioSystems, Warrington, England) on an ABI 3730 automated sequencer (PE Applied BioSystems). The sequences were edited with Sequencher TM 4.10.1 (Gene Codes Corporation, Ann Arbor, MI, USA) and aligned manually in MacClade v.4.06 ( Maddison & Maddison 2003Maddison DR, Maddison WR 2003. MacClade v.4.06: analysis of phylogeny and character evolution, Sinauer Associates, Sunderland, MA, USA. ). Sequence similarities were compared with those available in GenBank using Basic Local Alignment Search Tool (ncbi.nlm.nih.gov/genbank/) and compared with sequences available in Barcode of Life Data Systems (barcodinglife.com/). Sequence statistics were calculated using MEGA v.4 ( Kumar et al. 2008Kumar S, Dudley J, Nei M, Tamura K 2008. MEGA: a biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief Bioinform 9 : 299-306. ).
Description of fishing activities - To detect and describe the main fishing activities in the area, 180 structured interviews of active fishermen were performed; the time of departure and return from fishing, the preferred sites and times for fishing, the species of fish captured and the types of fishing methods were recorded. The fishermen also reported the control measures used to prevent mosquito bites and whether the measures seemed effective.
A total of 50 h of observations were made of fishermen in the field while they performed their fishing activities. The observation sites consisted of mangroves frequented by fishermen to set their nets, canoes with fishermen and boarding areas. Three trained persons recorded the times of departure and arrival of the fishermen, the fish caught and the fishing technique used. The places where the main fishing activities took place were georeferenced using a GPS system in the localities of La Ensenada, Juanchillo and Las Varas.
Natural infection - Parasite detection of Plasmodium species in female mosquitoes was performed using an ELISA ( Wirtz et al. 1985Wirtz RA, Burkot TR, Andre RG, Rosenberg R, Collins WE, Roberts DR 1985. Identification of Plasmodium vivax sporozoites in mosquitoes using an enzyme-linked immunosorbent assay. Am J Trop Med Hyg 34 : 1048-1054. , 1987Wirtz RA, Zavala F, Charoenvit Y, Campbell GH, Burkot TR, Schneider I, Esser KM, Beaudoin RL, Andre RG 1987. Comparative testing of monoclonal antibodies against Plasmodium falciparum sporozoites for ELISA development. Bull WHO 65 : 39-45. ). The procedure was adjusted following the guidelines of the standard protocol distributed with the ELISA kits (Centers for Disease Control and Prevention, Atlanta, GA, USA). Once identified, the head and thorax of each specimen were separated from the body and macerated in a vial of 0.5 mL. The remainder of each body (wings, legs and abdomens) was stored for taxonomic support. Each mosquito was tested individually or in pools of up to 10 specimens, provided that they belonged to the same species, came from the same site and were collected on the same date. Samples were tested in a 96-well ELISA plate along with seven negative controls (mosquitoes from a laboratory colony of Anopheles albimanus ) and two positive controls corresponding to pure circumsporozoite protein. The results were read in an ELISA reader (MRX DYNEX ® , Magellan Biosciences) with a 415 nm filter and were rechecked after 1 h. The cut-off used was two times the average of the negative control, following the directions of Beier et al. (1988)Beier JC, Asiago CM, Onyango FK, Koros JK 1988. ELISA absorbance cut-off method affects malaria sporozoite rate determination in wild Afrotropical Anopheles . Med Vet Entomol 2 : 259-264. . Positives samples in the first ELISA underwent a second test. Only the samples that were positive in both tests were ultimately considered positive.
RESULTS
Species identification - A total 6,382 anopheline mosquitoes were collected. Two species were found: An. neivai represented 81.2% (5,222 female mosquitoes) and An. albimanus represented 18.2% (1,160 female mosquitoes) ( Table I ).
Molecular confirmation - A total of 120 DNA sequences were obtained, corresponding to 30 sequences per collection site. Analysis of these sequences using the markers COI and ITS2 showed only one haplotype, confirming no intraspecific variation among the individuals analysed. The ITS2 fragment showed no length variability (546 bp) and the COI fragment exhibited a length of 650 pb. The COI sequences were compared with sequences of Barcode of Life Data Systems (boldsystems.org). This comparison showed that the sequences obtained had a high similarity to sequences of An. neivai : barcode record MBIB513-10 (99.69% similarity) and barcode record MBIB519-10 (99.54% similarity).
Relative abundance - An. albimanus was the most abundant species indoors and outdoors (73% and 76%, respectively), while An. neivai was the most abundant species in extradomiciliary sites such as mangroves (91%) and canoes (90%).
Human biting rates (HBR) - Indoors and outdoors - Indoors, An. neivai showed a peak between 05:00 pm-07:00 pm. This peak had a HBR of 15.6 mosquitoes/man/h (m/m/h), with a range of variation (RV) between 30 m/m/h-180 m/m/h. An. albimanus presented one peak between 05:00 pm-07:00 pm, more than twice that of An. neivai (HBR: 38.4 m/m/h), with decreasing activity throughout the night ( Fig. 2 ).
: man biting activity in indoors of Anopheles neivai and Anopheles albimanus (geometric mean: 95% confidence interval), municipality of Iscuandé, Las Varas (sampling for 24 h; n = 841).
Outdoors, both species demonstrated activity beginning at 05:00 pm. The peak of activity of An. albimanus (HBR: 35.4 m/m/h; RV: 21 m/m/h - 48 m/m/h) was at 07:00 pm and was more than twice that of An. neivai (HBR: 17.6 m/m/h; RV: 11 m/m/h-24 m/m/h), which occurred at 06:00 pm ( Fig. 3 ).
: man biting activity in peridomicile of Anopheles neivai and Anopheles albimanus (geometric mean: 95% confidence interval), municipality of Iscuandé, Las Varas (sampling for 24 h; n = 906).
Outside the home and in mangroves - In mangroves (wooden platforms), An. neivai showed the highest biting rates, with a peak at 06:00 pm (HBR: 64,9 m/m/h; RV: 30 m/m/h-180 m/m/h) and two small peaks at 09:00 pm and 11:00 pm. This species also exhibited activity all day. In mangroves, An. albimanus showed very low biting activity relative to An. neivai , with two small peaks at 09:00 pm and 11:00 pm ( Fig. 4 ).
: man biting activity in extradomicile (mangrove) of Anopheles neivai and Anopheles albimanus (geometric mean: 95% confidence interval) near of municipality of Iscuandé, Las Varas (sampling for 24 h; n = 3,237).
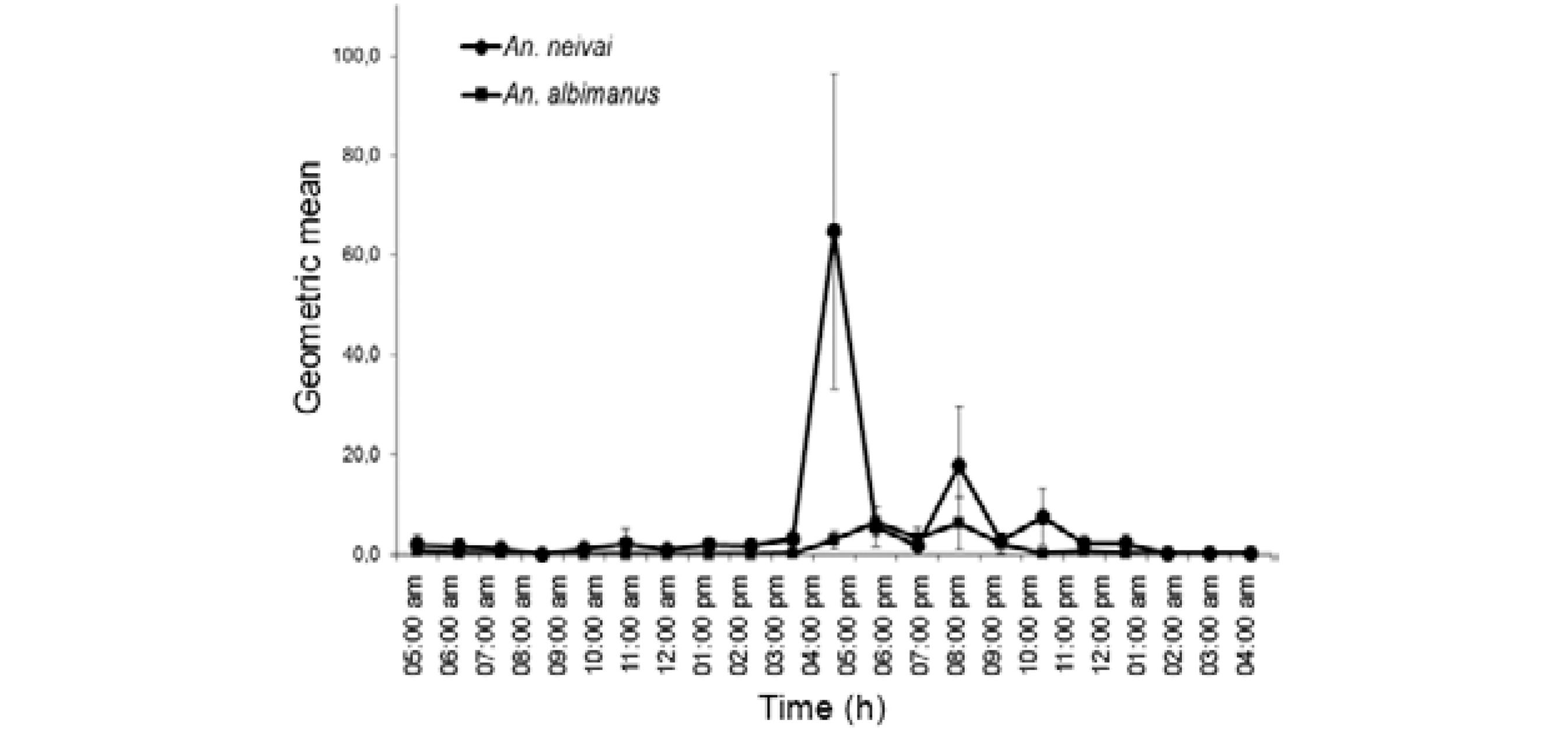
Canoes - In canoes, An . neivai was much more abundant than An. Albimanus, with a relative abundance of 90%. This mosquito displayed three peaks of activity: the first between 05:00 am-06:00 am (HBR: 8,3 m/m/h; RV: 3 m/m/h- 22 m/m/h), the second between 05:00 pm-07:00 pm (16,7 m/m/h; RV: 4 m/m/h-24 m/m/h) and the third between 08:00 pm-10:00 pm (12 m/m/h; RV: 6 m/m/h-30 m/m/h) ( Fig. 5 ). An. neivai and An . albimanus were detected up to 200 m from the coast in samplings carried out in canoes during fishing from 05:00 pm-07:00 pm.
: man biting activity in extradomicile (canoe) of Anopheles neivai and Anopheles albimanus (geometric mean: 95% confidence interval), municipality of Iscuandé, Las Varas (sampling for 24 h; n = 896).
Fisherman activities - Fishing in the studied area is mostly artisanal and consists of various activities that place the fishermen at risk of being bitten by Anopheles. Most of these activities are performed outside the home.
Piangua ( Anadara tuberculosa ) is a mollusc that is collected in the mangroves at low tide by women and children when they finish their schoolwork. This collection usually occurs between 07:00 am-10:00 am and between 01:00 pm-03:00 pm. These individuals must enter the mangroves to remove the shells that are attached to tree roots, thus exposing themselves to An. neivai . Similarly, native children catch crabs (Arthropoda: Crustacea: Decapoda), which are sold in the nearby markets, in mangroves. The capture of these arthropods can be performed all day and throughout the year. During this activity, children may be bitten by mosquitoes.
Fishermen use nets of several metres in length constructed of 2.5” mesh, known as trasmallo , to capture shrimp. This net is connected to the mangroves; at high tide, fish enter into flooded areas and then they become trapped in the nets at low tide. Fishermen assemble these nets between 05:00 am-01:00 pm, during which they may be bitten by An. neivai .
Another method utilises nets of 4” mesh, which are used to capture catfish ( Bagre pinnomaculatus ), snooks ( Centropomus undecimalis ), sierra ( Scomberomorus sierra ) and green jack ( Caranx caballus ), among other fish. This net is assembled in a similar manner, but between 06:00 pm-06:00 am. In both cases, fishermen may be bitten by mosquitoes when they are either in the mangrove to tie the net or in the canoe to gather the fish that have been caught.
Other fishing activities are performed exclusively by canoe and fishermen leave the canoes only to land. Many fishermen use homemade hooks that contain baits (known as calandros ) to capture fish such as snappers ( Lutjanus spp), Panama hake ( Merluccius angustimanus ), flathead sea catfish ( Arius planiceps and Arius platypogon ) and chihuil sea catfish ( Bagre panamensis ). Fishermen paddle their vessels around the mangroves and estuaries, searching for the best places to throw the calandro . Once they have found a good place, they wait patiently to catch the quota of fish required for their livelihood. This activity is usually performed between 5:00 am-07:00 pm.
Another important fishing activity is net fishing, where two or three fishermen work together to place the net into the sea and collect all the fish that have been caught. Depending on the type of fish they are seeking, they may stay only a few metres from the mangrove or go far offshore. This activity is performed over a broad time period, ranging from 04:00 am-12:00 am and it coincides with the three peaks of An. neivai biting activity.
Poor families that cannot afford an outboard motor, which would provide access to more fishing sites and reach at least 1,000 m offshore, use a small traditional canoe; thus, their mobility depends on the use of paddles. In such canoes, they navigate through the mangrove fringe at very low speeds, fishing with simple bait or small nets. Often, they spend several hours waiting at the edge of the mangrove for the fish to bite, an action that places them at risk of being bitten by An. neivai.
Natural infection - A total of 6,382 female mosquitoes were processed using the ELISA technique. Three specimens (0.05%) were infected with Plasmodium . Table II shows the field code, the species infected, the capture site, date and time and the Plasmodium species detected. Two females of An. neivai (2/5222 = 0.04%) were found infected with P. vivax VK247 and P. vivax VK210. One female An. albimanus (1/1160 = 0.09%) was infected with P. vivax VK210.
Entomological inoculation rate (EIR) - The EIR ( Warrell & Gilles 2002Warrell DA, Gilles HM 2002: Essential malariology , 4th ed., A Hodder Arnold Publication, London, 384 pp. ) was calculated from the product of the HBR (average of 24 h) and the sporozoite rate at each sampling site where mosquitoes were found infected with Plasmodium ( Table II ).
DISCUSSION
An important aim of this study was to determine whether An. neivai can reach fishing canoes and bite fishermen while they are performing their daily activities. It was possible to calculate the HBR for six samples of continuous 24-h sampling and for eight samples at peak activity, which indicates that this species reaches the boats and canoes not by accident, but rather as a result of “habitual” behaviour.
This study reports the presence of An. neivai and An. albimanus that bite fishermen in their canoes, demonstrating that mosquitoes can leave the swamp and fly hundreds of metres to bite fishermen. Mosquito activity was detected up to 200 m offshore and according to the fishermen themselves; mosquitoes can reach canoes located up to 500-800 m from shore.
The natural infection results ( Table II ) showed that three female Anopheles were found infected with P. vivax . Two (1 An. neivai and 1 An. albimanus ) were caught landing on fishermen in their canoes. The EIR values for each site were low, presumably due to the low biting rate observed in the early hours. Nevertheless, this result, coupled with the HBR in the canoes and mangroves at peak activity and confirmed by epidemiological data from the area, supports the argument that fishing is an “occupational” risk factor for contracting malaria.
An. neivai peaked at dawn in the canoes, which was not observed at any of the other sampling sites, even in the mangroves. It is possible that populations of An. neivai that are present in the mangroves feed at dawn on the fishermen in their canoes because there are no other humans at that time in the mangrove. Likewise, canoes concentrate people, which can enhance the attraction of mosquitoes. There are always between two-five fishermen in each boat. For example, while some fishermen are approaching the net, the others are removing the fish that have been caught in it. Fishing nets can weigh more than 200 kg; therefore, at least two people are required to move them.
The dawn peak activity of An. neivai (17.7 m/m/h) coincides with the boarding of fishermen, who carry out their fishing activities between 05:00 am-04:00 pm. At the time of boarding and even when they are in their canoes, they are at risk of being bitten by An. neivai .
In this study, An. neivai showed preferences for extradomiciliary biting activity, but it also showed constant intradomiciliary activity. Although most Kerteszia species are characterised by exophagic preferences ( Marrelli et al. 2007Marrelli MT, Malafronte RS, Sallum MAM, Natal D 2007. Kerteszia subgenus of Anopheles associated with the Brazilian Atlantic rainforest: current knowledge and future challenges. Malar J 2007, 6 : 127. ), there have been reports of some species of this subgenus that prefer anthropic environments over forest environments. Forattini et al. (1999)Forattini OP, Kakitani I, dos Santos RC, Ueno HM, Kobayashi KM 1999. Role of Anopheles (Kerteszia) bellator as malaria vector in southeastern Brazil (Diptera: Culicidae). Mem Inst Oswaldo Cruz 94 : 715-718. found that Anopheles (K) bellator was four times more abundant indoors than Anopheles (K) cruzii , which showed an inclination for sylvatic environments.
The few previous studies that have examined the Kerteszia subgenus in Colombia have identified little or no indoor activity. Quiñones et al. (1984)Quiñones M, Suárez M, Rodríguez A, Fleming G, Galvis L 1984. Comportamiento de Anopheles (Kerteszia) lepidotus Zavortink, 1973, y su incriminación como posible vector de malaria en el Departamento del Tolima, Colombia. Biomédica 4 : 5-13. did not detect indoor activity for Anopheles (K) pholidotus ( referred to as Anopheles lepidotus ). In relation to An. (K) neivai , Astaiza et al. (1988)Astaiza R, Murillo C, Fajardo P 1988. Biología de Anopheles (Kerteszia) neivai h., d. & k., 1913 (Diptera: Culicidae) en la Costa Pacífica de Colombia. Fluctuación de la población adulta. Rev Saude Publica 22 : 101-108. reported some domiciliary activity on the Pacific Coast during indoor sampling between 07:00 pm-05:00 am and they detected peak activity at 01:00 am. Solarte et al. (1996)Solarte Y, Hurtado C, Gonzalez R, Alexander B 1996. Man-biting Activity of Anopheles (Nyssorhynchus) albimanus and An . (Kerteszia) neivai (Diptera: Culicidae) in the Pacific lowlands of Colombia. Mem Inst Oswaldo Cruz 91 : 141-146. reported no significant indoor activity for this species in distinct settlements of the Naya River (Colombia).
It is possible that this species has inherent synanthropic potential or may be seeking new environments in which to feed due to external pressures, such as decreases of their original food sources or degradation of their natural environment. Moreover, many anophelines may show temporary or permanent changes in their biting behaviour in response to extrinsic factors, such as moon phases, rain, deforestation and the use of insecticides ( Reddy et al. 2011Reddy MR, Overgaard HJ, Abaga S, Reddy VP, Caccone A, Kiszewski AE, Slotman MA 2011. Outdoor host seeking behaviour of Anopheles gambiae mosquitoes following initiation of malaria vector control on Bioko Island, Equatorial Guinea. Malar J 10 : 184 ). The levels of domestication of mosquito species can vary by location and show different degrees of endophily and endophagy. These differences can be observed in the interactions between local ecological factors, such as the distance between houses and oviposition sites, the housing type, the time of year and biological-genetic specific factors such as heredity and age, which influence biting behaviour. Furthermore, it has been suggested that important differences in behaviour may indicate that the populations of interest are part of a species complex ( Rubio-Palis & Curtis 1992Rubio-Palis Y, Curtis CF 1992. Biting and resting behaviour of anophelines in western Venezuela and implications for control of malaria transmission. Med Vet Entomol 6 : 325-334. , Rubio-Palis et al. 2013Rubio-Palis Y, Bevilacqua M, Medina DA, Moreno JE, Cárdenas L, Sánchez V, Estrada Y, Anaya W, Martínez Á 2013. Malaria entomological risk factors in relation to land cover in the Lower Caura River Basin, Venezuela. Mem Inst Oswaldo Cruz 108 : 220-228. ).
As An. neivai breeds exclusively in the jungle and this species has exophilic preferences, females must travel between the forests and houses to achieve their food-oviposition requirements. Thus, the distance between dwellings and the mangrove forest is a key factor contributing to the development of endophagic behaviour in this species. Confirming this view, several studies have shown that dwelling closer to the forest is a risk factor for contracting malaria ( Rozendaal 1992Rozendaal JA 1992. Relations between Anopheles darlingi breeding habitats, rainfall, river level and malaria transmission rates in the rain forest of Suriname. Med Vet Entomol 6 : 16-22. , Stefani et al. 2011Stefani A, Hanf M, Nacher M, Girod R, Carme B 2011. Environmental, entomological, socioeconomic and behavioural risk factors for malaria attacks in Amerindian children of Camopi, French Guiana. Malar J 10 : 246. ).
The risk of being bitten by An. neivai in the dwellings is exacerbated because most of the dwellings in the study area are built of wood, have incomplete walls and lack glass windows. The peak indoor biting activity (06:00 pm-07:00 pm) creates a risk factor for the local population because they can be bitten by An. neivai at a time when most of them have not yet gone to bed. Stefani et al. (2011) Stefani A, Hanf M, Nacher M, Girod R, Carme B 2011. Environmental, entomological, socioeconomic and behavioural risk factors for malaria attacks in Amerindian children of Camopi, French Guiana. Malar J 10 : 246. found that children who go to bed after 07:00 pm have a higher risk of contracting malaria than those who go to bed earlier because the latter are protected by bednets at the time of peak anopheline biting activity.
An. neivai was more active at night outdoors than indoors and showed a bite rate lower than that reported by Astaiza et al. (1988)Astaiza R, Murillo C, Fajardo P 1988. Biología de Anopheles (Kerteszia) neivai h., d. & k., 1913 (Diptera: Culicidae) en la Costa Pacífica de Colombia. Fluctuación de la población adulta. Rev Saude Publica 22 : 101-108. , but higher than that reported by Solarte et al. (1996)Solarte Y, Hurtado C, Gonzalez R, Alexander B 1996. Man-biting Activity of Anopheles (Nyssorhynchus) albimanus and An . (Kerteszia) neivai (Diptera: Culicidae) in the Pacific lowlands of Colombia. Mem Inst Oswaldo Cruz 91 : 141-146. . An. neivai only showed one peak of activity (05:00 pm-06:00 pm), but it coincided with a variety of activities around the home, including chatting with neighbours, sharing work experiences, playing games and other tasks such as making or repairing fishing nets. The only option for inhabitants to avoid being bitten is to drive away insects with smoke from fires using the skin of coconut palms that are plentiful in the area. This measure, although inexpensive, is problematic because the smoke generated by burning coconut skin irritates the respiratory system. Furthermore, its effectiveness depends on the availability of this fruit in the zone and the combustion duration, which averages 15 min.
The forest activity of An. neivai has been reported previously ( Solarte et al. 1996Solarte Y, Hurtado C, Gonzalez R, Alexander B 1996. Man-biting Activity of Anopheles (Nyssorhynchus) albimanus and An . (Kerteszia) neivai (Diptera: Culicidae) in the Pacific lowlands of Colombia. Mem Inst Oswaldo Cruz 91 : 141-146. , Astaiza et al. 1988Astaiza R, Murillo C, Fajardo P 1988. Biología de Anopheles (Kerteszia) neivai h., d. & k., 1913 (Diptera: Culicidae) en la Costa Pacífica de Colombia. Fluctuación de la población adulta. Rev Saude Publica 22 : 101-108. ). However, this is the first time that biting activity has been measured for 24 continuous hours in the mangrove forest and related to the activities of fisherman. An. neivai presented a trimodal pattern of biting activity and this study has the highest reported biting activity for this species. Two of the three activity peaks coincided in time (between 06:00 pm-09:00 am and 09:00 pm-10:00 pm), as described by Solarte et al. (1996)Solarte Y, Hurtado C, Gonzalez R, Alexander B 1996. Man-biting Activity of Anopheles (Nyssorhynchus) albimanus and An . (Kerteszia) neivai (Diptera: Culicidae) in the Pacific lowlands of Colombia. Mem Inst Oswaldo Cruz 91 : 141-146. . The third peak, close to midnight, is reported here for the first time for this species. At the end of this peak (at 01:00 am), a female An. neivai infected with P. vivax was captured. An activity peak was not detected at sunrise. Previous reports ( Solarte et al. 1996Solarte Y, Hurtado C, Gonzalez R, Alexander B 1996. Man-biting Activity of Anopheles (Nyssorhynchus) albimanus and An . (Kerteszia) neivai (Diptera: Culicidae) in the Pacific lowlands of Colombia. Mem Inst Oswaldo Cruz 91 : 141-146. ) have described a small peak of activity for An. neivai between 06:00 am-07:00 am and studies investigating the subgenus Kerteszia indicate that this species has two peaks of activity in sylvatic areas: one at sunrise and another at sunset ( Forattini et al. 1996Forattini OP, Kakitani I, Massad E, Marucci D 1996. Studies on mosquitoes (Diptera: Culicidae) and anthropic environment. 11. Biting activity and blood-seeking parity of Anopheles (Kerteszia) in south-eastern Brazil. Rev Saude Publica 30 : 107-114. , Marrelli et al. 2007Marrelli MT, Malafronte RS, Sallum MAM, Natal D 2007. Kerteszia subgenus of Anopheles associated with the Brazilian Atlantic rainforest: current knowledge and future challenges. Malar J 2007, 6 : 127. ). All of the peaks of mosquito activity coincided with one or more times when fishing activities were being performed in the area.
Despite the low daytime activity of An. neivai reported in this study, women and children are primarily at risk of being bitten by this mosquito during the day, when they enter the mangroves to catch molluscs and crabs. These activities, which help them pay their school fees, are performed almost every day. They must remain within the mangrove forest for 3-6 h and are virtually unprotected against mosquitoes and other bloodsucking insects.
Similarly, fishermen are forced to spend several hours at the edge of mangroves as they set the nets and carefully remove fish that have fallen into them. To repel mosquitoes, they burn coconut husks or apply oil-derived substances to the skin, which, according to the fishermen, causes them discomfort and skin irritation.
This study has demonstrated a temporal and spatial coincidence between several fishing activities performed outdoors and peaks of An. neivai biting activity. This relationship, combined with the detection of infection by Plasmodium in female mosquitoes caught in canoes, suggests that An. neivai may be responsible for extradomiciliary transmission of malaria, mainly in mangroves and canoes. This type of malaria transmission is common in the Kerteszia subgenus and several of its species have been associated with malaria transmission in forest environments, which is known as bromeliad-malaria ( Ueno et al. 2007Ueno HM, Forattini OP, Kakitani I 2007. Vertical and seasonal distribution of Anopheles (Kerteszia) in Ilha Comprida, Southeastern Brazil. Rev Saude Publica 41 : 269-275. ). Similarly, Valderrama and Santander (1985)Valderrama FG, Santander AG 1985. Riesgo de infección extradomiciliar de la malaria en diferentes ámbitos sociológicos ocupacionales en Bajo Calima, Buenaventura, Colombia . MSc Thesis, University of Valle, Cali, 191 pp. found that 83% of malaria cases were contracted outside the home in a study on the Bajo Calima, where An. neivai is abundant.
Moreover, it has been suggested for over a decade that this species may be the primary malaria vector in some areas of the Pacific Colombian Coast, given its strong association with humans, its high densities, its constant presence in coastal areas ( Astaiza et al. 1988Astaiza R, Murillo C, Fajardo P 1988. Biología de Anopheles (Kerteszia) neivai h., d. & k., 1913 (Diptera: Culicidae) en la Costa Pacífica de Colombia. Fluctuación de la población adulta. Rev Saude Publica 22 : 101-108. , Solarte et al. 1996Solarte Y, Hurtado C, Gonzalez R, Alexander B 1996. Man-biting Activity of Anopheles (Nyssorhynchus) albimanus and An . (Kerteszia) neivai (Diptera: Culicidae) in the Pacific lowlands of Colombia. Mem Inst Oswaldo Cruz 91 : 141-146. ) and its natural infection with Plasmodium ( Carvajal et al. 1989Carvajal H, de Herrera MA, Quintero J, Alzate A, Herrera S 1989. Anopheles neivai : a vector of malaria in the Pacific low lands of Colombia. Trans R Soc Trop Med Hyg 83 : 609. , Gutiérrez et al. 2008Gutiérrez LA, Naranjo N, Jaramillo LM, Muskus C, Luckhart S, Conn JE, Correa M 2008. Natural infectivity of Anopheles species from the Pacific and Atlantic Regions of Colombia. Acta Tropica 107 : 99-105 ).
In the department of Nariño, most malaria control strategies have targeted intradomiciliary
mosquitoes through the provision of treated mosquito bednets. Municipal health services have
distributed free mosquito bednets and coverage reaches nearly 100% of the population.
However, this measure is not effective in preventing peri and extradomiciliary malaria
transmission in Nariño. Because Nariño is one of the eight poorest departments in the
country, where 56.1% of its inhabitants are under the poverty line ( DNP 2011DNP - Dirección Nacional de Planeación 2011. Indicadores de pobreza por
ingreso. Nueva metodología. Avalaible from: dnp.gov.co.
dnp.gov.co...
), it is likely that most people in the study area are not
accustomed to using commercial repellents because they cannot afford them. It is suggested
that low-cost commercial repellents against malaria be supplied by the departmental program
after implementing an education campaign and training the community to optimise repellent
use.
ACKNOWLEDGEMENTS
To Freddy Ruiz (Walter Reed Biosystematic Unit/Smithsonian Institution), for his support in generating DNA sequences, and to Lucio Góngora and the community of La Ensenada, for their hospitality.
REFERENCES
- Aponte SL, Díaz G, Pava Z, Echeverry DF, Ibarguen D, Rios M, Murcia LM, Quelal C, Murillo C, Gil P, Björkman A, Osorio L 2011. Sentinel network for monitoring in vitro susceptibility of Plasmodium falciparum to antimalarial drugs in Colombia: a proof of concept. Mem Inst Oswaldo 106 (Suppl. I): 123-129.
- Astaiza R, Murillo C, Fajardo P 1988. Biología de Anopheles (Kerteszia) neivai h., d. & k., 1913 (Diptera: Culicidae) en la Costa Pacífica de Colombia. Fluctuación de la población adulta. Rev Saude Publica 22 : 101-108.
- Barbieri AF, Sawyer DO 2007. Heterogeneidade da prevalência de malária em garimpos do norte de Mato Grosso, Brasil. Cad Saude Publica 23 : 2878-2886.
- Beier JC, Asiago CM, Onyango FK, Koros JK 1988. ELISA absorbance cut-off method affects malaria sporozoite rate determination in wild Afrotropical Anopheles . Med Vet Entomol 2 : 259-264.
- Carvajal H, de Herrera MA, Quintero J, Alzate A, Herrera S 1989. Anopheles neivai : a vector of malaria in the Pacific low lands of Colombia. Trans R Soc Trop Med Hyg 83 : 609.
- Clark J, Kelly K 1993. Human genetics, paleoenvironments and malaria: relationships and implications for the settlement of Oceania. Am Anthropol 95 : 612-630.
- Collins FH, Paskewitz SM 1996. A review of the use of ribosomal DNA (rDNA) to differentiate among cryptic Anopheles species. Insect Mol Biol 5 : 1-9.
- da Silva NS, da Silva-Nunes M, Malafronte RS, Menezes MJ, D’Arcadia RR, Komatsu NT, Scopel KK, Braga EM, Cavasini CE, Cordeiro JA, Ferreira MU 2010. Epidemiology and control of frontier malaria in Brazil: lessons from community-based studies in rural Amazonia. Trans R Soc Trop Med Hyg 104 : 343-350.
- DNP - Dirección Nacional de Planeación 2011. Indicadores de pobreza por ingreso. Nueva metodología. Avalaible from: dnp.gov.co.
» dnp.gov.co - Dolo G, Briet OJ, Dao A, Traore SF, Boaure M, Sogoba N 2004. Malaria transmission in relation to rice cultivation in the irrigated Sahel of Mali. Acta Trop 89 : 147-159.
- Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol 3 : 294-299.
- Forattini OP, Kakitani I, Massad E, Marucci D 1996. Studies on mosquitoes (Diptera: Culicidae) and anthropic environment. 11. Biting activity and blood-seeking parity of Anopheles (Kerteszia) in south-eastern Brazil. Rev Saude Publica 30 : 107-114.
- Forattini OP, Kakitani I, dos Santos RC, Ueno HM, Kobayashi KM 1999. Role of Anopheles (Kerteszia) bellator as malaria vector in southeastern Brazil (Diptera: Culicidae). Mem Inst Oswaldo Cruz 94 : 715-718.
- González R, Carrejo N 2009. Introducción al estudio taxonómico de Anopheles de Colombia. Claves y notas de distribución , 1st ed., Universidad del Valle, Cali, 237 pp.
- Gutiérrez LA, Naranjo N, Jaramillo LM, Muskus C, Luckhart S, Conn JE, Correa M 2008. Natural infectivity of Anopheles species from the Pacific and Atlantic Regions of Colombia. Acta Tropica 107 : 99-105
- Jaremin B, Nahorski W, Goljan J, Felczak-Korzybska I, Górski J, Myjak P, Kotłowski A 1993. Malaria as an occupational disease. Bull Inst Marit Trop Med Gdynia 44-45 : 43-50.
- Kumar S, Dudley J, Nei M, Tamura K 2008. MEGA: a biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief Bioinform 9 : 299-306.
- Linton YM, Harbach RE, Anthony TG, Chang MS, Asmad M 2001. Morphological and molecular identity of Anopheles (Cellia) sundaicus (Diptera: Culicidae), the nominotypical member of a malaria vector species complex in Southeast Asia. Syst Entomol 26 : 357-366.
- Maddison DR, Maddison WR 2003. MacClade v.4.06: analysis of phylogeny and character evolution, Sinauer Associates, Sunderland, MA, USA.
- Marrelli MT, Malafronte RS, Sallum MAM, Natal D 2007. Kerteszia subgenus of Anopheles associated with the Brazilian Atlantic rainforest: current knowledge and future challenges. Malar J 2007, 6 : 127.
- Mboera LEG, Senkoro KP, Mayala BK, Rumisha SF, Rwegoshora RT, Mlozi MR, Shayo EH 2010. Spatio-temporal variation in malaria transmission intensity in five agro-ecosystems in Mvomero district, Tanzania. Geospat Health 4 : 167-178.
- Montoya-Lerma J, Solarte YA, Giraldo-Calderón GI, Quiñones ML, Ruiz-López F, Wilkerson RC, González R 2011. Malaria vector species in Colombia - A review. Mem Inst Oswaldo Cruz 106 (Suppl. I): 223-238.
- Murillo C, Astaiza V, Fajardo P 1988. Biología de Anopheles (Kerteszia) neivai H., D. & K., 1913 (Diptera: Culicidae) en la Costa Pacífica de Colombia. I. Fluctuación de la población larval y caracteristicas de sus criaderos. Rev Saude Publica: 22 : 94-100.
- Olano V, Brochero H, Sáenz R, Quiñones M, Molina J 2001. Mapas preliminares de la distribución de Anopheles vectores de malaria en Colombia. Biomédica 21 : 402-403.
- Padilla JC, Alvarez G, Montoya R, Chaparro P, Herrera S 2011. Epidemiology and control of malaria in Colombia. Mem Inst Oswaldo Cruz 106 (Suppl. I): 114-122.
- Quiñones M, Suárez M, Rodríguez A, Fleming G, Galvis L 1984. Comportamiento de Anopheles (Kerteszia) lepidotus Zavortink, 1973, y su incriminación como posible vector de malaria en el Departamento del Tolima, Colombia. Biomédica 4 : 5-13.
- Rajasekhar M, Nandakumar NV 2000. Occupational malaria and health risk among select occupational health care employee groups in an urban hospital at Tirupati, AP. Indian J Malariol 37 : 53-60.
- Reddy MR, Overgaard HJ, Abaga S, Reddy VP, Caccone A, Kiszewski AE, Slotman MA 2011. Outdoor host seeking behaviour of Anopheles gambiae mosquitoes following initiation of malaria vector control on Bioko Island, Equatorial Guinea. Malar J 10 : 184
- Rozendaal JA 1992. Relations between Anopheles darlingi breeding habitats, rainfall, river level and malaria transmission rates in the rain forest of Suriname. Med Vet Entomol 6 : 16-22.
- Rubio-Palis Y, Bevilacqua M, Medina DA, Moreno JE, Cárdenas L, Sánchez V, Estrada Y, Anaya W, Martínez Á 2013. Malaria entomological risk factors in relation to land cover in the Lower Caura River Basin, Venezuela. Mem Inst Oswaldo Cruz 108 : 220-228.
- Rubio-Palis Y, Curtis CF 1992. Biting and resting behaviour of anophelines in western Venezuela and implications for control of malaria transmission. Med Vet Entomol 6 : 325-334.
- Ruiz F, Linton Y-M, Ponsonby DJ, Conn JE, Herrera M, Quiñones ML, Vélez ID, Wilkerson RC 2010. Molecular comparison of topotypic specimens confirms Anopheles (Nyssorhynchus) dunhami Causey (Diptera: Culicidae) in the Colombian Amazon. Mem Inst Oswaldo Cruz 105: 899-903.
- Sanabria H, Hernández A, Villafuerte A, Erazo P 2004. Percepción y condiciones de trabajo de personas que tuvieron malaria. Rev Peru Med Exp Salud Publica 21 : 210-216.
- Sevilla-Casas E 1993. Human mobility and malaria risk in the Naya River basin of Colombia. Social Sci Med 37 : 1155-1167.
- Solarte Y, Hurtado C, Gonzalez R, Alexander B 1996. Man-biting Activity of Anopheles (Nyssorhynchus) albimanus and An . (Kerteszia) neivai (Diptera: Culicidae) in the Pacific lowlands of Colombia. Mem Inst Oswaldo Cruz 91 : 141-146.
- Stefani A, Hanf M, Nacher M, Girod R, Carme B 2011. Environmental, entomological, socioeconomic and behavioural risk factors for malaria attacks in Amerindian children of Camopi, French Guiana. Malar J 10 : 246.
- Ueno HM, Forattini OP, Kakitani I 2007. Vertical and seasonal distribution of Anopheles (Kerteszia) in Ilha Comprida, Southeastern Brazil. Rev Saude Publica 41 : 269-275.
- Valderrama FG, Santander AG 1985. Riesgo de infección extradomiciliar de la malaria en diferentes ámbitos sociológicos ocupacionales en Bajo Calima, Buenaventura, Colombia . MSc Thesis, University of Valle, Cali, 191 pp.
- Warrell DA, Gilles HM 2002: Essential malariology , 4th ed., A Hodder Arnold Publication, London, 384 pp.
- Wirtz RA, Burkot TR, Andre RG, Rosenberg R, Collins WE, Roberts DR 1985. Identification of Plasmodium vivax sporozoites in mosquitoes using an enzyme-linked immunosorbent assay. Am J Trop Med Hyg 34 : 1048-1054.
- Wirtz RA, Zavala F, Charoenvit Y, Campbell GH, Burkot TR, Schneider I, Esser KM, Beaudoin RL, Andre RG 1987. Comparative testing of monoclonal antibodies against Plasmodium falciparum sporozoites for ELISA development. Bull WHO 65 : 39-45.
- Zavortink TJ 1973. Mosquitoes studies (Diptera: Culicidae). XXIX. A review of the subgenus Kerteszia of Anopheles . Contr Amer Ent Inst 9 : 1-59.
Publication Dates
-
Publication in this collection
6 Dec 2013
History
-
Received
10 May 2013 -
Accepted
31 Oct 2013