Abstracts
Mycetarotes is a small genus of the exclusively Neotropical fungus-growing ants, that includes M. parallelus (Emery), M. senticosus Kempf, M. acutus Mayhé-Nunes and M. carinatus Mayhé-Nunes. We hereby revise historical and recent information regarding Mycetarotes species for the first time, providing an identification key to workers, diagnoses, synoptic illustrated redescriptions of the species, including those of sexuals when known, updates of distributional records, and nest pictures of M. carinatus and M. parallelus. We comment the taxonomy and phylogenetic relationships among Mycetarotes and related genera, and on their geographical distribution. The available biological information on the genus is summarized.
Attini; biology; Myrmicinae; phylogeny; taxonomy
O gênero Mycetarotes inclui quatro espécies de formigas cultivadoras de fungos, exclusivamente encontradas na região Neotropical: M. parallelus (Emery), M. senticosus Kempf, M. acutus Mayhé-Nunes and M. carinatus Mayhé-Nunes. Apresentamos pela primeira vez um resumo sobre as informações disponíveis para as espécies do gênero, fornecendo uma chave para a identificação das operárias, diagnoses e redescrições sinópticas ilustradas das espécies, incluindo as dos alados, quando conhecidos, atualização dos registros de distribuição e fotografias dos ninhos de M. carinatus e M. parallelus. Comentamos a taxonomia e as relações de parentesco entre Mycetarotes e gêneros cognatos, e sobre sua distribuição geográfica. As informações biológicas disponíveis sobre o gênero também foram resumidas.
Attini; biologia; filogenia; Myrmicinae; taxonomia
SYSTEMATICS, MORPHOLOGY AND BIOGEOGRAPHY
Revisionary notes on the fungus-growing ant genus Mycetarotes Emery (Hymenoptera, Formicidae)
Notas sobre as formigas cultivadoras de fungos do gênero Mycetarotes Emery (Hymenoptera, Formicidae)
Antonio José Mayhé-NunesI; Carlos Roberto Ferreira BrandãoII
IDepartamento de Biologia Animal, Instituto de Biologia, Universidade Federal Rural do Rio de Janeiro, 23890-000 Seropédica-RJ, Brazil. Bolsista do CNPq. amayhe@ufrrj.br
IIMuseu de Zoologia da Universidade de São Paulo, Av. Nazaré 481, 04263-000 São Paulo-SP, Brazil. Bolsista do CNPq. crfbrand@usp.br
ABSTRACT
Mycetarotes is a small genus of the exclusively Neotropical fungus-growing ants, that includes M. parallelus (Emery), M. senticosus Kempf, M. acutus Mayhé-Nunes and M. carinatus Mayhé-Nunes. We hereby revise historical and recent information regarding Mycetarotes species for the first time, providing an identification key to workers, diagnoses, synoptic illustrated redescriptions of the species, including those of sexuals when known, updates of distributional records, and nest pictures of M. carinatus and M. parallelus. We comment the taxonomy and phylogenetic relationships among Mycetarotes and related genera, and on their geographical distribution. The available biological information on the genus is summarized.
Keywords: Attini; biology; Myrmicinae; phylogeny; taxonomy.
RESUMO
O gênero Mycetarotes inclui quatro espécies de formigas cultivadoras de fungos, exclusivamente encontradas na região Neotropical: M. parallelus (Emery), M. senticosus Kempf, M. acutus Mayhé-Nunes and M. carinatus Mayhé-Nunes. Apresentamos pela primeira vez um resumo sobre as informações disponíveis para as espécies do gênero, fornecendo uma chave para a identificação das operárias, diagnoses e redescrições sinópticas ilustradas das espécies, incluindo as dos alados, quando conhecidos, atualização dos registros de distribuição e fotografias dos ninhos de M. carinatus e M. parallelus. Comentamos a taxonomia e as relações de parentesco entre Mycetarotes e gêneros cognatos, e sobre sua distribuição geográfica. As informações biológicas disponíveis sobre o gênero também foram resumidas.
Palavras-chave: Attini; biologia; filogenia; Myrmicinae; taxonomia.
Mycetarotes is a small genus of the exclusively neotropical fungus growing ants, part of the so-called "lower attines", which study has been traditionally neglected in comparison with that of the "higher" Attini, the leaf-cutters Acromyrmex and Atta. However, a recent and strong impetus in the study of the basal Attini by different authors, may soon produce a sound history of estimated 50 million years of the positive relations between these ants and a particular cultivar of basidiomycete fungus, on which they and their larvae rely entirely for nourishment. The study of the basal attines is instrumental to the construction of this hypothesis.
The taxonomic history of Mycetarotes began when Emery (1905) described Cyphomyrmex parallelus, later transferred to Mycetarotes, then proposed as a subgenus (Emery 1913). Forel (1913) questioned the validity of Mycetarotes, but the name was nevertheless raised to generic rank by Borgmeier (1950). In the first Mycetarotes revision, Kempf (1960) further supported Borgmeier's proposal on the basis of studies on M. parallelus males and M. senticosus workers, and recognized two species in the genus. Two other species, M. acutus and M. carinatus, were later proposed by Mayhé-Nunes (1995). Lanziotti & Mayhé-Nunes (2001) redescribed the female of M. parallelus, and Mayhé-Nunes & Lanziotti (2004) first described gynes and males of M. carinatus. However, more than a century after the original description of the genus, the gynes and males of M. senticosus and M. acutus remain unknown.
We present, for the first time, illustrated synoptic descriptions of workers and those of known sexuals of the four known species, updates of distributional records, a summary of what is known on the biology of the species, and report and illustrate new observations, in special on nest architecture of M. carinatus and M. parallelus. We also comment on the taxonomic status and phylogenetic relationships among Mycetarotes and cognated genera.
Somolon et al. (2004) recently suggested that Mycetarotes is an ideal model taxon in South America for the study of coevolution of lower-attine ants and their cultivated fungi, paralleling the role of the Cyphomyrmex longiscapus species group in Central America (Schultz et al. 2002). Considering that attine diversity is highest in South America, where ant-fungus symbiosis is thought to have originated, Somolon et al. (2004) defended that adopting a model taxon could facilitate attine research regionally, stating that "investigations into the origin of this symbiosis should therefore focus on ants and fungi from South America." According to Somolon et al. (2004), Mycetarotes show five important model characteristics: i) some of its species may be locally abundant, and excavating their nests is relatively simple in comparison with other attines, ii) the species are easy to keep in laboratory conditions, iii) they show a basal position in the attine phylogeny, being thus more likely to have retained features of the attine ancestor, iv) different Mycetarotes species cultivate distinct fungal cultivars, suggesting that speciation in this ant genus may coincide with switches to novel cultivars (Mueller et al. 1998; Schultz et al. 2002); v) the cultivar of M. parallelus is capable of producing fruiting-body development, indicating that this cultivar may have close links to free-living, non symbiotic fungal populations.
Somolon et al. (2004) reported the first detailed study on nest architecture and colony size for two Mycetarotes species and integrated these findings with the scant available biological information published to date.
Mycetarotes may be locally abundant and always live in rather simple nests with one entrance leading to a superficial (5-18 cm below ground) single chamber, containing a unique fungus garden, one mated gyne, immatures and up to 100 or so workers. Sometimes it is possible to observe a small chamber at mid height, possibly a vestige of the founding chamber. Excavating nests and rearing Mycetarotes colonies have been considered relatively easy in comparison with the efforts required to excavate multiple chambered nests; also it has been considered relatively easy to keep live mature Mycetarotes colonies in the laboratory, in comparison with the larger populations of the other so callled "lower" attine genera. These qualities led Solomon et al (2004) to suggest that Mycetarotes, particularly M. parallelus, could be used as a model system in studies aiming to improve our understanding of the origin and history of the symbiosis between fungi and ants.
MATERIAL AND METHODS
The terminology here adopted follows Kempf (1960), partially modified by Mayhé-Nunes and Brandão (2005). Abbreviations for measurements and ratios are defined as follow: TL, total length; HL, head length (except mandibles); HW, head width (including eyes); IFW, inter frontal width (distance between the lateral margins of frontal lobes); ScL, scape length; WL, Weber's length (= alitrunk length); HfL, hind femora length; LFw, lenght of fore wing; LHw, lenght of hind wing; CI, cephalic index (HW/HL x 100); FLI, frontal lobes index, to compare among species the lateral expansion of the frontal lobes (IFW/HW x 100).
Nests were excavated using routine procedures as described by Schultz (1993). Other samples examined are deposited in the following institutions (curators between parentheses):
CECL - Coleção Entomológica Angelo Moreira da Costa Lima. Instituto de Biologia, Universidade Federal Rural do Rio de Janeiro, Seropédica, RJ, Brazil (Francisco Racca Filho)
INPA - Instituto Nacional de Pesquisas da Amazônia, Manaus, AM, Brazil (Célio U. Magalhães Filho)
MPEG - Museu Paraense Emilio Goeldi, Belém, PA, Brazil (Alexandre B. Bonaldo)
MZSP - Museu de Zoologia da Universidade de São Paulo, São Paulo, SP, Brazil (Carlos Roberto F. Brandão)
RESULTS
Mycetarotes Emery, 1913
Cyphomyrmex (in part): Emery, 1905: 161.
Atta (Mycocepurus) (in part): Forel, 1911: 293.
Cyphomyrmex (Mycetarotes) Emery, 1913: 251 (type species Cyphomyrmex parallelus, by original designation). Emery, 1922: 342-343.
Mycetarotes: Borgmeier, 1950: 384 (raised to genus). Kempf, 1960: 277 (revision; key). Kempf, 1972: 144 (catalog). Mayhé-Nunes, 1995: 197 (synopsis; key). Bolton, 1995: 36; 268 (catalog). Bolton, 2003: 199 (taxonomic history).
Diagnosis (workers): Monomorphic attine ants, 2.6-4.0 mm total length. Mandibles finely and longitudinally striolate, with 5-6 teeth on masticatory margin. Anterior margin of clypeus convex. Frontal area impressed. Cephalic capsule slightly longer than broad to as long as broad (CI 90-101). Lateral margins of frontal lobes little expanded laterally (FLI 35-47) and bluntly angulated or rounded. Frontal carinae subparallel and diverging caudad into two branches at level of vertex (weakly in M. parallelus) little before reaching the occipital margin; ending at some distance from the base of small spines variably prominent on the occipital margin. Preocular carinae curving towards the head's middle at the compound eyes level. Eyes convex in frontal view, surpassing notably the lateral margins of head, set near the middle of the lateral sides. A clearly delimited antennal scrobe absent. Antennal scapes, when laid backwards as much as possible, notably surpassing the occipital margin. Funiculus without a noticeable club. With the head seen in frontal view, occipital spines projected between the outer spines of occipital margin and supraocular spines, giving a tridentate appearance to the occipital corners. Dorsal surface of pronotum with four spines, in a transversal row; median spines largely spaced; pronotal inferior corners with a triangular tooth on each side; humeral angles commonly with a small tubercle on each side of pronotal dorsum, near its lateral border (sometimes absent in M. parallelus); first and second pairs of mesonotal spines always present. Petiole longer than broad, in dorsal view; petiolar dorsum with a pair of spines near the posterior border. Tergum I of gaster laterally marginate. Gaster longer than broad in dorsal view. Long hairs and pubescence scarce; short hairs distributed on tergum I of gaster and outer margin of mandibles, rarely on mesosoma, somewhat thicker and curved on legs and border of frontal lobes than in other body parts; decumbent hairs on scape and clypeus, also present and longer on hipostomal region than in other body parts, clypeal anterior border and masticatory margin of mandibles; pubescence confined to antennal funiculus.
Key for species of Mycetarotes (workers)
Tergum I of gaster with only two longitudinal ridges on its anterior two thirds, sometimes inconspicuous; promesonotum with six or seven projections, their apices varying from acute to blunt; propodeal spines as long as the other high mesosomal projections ........................................................................ 2
Frontal carinae above the frontal lobes subparallel; outer stem of frontal carinae weakly marked; mesosomal projections blunt; preocular carinae not reaching the frontal ones ............................................................................... 3
Mesonotum with three pairs of spines, the third mesonotal conspicuously projected; anterior margin of clypeus with a medial notch ............................................................................ M. senticosus Kempf
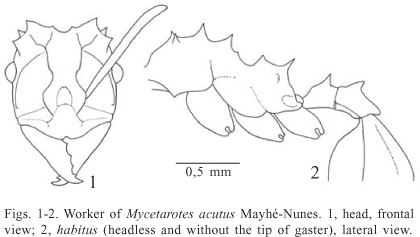
Mycetarotes acutus Mayhé-Nunes, 1995
Mycetarotes acutus Mayhé-Nunes, 1995: 198-200 (worker, Manaus, Amazonas, Brazil).
Mycetarotes acutus: Solomon et al., 2004: 334-335 (nest)
Type material: Holotype. BRAZIL: Amazonas State, Manaus, março 1990, E. F. Morato col. (holotype MZSP) (examined).
Diagnosis: Projections over the body notably slenderer and sharper than in other species of the genus. Preocular carinae almost touching the frontal ones, the latter constricted above frontal lobes. Postpetiolar dorsum not impressed posteriorly.
Description: Worker. Average of measurements in mm (n = 4): TL 3.4 (3.1-3.6); HL 0.90 (0.86-0.94); HW 0.87 (0.83-0.92); IFW 0.31 (0.29-0.34); ScL 0.97 (0.89-1.07); WL 1.23 (1.17-1.36); HfL 1.15 (1.12-1.17). Reddish-brown to yellowish-brown. Integument opaque, fine and indistinctly punctuated.
Head. Cephalic capsule slightly longer than broad to almost as long as broad; CI average 97 (94-99). Chewing border of mandible with 5 teeth. Median anterior border of clypeus with a small notch; median apron anteriorly with an inconspicuous denticle between the anterior insertion of frontal lobes and its anterior margin. Frontal lobes nearly round, with approximate lateral margins; FLI average 37 (35-39). Frontal carinae constricted above the frontal lobes; preocular carinae almost touching the frontal carinae. Supraocular projections acute and tubercle-like. Occipital margin medially round in full-face view, notched between the inner spines, and obliquely straight between this spines and the outer ones. All funicular segments longer than broad.
Mesosoma. With the pronotum in frontal view, the midpronotal spines are slightly shorter than the lateral ones. Mesonotal dorsum with three pairs of projections; the first two spines longer than the other mesosomal projections; third pair very small. Basal face of propodeum with two longitudinal ridges, mesially projected as conspicuous acute spines; propodeal spines slender, obliquely directed outwards.
Waist and gaster. Petiolar dorsum with a pair of spines directed upwards, separated from posterior border by a distance nearly equal to the spines length. Postpetiole with a tooth on each side of the lateral anterior margin, posterior margin notched; dorsal surface flat, without median impression. Tergum I of gaster longitudinally marginated by a low ridge on each side; longitudinal ridges on disc absent.
Gyne and Male. Unknown.
Comments: This species resembles M. senticosus in the arrangement of the mesosomal projections, the notched anterior margin of clypeus, and the shape of the petiolar spines. This similarity probably led to the identification of a sample of this species collected in Manaus as M. senticosus by Mueller et al. (1998), a move later rectified by Solomon et al. (2004). It differs from M. senticosus by having sharper body ridges and projections, a smaller third pair of mesonotal projections, by the shape of occipital margin, the constricted frontal carinae, longer preocular carinae, and posteriorly notched postpetiole, without median impression.
Biology: Solomon et al. (2004) excavated five nests near Manaus, Brazil, all with similar architecture, agreeing with recent observations in Melgaço, Pará by C. E. D. Sanhudo (pers. comm.). According to Solomon et al. (2004), the single entrance is surrounded by a turret made by excavated sand or clay pellets, with average height and width of 2.3-1.3 cm, respectively. The nests showed a single chamber, on average placed 11.6 cm (S.D. 6.1; n = 5) of depth bellow ground, measuring in width of 5.1 cm (S.D. 1.1; n = 5) and 5.0 cm in height (S.D. 2.0; n = 3), with the fungus gardens suspended from the roof by rootlets. The estimated population of one nest was 20-30 workers and the foraging substrate consisted of seeds, flower stems, and unidentified plant fibers.
Material examined: BRAZIL: Amazonas State, Manaus, Rs 1302, 12.xi.1993. A. B. Casimiro col. one worker (CECL). Pará State, Caxiuanã, ECF Penna, 24.iv.2004, C.E.D. Sanhudo col. 2, 4 and 5 workers (CECL, INPA, MPEG).
Mycetarotes carinatus Mayhé-Nunes, 1995
(Figs. 3-6)

Mycetarotes carinatus Mayhé-Nunes, 1995:200-202 (worker, Engenheiro Paulo de Frontin, Rio de Janeiro, Brazil; nest).
Mycetarotes carinatus: Mayhé-Nunes & Lanziotti, 2004:110 (female and male; nest).
Type material: Holotype and six paratype workers in MZSP; seven paratype workers in CECL (examined). BRAZIL: Rio de Janeiro State, Engenheiro Paulo de Frontin, 29.xii.1993, A. Mayhé col. (holotype and paratypes).
Diagnosis: Tergum I of gaster with four longitudinal ridges. Mesosomal projections stouter than in other species of the genus, bluntly pointed.
Description: Worker. Average of measurements in mm (n = 9): TL 2.8 (2.6-2.9); HL 0.80 (0.75-0.83); HW 0.77 (0.74-0.78); IFW 0.33 (0.32-0.37); ScL 0.76 (0.68-0.80); WL 1.02 (0.92-1.11); HfL 0.93 (0.89-0.98). Light brown to yellowish brown, with head slightly darker. Integument opaque, fine and indistinctly punctuated; gaster microscopically reticulated laterally in some specimens.
Head. Cephalic capsule slightly longer than broad to as long as broad; CI average 97 (94-100). Chewing border of mandible with 5 teeth, the basal ones small to indistinct. Median anterior border of clypeus without a small notch; median apron anteriorly with an inconspicuous tubercle between the anterior insertion of frontal lobes and its anterior margin. Frontal lobes subtriangular, with approximate to moderately approximate lateral margins; FLI average 43 (40-47). Frontal carinae without constriction above the frontal lobes, diverging caudad in two branches above eyes, reaching the base of the spines on occipital margin; preocular carinae curved inwards bellow eyes, falind out near the posterior orbit. Supraocular projections spine-shaped. Occipital margin in full-face view concave, notched between the inner spines, and shallowly concave between these spines and the outer ones. Only the five apical funicular segments longer than broad, I-V almost as long as broad.
Mesosoma. With the pronotum in frontal view, midpronotal spines almost as long as lateral ones. Mesonotal dorsum with three pairs of projections; the first two spines longer than the other mesosomal projections; third pair very small. Basal face of propodeum with two longitudinal ridges, medially projected as conspicuous spines; propodeal spines slender, obliquely and outwards directed.
Waist and gaster. Petiolar dorsum with a pair of spines directed upwards, placed away of posterior border, in a distance similar to length of spines. Postpetiole with small tubercle near each side of lateral anterior margin; posterior margin smooth and concave; dorsal surface deeply impressed. Tergum I of gaster longitudinally marginated by low ridge on each side, with two additional longitudinal ridges on disc.
Gyne (measurements in mm): TL 3.1; HL 0.85; HW 0.88; IFW 0.41; ScL 0.74; WL 1.11; LFw 3.29; LHw 2.45. Color, integument, pilosity, head, waist and gaster resembling those of conspecific workers, but differing as follow: cephalic capsule as long as broad (CI 103); interfrontal width a little larger than in other species in the genus (FLI 47); three ocelli on vertex bordered by short and curved carinae. Petiole as long as broad. Dorsum of the gaster's basal portion with a narrow transversal furrow, matching with posterior margin of postpetiole.
Mesosoma. Pronotum with a pair of strong and blunt humeral spines, obliquely directed, and with inferior spines pointed forwards and downwards, similar to the humeral ones; pair of midpronotal spines absent. Scutum without conspicuous divisions in dorsal view; anterior margin shallowly notched in the middle, and with a short and narrow furrow on each side; median posterior margin superficially impressed; parapsidial furrow visible in dorsal view, parapsis weakly excavate, with a low ridge delimiting the lateral free margin. Mesothoracic paraptera deeply impressed in the middle, with rounded lateral free margin. Scutellum with a pair of longitudinal blunt ridges, finishing posteriorly as short and blunt spines. Metathoracic paraptera concealed by scutellum in dorsal view. Propodeum with rounded lateral margins in dorsal view and with a pair of small tubercles between basal and declivous faces.
Wings. Microscopically hairy and infuscated. Fore wing with four closed cells (sub-marginal, costal, median and sub-median); marginal cell open. Pterostigma conspicuous. Hind wing with five veins, without closed cells; 6 hamuli on anterior margin.
Male (measurements in mm): TL 3.9; HL 0.65; HW 0.89; IFW 0.21; ScL 0.62; WL 1.38; LFw 2.98; LHw 2.07. Dark brown to black; inconspicuous and appressed minute hairs; integument finely reticulated, with rugulae on some parts of head and mesosoma.
Head. Cephalic capsule broader than long (CI 137). Mandible striated on dorsal surface and fully punctuated; masticatory margin with only 2 apical teeth; external margin straight with the apex curved inwards, with head in frontal view. Median margin of clypeus moderately convex, without anterior notch; surface of clypeus with two minute lateral tubercles between anterior margin and base of frontal lobe, followed medially above by wide and tranversally vaulted furrow. Frontal lobes aproximate (FLI 24), laterally rounded and dorsally pointing forwards. Frontal carina short in relation to that of other species in the genus, rather straight, obliquely directed inward. Preocular carinae curved above eye, not attaining the vertex. Very large and convex compound eyes, filling about 2/3 of the head's side. Three occelli on vertex, inserted into short and cylindrical projections. Antennae with 13 segments; scape surpassing the occipital margin by nearly 1/4 of its length, when laid backward as much as possible, longer than funicular segments I-III combined; funicular segment I slightly shorter than II. Occipital corner angulated with a single tooth on each side. Occipital margin notched in the middle, between the lateral ocelli.
Mesosoma. Pronotum with small humeral spine and a tumuliform projection close to the rounded inferior corner. Scutum with wide and deeply impressed Mayrian furrows separating three areas: an anterior prescutal and two lateral mesoscutal. Prescutum a little lower than mesoscutum in lateral view; parapsidial furrows on mesoscutum visible in dorsal view; parapsis deeply excavated, with lateral free margin delimited by a high ridge. Mesothoracic paraptera narrowed and deeply impressed medially in dorsal view, with thick and blunt projection on each side. Scutellum surmount by a pair of longitudinal blunt ridges, ending as two short, slim and blunt spines. Metathoracic paraptera concealed by the scutellum in dorsal view. Lateral margin of propodeum angulated in dorsal view, with a pair of small projections between basal and declivous faces.
Waist and gaster. Dorsum of petiole with a pair of blunt tubercles. In dorsal view, postpetiole very little broader than the petiole, shallowly impressed dorsally, with posterior margin notched. First gastric tergite with a longitudinal median furrow flanked by two inconspicuous ridges, lower than lateral ones.
Wings. Microscopically hairy and infuscated. Fore wing with three closed cells (marginal, sub-marginal and costal); median and sub-median cells partially open. Pterostigma conspicuous. Hind wing with five veins, without closed cells; 6 hamuli on anterior margin.
Comments: Mycetarotes carinatus is easily recognized by its blunt mesosomal projections and because it is the only species in the genus that has four longitudinal carinae on tergum I of the gaster. Among other Attini, however, some species of Cyphomyrmex and Trachymyrmex also present four longitudinal ridges on gaster, suggesting a homoplastic condition of the character state.
Biology: The nest excavated in the type locality (Fig. 6) by one of us (AJMN) was found close to a pasture in the glade of a secondary forest border (Mayhé-Nunes 1995). The narrow entrance was 3 mm in diameter and hidden under a partially rotten leaf, without any accumulated earth grains surronding ring. The roof of the single chamber was 9 cm bellow the ground surface, 7 cm wide and 4 cm in height; the pending gray-yellowish fungus garden had a volume of approximately 50 ml. Since the gyne was not found, it seems likely that the nest had an additional chamber not found, hampering accurate population count. During the nest excavation the workers were very active, but stray workers feigned death when disturbed. The chambers of the three nests observed in Minas Gerais were found in shallow depth, in clay soil covered by a secondary forest at Universidade Federal de Viçosa campus, in April 1996 (José Henrique Schoereder pers. comm.). One nest was transferred to the laboratory and produced gynes and males after six months. Two additional nests found in the same area had the entrance surrounded by a turret, the biggest one with 4 cm of height and 6 cm of diameter at the base (Fig. 5). In a third nest without an entrance mound, the workers were following two opposite trails to dispose earth grains and old fungal substrate almost 15 cm away from the nest entrance. A spider exuvia carried by a worker was the only foraging item observed in the field, but substrate discarded in the laboratory was similar to that observed for M. parallelus: seeds, decayed wood and insect feces. The observations in Minas Gerais and an additional worker captured in a primary forest (Reserva Biológica do Tinguá, RJ, Brazil) from litter sample submitted to Winkler extractor, revealed that M. carinatus can also live inside forests.
Material examined: BRAZIL: Minas Gerais State, Viçosa, "Mata da Biologia", [Universidade Federal de Viçosa campus], 2.iv.1996, J.H. Schoereder col. ~ 700 m alt. 4 workers, 5 gynes, 10 males (CECL), 2 gynes, 2 males (MZSP); ibidem, 15.v.1996, A. Mayhé col. 4 workers (CECL). Rio de Janeiro State, Nova Iguaçu, Reserva Biológica do Tinguá, 04.ii.2002, S.V. Ferreira col. Winkler sacks one worker (MZSP). São Paulo State, Botucatu, 11.iii.1988, L. C. Forti & Rinaldi, I. M. P. col. one worker (MZSP).
Mycetarotes parallelus (Emery, 1905)
(Figs. 711)

Cyphomyrmex parallelus Emery, 1905: 162-163 (worker, female, Coxipó, Mato Grosso, Brazil).
Atta (Mycocepurus) luederwaldti Forel, 1911: 293-294; Luederwaldti 1918: 68 (nest). Mycetarotes parallelus: Kempf, 1960: 279-282 (male, key, distribution, synonymy, biology); Kempf, 1972: 145 (catalog); Mayhé-Nunes, 1995: 202-203 (key, biology, nest); Schultz & Meier, 1995: 357 (larva); Bolton, 1995: 268 (catalog); Lanziotti & Mayhé-Nunes, 2001: 176 (female); Solomon et al., 2004: 334 (nest).
Type material: Worker and female in Emery´s collection at Museo Civico di Storia Naturale, Genoa, Italy (R. Poggi pers. comm.; not seen).
Diagnosis: Four spines on the mesonotum. Well developed petiolar spines, inserted very close to posterior border. Outer branches of frontal carinae vestigial.
Description: Worker. Average of measurements in mm (n = 8): TL 3.0 (2.7-3.2); HL 0.83 (0.74-0.91); HW 0.79 (0.72-0.88); IFW 0.33 (0.32-0.34); ScL 0.79 (0.74-0.88); WL 1.05 (0.94-1.12); HfL 1.00 (0.95-1.09). Reddish-brown to yellowish, with head and gaster darker in some specimens. Integument opaque, microscopically reticulated.
Head. Cephalic capsule slightly longer than broad to as long as broad; CI average 95 (90-101). Chewing border of mandible with 6 teeth, the basal ones small to indistinct. Median anterior border of clypeus without a small notch; Frontal lobes subtriangular, with approximate lateral margins; FLI average 42 (39-44). Frontal carinae without constriction above the frontal lobes, posteriorly split into two branches above eyes, the outer ones vestigial and the inner weakly marked in the base, reappearing in the vertex and almost reaching the occipital margin; preocular carinae curved inwards bellow eyes, fading out near the posterior orbit. Supraocular projections spine-shaped. Occipital margin in full-face view medially round, notched between the inner spines, and shallowly concave between these spines and the outer ones. Only the five apical funicular segments longer than broad, I-V almost as long as broad.
Mesosoma. With pronotum in frontal view, midpronotal spines shorter than lateral ones. Mesonotal dorsum with two pairs of projections, the first shorter than the second. Basal face of propodeum with two longitudinal ridges, medially projected as low and blunt projections; propodeal spines slender, directed obliquely backwards.
Waist and gaster. Petiolar dorsum with a pair of spines directed upwards, placed very close to the posterior margin. Postpetiole without a tooth near each side of the lateral anterior margin; posterior margin smoothly concave; dorsal surface with a deep circular impression. Tergum I of gaster longitudinaly marginated by low ridges; longitudinal ridges on disc absent.
Gyne (measurements in mm): TL 4.7; HL 1.13; HW 1.03; IFW 0.35; ScL 0.94; WL 1.35; LFw 3.80; LHw 2.83. Color, integument, pilosity, head, waist and gaster similar to those of the workers, differing as follow: cephalic capsule little longer than broad (CI 91); smaller interfrontal width than in conspecific workers (FLI 34); three ocelli on vertex bordered by short and curved carinae; petiole as long as broad.
Mesosoma. Pronotum with a pair of strong and acute humeral spines, obliquely directed forwards, and with the inferior spines down and forwards pointed, similar to the humeral ones; pair of midpronotal spines absent. Scutum without conspicuous divisions in dorsal view; anterior margin without a shallow median notch in the middle, but with a short and very narrow furrow on each side; parapsidial furrow inconspicuous in dorsal view, parapsis weakly excavate, with a low ridge limiting the lateral free margin. Mesothoracic paraptera deeply impressed in the middle, with lateral free margin notched medially in dorsal view. Scutellum with a pair of strong dorsal low tubercles, finishing posteriorly in a pair of acute spines. Metathoracic paraptera concealed by the scutellum in dorsal view. Propodeum with bluntly angulated lateral margins in dorsal view; between the basal and declivous faces with a pair of slim acute spines, directed obliquely outwards.
Wings. Microscopically hairy and infuscated. Fore wing with four closed cells (sub-marginal, costal, median and sub-median); marginal cell open. Pterostigma conspicuous. Hind wing with four veins, without closed cells; 6 hamuli on anterior margin.
Male (measurements in mm): TL 3.7; HL 0.65; HW 0.80; IFW 0.25; ScL 0.31; WL 1.16; LFw 2.86; LHw 2.07 Fuscous reddish-brown, head more infuscated than the rest of the body; integument finely reticulated; inconspicuous and appressed minute hairs.
Head. Cephalic capsule a little longer than broad (CI 123). Mandible faintly striated and fully punctuated on the dorsal surface; masticatory margin vestigialy denticulate, with 2 distinct apical teeth; external margin straight with the apex curved inwards, with the head in frontal view. Median margin of clypeus moderately convex, without anterior notch; surface of clypeus without projections between anterior margin and base of frontal lobe. Frontal lobes aproximate (FLI 31), laterally rounded and pointing forwardsdorsally . Frontal carina short, rather straight, obliquely directed inwards. Preocular carinae curved above eye, not attaining the vertex. Very large and convex compound eye, covering some 2/3 of the head's side. Three occelli on vertex, set on prominent tubercles. Antennae with 13 segmets; scape not surpassing the occipital margin, when laid backward as much as possible, shorter than funicular segments I-III combined; funicular segment I distinctly shorter than II. Occipital corner tridentate. Occipital margin shallowly notched in the middle, between the lateral ocelli.
Mesosoma. Pronotum with a strong humeral spine and a similar projection close to the inferior corner. Scutum with narrow and deeply impressed Mayrian furrows separating three areas: an anterior prescutal and two lateral mesoscutal. Prescutum with a shallow median longitudinal impression, laterally marginate above, before sloping down untill the sagital Mayrian furrow; parapsidial furrows on mesoscutum visible in dorsal view; parapsis deeply excavated, with lateral free margin limited by a high ridge. Mesothoracic paraptera deeply impressed, medially bisected by a sharp and short longitudinal carina, with notched free lateral margin. Scutellum with a pair of stout, raised, longitudinally carinate tuberosities, ending posteriorly in a pair of strong spines. Metathoracic paraptera concealed by the scutellum in dorsal view. Propodeum with bluntly angulated lateral margins in dorsal view; between the basal and declivous faces with a pair of strong acute spines, directed obliquely outwards.
Wings. Microscopically hairy and infuscated. Fore wing with five closed cells (marginal, sub-marginal, costal, median and sub-median). Pterostigma conspicuous. Hind wing with five veins, without closed cells; 6 hamuli on anterior margin.
Comments: M. parallelus workers are unmistakable because of the lack of the third pair of mesonotal projections. They share with M. carinatus the non-notched anterior border of the clypeus and the absence of a tooth on the antero-lateral border of the postpetiole. They can be easily separated from congeneric workers by the eight promesonotal spines, weaker outer frontal carinae branches, longer propodeal and petiolar spines than in any other species of the genus, and by the dorsal surface of postpetiole with a deep circular impression.
Biology: The first record of a nest of this species was by Luederwaldt (1918), who described briefly a small spherical shelter nearly 15 cm deep in the soil, with 4 cm in diameter, and with the fungus garden resting on the bottom of the chamber. Mayhé-Nunes (1995) excavated 13 nests in a dirt road bordering the edge of a forest at Universidade Federal de Viçosa campus, Minas Gerais State, Brazil. The biggest nest showed a conical mound around the entrance (Fig. 9), with the fungus garden hanging from the roof chamber at nearly 10 cm depth, 7 cm in wide and wiyh 4 cm in height (Fig. 10). Above this chamber there was a smaller one, probably built by the foundress gyne when establishing the nest (Fig. 11); the same pattern was observed by Solomon et al. (2004) in two nests dug in Argentina. They found also one nest in São Gabriel da Cachoeira, Amazonas State, which represents the northernmost geographic record for the genus. The average measurements of nests excavated by Solomon et al. (2004) were: 10.8 cm (S.D. 3.9; n = 18) depth bellow ground, 4.9 cm in wide (S.D. 2.0; n = 16) and with 4.2 cm in height (S.D. 3.2; n = 14), with the fungus gardens suspended from the roof chambers by rootlets. The estimated population was of approximately 100 workers; gynes and males were collected in only one São Gabriel nest, in August 1992. Unlike other species of the genus, M. parallelus seems to prefer open habitats (Kempf 1960; Mayhé-Nunes 1995; Solomon et al. 2004).
Material examined: BRAZIL: Amazonas State, Humaitá, ACAR, 22.iv.1975, V. P. da Silva col. one worker (MZSP). Bahia State, Encruzilhada, 980m, xi.1974, Seabra & Alvarenga col. 2 males (MZSP). Goiás State, Anápolis, 12.ii.1958, W. Kempf col. # 2322, one worker (MZSP); Jatai, xii.1972, F. M. Oliveira col. # [Kempf coll.] 8934, one male (MZSP). Mato Grosso State, Faz. Fortaleza, 19.ii.1976, J.L.M. Diniz col. 6 workers (MZSP). Minas Gerais State, Belo Oriente, 25.ii.1996, A. Mayhé col. 2 workers (CECL); Caraça, Sta. Bárbara, xi.1971, F. M. Oliveira col. one male (MZSP); Dionisio, 4.xii.1989, N.G. Campos col. one gyne (CECL); Pedra Azul, 800 m, xi.1971, Seabra & Alvarenga col. # [Kempf coll.] 9316, 3 males (MZSP); Três Poços, E. F. C. Br, 12.vii.1927, Borgmeier col. # 1591, one worker (MZSP); Viçosa, 18.ii.1993, A. Mayhé & M. Oliveira cols. 27 workers, one gyne, 4 males (CECL). Rio de Janeiro State, Campos, 28.iv.1983, C.R. Gonçalves col. one worker (CECL); Nova Iguaçu, Parque Municipal, 9.vi.2005, A. Mayhé col. one worker (CECL); [Seropédica] Escola Nacional Agronomia, Km 47, C. R. Gonçalves # 1328, 12 workers, one male (MZSP). São Paulo State, Araçatuba, x.1961, J. Lane col. one male (MZSP); Botucatu, Faz. Lajeado, 28.ix.1990, B. H. Dietz col. one worker (MZSP); Campo Limpo, 20.ii.1966, W. W. Kempf. col. 4 workers (MZSP); Guaratinguetá, 15.xi.1958, W. Kempf col. one worker (MZSP); Mairiporã (represa), 23.viii.2003, Klingenberg & Fernandes Lourenço col. 2 workers (MZSP); São Paulo, Ipiranga, H. Luederwaldt col. # MP 12692 syntypes of Atta (Mycocepurus) luederwaldti Forel, 1911, 8 workers (MZSP); São Paulo, 31.iii.1960, W. Kempf col. one worker (MZSP). ARGENTINA, Tucumán, 26.iii.1948, N. Kusnezov col. # 1425, 5 workes (MZSP).
Mycetarotes senticosus Kempf, 1960
Mycetarotes senticosus Kempf, 1960: 282 (worker, São Paulo, Brazil, key); Kempf, 1972: 145 (catalog); Mayhé-Nunes, 1995: 202 (key); Schultz & Meier, 1995: 352, 353, 357 (larva); Bolton, 1995: 268 (catalog).
Type material (workers): Holotype: São Paulo State, Federal Highway São Paulo-Curitiba km 40, 7.v.1960, W.W. Kempf col., and three paratypes in MZSP (examined). BRAZIL: Rio de Janeiro State, Petrópolis, 21.iv.1945, W.W. Kempf col. one individual. Santa Catarina State, Chapecó, xii.1957, F. Plaumann col. one individual.
Diagnosis: Resembling M. acutus Mayhé-Nunes, but the preocular carinae ends basally in relation to the frontal ones, with the later subparallel above frontal lobes, mesosomal projections stouter than in congeneric species and with blunt apices; postpetiole faintly impressed on dorsal surface.
Description: Worker (Kempf´s measurements in mm, n = 4): TL 3.7-4.0; HL 0.94-0.9; HW 0.87-0.92; IFW 0.33; ScL 0.91-0.94; WL 1.16-1.27. Yellowish brown. Integument opaque, finely reticulo-punctate.
Head. Cephalic capsule slightly longer than broad (CI 90). Chewing border of mandible with 6 teeth. Median anterior border of clypeus with a small notch; frontal lobes subtriangular (FLI 39). Frontal carinae without constriction above the frontal lobes, diverging caudad in two branches above eyes, the inner ones reaching the base of the spines on occipital margin, and the outer branch weaker. Supraocular projections bluntly tuberculate. Occipital margin in full-face view medially notched between the inner spines and the outer ones. All funicular segments longer than broad.
Mesosoma. With the pronotum in frontal view, midpronotal spines as long as lateral ones. Mesonotal dorsum with three pairs of projections; the first two spines are longer than the other mesosomal projections; third pair smaller. Basal face of propodeum with two longitudinal ridges, mesially projected as conspicuous spines; propodeal spines slender, directed obliquely backwards.
Waist and gaster. Petiolar dorsum with a pair of spines directed upwards, placed relatively away from the posterior border, in a distance nearly equal to the length of the spines. Postpetiole with a tooth near each side of the lateral anterior margin; posterior margin concave; dorsal surface vestigially impressed. Tergum I of gaster longitudinally marginated by low ridges; longitudinal ridges on disc absent..
Gyne and Male. Unknown.
Comments: See M. acutus Mayhé-Nunes.
Biology: Unknown. According to Kempf (1960: 283), the specimens collected by him "were taken as strays from forest floor cover."
Material examined: BRAZIL: Santa Catarina State, Nova Teutônia, vi.1960, F. Plaumann col. one worker (MZSP).
DISCUSSION
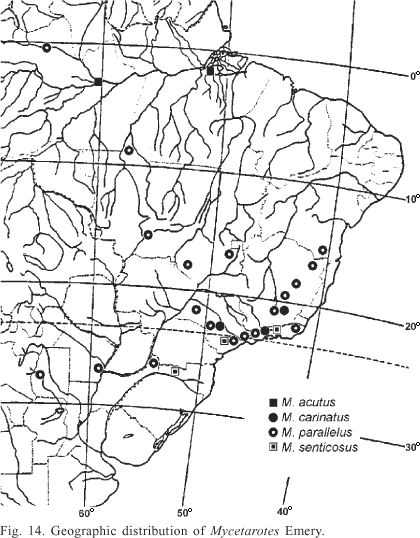
Mycetarotes distribution is restricted to South America, more commonly collected thus far in southeastern Brazil (Fig. 14). The south-north limits of its geographic distribution are respectively Tucumán, Argentina and São Gabriel da Cachoeira, Amazonas State, Brazil, matching the distribution of its most common species, M. parallelus, while M. acutus and M. carinatus have restricted known occurrences, respectively in northern and southeastern Brazil. M. senticosus has been collected in a few Brazilian localities in the southern and southeastern regions, always above 650 m of altitude.
Workers of Mycetarotes share some of the main diagnostic features of the Cyphomyrmex rimosus group (Kempf 1964), but clearly differ by the relatively smaller frontal lobes index and longer antennal scapes, frontal carinae diverging into two branches, presence of occipital spines, and bispinose petiolar dorsum. Chapela et al. (1994) proposed a close phylogenetic relationship between Cyphomyrmex and Mycetarotes. Schultz & Meier (1995), however, found a synapomorphy that supports the Mycetarotes clade parallelus-senticosus: the presence of spinules over the entire head of Mycetarotes larvae, while restricted to the clypeus and genae in Cyphomyrmex. Moreover, all larvae of Cyphomyrmex of the rimosus group included in the analysis have pronounced anterior abdominal ventromedian bosses, reduced in the Cyphomyrmex of the strigatus group, but completely absent in Mycetarotes. Villensen et al. (2002) accepted the topology of Schultz & Meier's cladogram, where Mycetarotes appears as the sister group of a clade encompassing Mycetosoritis, Cyphomyrmex and Mycetophylax.
Mycetarotes has been considered rare because of its scarcity in entomological collections (Mayhé-Nunes 1995). However, recent surveys show M. parallelus to be relatively widespread and common in central and eastern South America (Mayhé-Nunes & Jaffé 1998). Mycetarotes senticosus and M. carinatus appear to be truly uncommon, being known mostly in southeastern Brazil (Kempf 1972; Mayhé-Nunes 1995; Mayhé-Nunes & Lanziotti 2004). Mycetarotes acutus original description was based on a single worker specimen collected in Manaus, Brazil, and it may be mistaken by M. senticosus (Mayhé-Nunes, 1995). Solomon, et al. (2004) recognized the specimens they collected in Manaus as M. acutus, a species also recently collected by Carlos Eduardo D. Sanhudo in Melgaço, Pará State, Brazil, as confirmed by one of us (AJMN).
Solomon et al. (2004) described how similar the nests of three species of Mycetarotes are, not only in their architecture, with a single chamber, but without significant differences between average chamber depth, width, or height. They also estimated mature nest worker populations for M. parallelus (111 individuals) based on actual excavations, bigger than the average "lower"-attine colony size (approximately 350 individuals). This is accordance with the traditional view that more "basal" attine species preferably live in relatively small colonies (Schultz & Meier, 1995).
Acknowledgments. We thank Carlos Eduardo Dias Sanhudo (INPA graduate student) and Jacques H. C. Delabie (CEPLAC/CEPEC) for allowing us to study samples of M. acutus from Pará and Manaus, respectively. Guilherme Orsolon de Souza (UFRRJ graduate student) for helpful comments, Alan Neil Andersen (CSIRO Australia) and Alexandre Pires Aguiar (MZSP) for critically reading the manuscript. This research was supported by grants of the Conselho de Desenvolvimento Científico e Tecnológico CNPq, Brazil, to both authors (respectively # 305876/2004-0 and 303248/87).
Received 28/03/2006; accepted 20/09/2006
- Bolton, B. 1995. A new general catalogue of the ants of the world Harvard University Press, 504 p.
- Bolton, B. 2003. Synopsis and classification of Formicidae. The American Entomological Institute 71: 1-370.
- Borgmeier, T. 1950. Bemerkungen zu Dr. Creighton´s Werk "The Ants of North America". Revista de Entomologia 21: 381386.
- Chapela, I. H.; S. A. Rehner; T. R. Schultz & U. G. Mueller. 1994. Evolutionary history of the symbiosis between fungus-growing ants and their fungi. Science 266: 16911694.
- Emery, C. 1905. Studi sulle formiche della fauna neotropica. XXVI. Bolletino de la Societá Entomologica Italiana 37: 107194.
- Emery, C. 1913. Études sur les Myrmicinae. V-VII. Annales de la Société Entomologique de Belgique 57: 250262.
- Emery, C. 1922. Hymenoptera. Fam. Formicidae. Subfam. Myrmicinae. Genera Insectorum 174B: 1397.
- Forel, A. 1911. Ameisen des Herrn Prof. v. Iheringi aus Brasilien (São Paulo usw.) nebst einigen anderen aus Südamerika und Afrika (Hym.). Deutsche Entomologische Zeitschrift 1911: 285312.
- Forel, A. 1913. Fourmis d'Argentine, du Brésil, du Guatémala & de Cuba. Bulletin de la Société Vaudoise des Sciences Naturelles 49: 203250.
- Kempf, W. W. 1960. Review of the ant genus Mycetarotes Emery (Hymenoptera: Formicidae). Revista Brasileira de Biologia 20: 277283.
- Kempf, W. W. 1964. A revision of the Neotropical fungus-growing ants of the genus Cyphomyrmex Mayr. Part 1: Group of strigatus Mayr. Studia Entomologica 7: 144.
- Kempf, W. W. 1972. Catálogo abreviado das formigas da região Neotropical (Hym., Formicidae). Studia Entomologica 15: 3344.
- Lanziotti, A. M. & A. J. Mayhé-Nunes. 2001. Redescription of the female of Mycetarotes parallelus (Emery, 1905) (Hymenoptera: Formicidae). Acta Biologica Leopoldensia 23: 175179.
- Luederwaldt, H. 1918. Notas myrmecologicas. Revista do Museu Paulista 10: 2964.
- Mayhé-Nunes, A. J. 1995. Sinopse do gênero Mycetarotes (Hym., Formicidae), com a descrição de duas espécies novas. Boletín de Entomologia Venezolana 10: 197205.
- Mayhé-Nunes, A. J.& C. R. F. Brandão. 2005. Revisionary studies on the attine ant genus Trachymyrmex Forel. Part 2: the Iheringi group (Hymenoptera: Formicidae). Sociobiology 45: 271305.
- Mayhé-Nunes, A. J. & K. Jaffé. 1998. On the biogeography of Attini (Hymenoptera: Formicidae). Ecotropicos 11: 4554.
- Mayhé-Nunes, A. J.& A. M. Lanziotti. 2004. Description of the female and male of Mycetarotes carinatus (Hymenoptera: Formicidae). Revista de Biologia Tropical 52: 109114.
- Mueller, U. G.; S. A. Rehner & T. R. Schultz. 1998. The evolution of the agriculture in ants. Science 281: 20342038.
- Schultz, T. R. 1993. Stalking the wild attine. Notes from Underground 8: 710.
- Schultz, T. R. & R. Meier. 1995. A phylogenetic analysis of the fungus-growing ants (Hymenoptera: Formicidae: Attini) based on morphological characters of the larvae. Systematic Entomology 20: 337370.
- Schultz, T. R.; S. E. Solomon; U. G. Mueller; P. Villesen; J. J. Boosmsma; R. M. M. Adams & B. Norden. 2002. Cryptic speciation in the fungus-growing ants Cyphomyrmex longiscapus Weber and Cyphomyrmex muelleri Schultz and Solomon, new species (Formicidae, Attini). Insectes Sociaux 49: 331343.
- Solomon, S. E.; U. G. Mueller; T. R. Schultz; C. R. Currie; S. L. Price; A. C. Oliveira da Silva-Pinhati; M. Bacci Jr. & H. L. Vasconcelos. 2004. Nesting biology of the fungus growing ants Mycetarotes Emery (Attini, Formicidae). Insectes Sociaux 51: 333338.
- Villesen, P.; T. Murakami; T. R. Schultz; & J. J. Boomsma. 2002. Identifying the transition between single and multiple mating of queens in fungus-growing ants. Proceedings of the Royal Society of London 269: 14511458.

Publication Dates
-
Publication in this collection
30 Jan 2007 -
Date of issue
Dec 2006
History
-
Accepted
20 Sept 2006 -
Received
28 Mar 2006