Resumos
Bactérias diazotróficas endofíticas contribuem para o desenvolvimento das plantas por meio da fixação biológica do nitrogênio, produção e liberação de substâncias reguladoras do crescimento vegetal, podendo, assim, facilitar a revegetação de solos degradados por atividades antrópicas. No entanto, pouco se conhece sobre as populações destas bactérias em solos ou plantas de áreas de mineração. Objetivando avaliar o efeito de diferentes tipos de vegetação e tempo de reabilitação de áreas degradadas por mineração de bauxita na densidade e diversidade de algumas espécies de bactérias diazotróficas endofíticas, realizaram-se, em duas épocas, amostragens de solo, de dois ambientes distintos, submetidos a diferentes processos de reabilitação. A densidade, avaliada pelo número mais provável, utilizando os meios de cultura: NFb, JNFb e Fam, para Azospirillum brasilense e A. lipoferum, Herbaspirillum spp. e A. amazonense, respectivamente, variou de 0 a 2,0 x 10(4) bactérias por grama de solo e mostrou que o tipo de vegetação influiu nas populações destas bactérias. Foram encontradas densidades maiores em solos revegetados com gramíneas: braquiária (Brachiaria decumbens), capim-azevém (Lolium multiflorum) e capim-gordura (Melinis minutiflora). Contudo, estas densidades podem ser consideradas baixas, se comparadas às de solos agrícolas, e não apresentaram relação com o tempo de reabilitação da área. Foram encontrados 36 fenótipos culturais em meio batata, entre os 72 isolados obtidos dos três meios de cultura utilizados. A partir destes, foram formados sete grandes grupos com similaridade superior ou igual a 63 %, dos quais cinco, representando 62,5 % do total de isolados obtidos, continham as estirpes-tipo de Burkholderia brasilensis, Herbaspirillum seropedicae e Azospirillum spp. (A. brasilense, A. amazonense, A. lipoferum, A. irakense). Apesar da baixa densidade, este grupo de bactérias apresentou alta diversidade fenotípica no ambiente estudado.
Azospirillum spp.; Herbaspirillum; Burkholderia; fixação biológica de nitrogênio; gramíneas
Diazotrophic endophytic bacteria enhance plant growth through biological nitrogen fixation and production and release of plant growth regulating substances, which facilitate the revegetation of areas degraded by human activities. However, little is known about populations of such bacteria in soils or plants of mining areas. Aiming to study the effects of different vegetation types and rehabilitation periods on some endophytic diazotrophic bacteria species, soil samples were collected under two environmental conditions ("Campo" and "Serra") in bauxite mined areas undergoing different rehabilitation processes. Population densities were evaluated by the most probable number method in media (NFb, JNFb, and Fam for Azospirillum brasilense and A. lipoferum, Herbaspirillum spp. and A. amazonense, respectively) and ranged from 0 to 2.0 x 10(4) bacteria g-1 soil. The vegetation types affected the diazotrophic populations. Highest densities were found in mined soils revegetated with the grass species brachiaria (Brachiaria decumbens), rye grass (Lolium multiflorum) and capim-gordura (Melinis minutiflora). However, these densities are considered low compared to those found in agricultural soils. No relationship was found between the rehabilitation time and population density. Thirty-six cultural phenotypes were found on potato medium among 72 isolates out of all three culture media. At a similarity of at least 63 %, these isolates formed seven great groups, five of which (comprising 62.5 % of the total isolates) contained the type strains of Burkholderia brasilensis, Herbaspirillum seropedicae, and Azospirillum spp. (A. brasilense, A. amazonense, A. lipoferum, A. irakense). In spite of its low density, this group of bacteria presented a high phenotypic diversity in the studied environment.
Azospirillum spp.; Herbaspirillum; Burkholderia; biological nitrogen fixation; grasses
SEÇÃO III - BIOLOGIA DO SOLO
Densidade e diversidade fenotípica de bactérias diazotróficas endofíticas em solos de mineração de bauxita, em reabilitação
Density and phenotypic diversity of endophytic nitrogen fixing bacteria in soils under rehabilitation after bauxite mining
R. MelloniI; R. S. A. NóbregaII; F. M. S. MoreiraIII; J. O. SiqueiraIII
IProfessor do Departamento de Mecânica da Universidade Federal de Itajubá UNIFEI. CEP 37500-903. Itajubá (MG).E-mail: rmelloni@iem.efei.br
IIDoutoranda em Solos e Nutrição de Plantas, Departamento de Ciência do Solo, Universidade Federal de Lavras UFLA. Caixa Postal 37, CEP 37200-000 Lavras (MG). Bolsista do CNPq. E-mail: rafaelanobrega@ufla.com.br
IIIProfessor do Departamento de Ciência do Solo, UFLA. Bolsista CNPq. E-mail: fmoreira@ufla.br; siqueira@ufla.br
RESUMO
Bactérias diazotróficas endofíticas contribuem para o desenvolvimento das plantas por meio da fixação biológica do nitrogênio, produção e liberação de substâncias reguladoras do crescimento vegetal, podendo, assim, facilitar a revegetação de solos degradados por atividades antrópicas. No entanto, pouco se conhece sobre as populações destas bactérias em solos ou plantas de áreas de mineração. Objetivando avaliar o efeito de diferentes tipos de vegetação e tempo de reabilitação de áreas degradadas por mineração de bauxita na densidade e diversidade de algumas espécies de bactérias diazotróficas endofíticas, realizaram-se, em duas épocas, amostragens de solo, de dois ambientes distintos, submetidos a diferentes processos de reabilitação. A densidade, avaliada pelo número mais provável, utilizando os meios de cultura: NFb, JNFb e Fam, para Azospirillum brasilense e A. lipoferum, Herbaspirillum spp. e A. amazonense, respectivamente, variou de 0 a 2,0 x 104 bactérias por grama de solo e mostrou que o tipo de vegetação influiu nas populações destas bactérias. Foram encontradas densidades maiores em solos revegetados com gramíneas: braquiária (Brachiaria decumbens), capim-azevém (Lolium multiflorum) e capim-gordura (Melinis minutiflora). Contudo, estas densidades podem ser consideradas baixas, se comparadas às de solos agrícolas, e não apresentaram relação com o tempo de reabilitação da área. Foram encontrados 36 fenótipos culturais em meio batata, entre os 72 isolados obtidos dos três meios de cultura utilizados. A partir destes, foram formados sete grandes grupos com similaridade superior ou igual a 63 %, dos quais cinco, representando 62,5 % do total de isolados obtidos, continham as estirpes-tipo de Burkholderia brasilensis, Herbaspirillum seropedicae e Azospirillum spp. (A. brasilense, A. amazonense, A. lipoferum, A. irakense). Apesar da baixa densidade, este grupo de bactérias apresentou alta diversidade fenotípica no ambiente estudado.
Termos de indexação:Azospirillum spp., Herbaspirillum, Burkholderia, fixação biológica de nitrogênio, gramíneas.
SUMMARY
Diazotrophic endophytic bacteria enhance plant growth through biological nitrogen fixation and production and release of plant growth regulating substances, which facilitate the revegetation of areas degraded by human activities. However, little is known about populations of such bacteria in soils or plants of mining areas. Aiming to study the effects of different vegetation types and rehabilitation periods on some endophytic diazotrophic bacteria species, soil samples were collected under two environmental conditions ("Campo" and "Serra") in bauxite mined areas undergoing different rehabilitation processes. Population densities were evaluated by the most probable number method in media (NFb, JNFb, and Fam for Azospirillum brasilense and A. lipoferum, Herbaspirillum spp. and A. amazonense, respectively) and ranged from 0 to 2.0 x 104 bacteria g-1 soil. The vegetation types affected the diazotrophic populations. Highest densities were found in mined soils revegetated with the grass species brachiaria (Brachiaria decumbens), rye grass (Lolium multiflorum) and capim-gordura (Melinis minutiflora). However, these densities are considered low compared to those found in agricultural soils. No relationship was found between the rehabilitation time and population density. Thirty-six cultural phenotypes were found on potato medium among 72 isolates out of all three culture media. At a similarity of at least 63 %, these isolates formed seven great groups, five of which (comprising 62.5 % of the total isolates) contained the type strains of Burkholderia brasilensis, Herbaspirillum seropedicae, and Azospirillum spp. (A. brasilense, A. amazonense, A. lipoferum, A. irakense). In spite of its low density, this group of bacteria presented a high phenotypic diversity in the studied environment.
Index terms:Azospirillum spp., Herbaspirillum, Burkholderia, biological nitrogen fixation, grasses.
INTRODUÇÃO
Bactérias diazotróficas endofíticas são aquelas que fixam N2 atmosférico e colonizam o interior de tecidos vegetais sem causar sintomas de doenças (Döbereiner, 1992). Dentre as mais estudadas, destacam-se aquelas pertencentes aos gêneros Azospirillum, Herbaspirillum, Acetobacter, Burkholderia e Azoarcus. Bactérias do gênero Azospirillum são consideradas endofíticas facultativas (Baldani et al., 1997), pois, além de colonizarem os hospedeiros, podem sobreviver no solo na forma de cistos (Bashan & Holguin, 1997) e, ou, utilizar poli-b-hidroxibutirato como fonte de carbono e energia (Baldani et al., 1999) na ausência destes. Burkholderia é outro gênero considerado endofítico, cuja capacidade diazotrófica foi descoberta recentemente (Gillis et al., 1995; Baldani et al., 1997). Já o gênero Herbaspirillum é considerado endofítico obrigatório, por apresentar baixa sobrevivência no solo (Olivares et al., 1996). Apesar de a especificidade com relação a seus hospedeiros ser baixa (Baldani et al., 1997), a associação dessas bactérias fixadoras de nitrogênio é mais freqüente com gramíneas (Magalhães & Döbereiner, 1984; Bashan & Holguin, 1997; Baldani et al., 1999).
Os microrganismos diazotróficos endofíticos podem desempenhar importante papel na reabilitação e sustentabilidade dos ecossistemas, uma vez que incorporam N por meio da fixação biológica em quantidades que podem variar de 25 a 50 kg ha-1 ano-1 de N e produzem e liberam substâncias reguladoras do crescimento vegetal, como auxinas, giberelinas e citocininas, as quais contribuem para melhorar a nutrição mineral e utilização de água pelas plantas (Bazzicalupo & Okon, 2000). Por estes e outros efeitos, a introdução prévia de gramíneas em áreas desprovidas de vegetação é muito vantajosa. No entanto, Bashan & Holguin (1997) e Baldani et al. (1999) relataram que a ocorrência e a atividade destas bactérias no solo e na planta são fortemente influenciadas por estresses físicos (baixa umidade e alta temperatura), químicos (acidez e baixos teores de nutrientes e carbono) e biológicos (espécie vegetal não-hospedeira), os quais são comuns em solos de mineração de bauxita, razão por que podem influenciar as populações destas bactérias.
Embora gramíneas sejam freqüentemente plantadas ou semeadas em áreas em processo de reabilitação, não há estudos sobre bactérias diazotróficas associativas nestas e, ou, em outras espécies vegetais utilizadas com tal finalidade em áreas de mineração. Os objetivos do presente estudo foram: avaliar a densidade e a diversidade fenotípica de algumas espécies de bactérias diazotróficas endofíticas, em solos de mineração de bauxita, sob diferentes tipos de vegetação, incluindo gramíneas, bem como o efeito do tempo de reabilitação e das estações do ano.
MATERIAL E MÉTODOS
De acordo com o tempo de reabilitação e tipo de vegetação após a mineração de bauxita, selecionaram-se, para o presente estudo, 15 áreas, originalmente sob Cambissolo distrófico de baixa fertilidade, cujas características edáficas químicas e físicas encontram-se em Melloni (2001). Todas as áreas estão localizadas na empresa de mineração Alcoa Alumínio S/A em Poços de Caldas (MG), em dois ambientes distintos, identificados como Campo e Serra. Destas áreas, sete foram selecionadas no Campo, com vegetação original típica de campo tropical, composta por espécies herbáceas e arbustivas, e oito, na Serra, de relevo mais montanhoso e com cobertura vegetal tipo capoeira e mata. As áreas foram mineradas nas últimas décadas e submetidas a procedimentos variados de reabilitação, encontrando-se atualmente com diferentes tipos de vegetação: gramíneas, leguminosas herbáceas, bracatinga, espécies arbóreas nativas e Eucalyptus saligna, com diferentes idades e estádios de reabilitação (Quadro 1).
De cada área, coletaram-se três amostras de solo próximas às raízes, compostas de dez subamostras retiradas na profundidade de 0-10 cm, em duas estações do ano: verão (março de 1999) e primavera (dezembro de 1999). Estas coletas foram feitas após uma avaliação semiquantitativa da presença de diazotróficos, em amostras de solo de todas as áreas em agosto de 1998, pelo método de cultura de enriquecimento nos mesmos meios utilizados para contagem (cinco culturas por meio por área). Estas épocas foram escolhidas, visto que representavam o período de seca (agosto) e o início e fim da estação chuvosa (março e dezembro), períodos distintos de atividade biológica especialmente no que se refere ao vigor e ciclo da vegetação.
As amostras foram acondicionadas em sacos plásticos, mantidas em caixas térmicas e transportadas ao laboratório no prazo máximo de 24 h, quando foram submetidas à contagem de bactérias endofíticas, por meio do método do número mais provável (NMP). Realizaram-se diluições sucessivas das amostras de solo em água, de 10-1 (10 g solo: 90 mL H2O esterilizada) a 10-5, antes da inoculação de 0,1 mL destas suspensões em frascos que continham 4 mL dos meios de cultura semi-sólidos que favorecem o crescimento de Azospirillum spp. (A. lipoferum e A. brasilense) (NFb), A. amazonense (Fam) e Herbaspirillum spp. (JNFb), preparados conforme Döbereiner (1980), Magalhães (1983) e Baldani et al. (1992), respectivamente, com três repetições por diluição. Estas culturas foram mantidas sob incubação por sete dias a 28 ºC, avaliando-se, posteriormente, a presença ou ausência de crescimento bacteriano pela formação de película próxima à superfície no meio.
Utilizou-se o programa MPNES (Woomer et al., 1990) para determinação do NMP destas bactérias por grama de amostra de solo, seguindo-se a transformação dos dados para log (NMP + 1) e comparação das áreas pelo teste Duncan a 5 % de significância, por meio do programa estatístico SANEST (Zonta et al., 1984). Cada área amostrada foi considerada tratamento (com três repetições por área/tratamento), sendo todas analisadas em delineamento inteiramente ao acaso.
Para obtenção dos isolados e avaliação da diversidade fenotípica, transferiu-se, com alça de platina, uma porção de películas crescidas nas culturas utilizadas para avaliação semiquantitativa e contagem para os mesmos meios de cultura semi-sólidos (JNFb, NFb e Fam). As novas películas formadas foram repicadas para estes meios, porém, com consistência sólida (15 g L-1 de ágar acrescidos de extrato de levedura), conforme descrito nas referências já citadas. Após cinco dias ou mais de incubação a 28 ºC, colônias isoladas foram transferidas novamente para meios de cultura semi-sólidos com posterior verificação de crescimento, indicado pela formação de película típica. Para a purificação final, porções das novas películas formadas foram riscadas no meio de cultura batata (Döbereiner et al., 1995) e, após sete dias de incubação a 28 ºC, caracterizadas quanto à cor, diâmetro e consistência das colônias. As estirpes-tipo de Azospirillum lipoferum (BR11080T), A. brasilense (BR11101T), A. amazonense (BR11140T), A. irakense (CIP 103311; KBC1), Burkholderia brasilensis (BR 11340T) e Herbaspirillum seropedicae (BR11175T) foram também avaliadas quanto às mesmas características. A estirpe-tipo de Burkholderia foi incluída como referência, pois foi relatado seu isolamento em meios similares aos utilizados neste estudo (Baldani, 1996).
Todos os isolados e estirpes-tipo foram agrupados em dendrograma de similaridade pelo método Complete Linkage e Distância Euclidiana (Everitt, 1993), por meio do programa SAS INSTITUTE 5.0. Para isso, atribuíram-se os seguintes valores às características morfológicas: (a) diâmetro médio das colônias: 1 (< 1 mm), 2 (1-2 mm), 3 (2-3 mm), 4 (> 3 mm); (b) cor: 1 (creme), 2 (creme centro-escuro), 3 (creme-claro), 4 (rosa), 5 (creme-transparente), 6 (branco); (c) consistência das colônias: 1 (média), 2 (mole) e 3 (dura/quebradiça).
RESULTADOS E DISCUSSÃO
Em ambas as épocas, os maiores valores de densidade total de bactérias diazotróficas endofíticas foram obtidos nas amostras de solo das áreas revegetadas com gramíneas (Quadro 2), as quais são consideradas plantas hospedeiras comuns destas bactérias (Magalhães & Döbereiner, 1984; Baldani et al., 1999). No verão, em áreas com aproximadamente seis meses de reabilitação com braquiária ou capim-gordura, registraram-se densidades totais de 3,1 x 102 bactérias g-1 solo, no campo, e de 3,0 x 103 bactérias g-1 solo, na serra, indicando a rápida proliferação destas bactérias no solo, após a atividade de mineração e início da reabilitação. Isto ocorreu provavelmente pela fácil disseminação destas bactérias por meio das sementes das espécies plantadas na área e pela maior afinidade destas bactérias pela rizosfera de gramíneas.
No entanto, estes valores foram relativamente pouco alterados em outras áreas com maior tempo de reabilitação, tanto na primavera quanto no verão. O valor máximo obtido foi de 2,0 x 104 bactérias g-1 solo em área de campo (verão) reabilitada com capim-azevém e Eupathorium quatro anos. Apesar de estar a densidade diretamente relacionada com a espécie vegetal associada (Baldani et al., 1999), estes valores podem ser considerados baixos, quando comparados aos encontrados para Azospirillum spp. em raízes e solo de outras gramíneas, como milho e sorgo no Sudeste (106 a 107 bactérias g-1 solo) (Döbereiner, 1978, citado por Döbereiner et al., 1995), em raízes não lavadas de pupunha e braquiária na Amazônia (105 bactérias g-1 solo) (Magalhães & Döbereiner, 1984), em raízes desinfestadas superficialmente e na rizosfera de espécies de braquiária em Goiás e Minas Gerais (103 a 107 bactérias g-1 solo) (Reis Jr. et al., 2000).
Uma possível explicação para a baixa densidade nas áreas mineradas é que estas bactérias apresentavam baixa ocorrência no ambiente original. De fato, na área da serra, estas não foram encontradas e no campo tiveram ocorrência rara. Nas áreas M10, B14, E16, M18 e RF-SE (da serra) e E3 e E16 (do campo), registraram-se os menores valores de densidade, sendo ausentes em várias amostras. Isto pode estar relacionado com a ausência de plantas hospedeiras (gramíneas) nestas áreas e, ou, com a baixa disponibilidade de nutrientes nestes solos. Conforme Melloni (2001), esses solos apresentaram baixos teores de matéria orgânica, P, K e baixa saturação por bases. Isto corrobora os resultados de Magalhães & Döbereiner (1984) que relataram baixa densidade destas bactérias, sendo muitas vezes nula em solos desprovidos de gramíneas e com baixa fertilidade, como os de campinas e florestas de terra firme na Amazônia.
Estes resultados também podem indicar que estes microrganismos, a exemplo do que ocorre com as simbioses rizóbios e leguminosas, são importantes principalmente nos estádios iniciais da sucessão da vegetação, os quais apresentam deficiência de N. Nos ecossistemas em clímax ou próximos a este, a fixação biológica de N2 não é estimulada ou favorecida porque as demandas por N são supridas por uma ciclagem mais eficiente (Moreira, 1994).
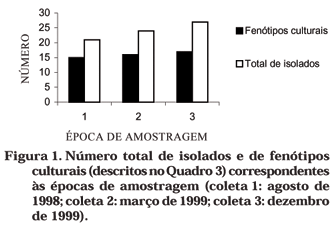
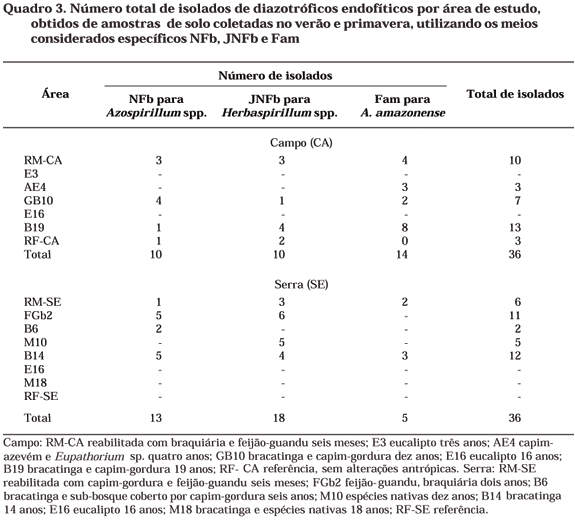
O número total de isolados por área e por coleta (época de amostragem) encontra-se, respectivamente no quadro 3 e na figura 1. Foram obtidos isolados de todos os meios utilizados e da maioria das áreas estudadas onde se registrou a presença de diazotróficos (Quadros 2 e 3). Assim como observado para a densidade de endofíticos, em áreas cobertas por capim-gordura e braquiária (RM-CA, GB10 e B19 do campo e FGb2, da serra), obteve-se o maior número de isolados, com valores superiores aos das áreas de referência. A revegetação com gramíneas contribuiu para maior multiplicação destas bactérias no solo e, conseqüentemente, para maior probabilidade de isolamento. Embora tenha sido registrada a ocorrência destas bactérias em solos com eucalipto, na primavera (Quadro 2), não se obtiveram isolados nestas áreas graças à contaminação e descarte de culturas cuja pureza foi comprometida. O número de fenótipos culturais, em relação ao número total de isolados, não variou entre as épocas amostradas (Figura 1), indicando estabilidade quanto à riqueza fenotípica deste grupo de bactérias em áreas de mineração.
Foi observada uma grande diversidade fenotípica cultural, sendo encontrados trinta e seis fenótipos (Quadro 4). Três destes fenótipos foram exclusivos de estirpes-tipo de A. brasilense, A. lipoferum, e H. seropedicae. Fenótipos das estirpes de A. irakense e A. amazonense foram similares aos isolados UFLA36 e UFLA61, respectivamente. Seis isolados tiveram fenótipo semelhante ao de Burkholderia brasilense. Os 30 fenótipos restantes não apresentaram similaridade cultural com nenhuma das estirpes-tipo.
Em cerca de 84 % de similaridade entre fenótipos culturais de isolados e de estirpes-tipo de espécies conhecidas (Figura 2), encontram-se os grupos de fenótipos listados no quadro 4, os quais apresentam alto grau de discriminação de suas características culturais. Diminuindo o nível de similaridade para cerca de 63 %, verifica-se que houve maior coerência taxonômica entre os grupos com base nas estirpes-tipo. Dos oito grandes grupos formados, cinco apresentaram similaridade com estirpes-tipo. Os quatro grupos que continham as quatro estirpes-tipo de a-Proteobacteria das espécies do gênero Azospirillum compreenderam 36 isolados. O quinto grupo, que compreendeu as estirpes-tipo de espécies de b-Proteobacteria dos gêneros Herbaspirillum e Burkholderia, continha 17 isolados. Os três grupos sem estirpes-tipo continham 19 isolados que podem pertencer a outras espécies de diazotróficos. Apesar de os métodos moleculares serem considerados essenciais à identificação destas bactérias (Kirchhof et al., 1997), estes resultados indicam que características culturais são também aplicáveis em estudos exploratórios da diversidade destas bactérias em diferentes ambientes.
Mesmo ausente ou ocorrendo em baixa densidade no ambiente original (áreas de referência), estas bactérias podem ter sobrevivido nos solos reabilitados graças à capacidade de formar estruturas de resistência, como cistos, quando em condições estressantes (Bashan & Holguin, 1997), e à possibilidade de fixar N2 na ausência de fonte exógena de C (Okon & Itzigsohn, 1992). Estudos têm mostrado ainda que estas espécies são capazes de acumular poli-b-hidroxibutirato, um material de reserva que lhes permite resistir a condições de estresses ambientais (Döbereiner, 1991; Baldani, 1996) e que, portanto, possibilitaria a sua sobrevivência em solos minerados.
Estes resultados mostram também que, embora o maior número de isolados tenha sido obtido com a utilização do meio JNFb (Quadro 4), a análise de similaridade (Figura 2) indica que estes não necessariamente apresentaram maior similaridade à Herbaspirillum. A falta de especificidade do meio JNFb, já considerada por Baldani et al. (1999), indica que o meio JNFb permite o crescimento de outras bactérias endofíticas, principalmente aquelas capazes de tolerar a maior acidez inicial deste meio, como Azospirillum. Do mesmo modo, Magalhães & Döbereiner (1984) mostraram que, mesmo sendo as espécies de Azospirillum - favorecidas nos meios NFb e Fam, outros microrganismos diazotróficos podem crescer nestes meios, como ocorreu no presente estudo.
CONCLUSÕES
1. Bactérias diazotróficas endofíticas não ocorreram na área de referência da serra (mata) e ocorreram em baixas densidades na referência do campo.
2. A reabilitação do solo com as gramíneas: capim-gordura, azevém e braquiária favoreceu a ocorrência de bactérias diazotróficas endofíticas em áreas de mineração de bauxita reabilitadas. No entanto, estas densidades são baixas quando comparadas àquelas geralmente encontradas em áreas agrícolas.
3. O tempo de reabilitação e a variação sazonal não influenciaram a densidade e a diversidade de diazotróficos endofíticos nas diversas áreas estudadas.
4. Houve grande diversidade fenotípica cultural entre os isolados obtidos das diferentes áreas em reabilitação e parte significativa destes não apresentou similaridade com as estirpes-tipo das espécies de diazotróficos pesquisadas.
AGRADECIMENTOS
Os autores agradecem à Embrapa-Agrobiologia, na pessoa da pesquisadora Rosa Maria Pittard, pelo fornecimento das estirpes-tipo, e ao Prof. Augusto Ramalho de Moraes, pelo auxílio nas análises estatísticas.
LITERATURA CITADA
Recebido para publicação em agosto de 2003 e aprovado em novembro de 2003.
Trabalho financiado pelo convênio FAEPE/ALCOA e FAPEMIG.
- BALDANI, V.L.D. Efeito da inoculação de Herbaspirillum spp. no processo de inoculação e infecção de plantas de arroz e, ocorrência e caracterização parcial de uma nova bactéria diazotrófica. Rio de Janeiro, Universidade Federal Rural do Rio de Janeiro, 1996. 265p. (Tese de Doutorado)
- BALDANI, V.L.D.; BALDANI, J.I.; OLIVARES, F.L. & DÖBEREINER, J. Identification and ecology of Herbaspirillum seropedicae and closely related Pseudomonas rubrisubalbicans Symbiosis, 13:65-73, 1992.
- BALDANI, V.L.D.; OLIVEIRA, E.; BALOTA, E.; BALDANI, J.I.; KIRCHHOF, G. & DÖBEREINER, J. Burkholderia brasilensis sp. nov., uma nova espécie de bactéria diazotrófica endofítica. An. Acad. Bras. Ci., 69:116, 1997.
- BALDANI, J.I.; AZEVEDO, M.S.; REIS, V.M.; TEIXEIRA, K.R.S.; OLIVARES, F.L.; GOI, S.R.; BALDANI, V.L.D. & DÖBEREINER, J. Fixação biológica de nitrogênio em gramíneas: avanços e aplicações. In: SIQUEIRA, J.O.; MOREIRA, F.M.S.; LOPES, A.S.; GUILHERME, L.R.G.; FAQUIN, V.; FURTINI NETO, A.E. & CARVALHO, J.G., eds. Inter-relação fertilidade, biologia do solo e nutrição de plantas. Viçosa, SBCS/UFLA/DCS, 1999. p.621-666.
- BASHAN, Y. & HOLGUIN, G. Azospirillum-plant relationships: enviromental and physiological advances (1990-1996). Can. J. Microbiol., 43:103-121, 1997.
- BAZZICALUPO, M. & OKON, Y. Associative and endophytic symbiosis. In: PEDROSA, F.; HUNGRIA, M.; YATES, M.G. & NEWTON, W.E., eds. Nitrogen fixation: from molecules to crop productivity. Dordrecht, Kluwer Academic Publishers, 2000. p.409-410.
- DÖBEREINER, J. Forage grasses and grain crops. In: BERGERSEN, F.J., ed. Methods for evaluating biological nitrogen-fixation. New York, John Wiley, 1980. p.535-555.
- DÖBEREINER, J. The genera Azospirillum and Herbaspirillum In: BALLOWS, A.; TRÜPER, H.G.; DWORKIN, M.; HARDER, W. & SHLEIFER, K., eds. The Prokaryotes, 2.ed. New York, Springer-Verlag, 1991. p.2236-2253.
- DÖBEREINER, J. Recent changes in concepts of plant bacteria interactions: endophytic N2 fixing bacteria. Ci. Cult., 44:310-313, 1992.
- DÖBEREINER, J.; BALDANI, V.L.D. & BALDANI, J.I. Como isolar e identificar bactérias diazotróficas de plantas não-leguminosas. Brasília, EMBRAPA-SPI e Seropédica, EMBRAPA-CNPAB, 1995. 60p.
- EVERITT, B.S. Cluster analysis. New York, John Wiley, 1993. 170p.
- GILLIS, M.; VAN, T.V.; BARDIN, R.; GOOR, M.; HEBBAR, P.; WILLEMS, A.; SEGERS, P.; KERSTERS, K.; HEULIN, T. & FERNADEZ, M.P. Polyphasic taxonomy in the genus Burkholderia leading to an emend description of the genus and proposition of Burkholderia vietnamiensis sp.nov. for N2-fixing isolates from rice in Vietnam. Inter. J. System. Bacteriol., 45:274-289, 1995.
- KIRCHHOF, G.; SCHLOTER, M.; ABMUS, B. & HARTMANN, A. Molecular microbial ecology approaches applied to diazotrophs associated with non-legumes. Soil Biol. Biochem., 29:853-862, 1997.
- MAGALHÃES, F.M.M. Caracterização e distribuição de uma nova espécie de bactéria fixadora de nitrogênio. Manaus, Universidade do Amazonas, 1983. 89p. (Tese de Mestrado)
- MAGALHÃES, F.M.M. & DÖBEREINER, J. Ocorrência de Azospirillum amazonense em alguns ecossistemas da Amazônia. R. Microbiol., 15:246-252, 1984.
- MELLONI, R. Densidade e diversidade de bactérias diazotróficas e fungos micorrízicos arbusculares em solos de mineração de bauxita. Lavras, Universidade Federal de Lavras, 2001. 173p. (Tese de Doutorado)
- MOREIRA, F.M.S. Fixação Biológica do nitrogênio em espécies arbóreas. In: ARAUJO, R.S. & HUNGRIA, M., eds. Microrganismos de importância agrícola. Brasília, Empresa Brasileira de Pesquisa Agropecuária, 1994. p.121-149.
- OKON, Y. & ITZIGSOHN, R. Poly-b-hydroxybutyrate metabolism in Azospirillum brasilense and the ecological role of PHB in the rhizosphere. FEMS Microbiol. Lett., 103:131-139, 1992.
- OLIVARES, F.L.; BALDANI, V.L.D.; REIS, V.M.; BALDANI, J.I. & DÖBEREINER, J. Occurrence of the endophytic diazotrophs Herbaspirillum spp. in roots, stems, and leaves, predominantly of Gramineae. Biol. Fertil. Soils, 21:197-200, 1996.
- REIS Jr., F.B.; SANTOS, S.T.; TEIXEIRA, K.R.S.; REIS, V.M. & DÖBEREINER, J. N2-fixing bacteria associated with Brachiaria pastures. In: PEDROSA, F.; HUNGRIA, M.; YATES, M.G. & NEWTON, W.E., eds. Nitrogen Fixation: from molecules to crop productivity. Dordrecht, Kluwer Academic Publishers, 2000. p.432.
- SAS INSTITUTE - Statistic Analysis. Versão 5.0, Cary, 1995.
- WOOMER, P.; BENNET, J. & YOST, R. Overcoming the inflexibility of most-probable-number procedures. Agron. J., 82:349-353, 1990.
- ZONTA, E.P.; MACHADO, A.A. & SILVEIRA JÚNIOR, P. Sistemas de análise estatística para microcomputadores (SANEST). Pelotas, Universidade Federal de Pelotas, 1984. 151p.
Datas de Publicação
-
Publicação nesta coleção
21 Jun 2004 -
Data do Fascículo
Fev 2004
Histórico
-
Recebido
Ago 2003 -
Aceito
Nov 2003