Abstracts
Nyssomyia intermedia (Lutz & Neiva) and Nyssomyia neivai (Pinto) are morphologically very close and both present great variation in some structures. The objective of this study is a description of the variation among the females of these species in populations from the States of Minas Gerais and São Paulo. The morphological structures studied were the number of horizontal teeth in the cibarium and the number of rings and the shape of the terminal knob of the spermathecae. The spermatheca rings are significantly more numerous in N. intermedia than in N. neivai and the simple shape of the terminal knob predominated in both species. Regarding the cibarium, eight to eleven teeth have been found in both species, with up to twelve teeth in the latter. The number of horizontal teeth and the shape of the terminal knob of the spermathecae were variable throughout the populations of both species and all structures were polymorphic in the populations studied.
Sandflies; leishmaniosis; vectors
Nyssomyia intermedia (Lutz & Neiva) e Nyssomyia neivai (Pinto) são muito semelhantes morfologicamente e ambas apresentam grande variação em algumas estruturas. O objetivo deste estudo foi descrever a variação ocorrida em fêmeas destas espécies, provenientes dos Estados de Minas Gerais e São Paulo. As estruturas morfológicas estudadas foram o número de dentes horizontais no cibário e o número de anéis e forma da cabeça da espermateca. O número de anéis nas espermatecas foi significativamente maior em N. intermedia e forma da cabeça da espermateca simples predominou em ambas. Com relação ao cibário, foi encontrada uma variação de oito a onze dentes horizontais nas duas espécies, chegando a doze em N. intermedia. O número de dentes horizontais e a forma da cabeça da espermateca foram variáveis em todas as populações estudadas e todas as estruturas pesquisadas foram consideradas polimórficas.
Flebotomíneo; leishmaniose; vetores
BIOLOGY, ECOLOGY AND DIVERSITY
Polymorphism, inter-population and inter-specific variation in Nyssomyia intermedia (Lutz & Neiva) and Nyssomyia neivai (Pinto) (Diptera, Psychodidae, Phlebotominae)
Polimorfismo, variação interpopulação e interespecífica em Nyssomyia intermedia (Lutz & Neiva) e Nyssomyia neivai (Pinto) (Diptera, Psychodidae, Phlebotominae)
José Dilermando Andrade FilhoI; Eunice A. Bianchi GalatiII; Alda Lima FalcãoI
ILaboratório de Leishmanioses, Centro de Pesquisas René Rachou/Fiocruz, Av. Augusto de Lima, 1715, Barro Preto, 30190-002 Belo Horizonte-MG, Brasil. jandrade@cpqrr.fiocruz.br
IIDepartamento de Epidemiologia, Faculdade de Saúde Pública, Universidade de São Paulo, Av. Dr. Arnaldo, 715, 01246-904, São Paulo-SP, Brasil. egalati@usp.br
ABSTRACT
Nyssomyia intermedia (Lutz & Neiva) and Nyssomyia neivai (Pinto) are morphologically very close and both present great variation in some structures. The objective of this study is a description of the variation among the females of these species in populations from the States of Minas Gerais and São Paulo. The morphological structures studied were the number of horizontal teeth in the cibarium and the number of rings and the shape of the terminal knob of the spermathecae. The spermatheca rings are significantly more numerous in N. intermedia than in N. neivai and the simple shape of the terminal knob predominated in both species. Regarding the cibarium, eight to eleven teeth have been found in both species, with up to twelve teeth in the latter. The number of horizontal teeth and the shape of the terminal knob of the spermathecae were variable throughout the populations of both species and all structures were polymorphic in the populations studied.
Keywords: Sandflies; leishmaniosis; vectors.
RESUMO
Nyssomyia intermedia (Lutz & Neiva) e Nyssomyia neivai (Pinto) são muito semelhantes morfologicamente e ambas apresentam grande variação em algumas estruturas. O objetivo deste estudo foi descrever a variação ocorrida em fêmeas destas espécies, provenientes dos Estados de Minas Gerais e São Paulo. As estruturas morfológicas estudadas foram o número de dentes horizontais no cibário e o número de anéis e forma da cabeça da espermateca. O número de anéis nas espermatecas foi significativamente maior em N. intermedia e forma da cabeça da espermateca simples predominou em ambas. Com relação ao cibário, foi encontrada uma variação de oito a onze dentes horizontais nas duas espécies, chegando a doze em N. intermedia. O número de dentes horizontais e a forma da cabeça da espermateca foram variáveis em todas as populações estudadas e todas as estruturas pesquisadas foram consideradas polimórficas.
Palavras-chave: Flebotomíneo; leishmaniose; vetores.
Nyssomyia intermedia (Lutz & Neiva, 1912) and Nyssomyia neivai (Pinto, 1926) are very close species, separated only by the females' spermatheca structures and the male genital filament tips (Marcondes 1996, Andrade Filho et al. 2003). The biology of the two species also shows a difference in the time of development of the second and third larval instars (Andrade Filho et al. 2004).
Both species are suspected of transmitting cutaneous leishmaniasis due to their very high frequencies at foci of the diseases in southern and southeastern regions of Brazil (Andrade Filho 2003) and to the fact that they have been found naturally infected by flagellates by Rangel et al. (1984) (N. intermedia), Forattini et al. (1972) and Casanova et al. (1995) (N. neivai, presumably).
Both species present great inter and intra-populational polymorphism. Thus, for N. intermedia and N. neivai, six different palpal formulae (Marcondes et al. 1998); the number of horizontal teeth of cibarium varying from 8 to 12 and great variation in the number of spermatheca rings (Marcondes 1996, Marcondes 1997, Marcondes et al. 1998, Andrade Filho et al. 2003) have been observed. The dimensions of N. intermedia are influenced by altitude, latitude and period of capture (Marcondes et al. 1999).
This study deals with the morphological variation inter-population, inter-specific and polymorphism of the females of several populations of N. intermedia and N. neivai.
MATERIAL AND METHODS
Five populations of each species were analyzed, being two of the populations of N. intermedia captured in Lassance (19º54'S; 44º48'W), in the Brazilian state of Minas Gerais, one captured in an area with "cerrado" vegetation and the other in a chicken house, situated about 5 km apart. The third population was from the peridomicile of a house in Além Paraíba, Minas Gerais (21º53'S; 42º42'W). The two other populations were both from Iporanga (24º37'S; 48º35'W), in São Paulo State, one from the peridomicile of a house and the other from a pigsty situated close to a wooded area (Atlantic forest), these two sites being about 700 m apart.
Populations of N. neivai were also sampled from both Iporanga sites, as well as from the peridomicile of a house in Corinto, on the banks of the Rio das Velhas in Minas Gerais (17º58'S; 49º32'W). Other populations of N. neivai sampled were from Barra do Rio Cabo Verde and Sapucaí, Alfenas, Minas Gerais (21º18'S; 46ºW) and finally from Conchal, São Paulo (22º18'S; 47º13'W). Specimens of this last population were kindly provided by Dr. Cláudio Casanova.
The specimens were observed under an optical microscope, Olympus®, CH-2 model, after clearing by method described by Forattini (1973) and mounting on microscope slides in Balsam of Canada resin or Berlese's fluid.
Cibarium and spermathecae were compared. The number of horizontal teeth in the cibarium was counted and the numbers of rings on the two spermathecae were counted and the shape of the terminal knob of the spermathecae classified as simple, simple/bilobed, bilobed, bilobed/trilobed or trilobed.
For the statistical test, a variance analysis (ANOVA) was carried out, with 5% probability to compare the two species. The three structures studied were analyzed isolated and compared as between the two species and each population. A score was attributed for the shape of the terminal knob of the spermathecae: simple (1), simple/bilobed (2), bilobed (3), bilobed/trilobed (4).
The classification utilized is that proposed by Galati (2003).
RESULTS
Inter-specific Variation
Four hundred and two females of N. intermedia and 196 of N. neivai were analyzed. The number of rings on each spermatheca of N. intermedia was considerably greater than that on each of N. neivai (Fig. 1). For the former species, there was a variation of from eight to 17, with a mean of 11.5 (+ 1.5), the most frequent numbers being 11 (26.8%), 12 (22.5%) and ten (17.9%), while for N. neivai, the mean was 8.5 (+ 1.1), varying from six to 11 rings, the most common numbers being eight (35.9%) and nine rings (29.7%). ANOVA showed a highly significant difference (Fobs. = 1.132.0; Ftab. = 3.84).
When the number of spermatheca rings was added up (Fig. 2), the dominant values were 21 (14.4%) and 23 (17.4%) for N. intermedia and 15 (18.9%) and 17 (32.4%) for N. neivai, with a variation from 17 to 33 rings (excluding 30 and 32) for the former species and 13 to 22 for the latter.
The prevalent shape of the terminal knob of the spermathecae of N. intermedia was simple (45.8%), as also of N. neivai (Fig. 3), in this latter species, however, this shape appeared much more frequent (89.3%). The bilobed shape presented in 31.8% of N. intermedia specimens, while in N. neivai, it was rarer, presenting in only 3.4%. The simple/bilobed shape was also more frequent in N. intermedia than in N. neivai, appearing in 20.1% in the former and in 7.3% in the latter. No trilobed terminal knob of the spermatheca was found in these species and bilobed/trilobed appeared only in N. intermedia, with a much lower frequency (2.3%). ANOVA showed a significant difference (Fobs. = 105.4; Ftab. = 3.84).
The horizontal teeth of the cibaria varied from eight to twelve in N. intermedia and from eight to eleven in N. neivai (Fig. 4), and the predominant number for this species was ten (49.2%), followed by eight (36.6%), nine (13.7%) and eleven (0.6%). The mean for this species was 9.1 (+ 0.9). In N. intermedia, the mean was of 8.9 (+ 1.0) horizontal teeth in the cibarium, with eight teeth being the most frequent (49.2%), followed by ten (34.6%), nine (14.6%), eleven (1.3%) and twelve (0.3%). ANOVA showed a highly significant difference (Fobs. = 99.3; Ftab. = 3.84).
Polymorphism
Nyssomyia intermedia
Além Paraíba
Seventy five females in this population were analyzed. The number of spermatheca rings in this population varied from 8-15 (mean 11.3 + 1.50). The number of rings in the two spermathecae differed in 69.9% of the specimens. Eleven rings was the most prevalent number (26.0%), followed by 10 (24.7%) and 12 (17.8%). When the rings of a pair of spermathecae were added up, the total number varied from 17-29, specimens with 23 (17.8%), 20 (16,4%) and 21 (16.4%) predominating. The most prevalent form of terminal knob of the spermathecae was simple (50.7%), although simple/bilobed (21.9%) and bilobed (27.4%) also occurred. The mean number of horizontal teeth in the cibarium was 8.12 and 90.7% of the specimens presenting eight teeth, the remainder having nine (6.7%) or 10 (2.7%).
Lassance (chicken house)
In all 109 females of the Lassance chicken house population analyzed, 72.2% showed different numbers of rings in their two spermathecae. The number of spermatheca rings varied from 9-17 (mean 11.9 + 1.42), 12 (28.3%) the most prevalent, followed by 11 (23.5%) and 13 (19.4%). When the number of rings in the two spermathecae was added together, the total number varied from 19-33, the highest prevalence being of 25 (15.7%), followed by 26 (14.8%), 23 (13.9%) and 24 (13.0%). The simple terminal spermatheca knob predominated (52.5%), followed by the simple/bilobed (22.4%), bilobed (21.5%) and bilobed/trilobed (1.9%) forms. The number of horizontal cibarial teeth varied from 8-11 (mean 9.5+ 0.80), the form with 10 (60.0%) being the predominant number, followed by nine (21.1%), eight (16.7%) and 11 (2.2%).
Lassance (cerrado)
Of the 99 females of this population studied, 73.2% had different numbers of rings in the two spermathecae. The number of rings per spermatheca varied from 8-16 (mean 11.2 + 1.54), with 11 (29.8%), 10 (21.2%) and 12 (15.7%) predominating. The sum of spermathecal rings was 17-31, the greatest occurrence being of 22 (16.3%), 24 (14.3%), 20 (13,3%) and 23 (13.3%). The simple form of the spermatheca terminal knob was commonest (60.2%), followed by bilobed (20.4%) and simple/bilobed (19.4). The number of horizontal cibarial teeth in specimens of this population varied from 8-12 (mean 9.7 + 0.74), the highest prevalence being of 10 (67.4%), followed by nine (18.5%), eight (9.8%), 11 (3.3%) and 12 (1.1%).
Iporanga (Peridomicile)
Only 20 females of this population were analyzed and in 88.9% of these the number of rings of the spermathecae varied. The bilobed form of the terminal knob of the spermathecae predominated (60.0%), followed by the simple (25.0%), simple/bilobed (10.0%) and bilobed/trilobed (5.0%) morphs. The number of spermatheca rings varied from 8-14 (mean 11.29 + 1.39), the highest prevalence being of 11 (36.9%) and 12 (18.4%). When the rings of the pairs of spermathecae were added together, the highest prevalence was of 23 (27.8%), followed by 21, 22 and 27 (all 16.7%). Sand flies of this population had 8-10 horizontal teeth (mean 8.9 + 1.06), half of the specimens having eight, 30.0% having nine and 20.0% 10 teeth.
Iporanga (pigsty)
A large number of specimens of this population were captured at this site and 100 females analyzed. The number of rings on the two spermathecae differed in 79% of the specimens, varying from 8-17 (mean 11.6 + 1.46), with the highest prevalence (26.0%) being of 11, followed by 12 (24.0%), 10 (16.5%) and 13 (13.0%). Taking both spermathecae together the total number varied from 17-31. There were no specimens with a total of 30 and the most prevalent forms had 23 (24.0%) and 21 (18.0%) rings. The predominant form of the terminal knobs of the spermathecae was the bilobed (51.0%), followed by simple (26.0%), simple/bilobed (19.0%) and bilobed/trilobed (4.0%). The mean number of horizontal cibarial teeth in members of this population was 8.2 + 0.59, with most specimens (83.8%) presenting eight and the remaining nine or 10 teeth (8.1% each).
Nyssomyia neivai
Corinto
In all, the 51 females of this species captured at Corinto using an illuminated. Shannon trap and by aspirating from the external and internal walls of a house located about 15m away, were analyzed.
The number of rings differed in the two spermathecae in 67.4% of the specimens examined. The mean number of spermatheca rings was 8.6 (+ 1.07), the most prevalent number being eight (35.7%) and nine (32.7%). Specimens with seven or ten annulations were less prevalent (12.3 and 19.4%, respectively). When the numbers on both spermathecae of a pair were added up, a total of from 14-20 annulations was found, the majority of specimens having 17 (36.7%). The simple terminal knob of the spermathecae predominated (92.2%), although the simple/bilobed (5.9%) and bilobed (2.0%) forms were also present. The number of horizontal cibarial teeth varied from 8-10 (mean 9.6 + 0.79), the latter number predominating (77.1%).
Alfenas
Although the specimens of this population were collected and mounted on slides in 1958, we found them to be in good condition and 54 of them were studied. The number of rings differed between the spermathecae in 87.5% of the specimens. The number of spermathecal rings varied from 6-11 (mean 8.13 + 1.06), with eight (38.5%), nine (26.0%) and seven annulations (24.0%) predominating. When the numbers on the two spermathecae were added together, 17 (37.5%) and 15 (31.3%) annulations presented the greatest prevalence, although the number ranged from 13-22. The simple form of the terminal knob of the spermathecae was predominant (94.4%), followed by bilobed (3.7%) and simple/bilobed (1.9%). The number of horizontal cibarial teeth ranged from 8-10 (mean 9.2 + 0.87), predominantly 10 (52.0%), followed by eight (28.0%) and nine (20.0%).
Iporanga (pigsty)
Thirty-four females from this population of N. neivai were analyzed, of which 81.8% had different numbers of rings on the two spermathecae. The number varied from 6-11 (mean of 8.6 + 1.07), with eight (36.9%) and nine (30.8%) rings predominating. The sum of the number of rings on a pair of spermathecae varied from 13-21, with 17 predominating (31.2%). The simple form of the terminal knob of the spermathecae was the most commonest (81.8%), followed by simple/bilobed (15.2%) and bilobed (3.0%). The number of horizontal cibarial teeth in the cibarium was eight (50.0%), nine (20.6%) or 10 (29.4%).
Iporanga (peridomicile)
Thirty-nine females of this population were analyzed and the number of annulations was different between the two spermathecae in 87.5% of the specimens. The number of annulations varied from 7-11 (mean 8.7 + 1.12), with eight (32.1%) or nine (29.5%) predominating. The sum of spermathecal rings varied from 14-22, being predominantly 17 (28.2%) and 15 or 18 (both 12.8%). The simple spermathecal terminal knob occurred most frequently in this population, (84.6%), followed by the simple/bilobed (10.3%) and bilobed (5.1%) forms. The number of horizontal cibarial teeth varied from 8-11 (mean 8.6 + 0.90), predominantly eight (63.9%) followed by nine (13.9%), 10 (19.4%) and 11 (2.8%).
Conchal
Only 18 females from Conchal were examined, 76.5% of them presenting different numbers of rings on their two spermathecae. The number of rings varied from 7-10 (mean 8.5 + 0.99), predominantly eight (35.3%) and nine (29.4%). The sum of the number of rings of the two spermathecae was 14-19, this latter value being the most frequent (29.4%), followed by 15 (23.5%) and 17 (17.7%). The simple form of the spermathecal terminal knob was the most prevalent (88.9%), the remaining specimens (11.1%) having the simple/bilobed type. The bilobed form was not found. The number of horizontal cibarial teeth varied from 8-10 (mean 9.4 + 0.91), the most prevalent being 10 (66.7%), followed by eight (26.7%) and finally, nine (6.7%).
Inter-populational variation
Nyssomyia intermedia
All the N. intermedia populations most commonly had 11 spermathecal rings, except for that of the Lassance chicken house, for which the 12 rings predominated and none of whose specimens had eight. The other Lassance population (cerrado) lacked specimens of 17 spermathecal rings. The Além Paraíba and Iporanga peridomiciliary populations did not include specimens with 16 or17 rings. This latter population had not, however, specimens exhibiting 15 rings (Fig. 5). The analysis of variance for the number of spermathecal rings is shown in Table I. The Lassance chicken house sand flies differed significantly from all the other populations studied, whereas the Iporanga peridomicile, except from this latter, not differed of all other populations.
Prevalence values for the shape the spermathecal terminal knob in the study populations are given in Fig. 6. The simple form predominated in more than half of the specimens of all the Minas Gerais populations. This frequency was approximately double that found in the São Paulo populations. The opposite occurred in populations of this latter state, with a marked predominance of the bilobed form. In the Iporanga population this form appeared almost three times as often as in that of Lassance (cerrado) and was approximately twice as common as in the other populations studied.
The prevalence of the simple/bilobed form (Fig. 6) varied from 19.0 -22.4% in the Minas Gerais and Iporanga (pigsty) populations but was much lower (10.0%) in the others. Of the Minas Gerais populations only the one from Lassance (chicken house) presented the bilobed/trilobed shape, although this morph was present in both those of São Paulo. The two Iporanga populations differed significantly from those of Minas Gerais, except when the peridomiciliary one was compared with that of Além Paraíba. This latter was significantly different from the Lassance (cerrado) population (Table II).
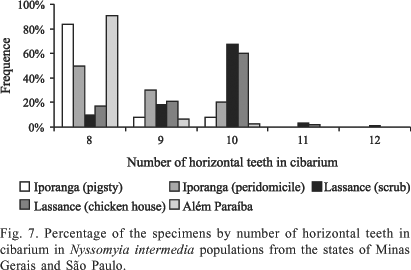
The prevalence of N. intermedia cibaria with eight, nine and ten horizontal teeth are shown in Fig. 7. The number of horizontal cibarial teeth showed considerable variation among the populations. Of the three from Minas Gerais, eight horizontal teeth predominated only in Além Paraíba (90.67%). This form was also commonest in the two São Paulo populations, at 50.0% for Iporanga (peridomicile) and 83.8% for Iporanga (pigsty). The latter frequency was almost nine times higher than that in the Lassance (cerrado) population (9.8%). In both Lassance populations, the greater number of specimens had ten horizontal teeth. Specimens with 11 horizontal teeth were present only in the Lassance populations, both of the chicken house and cerrado, the latter being one presenting specimens with 12 teeth.
The Iporanga (peridomicile) population varied significantly from all the others with regard to the number of teeth in cibarium, the population collected in the pigsty being similar only to that of Além Paraíba. The two Lassance populations did not differ significantly with regard to this character (Table III).
Nyssomyia neivai
The variation in the number of spermathecal rings for N. neivai is shown in Fig. 8. The number of spermathecal rings in this species varied from 6-11 in two of the populations studied, i.e., those of Alfenas and Iporanga (pigsty). There were no specimens with six rings in sand flies from Corinto, Conchal or Iporanga (peridomicile), but the 11-ring form was absent only from the Corinto and Conchal populations.
In all the populations the predominant form had eight rings, the prevalence varying from 32.0-38.5%. The 9-ring morph was next the most frequent, at between 26.0-32.6% in all populations studied (Fig. 8).
The Conchal population did not differ significantly from any other, while that of Alfenas differed from all the rest. No significant variation was seen among the remaining populations with respect to this character (Table IV).
A large majority of the specimens showed the simple form of spermathecal terminal knob (Fig. 9) and there was little variation among the populations, the frequency ranging from 81.8- 94.4%. The three São Paulo populations had frequencies of the simple/bilobed form greater than those of Minas Gerais, the bilobed morph being rare in four of the populations and absent in that of Conchal. There was no significant difference in this among the populations studied (Table V).
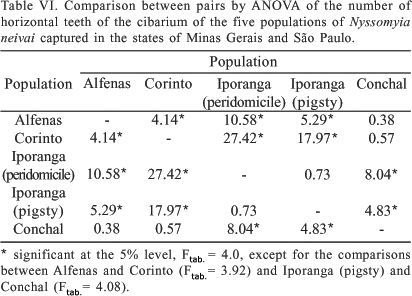
The large variation encountered in the populations studied with respect to the number of horizontal cibarial teeth is shown in Fig. 10. In the two Iporanga populations more than half of the specimens had eight teeth, the morph with 11 teeth occurring only in the peridomiciliary one. In Conchal, Alfenas and Corinto the predominant number of horizontal teeth was ten, present in more than 50% of the specimens of these populations. The highest frequency for nine horizontal teeth was found among the Iporanga (pigsty) and Alfenas populations, at 20.6% and 20.0%, respectively, and the lowest in Corinto, where this morph was represented in only 4.2% of the specimens studied.
With respect to the number of horizontal cibarial teeth, the Alfenas population differed significantly from all the others, except that of Conchal. The Iporanga populations were identical in this character (Table VI).
DISCUSSION
The two species, N. intermedia and N. neivai, are morphologically very close. This fact will undoubtedly lead to further mistakes in their identification and reflect on relative importance attributed to them in the transmission of leishmaniasis.
The variation found for N. intermedia was higher than that found for N. neivai. The existing variation seems to be limited to the two species, indicating the non-existence of a third. According to Marcondes (1997), these two species diverged from each other about 250,000 years ago, as a result of an allopatric speciation.
N. intermedia presents a greater mean number of spermatheca rings than does N. neivai, since, in the latter, the maximum number of rings was 11, which seems to be the upper limit for this species, whereas in the former about 48.2% of the spermathecae analyzed presented a value equal to or higher than 12 rings, 17 being the highest upper limit so far found. The lowest limit of spermatheca rings for N. neivai was six, and 16.4% of the specimens studied had either six or seven rings, while in N. intermedia the minimum value was eight, one less than Marcondes (1996) had found. A great overlap of the values occurs in this structure when the number of rings is between eight and eleven, and in 61.9% of N. intermedia and N. neivai specimens, this is what in fact occurred.
The total number of rings of the two spermathecae varies greatly; this index is greater for N. intermedia. The overlap range is, however, smaller than that of each species alone. The common range, which includes 49.3% of the specimens of the two species, presents values between 17 and 22 rings.
In spite of the value overlap shown above, these data may be useful in distinguishing the two species, as when the extreme values of the total number of rings on the spermathecae are considered, they serve to indicate the difference clearly. Those specimens presenting 13-16 surely belong to N. neivai and those presenting 23-33 belong to N. intermedia.
The simple terminal knob of the spermatheca was dominant in both species, though with quite different values, 45.8% for N. intermedia and 89.3% for N. neivai. When this structure is bilobed, it is most frequently found in N. intermedia (31.8%), being very rare in N. neivai (3.4%), and this aided in the correct identification of this latter species (Andrade Filho et al. 2003).
Marcondes (1996) and Marcondes et al. (1998) used the term "asymmetric" or "truncated/asymmetric", to classify the spermatheca terminal knob. In addition to this term, these authors combined to simple, bilobed and trilobed. Asymmetry has not been considered here, as there are several levels of asymmetry, both for the simple head and for the bilobed one.
Cibarium teeth are still more similar as between these species. Marcondes (1996) found a mean of 10.2 horizontal teeth for N. neivai collected in Bolivia and 8.2 for N. intermedia from Além Paraíba. However, in the populations studied here the mean of the former was 9.1 and of the second 8.9, thus being very close to each other, which indicates that this number is variable according to the population of each species studied, and it is not sufficient to distinguish between the two species.
The asymmetries (or anomalies) in phlebotomines have already also been reported for several other species (Coutinho & Barretto 1940, Sherlock 1958, Ximenes et al. 2002, Andrade Filho et al. 2004). They usually occur in even numbered structures, and the examination of the corresponding region helps in species identification. Nine teeth in the cibarium were found in 14.63% of N. intermedia specimens and in 15.3% of those of N. neivai. Marcondes (1999) considered this number an anomaly; nevertheless, due to the great number of specimens with this number of horizontal teeth, this is more likely to be a naturally existing variable in this species.
According to Futuyma (1993), polymorphism is the existence, within a given population, of two or more genotypes for a particular characteristic, with the greatest frequency rarely exceeding 1%. Phenotypical variation within a population may or may not be genetically based. In this study, a high degree of polymorphism was seen in the three structures studied in all the populations, given that all of them had one value in which the variation exceeded 1%.
There is large genetic and phenotypical variation in several species of phlebotomines, both in the American continents and in the Old World (Lane & Rahman 1980, Azevedo et al. 2000, Martín-Sanchez et al. 2000). In N. intermedia and N. neivai these variations also affect behavior, seeing that although both species occur in forest (Andrade Filho et al. 1998, Domingos et al. 1998) they may also be captured in urban areas, in domestic animal shelters and even indoors (Gomes et al. 1978, Mayrink et al. 1979, Oliveira et al. 1995, Andrade Filho et al. 1997). The polymorphism and morphological variation observed by the present authors as well as others (Marcondes et al. 1998) may contribute to the adaptation of these species to such diverse environments, including the anthropic ones.
Despite the low dispersal capacity of N. neivai (about 70m), it can reach 120m (Casanova 2001) eight nights after release and there appears to be considerable genetic flow between populations, principally between those that are geographically closer. The great polymorphism seen here could only be achieved by genetic crossing between populations. Despite probable genetic flow there was significant variation between some of the populations in our study. The form of the terminal knob of the spermatheca varied considerably as between the Minas Gerais and São Paulo populations, as did the number of horizontal cibarial teeth, the Além Paraíba population being closest to that of São Paulo in this respect.
Of the characters studied, those that presented greatest intra-populational polymorphism were those relating to the spermathecae. This may be because these characters presented the smallest variation among the populations, since they are already expressing their whole range of variation. The morphs with 10-13 annulations in N. intermedia and of that with 7-10 in N. neivai seem to be more frequent.
In addition to the genus Nyssomyia, several other groups of phlebotomines presented spermathecae with annulations, although without this range of variation.
The relationship of each shape of the terminal knob of the spermathecae the face to environmental selective pressures are unknown. Diverse shapes of terminal knob are found in phlebotomines, but only N. intermedia and N. neivai show so great variation, there being no records of other species presenting the bilobed and trilobed forms.
The terminal knob of the spermathecae was highly varied among the populations of N. intermedia, maintaining a more uniform pattern among those of N. neivai. The São Paulo populations of N. intermedia presented the greatest index of bilobed forms, agreeing with data of Marcondes et al. (1998) who found this morph to be predominant in insects of the Southeast and Northeast regions of Brazil. This could not be verified for the Minas Gerais populations, in all of which more than half of the specimens studied had the simple form of the terminal knob.
In all the studied populations of these two species, forms with eight or ten horizontal cibarial teeth predominated. These structures are a continuation of the hypopharynx and the labrum (Forattini 1973) and are involved in the process of blood meal digestion, probably initiating the mechanical breakdown of the cells. It is not known for certain what adaptive advantage would be obtained by a population with ten horizontal teeth over one with eight teeth. It may be that environmental selection pressure, acting through the mammalian hosts, favors a particular number of horizontal teeth, leading this form to predominate in a given population. Other species such as Pintomyia monticola (Costa Lima, 1932) and Psathyromyia shannoni (Dyar, 1929) present variation in the number of horizontal cibarial teeth, although this variation is very rare, not affecting 1% of the specimens (Andrade Filho unpublished results).
Seasonal variation may significantly influence the number of variations. Marcondes et al. (1999) studied material collected during the months of June and December from Venda Nova do Imigrante in the Brazilian state of Espírito Santo, noting small differences in the dimensions. Other populations may thus present different patterns to those observed in this study.
Received 17/05/2005; accepted 31/08/2006
- Andrade Filho, J. D. 2003. Morfologia do Complexo Nyssomyia intermedia (Diptera: Psychodidae: Phlebotominae), Thesis, Universidade de São Paulo, São Paulo, 152 pp.
- Andrade Filho, J. D.; A. P. S. Carneiro; M. L. N. Lima; R. M. Santiago; M. A. Gama; C. A Santos; A. L. Falcão & R. P. Brazil. 1997. Flebotomíneos de Timóteo, Estado de Minas Gerais, Brasil (Diptera: Psychodidae). Cadernos de Saúde Pública 13: 767770.
- Andrade Filho, J. D.; G. M. L. Carvalho; L. Saraiva & A. L. Falcão. 2004. Bilateral anomaly in the style of Micropygomyia schreiberi (Martins, Falcão & Silva) (Diptera: Psychodidae). Revista Brasileira de Entomologia 48: 583585.
- Andrade Filho, J. D.; E. A. B. Galati & A. L. Falcão. 2003. Redescription of Nyssomyia intermedia (Lutz & Neiva, 1912) and Nyssomyia neivai (Pinto, 1926) (Diptera: Psychodidae). Memórias do Instituto Oswaldo Cruz 98: 10591065.
- Andrade Filho, J. D.; E. A. B. Galati & A. L. Falcão. 2004. Biology of the first generation of a laboratory colony of Nyssomyia intermedia (Lutz & Neiva, 1912) and Nyssomyia neivai (Pinto, 1926) (Diptera: Psychodidae). Memórias do Instituto Oswaldo Cruz 99: 597601.
- Andrade Filho, J. D.; M. L. N. Lima; A. L. Falcão & R. P. Brazil. 1998. Sazonalidade dos flebotomíneos (Diptera, Psychodidae) dos arredores da Gruta da Lapinha, município de Lagoa Santa, Minas Gerais, Brasil. Revista Brasileira de Entomologia 42: 9395.
- Azevedo, A. C. R.; F. A. Monteiro; P. H. Cabello; N. A. Souza; M. G. Rosa-Freitas & E. F. Rangel. 2000. Studies on populations of Lutzomyia longipalpis (Lutz & Neiva, 1912) (Diptera: Psychodidae: Phlebotominae) in Brazil. Memórias do Instituto Oswaldo Cruz 95: 305322.
- Casanova, C. 2001. Avaliação do potencial biológico de Lutzomyia intermedia s.l. como vetor da leishmaniose tegumentar americana na zona rural do Município de Conchal, Estado de São Paulo, Thesis, Universidade de São Paulo, São Paulo, 99 pp.
- Casanova, C.; R. C. Mayo; O. Rangel; L. M. Mascarini; M. G. Pignatti; E. A. B. Galati & A. C. Gomes. 1995. Natural Lutzomyia intermedia (Lutz & Neiva) infection in the valley of the Mogi Guaçú River, State of São Paulo, Brazil. Bolletín Direccíon Malariología e Saneamiento Ambiental 35: 7784.
- Coutinho, J. O. & M. P. Barretto. 1940. Contribuição para o conhecimento do flebótomos de São Paulo. III. Descrição do macho de Phlebotomus alphabeticus Fonseca, 1936 e de Phlebotomus pascalei n. sp. (Diptera: Psychodidae). Annais da Faculdade de Medicina da Universidade de São Paulo 16: 193206.
- Domingos, M. F.; G. C. Carreri-Bruno; R. M. C. Ciaravolo; E. A. B. Galati; D. M. V. Wandeley & F. M. A. Corrêa. 1998. Leishmaniose tegumentar americana: flebotomíneos de área de transmissão, no município de Pedro de Toledo, região sul do estado de São Paulo, Brasil. Revista da Sociedade Brasileira de Medicina Tropical 31: 425432.
- Forattini, O. P. 1973. Entomologia Médica, São Paulo, Blücher Ltda., 658 pp.
- Forattini, O. P.; D. B. G. Pattoli; E. X. Rebello & A. O. Ferreira 1972. Infecção natural de flebotomíneos em foco enzoótico de leishmaniose no estado de São Paulo, Brasil. Revista de Saúde Pública 6: 431433.
- Gomes, A. C.; E. X. Rabello & E. A. B. Galati. 1978. Flebotomíneos encontrados em galinheiros experimentais nos estado de São Paulo e Minas Gerais (Brasil) e algumas observações ecológicas. Revista de Saúde Pública 12: 403407.
- Lane, R. P. & S. J. Rahman. 1980. Variation in the ascoids of the sandfly Phlebotomus argentipes in a population from Patna, Northern India. Journal Communicable Diseases 12: 216218.
- Marcondes, C. B. 1996. A redescription of Lutzomyia (Nyssomyia) intermedia (Lutz & Neiva, 1912), and resurrection of L neivai (Pinto, 1926) (Diptera, Psychodidae, Phlebotominae).Memórias do Instituto Oswaldo Cruz 91: 457462.
- Marcondes, C. B. 1997. Morfometria e DNA mitocondrial de populações sul americanas de Lutzomyia (Nyssomyia) intermedia (Lutz & Neiva, 1912) (Diptera, Psychodidae, Phlebotominae), Thesis, Universidade Federal do Paraná, Curitiba, 260pp.
- Marcondes, C. B. 1999. Anomalies of Lutzomyia intermedia (Lutz & Neiva, 1912) (Diptera, Psychodidae, Phlebotominae). Memórias do Instituto Oswaldo Cruz 94: 365366.
- Marcondes, C. B.; A. L. Lozovei; A. Falqueto; R. P. Brazil; E. A. B. Galati; G. M. Aguiar & N. A. Souza. 1999. Influence of altitude, latitude and season of collection (Bergmann's Rule) on the dimension of Lutzomyia intermedia (Lutz & Neivai, 1912) (Diptera, Psychodidae, Phlebotominae). Memórias do Instituto Oswaldo Cruz 94: 693700.
- Marcondes, C. B.; A. L. Lozovei & E. A. B. Galati. 1998. Variações regionais e interespecíficas na morfologia de insetos do complexo Lutzomyia intermedia (Diptera, Psychodidae, Phlebotominae). Revista de Saúde Pública 32: 519525.
- Martín-Sanchez, J.; M. Gramiccia; B. Pesson & F. Morillas-Marquez. 2000. Genetic polymorphism in sympatric of the genus Phlebotomus, with special reference to Phlebotomus perniciosus and Phlebotomus longicuspis (Diptera, Phlebotominae). Parasite 7: 247254.
- Mayrink, W.; P. Williams; M. V. Coelho; M. Dias; A. V. Martins; P. A. Magalhães; C. A. Costa; A. R. Falcão; M. N. Melo & A. L. Falcão. 1979. Epidemiology of dermal leishmaniasis in the Rio Doce Valley, State of Minas Gerais, Brazil. Annals Tropical Medicine and Parasitology 73: 123137.
- Oliveira, S. M. P.; R. C. H. Afonso; C. M. G. Dias & R. P. Brazil. 1995. Estudo da fauna de flebotomíneos (Diptera: Psychodidae) em Santa Cruz, município do Rio de Janeiro, Brasil. Revista Brasileira de Entomologia 39: 547551.
- Rangel, E. F.; N. A. Souza; E. D. Wermelinger & A. F. Barbosa. 1984. Infecção natural de Lutzomyia intermedia Lutz & Neiva, 1912 em área endêmica de leishmaniose tegumentar no estado do Rio de Janeiro. Memórias do Instituto Oswaldo Cruz 79: 395396.
- Sherlock, I. A. 1958. Anomalias de "Phlebotomus longipalpis" Lutz & Neiva, 1912 (Diptera, Psychodidae). Revista Brasileira de Biologia 18: 433437.
- Ximenes, M. F. F. M.; E. G. C. Bermúdez & R. A. Freitas. 2002. Morphological genitalic anomaly in Lutzomyia longipalpis sensu lato (Lutz & Neivai, 1912) collected in the State of Rio Grande do Norte, Brazil. Entomotropica 17: 183184.
Publication Dates
-
Publication in this collection
27 Oct 2006 -
Date of issue
Sept 2006
History
-
Accepted
31 Aug 2006 -
Received
17 May 2005