Resumos
A deficiência tardia de potássio na cultura do algodoeiro tem ocorrido com freqüência nas regiões do cerrado brasileiro. Um dos motivos poderia ser atribuído à baixa disponibilidade de água nessa época. Assim, procurou-se quantificar a contribuição da difusão e do fluxo de massa no suprimento de K às raízes do algodoeiro de acordo com a disponibilidade do nutriente e de água. Para tanto, realizou-se um experimento em vasos em casa de vegetação, utilizando amostras da camada arável (0-20 cm) de um Latossolo Vermelho típico, com 330 mg kg-1 de argila. O experimento constou de duas doses de potássio (15 e 121 mg dm-3 ), na forma de KCl, e quatro conteúdos de água (-0,03; -0,1; -0,5 e -1,0 MPa). As plantas foram colhidas aos 53 dias da emergência. A difusão foi o principal mecanismo de transporte de K no solo, variando de 72 a 96 % do total absorvido pelo algodoeiro. A influência do conteúdo de água do solo sobre os mecanismos de transporte de K foi maior em solos com maior concentração deste nutriente, razão por que o fluxo de massa cresce em importância em solos mais secos.
Gossypium hirsutum; nutrição mineral; solução do solo; transporte de K
Late season potassium (K) deficiency has been observed quite frequently for cotton crops in the Brazilian cerrado region. One possible reason for such a problem could be the low water availability at this period of the season. Thus, an experiment was conducted in order to quantify the relative contribution of mass flow and diffusion in supplying K to cotton roots, as affected by soil water and K availability. The arable layer of a typic Red Latosol (Haplortox), with 630 mg kg-1 sand, 40 mg kg-1 silt, and 330 mg kg-1 clay was sampled and filled into 5 L pots. The treatments were two K rates (15 and 121 mg dm-3), applied as potassium chloride, and four levels of soil water (-0.03, -0.1, -0.5, and -1.0 MPa). Two cotton plants were grown in each pot and harvested 53 days after plant emergence. Diffusion was the main transport mechanism of K to cotton roots, contributing from 72 to 96 % of the total uptake. The effect of soil water upon the K transport to the root surface was greater in soils with a higher K concentration. That is the reason why mass flow is more important in dryer soils.
Gossypium hirsutum; mineral nutrition; soil solution; K transport
SEÇÃO IV - FERTILIDADE DO SOLO E NUTRIÇÃO DE PLANTAS
Importância do fluxo de massa e difusão no suprimento de potássio ao algodoeiro como variável de água e potássio no solo(1) (1) Trabalho apresentado no XXVIII Congresso Brasileiro de Ciência do Solo, Londrina (PR), de 01 a 06 de Julho de 2001.
Importance of mass flow and diffusion on the potassium supply to cotton plants as affected by soil water and potassium
R. H. OliveiraI; C. A. RosolemII; R. M. TrigueiroI
IEstudante de Pós-Graduação, Departamento de Produção Vegetal, Faculdade de Ciências Agronômicas FCA/UNESP. Caixa Postal 237, CEP 18603-970 Botucatu (SP). E-mail: rosahonorato@fca.unesp.br
IIProfessor Titular do Departamento de Produção Vegetal, FCA/UNESP. Bosista do CNPq. E-mail: rosolem@fca.unesp.br
RESUMO
A deficiência tardia de potássio na cultura do algodoeiro tem ocorrido com freqüência nas regiões do cerrado brasileiro. Um dos motivos poderia ser atribuído à baixa disponibilidade de água nessa época. Assim, procurou-se quantificar a contribuição da difusão e do fluxo de massa no suprimento de K às raízes do algodoeiro de acordo com a disponibilidade do nutriente e de água. Para tanto, realizou-se um experimento em vasos em casa de vegetação, utilizando amostras da camada arável (0-20 cm) de um Latossolo Vermelho típico, com 330 mg kg-1 de argila. O experimento constou de duas doses de potássio (15 e 121 mg dm-3 ), na forma de KCl, e quatro conteúdos de água (-0,03; -0,1; -0,5 e -1,0 MPa). As plantas foram colhidas aos 53 dias da emergência. A difusão foi o principal mecanismo de transporte de K no solo, variando de 72 a 96 % do total absorvido pelo algodoeiro. A influência do conteúdo de água do solo sobre os mecanismos de transporte de K foi maior em solos com maior concentração deste nutriente, razão por que o fluxo de massa cresce em importância em solos mais secos.
Termos de indexação: Gossypium hirsutum, nutrição mineral, solução do solo, transporte de K.
SUMMARY
Late season potassium (K) deficiency has been observed quite frequently for cotton crops in the Brazilian cerrado region. One possible reason for such a problem could be the low water availability at this period of the season. Thus, an experiment was conducted in order to quantify the relative contribution of mass flow and diffusion in supplying K to cotton roots, as affected by soil water and K availability. The arable layer of a typic Red Latosol (Haplortox), with 630 mg kg-1 sand, 40 mg kg-1 silt, and 330 mg kg-1 clay was sampled and filled into 5 L pots. The treatments were two K rates (15 and 121 mg dm-3), applied as potassium chloride, and four levels of soil water (-0.03, -0.1, -0.5, and -1.0 MPa). Two cotton plants were grown in each pot and harvested 53 days after plant emergence. Diffusion was the main transport mechanism of K to cotton roots, contributing from 72 to 96 % of the total uptake. The effect of soil water upon the K transport to the root surface was greater in soils with a higher K concentration. That is the reason why mass flow is more important in dryer soils.
Index terms:Gossypium hirsutum, mineral nutrition, soil solution, K transport.
INTRODUÇÃO
A disponibilidade de nutrientes na rizosfera depende, dentre outros fatores, da concentração do nutriente na solução do solo e do transporte deste para a superfície radicular (Chen & Gabelman, 2000). A concentração do nutriente na solução do solo é de importância primária para o suprimento deste nutriente para a raiz (Marshner, 1995).
Os nutrientes são transportados até às raízes por meio de três mecanismos: intercepção radicular, difusão e fluxo de massa. A intercepção radicular ocorre quando os nutrientes são interceptados pelas raízes durante o processo de crescimento; somente pequena percentagem do nutriente total requerido é suprida por este processo (Marshner, 1995). Entretanto, o mecanismo de intercepção radicular não tem sido considerado no transporte de nutrientes, pois, atualmente, não é aceita a possibilidade de trocas diretas entre as partículas do solo e as raízes das plantas, havendo necessidade do meio líquido para que a absorção pelas raízes ocorra (Ruiz et al., 1999). A difusão é caracterizada pelo movimento térmico ao acaso de íons em direção à raiz, em virtude do gradiente de concentração gerado na superfície radicular (Barber, 1984), enquanto o fluxo de massa está associado ao gradiente de potencial hídrico provocado pela absorção de água pelas plantas (Silva et al., 1998).
O K é bastante móvel no solo e sua absorção é altamente seletiva (Chen & Gabelman, 2000). Embora não sejam conhecidas as funções desse nutriente, sabe-se que ele ativa mais de 60 enzimas. Suas principais funções estão ligadas a processos vitais, tais como: fotossíntese, translocação e balanço iônico (Marschner, 1995). A difusão é o principal mecanismo de transporte do K até a raiz; contudo, o fluxo de massa pode ter contribuição significativa no processo quando a concentração de K na solução do solo é elevada (Ruiz et al., 1999).
O algodoeiro é uma cultura bastante exigente e muito responsiva a K em termos de produtividade e qualidade de fibra, produzindo mais quando em solo de alta fertilidade, sob condições adequadas de umidade do solo, altas temperaturas e alta intensidade luminosa (Grespan & Zancanaro, 1999). Apesar disso, é uma cultura que apresenta uma taxa diária de consumo hídrico relativamente baixa (cerca de 6,5 mm dia-1, na fase de maior demanda transpiratória), mesmo em climas quentes e com suprimento adequado de água no solo. No caso de variedades precoces, o consumo hídrico durante o ciclo vegetativo não ultrapassa 450 mm (Azevedo et al., 1993). Em variedades modernas, no final do ciclo, é comum ocorrer deficiência de K. Normalmente, o final do ciclo do algodoeiro coincide com épocas de menor pluviosidade e, portanto, de menos água disponível no solo. Como a maior parte do K é transportada até à raiz por difusão, processo altamente dependente da água do solo, à medida que a planta desenvolve e o solo seca, diminui o teor do nutriente que pode ser difundido, justificando, possivelmente, a deficiência observada ao final do ciclo da planta.
O objetivo do presente trabalho foi avaliar a contribuição da difusão e do fluxo de massa no suprimento de K para a cultura do algodão de acordo com a água disponível e com a disponibilidade de K no solo.
MATERIAL E MÉTODOS
Realizou-se um experimento em condições de casa de vegetação, no Departamento de Produção Vegetal, FCA/Unesp, Botucatu, no período de outubro a dezembro de 2000. O solo utilizado foi um Latossolo Vermelho distroférrico típico (EMBRAPA, 1999), com 630 mg kg-1 de areia, 40 mg kg-1 de silte e 330 mg kg-1 de argila, com as seguintes características de fertilidade, determinadas segundo indicações de Raij & Quaggio (1983): pH em CaCl2 0,01 mol L-1 de 4,1; 18 g dm-3 de M.O.; 1 mg dm-3de Presina; 75 mmolc dm-3 de H + Al; 4 mmolc dm-3 de Ca2+; 1 mmolc dm-3 de Mg2+; 0,1 mmolc dm3 de K+; 5 mmolc dm-3 de soma de bases; CTC pH 7,0 de 80 mmolc dm-3 e 6% de V. Foi realizada calagem para elevação da saturação por bases para 60%, seguida de incubação úmida (aproximadamente 80% da capacidade de retenção de água do solo) por 15 dias. Após este período, foi realizada a adubação com N (50 mg dm-3) e P (150 mg dm-3), nas formas de sulfato de amônio e superfosfato simples. Nessa mesma época, foram aplicadas as doses de K que, juntamente com os conteúdos de água, constituíram os tratamentos.
O experimento foi instalado em esquema fatorial 2 x 4, com duas doses de K (15 e 121 mg dm-3), como cloreto de potássio, e quatro potenciais de água no solo (-0,03; -0,1; -0,5 e -1,0 MPa), correspondendo a 108, 92, 81 e 77 g kg-1 de água no solo, com quatro repetições, arranjadas em blocos casualizados. Utilizaram-se vasos de polietileno com capacidade para 5 kg de solo. Em duas repetições de cada tratamento, foram instaladas cápsulas porosas de porcelana para extração de solução do solo. As cápsulas tinham 100 mm de comprimento e 20 mm de diâmetro externo e foram instaladas, verticalmente, a partir do fundo do vaso.
Semeou-se o algodão (Gossypium hirsutum cv. Delta Opal), colocando-se cinco sementes por vaso. Dez dias após a emergência das plântulas, foi realizado o desbaste, deixando-se duas plantas por vaso, quando foram estabelecidos os conteúdos de água. O controle da umidade foi feito por meio de pesagens diárias e reposição da água evapotranspirada. O tempo para que cada potencial fosse atingido variou conforme a temperatura no período. Da semeadura até 18 dias após a emergência, a umidade do solo nos vasos foi corrigida a -0,03 MPa, a cada dois dias, quando os tratamentos passaram a ser considerados. Além dos vasos com plantas, foram preparados dois vasos, sem plantas, para cada conteúdo de água, para monitorar a perda de água e estimar a evaporação.
A extração da solução do solo foi realizada em três épocas ao longo do experimento, em intervalos de 15 dias, sendo a primeira aos 33 dias da emergência das plantas. As coletas de solução do solo, entre 50 e 100 mL, foram realizadas com auxílio de uma bomba a vácuo, na manhã seguinte à elevação da umidade do solo a -0,03 MPa. A solução foi filtrada, acondicionada em frascos de vidros e armazenada sob refrigeração. Em cada coleta o K foi determinado por leitura direta em ICP (Induced Coupled Plasma).
As plantas foram colhidas aos 63 dias da emergência, divididas em parte aérea e raízes e secas em estufa de circulação forçada a 60 °C, onde permaneceram até atingir peso constante. Após a secagem, o material foi pesado, moído e submetido à análise para determinação dos teores de K, segundo Malavolta et al. (1997). Antes da secagem, foi tomada uma amostra de aproximadamente 30% do sistema radicular, para determinar o comprimento e o diâmetro, utilizando-se o software Winrhizo, versão 3.8.
Para calcular a quantidade de água transpirada, subtraiu-se a quantidade evaporada do volume total aplicado em cada vaso. O K absorvido, suprido por fluxo de massa, foi obtido, multiplicando-se a concentração do K na solução do solo (média entre duas extrações sucessivas) pelo volume de água transpirado no período. A difusão foi calculada subtraindo-se o valor correspondente do K transportado por fluxo de massa do K total acumulado na planta (Becker & Maures, 1986; Barber, 1894).
Os resultados foram submetidos à análise de variância, utilizando o teste de Tukey (p < 0,05). Quando apropriado, foram ajustadas curvas de regressão, escolhendo-se a equação matemática de melhor ajuste aos dados obtidos.
RESULTADOS E DISCUSSÃO
Embora Rossetto et al. (1995) tenham observado maior volume de raízes de soja em solos mais pobres em K, no presente trabalho, não houve efeito significativo da disponibilidade do nutriente no comprimento ou diâmetro radicular. Os conteúdos de água também não tiveram efeito no comprimento (média de 27,1 m por planta) ou diâmetro (média de 0,48 mm) radicular do algodoeiro.
Não foi observado efeito (p < 0,05) dos conteúdos de água no solo sobre a produção de matéria seca. Entretanto, a matéria seca das plantas que receberam maior dose de K (10,5 g em média) foi maior que as com menor disponibilidade do nutriente (8,9 g em média).
Não houve modificação dos teores de K na matéria seca da parte aérea do algodoeiro com a redução na disponibilidade de água no solo. Na dose de 121 mg dm-3, o teor médio foi de 26 g kg-1, enquanto no menor nível de K o teor médio foi menor (14 g kg-1). Os teores de K na matéria seca de raiz (Figura 1a) aumentaram até -0,5 MPa com 121 mg dm3 de K no solo e até -1,0 MPa com 15 mg dm-3 de K no solo. Na presença de maior dose de K, os teores do nutriente sempre foram maiores.

Os conteúdos de água não influenciaram o conteúdo de K na parte aérea do algodoeiro, embora o conteúdo na dose de 121 mg dm-3 (116,5 mg planta-1) tenha sido maior que aquele obtido na dose de 15 mg dm-3 (68 mg planta-1), o que é justificado pelo aumento na concentração do nutriente na solução do solo. É possível que este comportamento seja devido à baixa taxa de consumo hídrico do algodoeiro, mesmo com suprimento adequado de água no solo, conforme sugeriram Azevedo et al. (1993). A menor dose de K aplicada no solo proporcionou acúmulo de K na raiz com a redução na disponibilidade de água (Figura 1b), evidenciando, provavelmente, um efeito de concentração, uma vez que não houve efeito na produção de matéria seca.
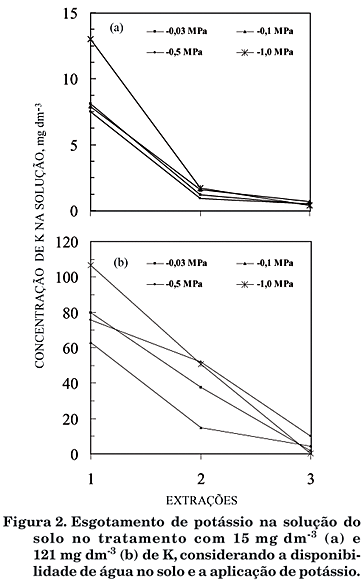
A concentração de K na solução do solo caiu expressivamente a cada extração sucessiva, para todos os conteúdos de água (Figura 2ab). O fenômeno deveu-se, pelo menos em parte, à demanda inicial da cultura pelo nutriente. Outra explicação seria que o K não permanece durante muito tempo em solução, tendendo a ficar adsorvido nos colóides do solo e nos resíduos de cargas da parede e na membrana celular das plantas. De acordo com Rosolem et al. (1993), as trocas entre K trocável e K solução são rápidas, de maneira que o K trocável é prontamente disponível às plantas. Desta forma, para a dose de 15 mg dm-3, observou-se um esgotamento de K na solução do solo antecipadamente ao esgotamento que ocorreu na maior dose de K. Além disso, os tratamentos com menores potenciais de água (-0,5 e -1,0 MPa) apresentaram maiores teores do nutriente. Assim, o efeito da água na concentração de K na solução do solo é mais importante em situação de alta disponibilidade do nutriente. O método utilizado no trabalho leva a um decréscimo momentâneo no K solução que, no entanto, representa pequena parte do K disponível. Considerando que, nos cálculos, foi considerada a média entre duas amostragens e considerando ainda que a quantidade de K extraído do solo representa menos de 3% do total absorvido pela planta, este pode ser considerado um erro desprezível.
Quanto aos mecanismos de suprimento de K às raízes, não foi observado efeito dos conteúdos de água na menor dose de K (Figura 3ab). Contudo, com maior disponibilidade do nutriente, houve um decréscimo no suprimento por difusão (Figura 3a), à medida que o solo estava mais seco, acompanhado de um aumento da contribuição do fluxo de massa (Figura 3b). Isso deve ter ocorrido em virtude do aumento da concentração de K na solução do solo (Figura 2ab). Este é um aspecto importante que atesta não só a relevância da concentração do nutriente na solução do solo para o suprimento à raiz (Marshner, 1995), mas também sua dependência do potencial matricial. A baixa disponibilidade de água no solo pode afetar a absorção de K em decorrência da restrição do crescimento radicular e da capacidade de absorção da raiz, assim como da menor mobilidade do K no meio (Seiffert et al., 1995). Ademais, a taxa de crescimento radicular é tida como o fator que mais influencia a absorção de K (Silberbush & Barber, 1983), de modo que, no caso de restrição ao crescimento radicular, a contribuição do fluxo de massa para o suprimento de K passaria a ser bastante representativa. Entretanto, no presente experimento, não foi observado efeito na morfologia radicular, levando à inferência de que a modificação na importância dos mecanismos de contato tenha sido uma conseqüência de modificações na cinética de absorção (Epstein, 1976).
É importante lembrar que, no período do experimento, a exigência da planta em água foi relativamente baixa (Grimes & El-Zick, 1990). Como conseqüência deste fato, não foi observada diferença na quantidade de água transpirada, seja em função da disponibilidade de K, seja de água (média de 2.730 cm3 por planta).
Considerando a equação da difusão de K no solo, a adição do nutriente eleva sua concentração na solução, aumentando o gradiente de concentração (dC/dx, em que C é concentração e x distância), aumentando a difusão (Ching & Barber, 1979). Por outro lado, a diminuição da água disponível levaria a uma diminuição na difusão. Maior conteúdo de água no solo favorece a difusão pelo seu efeito direto sobre o coeficiente de difusão e por reduzir a tortuosidade do caminho difusivo, aumentando, também, o valor do coeficiente de difusão (Barber, 1984). Uma redução do conteúdo de água no solo de 0,35 cm3 cm-3 para 0,3 cm3 cm-3 faz a difusão decrescer de 1,45.10-11 para 8,65.10-12 moles de K cm-2 s-2 (Hylander et al., 1999).
Outro fato a ser considerado é a distância média entre as raízes. Aceita-se que, quando a distância média entre as raízes é menor que 4 mm, pode haver competição por K (Yamaguchi & Tanaka, 1990). No presente caso, a distância média entre raízes, calculada como 1/(p L)1/2, sendo L densidade de comprimento, em cm cm-3 (Baldwin et al., 1973), foi sempre menor que 4,8 mm, permitindo a inferência de que, no caso, ocorreu competição pelo K difundido entre raízes vizinhas. A proporção do K transportado para as raízes por difusão decresce com o esgotamento do nutriente na rizosfera (Hylander et al., 1999), de modo que uma grande concentração radicular leva a esgotamento mais rápido do K solo na vizinhança das raízes (Seiffert et al., 1995). Outros trabalhos têm abordado a importância da taxa de crescimento da raiz, do comprimento radicular e da área superficial na absorção de K pelas plantas (Chen & Gabelman, 2000), mas, no presente caso, não houve modificação dessas características.
Assim, com a menor dose de K, a disponibilidade era tão baixa (Figura 2a) que limitou a absorção em grau maior que a disponibilidade de água. Quando a umidade não foi limitante, sem grande modificação na taxa transpiratória, a maior concentração de K na solução do solo mais seco proporcionou maior transporte do nutriente às raízes, fato também observado por Hylander et al. (1999).
Essa inferência foi confirmada pelos dados apresentados na figura 4. Observa-se que a difusão contribuiu, em média, com 96% do suprimento de K para as raízes de algodão, contra aproximadamente 3,5% do fluxo de massa, quando havia menos K no solo (Figura 4a). No caso de maior teor de K disponível (Figura 4b), a média da difusão caiu para cerca de 80%, e a contribuição de fluxo de massa passou de 3,5 para 19%. Por outro lado, a menor disponibilidade de água, aliada à grande concentração radicular, levou ao menor transporte do K por difusão. Os resultados para fluxo de massa estão próximos aos obtidos por Rosolem et al. (2001), quando observaram contribuição de 23,7% com uma dose de 60 mg kg-1 de K.

Barber (1984), estudando plantas de milho, observou que a difusão, em solo rico em K, contribuiu em média com cerca de 80% da quantidade do nutriente absorvido pela planta, o que mostra que, embora o algodão e o milho tenham sistemas radiculares muito diferentes, isso não altera a importância relativa dos mecanismos de transporte do K no solo. Em ambas as concentrações de K na solução do solo, a difusão foi o processo dominante no suprimento deste nutriente para as raízes, resultados condizentes com os obtidos em outros trabalhos (Ruiz et al., 1999).
CONCLUSÕES
1. A difusão foi o principal mecanismo de transporte do K no solo, contribuindo com 72 a 96% do total absorvido pelo algodoeiro.
2. A maior disponibilidade de K na solução do solo proporcionou aumento da contribuição da difusão para o transporte de K às raízes do algodoeiro.
3. A influência da água do solo nos mecanismos de transporte do K foi maior em solos que continham maiores concentrações de K em solução, tendo o fluxo de massa crescido em importância no caso de solo mais seco.
LITERATURA CITADA
Recebido para publicação em agosto de 2002 e aprovado em dezembro de 2003.
- AZEVEDO, P.V.; RAO T.V.R.; AMORIM NETO, M.S.; PEREIRA, J.R.C.; ESPÍDOLA SOBRINHO, J. & MACIEL, G.F. Necessidades hídricas da cultura do algodoeiro. Pesq. Agropec. Bras., 28:863-870, 1993.
- BALDWIN, J.P.; NYE, P.H. & TINKER, P.B. Uptake of solutes by multiple root and systems from soil. III. A model for calculating the solute systems developing in a finite volume of soil. Plant Soil, 38:621-35, 1973.
- BARBER, S.A. Soil nutrient biovailability: a mechanistic approach. New York, John & Wiley, 1984. 398p.
- BECKER, M. & MAURER, E.J. Morfologia de raízes, suprimento e influxo de potássio em plantas de milho. R. Bras. Ci. Solo, 10:259-263, 1986.
- CHEN, J. & GABELMAN, W.H. Morphological and physiological characteristics of tomato roots associated with potassium-acquisition efficiency. Sci. Hortic., 83:213-225, 2000.
- CHING, P.C. & BARBER, S.A. Evaluation of temperature effects on potassium uptake by corn. Agron. J., 71:1040-1044, 1979.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Centro de Pesquisa de Solos (Rio de Janeiro, RJ). Sistema Brasileiro de Classificação de Solos. Rio de Janeiro, 1999. 412p.
- EPSTEIN, E. Kinetics of ion transport and the carrier concept. In: LÜTTGE, U. & PITMAN, M.G., eds. Transport in plants. Encyclopedia of plant physiology. New York, Berlin, Springer-Verlag, 1976. p.70-94.
- GRESPAN, S.L. & ZANCANARO, L. Nutrição e adubação do algodoeiro no Mato Grosso. Rondonópolis, FMT/EMBRAPA, 1999. p.95 (Boletim, 3)
- GRIMES, D.W. & EL-ZICK, K.M. Cotton. In: STEWART, B.A. & NIELSEN, D.R., eds. Irrigation of agricultural crops. Madison, American Society of Agronomy, 1990. p.712-773.
- HYLANDER, L.D.; AE, N.; HATTA, T. & SUGIYAMA, M. Exploitation of K near roots of cotton, maize, upland rice and soybean grown in an Ultisol. Plant Soil, 208:33-41, 1999.
- MALAVOLTA, E.; VITTI, G.C. & OLIVEIRA, S.A. Avaliação do estado nutricional das plantas: princípios e aplicações. Piracicaba, POTAFOS, 1997. 319p.
- MARSCHNER, H. Mineral nutrition of higher plants. 2.ed. Londres, Academic Press, 1995. p.347-364.
- RAIJ, B. van & QUAGGIO, J.A. Métodos de análises de solo para fins de fertilidade. Campinas, Instituto Agronômico de Campinas, 1983. 31p. (Boletim Técnico, 81)
- ROSOLEM, C.A.; BESSA, A.M. & PEREIRA, H.F.M. Dinâmica do potássio no solo e nutrição potássica da soja. Pesq. Agropec. Bras., 28:1045-1054, 1993.
- ROSOLEM, C.A.; ESTEVES, J.A.F. & SILVA, R.H. Significance of mass flow and diffusion in supplying K to cotton roots as affected by limiting and K rates. In: INTERNATIONAL SYMPOSYUM ON PLANT-SOIL INTERATIONS AT LOW pH, 5., 2001, Abstracts. South Africa, Kualazulu-Natal, Departament of Agriculture and Environmental Affairs, 2001. p.48.
- ROSSETTO, C.A.V.; FERNANDES, D.M.; ISHIMURA, I. & ROSOLEM, C.A. Diferentes respostas de cultivares de soja ao potássio. Pesq. Agropec. Bras., 38:1225-1231, 1995.
- RUIZ, H.A.; MIRANDA, J. & CONCEIÇÃO, J.C.S. Contribuição dos mecanismos de fluxo de massa e de difusão para o suprimento de K, Ca, Mg às plantas de arroz. R. Bras. Ci. Solo, 23:1015-1018, 1999.
- SEIFFERT, S.; KASELOWSKY, J.; JUNGK, A. & CLAASSEN, N. Observed and calculated potassium uptake by maize as affected by soil water content and bulk density. Agron. J., 87:1070-1077, 1995.
- SILBERBUSH, M. & BARBER, S.A. Sensitivity analysis of parameters used in simulating uptake with a mechanistic mathematic model. Agron. J., 75:851-854, 1983.
- SILVA, D.J.; ALVAREZ, V.H. & RUIZ, H.A. Fluxo de massa e difusão de enxofre para raízes de milho em solos ácidos de Minas Gerais. R. Bras. Ci. Solo, 22:109-114, 1998.
- YAMAGUCHI, J. & TANAKA, A. Quantitative observation on the root systems of various crops growing in the field. Soil Sci. Plant Nutr., 36:483-493, 1990.
Datas de Publicação
-
Publicação nesta coleção
30 Jul 2004 -
Data do Fascículo
Jun 2004
Histórico
-
Recebido
Ago 2002 -
Aceito
Dez 2003