Resumos
O presente estudo objetivou avaliar o impacto da queima tradicional de campo nativo e do monocultivo de Pinus sp. em Lages (SC), no CO total do solo (COT), carbono da biomassa microbiana (CBM), respiração basal (C-CO2), quociente metabólico microbiano (qCO2) e relação CBM:COT. Foram selecionadas quatro áreas representativas da região, de 0,5 ha cada, sendo: (a) Campo nativo, sem queima nos últimos 50 anos (CN); (b) Campo nativo submetido à queima tradicional (CNQ), ambas, pastagens naturais; (c) Mata natural com predominância de Araucaria angustifolia (MATA); e (d) Reflorestamento de Pinus taeda com oito anos (PINUS). Para tanto, coletaram-se seis amostras, compostas de nove subamostras de solo em cada área, na profundidade de 0-5 cm, em dezembro de 2002. Para avaliar o CBM, foi utilizado o método da fumigação-extração. O C-CO2 foi determinado em laboratório. Os maiores valores de liberação de C-CO2 foram encontrados na MATA e no PINUS, seguidos de CNQ e CN, respectivamente. Os maiores valores de CBM, COT e relação CBM:COT foram encontrados na MATA, não tendo as demais áreas estudadas apresentado diferenças entre si. O qCO2 foi maior no PINUS, seguido de CNQ, em comparação com MATA e CN. A análise multivariada mostrou ser uma ferramenta auxiliar importante ao discriminar o CBM como sendo o atributo que mais contribuiu na separação entre as áreas estudadas.
bioindicador de qualidade; CO total; C-biomassa; quociente metabólico
The objective of this study was to evaluate the impact of burning native pastures and of cultivating Pinus sp in monoculture in Lages, State of Santa Catarina, Brazil. Total organic carbon (TOC), microbial biomass carbon (MBC), microbial basal respiration (C-CO2), the microbial metabolic quotient (qCO2), and the ratio MBC/TOC were analyzed. Four representative areas of the region with 0.5 ha each were selected: (a) native pasture without burning for at least 50 years (NP); (b) native pasture burnt off every year (NPF); (c) natural forest (NF) with predominance of Araucaria angustifolia and (d) eight-year-old Pinus taeda monoculture (PINUS). Data were obtained from six replicate samples composed of nine sub-samples each, from a depth of 0-5 cm, in the four studied areas. The fumigation-extraction method was used for the evaluation of MBC. C-CO2 evolution was obtained under laboratory conditions. The highest C-CO2 values were detected in NF and PINUS, followed by NPF and NP, respectively. The highest values of MBC, TOC and MBC:TOC were detected in NF, but the other areas did not present significant differences among each other. The qCO2 rate was higher in PINUS followed by NPF in comparison with NF and NP. Multivariate analysis was an important auxiliary tool for suggesting MBC as an attribute to discriminate the studied areas.
soil quality bioindicator; total organic carbon; soil microbial biomass carbon; metabolic quotient
SEÇÃO III - BIOLOGIA DO SOLO
Efeito do monocultivo de Pinus e da queima do campo nativo em atributos biológicos do solo no Planalto Sul Catarinense1 1 Parte da Tese de Mestrado do primeiro autor, apresentada ao Programa de Pós-graduação em Ciência do Solo, Centro de Ciências Agroveterinárias (CAV), Universidade do Estado de Santa Catarina - UDESC. Trabalho apresentado no XXIX Congresso Brasileiro de Ciência do Solo, Ribeirão Preto (SP), 13 a 18 de julho de 2003. Executado com recursos da CAPES/UDESC.
Effects of native pasture burning and Pinus monoculture on changes in soil biological attributes on the Southern Plateau of Santa Catarina - Brazil
Dilmar BarettaI; Julio Cesar Pires SantosII; Samuel Ribeiro FigueiredoIII; Osmar Klauberg-FilhoII
IPós-Graduando em Solos e Nutrição de Plantas, Escola Superior de Agricultura "Luiz de Queiroz", Universidade de São Paulo - USP/ESALQ. Av. Pádua Dias 11, Caixa Postal 9, CEP 13418-900 Piracicaba (SP). Bolsista do CNPq. E-mail: baretta@esalq.usp.br
IIProfessor do Departamento de Solos, Universidade do Estado de Santa Catarina - UDESC. Caixa Postal 281, CEP 88520-000 Lages (SC). E-mails: a2jcps@cav.udesc.br; a2okf@cav.udesc.br
IIIPós-Graduando em Ciência do Solo, Universidade Federal do Rio Grande do Sul - UFRGS. Caixa Postal 776, CEP 90001-970 Porto Alegre (RS). Bolsista do CNPq. E-mail: s.r.figueiredo@bol.com.br
RESUMO
O presente estudo objetivou avaliar o impacto da queima tradicional de campo nativo e do monocultivo de Pinus sp. em Lages (SC), no CO total do solo (COT), carbono da biomassa microbiana (CBM), respiração basal (C-CO2), quociente metabólico microbiano (qCO2) e relação CBM:COT. Foram selecionadas quatro áreas representativas da região, de 0,5 ha cada, sendo: (a) Campo nativo, sem queima nos últimos 50 anos (CN); (b) Campo nativo submetido à queima tradicional (CNQ), ambas, pastagens naturais; (c) Mata natural com predominância de Araucaria angustifolia (MATA); e (d) Reflorestamento de Pinus taeda com oito anos (PINUS). Para tanto, coletaram-se seis amostras, compostas de nove subamostras de solo em cada área, na profundidade de 0-5 cm, em dezembro de 2002. Para avaliar o CBM, foi utilizado o método da fumigação-extração. O C-CO2 foi determinado em laboratório. Os maiores valores de liberação de C-CO2 foram encontrados na MATA e no PINUS, seguidos de CNQ e CN, respectivamente. Os maiores valores de CBM, COT e relação CBM:COT foram encontrados na MATA, não tendo as demais áreas estudadas apresentado diferenças entre si. O qCO2 foi maior no PINUS, seguido de CNQ, em comparação com MATA e CN. A análise multivariada mostrou ser uma ferramenta auxiliar importante ao discriminar o CBM como sendo o atributo que mais contribuiu na separação entre as áreas estudadas.
Termos de indexação: bioindicador de qualidade, CO total, C-biomassa, quociente metabólico.
SUMMARY
The objective of this study was to evaluate the impact of burning native pastures and of cultivating Pinus sp in monoculture in Lages, State of Santa Catarina, Brazil. Total organic carbon (TOC), microbial biomass carbon (MBC), microbial basal respiration (C-CO2), the microbial metabolic quotient (qCO2), and the ratio MBC/TOC were analyzed. Four representative areas of the region with 0.5 ha each were selected: (a) native pasture without burning for at least 50 years (NP); (b) native pasture burnt off every year (NPF); (c) natural forest (NF) with predominance of Araucaria angustifolia and (d) eight-year-old Pinus taeda monoculture (PINUS). Data were obtained from six replicate samples composed of nine sub-samples each, from a depth of 0-5 cm, in the four studied areas. The fumigation-extraction method was used for the evaluation of MBC. C-CO2 evolution was obtained under laboratory conditions. The highest C-CO2 values were detected in NF and PINUS, followed by NPF and NP, respectively. The highest values of MBC, TOC and MBC:TOC were detected in NF, but the other areas did not present significant differences among each other. The qCO2 rate was higher in PINUS followed by NPF in comparison with NF and NP. Multivariate analysis was an important auxiliary tool for suggesting MBC as an attribute to discriminate the studied areas.
Index terms: soil quality bioindicator, total organic carbon, soil microbial biomass carbon, metabolic quotient.
INTRODUÇÃO
Na região serrana, mais precisamente no Planalto Sul Catarinense, práticas como a retirada da mata nativa de Araucaria sp para a introdução de espécies exóticas, principalmente o Pinus sp, bem como a queima do campo nativo como prática tradicional de manejo para a pecuária extensiva, estão em franco crescimento e são objeto de alguns estudos da Ciência do Solo, por acreditar-se que alteram significativamente os atributos químicos (Heringer & Jacques, 2002; Jacques, 2002; Rheinheimer et al., 2003) e biológicos do solo (Fernandes, 1997).
O monocultivo de pinus proporciona um ambiente uniforme, modifica a diversidade de substrato para a biota do solo, alterando, por sua vez, alguns atributos biológicos, como, por exemplo, C-biomassa e respiração microbiana do solo, além de alterar a densidade e diversidade da fauna edáfica (Baretta et al., 2003).
A queima de campos no sul do País aparece como prática de manejo tradicional das pastagens, sendo amplamente difundida em Santa Catarina e no Rio Grande do Sul (Rheinheimer et al., 2003), principalmente em razão da topografia acidentada e do afloramento de rochas, que dificultam o manejo do pasto acumulado durante a época da seca (final do inverno). A temperatura no interior do solo não se eleva muito durante a queima tradicional, mas a perda de água por evaporação pode ser muito elevada (Debano et al., 1979; Heringer & Jacques, 2002), afetando a camada superficial (até 2 cm). Além disso, acelera a mineralização da matéria orgânica do solo, libera nutrientes, como N e P, para a fase líquida do solo, deixando o N susceptível a perdas por lixiviação e volatilização (Mroz et al., 1980), elimina a cobertura vegetal (Bertol et al., 1997; Heringer & Jacques, 2002) e facilita as perdas de solo e nutrientes por erosão hídrica (Bertol et al., 1997). No entanto, é importante conscientizar os pecuaristas do grande desafio dos trabalhos nesta área (Rheinheimer et al., 2003), para, posteriormente, montar um modelo qualitativo e uma possível proposta de manejo sustentável das pastagens da região (Fernandes, 1997).
A perturbação antrópica de qualquer sistema estável (solo + cobertura vegetal) tende a causar mais perdas do que ganhos de C, acarretando perda da qualidade do solo ao longo do tempo. Tais perdas são devidas à liberação de CO2 na respiração, provenientes da decomposição da matéria orgânica do solo por hidrólise microbiana, da lixiviação e perdas de compostos por erosão hídrica, sendo estas duas últimas vias de magnitude menor em solos subtropicais (Bayer, 1999).
A biomassa microbiana constitui a maior fração ativa da matéria orgânica do solo e, por isso, é sensível em inferir mudanças nos níveis de matéria orgânica, causadas pelo manejo de plantas, de adubação (Marchiori Júnior & Melo, 2000) e práticas de cultivo (Gama-Rodrigues, 1999; Pascual et al., 2001) e pelo sistema agrícola adotado (Balota et al., 1998). Assim, o estudo da biomassa microbiana e a sua atividade podem servir como indicadores de qualidade do solo (Doran & Parkin, 1994; Kaiser et al., 1995; Daniel et al., 1999), auxiliando na avaliação e no estabelecimento de um novo equilíbrio biodinâmico do solo, além de fornecer subsídios para o planejamento do uso da terra (D'Andréa et al., 2002).
O presente estudo teve o objetivo de avaliar o impacto da queima tradicional de campo nativo e do monocultivo de pinus sobre a qualidade do solo, por meio de seus efeitos nos teores de CO total do solo (COT), carbono da biomassa microbiana (CBM), respiração basal (C-CO2) e relação CBM:COT, em comparação com áreas de campo nativo não submetido à queima e de mata de araucária.
MATERIAL E MÉTODOS
O estudo foi realizado na localidade de Coxilha Rica, município de Lages (SC), que se localiza a 27 ° 49 ' latitude sul e 50 ° 20 ' longitude oeste, com altitude média de 935 m acima do nível do mar, na região do Planalto Sul Catarinense. O clima da região é mesotérmico úmido com inverno chuvoso e verão ameno (Cfb, conforme a classificação de Köppen). A precipitação pluvial média anual é de 1.600 mm, com 123 a 140 dias no ano, com chuvas bem distribuídas ao longo do ano, e temperatura média anual variando de 3,8 ºC a 15,8 °C (Figura 1).
Para este estudo, foram selecionadas quatro áreas representativas da região, localizadas em altitudes e posições semelhantes de relevo, sendo caracterizadas, a seguir: (a) Campo nativo (CN); (b) Campo nativo submetido à queima (CNQ), (c) Monocultivo de Pinus taeda (PINUS) e (d) Mata natural com predominância de Araucaria angustifolia (MATA).
Os campos, segundo o zoneamento de Gomes (1990), são pastagens naturais do tipo "palha fina", compostas principalmente por espécies de ciclo estival, com predominância de Paspalum notatum e Paspalum plicatulum, exploradas basicamente por animais bovinos, comportando as maiores lotações no verão (máximo uma unidade animal ha-1) e as menores no inverno (0,2-0,5 unidade animal ha-1), graças ao efeito de sazonalidade de produção da pastagem. O CN não sofreu queimadas nos últimos 50 anos (situação difícil de encontrar na região) e o CNQ foi submetido à queima tradicional na região, realizada no final do mês de agosto de 2002. O PINUS, por sua vez, encontra-se com oito anos de idade, e, assim como na MATA, houve entrada de bovinos durante o ano inteiro, comportando uma lotação média de 0,3 unidade animal ha-1. O solo é classificado como Latossolo Bruno (Embrapa, 1999), profundo, alto teor de matéria orgânica e textura argilosa (Quadro 1).
Para análise da biomassa microbiana e sua atividade (respiração basal), no dia 10 de dezembro de 2002 (100 dias após a queima do CN), foram coletadas seis amostras, compostas de nove subamostras, em uma área de 0,5 ha, na profundidade de 0-5 cm. As amostras foram acondicionadas em sacos plásticos e transportadas em caixas de isopor com gelo para o laboratório de Microbiologia do Solo, no Centro de Ciências Agroveterinárias (CAV), onde foram mantidas em geladeira até serem analisadas. Para as análises, as amostras foram homogeneizadas e passadas em peneira com malha de 2 mm, uma subamostra foi retirada para determinação da umidade, e as seguintes características químicas determinadas: pH em água, índice SMP, teores de P, de Al, de Ca e Mg trocáveis, conforme método descrito em Tedesco et al. (1995).
Para avaliar o C da biomassa microbiana (CBM) das amostras de solo, foi utilizado o método de fumigação-extração (Vance et al., 1987). O CO total (COT) do solo foi determinado por oxidação úmida com dicromato de K (K2Cr2O4 0,75 mol L-1) e ácido sulfúrico (H2SO4 0,5 mol L-1), conforme descrito por Tedesco et al. (1995). A respiração basal (C-CO2) foi avaliada por meio da determinação da respiração microbiana em laboratório (Jäggi, 1976). A partir dos resultados de CBM e COT, foi calculada a relação entre as duas características, expressa como a percentagem de C microbiano em relação ao C total do solo (Anderson, 1994). Utilizaram-se os resultados de C-CO2 e do CBM para calcular o quociente metabólico (qCO2), que representa a quantidade de C-CO2 liberada em determinado tempo, por unidade de C microbiano (µg C-CO2 h-1/µg C-biomassa g-1 solo seco).
Os dados foram submetidos à análise de variância (ANOVA) pelo programa SAEG (Universidade Federal de Viçosa), e as médias foram comparadas pelo teste de Duncan (P < 0,05). Adicionalmente, realizou-se uma análise multivariada dos dados, utilizando-se o programa SAS versão 6.12 (SAS, 1990), por meio da Análise Canônica Discriminante (ACD), para identificar as características mais relevantes na discriminação das diferentes áreas amostradas. Este tipo de análise também permite separar as áreas de amostragem com base em funções canônicas discriminantes. Quando verificada diferença significativa entre as áreas estudadas, por meio da ACD, foi realizado teste de comparação de médias nos valores dos coeficientes canônicos homogeneizados (CCH), nas distintas funções canônicas, por meio do teste de LSD (P < 0,05), conforme Cruz-Castilho et al. (1994). Também foram efetuadas análises de correlação simples de Pearson para COT, CBM, C-CO2 e qCO2.
RESULTADOS E DISCUSSÃO
Carbono orgânico total (COT) e carbono da biomassa microbiana (CBM)
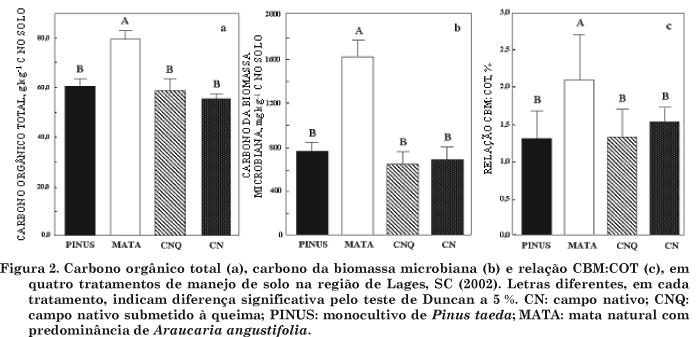
O maior teor de COT e CBM foi observado na mata (MATA), 80 g kg-1 de C no solo e 1.600 mg kg-1 de C no solo, respectivamente (Figura 2a e 2b). O COT na mata de araucária foi 40 % maior que o observado nas demais áreas estudadas, enquanto o CBM foi 140 % maior. Fialho et al. (1991), analisando química e fisicamente as propriedades de um Latossolo Vermelho-Amarelo sob mata natural, pastagem e eucalipto com cerca de dezoito anos de idade, no município de Viçosa (MG), encontraram comportamento semelhante do COT sob mata, sendo as quantidades de C no solo maiores na mata do que na pastagem e no eucalipto, até à profundidade de 0-8 cm. Marchiori Júnior & Mello (2000), trabalhando em um Latossolo Roxo, nos tratamentos mata natural (condição natural de cobertura do solo), café até 1994; mata natural até 1976, café até 1991 e milho até 1994; mata até 1940, café até 1960, citrus até 1978, e cana-de-açúcar até 1994, encontraram na mata natural maiores valores de C orgânico no solo (37,2 g kg-1 de C) que nos demais tratamentos estudados.
Os valores de COT e CBM para campo nativo (CN), campo nativo queimado (CNQ) e monocultivo de pinus (PINUS) não diferiram entre si (Figura 2a e 2b). Por outro lado, Cattelan & Vidor (1990), estudando as flutuações da biomassa, atividade e população microbiana do solo submetido a sete sistemas de cultivo em Eldorado do Sul (RS), encontraram na média de doze avaliações uma maior biomassa (C) no campo nativo (61,5 mg 100 g de C no solo), na profundidade de 0-5 cm. Esta maior biomassa microbiana na pastagem (campo nativo), encontrada por Cattelan & Vidor (1990), está de acordo com os dados obtidos por Geraldes et al. (1995), que encontraram maior biomassa microbiana, na profundidade de 0-10 cm, em pastagem de quatro anos na Amazônia (754 mg kg-1 de C no solo). D'Andréa et al. (2002), estudando os atributos biológicos indicadores de qualidade de um Latossolo Vermelho de Morrinhos (GO), encontraram maiores diferenças de CBM na camada superficial (0-10 cm), apresentando o cerrado nativo maiores valores de carbono microbiano (1.310 mg kg-1 de C), comparado com pastagem, plantio direto, plantio direto com histórico de gradagem superficial, plantio convencional de longa duração e plantio convencional após pastagem.
A prática da queima tradicional do campo nativo (CNQ) não alterou o COT e CBM, quando comparada com CN e PINUS (Figura 2a e 2b). Em contraposição a esses resultados, Cerri et al. (1985) observaram em um Latossolo Amarelo distrófico textura argilosa, sob condições de mata natural (Floresta da Amazônia), que o desmatamento e a queima da vegetação natural resultaram na redução de 66 % da biomassa microbiana inicial, que desapareceu totalmente nos primeiros 10 cm de profundidade do solo. A diminuição da população microbiana na camada subsuperficial do solo, em determinados sistemas, pode ser explicada, principalmente, pelas piores condições para o desenvolvimento de microrganismos, como, por exemplo, menor aeração do solo e menor disponibilidade de matéria orgânica facilmente decomponível (Luizão et al., 1991). A maior concentração e as diferenças na biomassa microbiana, encontradas na camada superficial do solo, também foram relatadas por Geraldes et al. (1995) e D'Andréa et al. (2002). A queima tradicional do campo nativo de curta duração pode causar inibição da biomassa microbiana e trazer problemas graves no ecossistema. Por outro lado, pode haver uma adaptação da microbiota, e produtos novos podem ser gerados pelas plantas, principalmente pelas raízes (Sauerbeck et al., 1982), e a biomassa reaparecer logo nos primeiros anos que seguem à queima.
Encontrou-se um alto índice de correlação de Pearson (r = 0,94, P < 0,05) entre CBM e COT, o que indica que o CBM acompanhou as mudanças na concentração da matéria orgânica e não foi capaz de discriminar possíveis diferenças nos ecossistemas, devidas a outros fatores, oriundos da atividade antrópica, como o manejo por fogo (CNQ) ou monocultivos de espécies florestais exóticas, como, por exemplo, o Pinus taeda (PINUS). Nesse sentido, Cattelan & Vidor (1990) encontraram uma relação positiva entre os teores de matéria orgânica e a biomassa microbiana em diferentes sistemas de cultura estudados, resultado de efeitos diretos e indiretos de variações na umidade e temperatura do solo, tornando, assim, o ambiente edáfico mais adequado para os microrganismos (Luizão et al., 1991). Assim sendo, em solos sob vegetação natural (MATA), geralmente não ocorrem variações nos conteúdos de matéria orgânica, ao longo do tempo, em razão, principalmente, da igualdade das quantidades de material orgânico adicionadas e perdidas, caracterizando um estado estacionário "steady state" (Addiscott, 1992).
Relação entre carbono da biomassa (CBM) e CO total (COT)
A contribuição do C da biomassa microbiana para o COT do solo variou de 1,3 a 2,2 %. A relação CBM:COT (Figura 2c) teve o mesmo comportamento da biomassa microbiana (Figura 2b), apresentando a mesma tendência, ou seja, a mata mostrou maior relação CBM:COT, com cerca de 2,2 %, seguida dos demais sistemas estudados, que não diferiram entre si. Essa relação tem sido reportada como medida da qualidade da matéria orgânica do solo (Wardle & Hungria, 1994), expressando a eficiência da biomassa microbiana em utilizar o C orgânico do solo. Também é considerada boa indicadora das alterações dos processos no solo, e aqueles que apresentarem valores maiores ou menores podem expressar ocorrência de acúmulo ou perda de C do solo, respectivamente. Na literatura, o valor 2,2 % é citado por Jenkinson & Ladd (1981) como sendo o nível no qual estaria ocorrendo equilíbrio. No entanto, essa relação pode sofrer influência do manejo do solo (Balota et al., 1998) e do grau de estabilização do CO (Sparling, 1992).
Respiração basal (C-CO2) e quociente metabólico microbiano (qCO2)
Com base nos resultados de C-CO2, verificou-se que as maiores perdas de C-CO2 por atividade microbiana ocorreram no PINUS e MATA seguidas de CNQ e CN (Figura 3b). Quando se considera a respiração (C-CO2) por unidade de C microbiano, percebe-se que a maior atividade de mineralização do C orgânico do solo ocorreu no sistema PINUS e CNQ, em que houve ação antrópica (Figura 3a). Solos sob interferência antrópica, como é o caso do PINUS e CNQ, apresentam mudanças na composição e atividades metabólicas específicas, considerando o estresse da população (Moreira & Siqueira, 2002).
Os sistemas de MATA e CN apresentaram os menores valores de qCO2 (6 e 5 µg C-CO2 µg CBM-1 h-1), respectivamente, enquanto PINUS (17 µg C-CO2 µg CBM-1 h-1) e CNQ (10 µg C-CO2 µg CBM-1 h-1) apresentaram os maiores valores de qCO2, sendo o do PINUS ainda maior que o do CNQ (Figura 3a). Maiores valores de qCO2 indicam maiores perdas de C no sistema na forma de CO2 por unidade de C microbiano, razão por que PINUS e CNQ apresentaram as maiores perdas possivelmente decorrentes de microrganismos de crescimento rápido. A MATA e CN, ao contrário, apresentaram menores valores de qCO2, sendo, portanto, sistemas mais conservativos (sustentáveis), com menores perdas de C e de maior sustentabilidade, no que se refere ao fluxo de C. Assim, o quociente metabólico permitiu discriminar os sistemas quanto à sustentabilidade de C no solo.
A melhoria temporária de alguns atributos químicos proporcionados pela queima, como, por exemplo, aumento de K, Ca2+ + Mg2+ e valores de pH e redução nos teores de Al trocável (Rheinheimer et al., 2003), permite que uma população de oportunistas se multiplique rapidamente, gerando desequilíbrio e, conseqüentemente, maiores perdas de C no sistema na forma de CO2. Com o passar do tempo, a queima resulta em maiores teores e saturação de Al3+ e maior acidez potencial, bem como em menores teores de Mg2+ na camada superficial do solo (Heringer et al., 2002; Jacques, 2002). Assim sendo, a prática da queima tradicional do campo nativo, nas condições estudadas, mostrou-se deletéria, visto que aumentou a taxa respiratória da microbiota, elevando as perdas de C na forma de C-CO2 (Figura 3a) em relação ao campo nativo não submetido à queima. Além deste efeito, outros têm sido relatados, como, por exemplo, aumentos da temperatura, diminuição da umidade (Cattelan & Vidor, 1990) e elevação do pH, impedindo que a biomassa microbiana utilize o CO, pelo menos, na camada superficial do solo (Cerri et al., 1985).
Embora os estoques totais de COT não tenham sido afetados entre PINUS, CNQ e CN (Figura 2a), aumentos nos valores de qCO2 no PINUS e CNQ (Figura 3a) são indicativos de alto requerimento energético desta população, e, conseqüentemente, maior taxa de decomposição da matéria orgânica do solo, o que poderá refletir-se em decréscimos futuros nos estoques e na qualidade da matéria orgânica do solo, comprometendo a ciclagem de nutrientes (Gama-Rodrigues, 1999).
Apesar de alguns pesquisadores questionarem o uso de qCO2 como bioindicador de distúrbios e estresse no meio, por não distinguir esses efeitos (Yan et al., 2003), a maioria dos estudos tem aplicado extensamente o qCO2 na avaliação de regime de cultivo (Anderson & Domsch, 1990), gradiente de poluição (Ohtonen, 1994), efeito da temperatura (Anderson & Gray, 1991) e ecossistemas de floresta (Anderson & Domsch, 1993).
Estudos aprofundados indicam que, em solos de florestas, o pH do solo também exerce uma função controladora sobre as taxas respiratórias da biomassa microbiana, afetando as relações qCO2 e CBM:COT (Anderson & Domsh, 1993; Anderson, 1999, 2003), razão pela qual devem ser estudados separadamente. Apesar de não se ter realizado análise de correlação, percebe-se que em pH baixo o qCO2 tende a ser maior (PINUS) que em pH mais elevado (MATA) (Quadro 1 e Figura 3a). PINUS e MATA, como têm pH do solo, respectivamente, de 4,4 e 5,6 (Quadro 1), tiveram um comportamento sobre a relação CBM:COT e qCO2 de aumento e redução, respectivamente (Figuras 2c e 3a). Nesse sentido, Anderson (1999) observou que o qCO2 se comporta inversamente proporcional, decorrente de um gradiente crescente de pH, tendo como referência quatro áreas de florestas nos Países Baixos. Assim sendo, as diferenças significativas na atividade microbiana entre os sistemas MATA e PINUS parecem fortemente relacionadas com o pH e com a condição ácida do solo (Anderson & Domsch, 1993). No monocultivo de Pinus taeda, entretanto, ainda há necessidade de maiores pesquisas neste sentido.
Enquanto o aumento do qCO2 no sistema PINUS tem como fator controlador o pH do solo, o incremento do qCO2 do CN para o CNQ, possivelmente, se deve, dentre outros fatores, à prática da queima tradicional desta pastagem (Figura 3a). Além disso, valores de qCO2 altos mostram que o manejo usado provoca degradação do solo, principalmente pela perda de matéria orgânica do solo, como demonstrado por Pascual et al. (2001). Assim, a queima tradicional do campo nativo pode proporcionar um estresse no sistema solo, estimular as populações de microrganismos de crescimento rápido e interromper bruscamente a sucessão de ecossistemas por ciclos naturais, conforme o modelo proposto por Odum (1969), gerando uma biomassa microbiana qualitativamente alterada e imatura e um solo longe de apresentar a desejada qualidade do solo.
Análise multivariada canônica discriminante (ACD)
A análise multivariada canônica discriminante (ACD) demonstrou na primeira e segunda função discriminante canônica (FDC1 e FDC2) uma alta correlação canônica, sendo as duas funções ajustáveis para explicar as variações encontradas para os valores de CBM, COT, C-CO2 e qCO2, nas áreas estudadas. A relação CBM:COT é uma relação de divisão direta e altamente influenciada pelos valores de CBM e, possivelmente, infringe um fenômeno tendencioso sobre a análise multivariada; dessa forma, esta relação não foi considerada na avaliação. Analisando os valores de "eigenvalues", observou-se que a FDC1 explicou 88 % da variação encontrada nas áreas estudadas, dentro dos atributos biológicos analisados, tendo a FDC2 explicado apenas 10 % da variação (Figura 4).
O teste estatístico multivariado de Wilk's Lambda indicou haver uma diferença altamente significativa (P < 0,0001) entre os quatro sistemas analisados. O teste de LSD a 5 % de significância da média de todos os coeficientes canônicos homogeneizados (CCH) mostrou dentro da FDC1 e FDC2 que todos os sistemas foram diferentes entre si (Quadro 2). Na FDC1, o sistema MATA obteve a maior média seguida de PINUS, CNQ e CN, enquanto, na FDC2, o sistema CN obteve a maior média, seguido dos sistemas MATA, CNQ e PINUS, respectivamente (Quadro 2).
Os valores do coeficiente de correlação (r) refletem informações univariadas (Quadro 3) e mostram a contribuição individual de cada atributo estudado na separação das áreas. O coeficiente canônico homogeneizado (CCH) apresenta valores relativos à contribuição conjunta das características para a distinção das áreas (Quadro 3). No entanto, costuma-se adotar o índice conhecido como coeficiente da razão paralela discriminante (CRPD) que leva em consideração os pesos dos dois índices anteriores, pois é um produto dos mesmos (CRPD = r x CCH).
O CBM apresentou dentro da FDC1 (Quadro 3) os maiores valores de r e CCH, apresentando, conseqüentemente, um maior CRPD, o que indica que esta característica responde, no somatório de todas as variáveis, por praticamente toda a separação entre as áreas avaliadas. Nesse sentido, a biomassa microbiana tem sido reportada freqüentemente como um indicador das mudanças que podem ocorrer ao longo do tempo, com relação a propriedades do agroecossistema (Kaiser et al., 1995). Por outro lado, em alguns trabalhos, os atributos biológicos recebem menos ênfase que as propriedades químicas e físicas, principalmente porque tais atributos são mais difíceis de medir e o método para sua determinação é mais oneroso (Parr et al., 1992).
O qCO2 apresentou valores de CCH semelhantes ao CBM dentro da FDC1 (Quadro 3); entretanto, com um r muito baixo (-0,0214), culminando em um CRPD insignificante na função indicada. Por outro lado, dentro da FDC2, os maiores valores de qCO2 e o C-CO2 foram encontrados no CRPD. O COT apresentou o maior valor de CCH dentro da FDC2.
A figura 4 apresenta as relações entre FDC1 e FDC2 das áreas sobre os coeficientes homogeneizados. Pode-se observar que a FDC1 separa os sistemas de campos nativos (CN e CNQ) dos sistemas florestais (MATA e PINUS), enquanto a FDC2 separa os sistemas naturais (MATA e CN) de sistemas manejados antropicamente (PINUS e CNQ). Apesar de a FDC2 explicar apenas 10 % da variação, discrimina os possíveis efeitos negativos que a prática da queima do campo nativo e o monocultivo de Pinus taeda possam exercer sobre a qualidade e sustentabilidade do C do solo. Desta forma, o CBM discriminou grupos gerais de sistemas, enquanto o C-CO2 e o qCO2 discriminaram interferências externas aos sistemas. De maneira geral, os atributos analisados mostraram-se eficientes para a separação entre as áreas (Figura 4).
Na última década, houve uma grande demanda por indicadores precisos, amplos e reproduzíveis, que detectem mudanças na qualidade do solo (Anderson, 2003). Ainda, segundo o mesmo autor, aspectos funcionais e de sustentabilidade da fertilidade do solo são amplamente governados pela atividade microbiana, isto, por sua vez, comprova a necessidade por indicadores microbiológicos de qualidade do solo. Os resultados obtidos mostram que devem ser avaliados todos os indicadores ecofisiológicos descritos no trabalho de Anderson (2003), de forma conjunta, e não separadamente, pois, desse modo, fornecem uma maior riqueza de informações para a tomada de decisões dentro da ecologia e sustentabilidade de ecossistemas complexos.
CONCLUSÕES
1. Os maiores valores de liberação de C-CO2 foram encontrados na MATA e no PINUS, seguidos de CNQ e CN, respectivamente.
2. Os maiores valores de CBM, COT e relação CBM:COT foram encontrados na MATA, não apresentando diferenças as demais áreas entre si.
3. O qCO2 foi maior no PINUS, seguido de CNQ, em comparação com MATA e CN.
4. A análise multivariada mostrou ser uma ferramenta auxiliar importante ao discriminar o CBM como sendo o atributo que mais contribuiu para a separação entre as áreas avaliadas.
AGRADECIMENTOS
Dilmar Baretta e Samuel Ribeiro Figueiredo agradecem à CAPES e ao CNPq, respectivamente, pela bolsa de estudos; ao PAP/UDESC, pelo financiamento do projeto; ao Professor do CAV, Ph.D. Cassandro Vidal Talamini do Amarante, pelo auxílio na análise estatística multivariada; à Professora da ESALQ, Dra. Elke J.B.N. Cardoso, pelas sugestões no manuscrito, e aos proprietários das áreas utilizadas.
LITERATURA CITADA
Recebido para publicação em outubro de 2003 e aprovado em junho de 2005.
- ADDISCOTT, T.M. Entropy and sustainability. Eur. J. Soil Sci., 46:161-168, 1992.
- ANDERSON, T.H. & DOMSCH, K.H. Aplication of eco-physiological quotients (qCO2 and qD) on microbial biomass from soils of different cropping histories. Soil Biol. Biochem., 22:251-255, 1990.
- ANDERSON, T.H. & DOMSCH, K.H. The metobolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environment conditions, such as pH, on the microbial biomass of forest soils. Soil Biol. Biochem., 25:393-395, 1993.
- ANDERSON, T.H. Einfluss des Standortes auf die Gesellschaftsstruktur von Mikroorganismen und deren Leistungen unter besonderer Berücksichtigung der C-Nutzung. In: BREDEMEIER, M. & WIEDEY, G.A., eds. Verbundprojekt Veränderungsdynamik von Waldökosystemen, Teil 2. Göttingen, Forschungszentrum Waldökosysteme, 1999. p.385-394.
- ANDERSON, T.H. & GRAY, T.R.G. The influence of soil organic carbon on microbial growth and survival In: WILSON, W.S., ed. Advances in soil organic matter research: the impact on agriculture & the environment. Melksham, Redwood Press, 1991. p.253-260.
- ANDERSON, T.H. Microbial eco-physiological indicators to assess soil quality. Agric. Ecosys. Environ., 98:285-293, 2003.
- ANDERSON, T.H. Physiological analysis of microbial communities in soil: Applications and limitations. In: RITZ, K.D. & GILLER, K.E., eds. Beyond de biomass. London, British Society of Soil Science, 1994. p.67-76.
- BALOTA, E.L.; COLOZZI-FILHO, A.; ANDRADE, D.S. & HUNGRIA, M. Biomassa microbiana e sua atividade em solos sob diferentes sistemas de preparo e sucessão de culturas. R. Bras. Ci. Solo, 22:641-649, 1998.
- BARETTA, D.; SANTOS, J.C.P.; MANFROI, A.F.; TASCA, F.A.; DOMINGOS, M.D.; KLAUBERG-FILHO, O. & MAFRA, A.L. Diversidade da fauna edáfica em mata nativa, floresta de pinus e campo nativo. In: CONGRESSO BRASILEIRO DE CIÊNCIA DO SOLO, 29., Ribeirão Preto, 2003. Resumos expandidos. Ribeirão Preto, SBCS/UNESP, 2003. p.1-4. CD ROM.
- BAYER, C. Dinâmica da matéria orgânica do solo. In: SANTOS, G.A. & CAMARGO, F.A.O., eds. Fundamentos da matéria orgânica do solo-Ecossistemas tropicais e subtropicais. Porto Alegre, Gênesis, 1999. p.9-26.
- BERTOL, I. COGO, N.P & LEVIEN, R. Erosão hídrica em diferentes preparos do solo após as colheitas de milho e trigo, na presença e ausência dos resíduos culturais. R. Bras. Ci. Solo, 21:409-418, 1997.
- CATTELAN, A.J. & VIDOR, C. Flutuação na biomassa, atividade e população microbiana do solo, em função de variações ambientais. R. Bras. Ci. Solo, 14:133-142, 1990.
- CERRI, C.C.; VOLKOFF, B. & EDUARDO, B.P. Efeito do desmatamento sobre a biomassa microbiana em Latossolo Amarelo da Amazônia. R. Bras. Ci. Solo, 9:1-4, 1985.
- CRUZ-CASTILHO, J.G.; GANESHANANDAM, S.; MACKAY, B.R.; LAWES, G.S.; LAWOKO, C.R.O.O. & WOOLLEY, D.J. Applications of canonical discriminat analysis in horticultural research. HortSci., 29:1115-1119, 1994.
- D'ANDRÉA, A.F.; SILVA, M.L.N.; CURI, N.O.; SIQUEIRA, J.O. & CARNEIRO, M.A.C. Atributos biológicos indicadores da qualidade do solo em diferentes sistemas de manejo do solo na região do cerrado no sul do Estado de Goiás. R. Bras. Ci. Solo, 26:913-923, 2002.
- DANIEL, O.; COUTO, L.; SILVA, E.; JUCKSCH, I.; GARCIA, R. & PASSOS, C.A.M. Sustentabilidade em sistemas agroflorestais: Indicadores biofísicos. R. Árvore, 23:381-392, 1999.
- DeBANO, L.; EBERLEIN, G. & DUNN, P. Effects of burning on Chaparral soils: I - Soil Nitrogen. Soil Sci. Soc. Am. J., 43:504-509, 1979.
- DORAN, J.W. & PARKIN, T.B. Defining and assessing soil quality. In: DORAN, J.W.; COLEMAN, D.C.; BEZDICEK, D.F & STEWART, B.A., eds. Defining soil quality for a sustainable environment. Madison, Soil Science Society of America, 1994, p.3-21.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Centro Nacional de Pesquisa de Solos. Sistema brasileiro de classificação de solos. Brasília, Rio de Janeiro, 1999. 412p.
- FERNANDES, V.B.B. A queima de campos nativos no planalto catarinense, em especial no Município de Lages: Subsídios para uma proposta de gerenciamento ecológico. Porto Alegre, Universidade Federal do Rio Grande do Sul, 1997. 111p. (Tese de Mestrado)
- FIALHO, J.F.; BORGES, H.C. & BARROS, N.F. Cobertura vegetal e as características químicas e físicas e atividade da microbiota de um Latossolo Vermelho-Amarelo distrófico. R. Bras. Ci. Solo, 15:21-28, 1991.
- GAMA-RODRIGUES, E.F. Biomassa microbiana e ciclagem de nutrientes. In: SANTOS, G.A. & CAMARGO, F.A.O., eds. Fundamentos da matéria orgânica do solo-Ecossistemas tropicais e subtropicais. Porto Alegre, Gênesis, 1999. p.227-244.
- GERALDES, A.P.A.; CERRI, C.C. & FEIGL, B.J. Biomassa microbiana de solo sob pastagens na Amazônia. R. Bras. Ci. Solo, 19:55-60, 1995.
- GOMES, K.E. Zoneamento das pastagens naturais do planalto catarinense. In: RENIÃO DO GRUPO TÉCNICO REGIONAL DO CONESUL EM MELHORAMENTO E UTILIZAÇÃO DOS RECURSOS FORRAGEIROS DAS ÁREAS TROPICAL E SUBTROPICAL, 11., Lages, 1990. Relatório. Lages, 1990. p.304-314.
- HERINGER, I. & JACQUES, A.V.A. Características de um Latossolo Vermelho sob pastagem natural sujeita à ação prolongada do fogo e de práticas alternativas de manejo. Ci. Rural, 32:309-314, 2002.
- JACQUES, A.V.A. A queima das pastagens naturais - efeitos sobre o solo e a vegetação. Ci. Rural, 33:177-181, 2002.
- JÄGGI, W. Die Bestimmung der CO2-Biulding als Maâ der bonbodenbiologischen Aktivität. Schwiez Landwirtschaft Forchung Band 15, Heft, 314:317-380, 1976.
- JENKINSON, D.S. & LADD, J.N. Microbial biomass in soil: Measurement and turnover. In: PAUL, E.A. & LADD, J.M., eds. Soil Biochem., 5:415-471, 1981.
- KAISER, E.A.; MARTENS, R. & HEINEMEYER, O. Temporal changes in soil microbial biomassa carbon in an arable soil: consequences for soil sampling. Plant Soil, 179:287-295, 1995.
- LUIZÃO, R.C.; CAMPISTA, D. & COSTA, E.S. Efeito da idade e do manejo das pastagens da Amazônia Central sobre a biomassa microbiana e suas atividades no solo. In: SIMPÓSIO BRASILEIRO SOBRE MICROBIOLOGIA DO SOLO, 2., São Paulo, 1991, Anais. São Paulo, Sociedade Brasileira de Ciência do Solo, 1991, p.49.
- MARCHIORI JÚNIOR, M. & MELO, W.J. Alterações na matéria orgânica e na biomassa microbiana em solo de mata natural submetido a diferentes manejos. Pesq. Agropec. Bras., 35:1177-1182, 2000.
- MOREIRA, F.M.S. & SIQUEIRA, J.O. Microbiologia e bioquímica do solo. Lavras, Universidade Federal de Lavras, 2002. 625p.
- MROZ, G.; JURGENSEN, M.; HARVEY, A. & LARSEN, M. Effects of fire on nitrogen in forest floor horizons. Soil Sci. Soc. Am. J., 44:235-242, 1980.
- ODUM, E.P. The strategy of ecosystem development. Science, 64:262-270, 1969.
- OHTONEN, R. Accumulation of organic matter along a pollution gradient: Application of Odum's theory of ecosystem energetics. Microb. Ecol., 27:43-55, 1994.
- PARR, J.F.; PAPENDICK, R.I.; HORNICK, S.B. & MEYER, R.E. Soil quality: attributes and relationship to alternative and sustainable agriculture. Am. J. Altern. Agric., 7:5-10, 1992.
- PASCUAL, J.A.; ROS, M.; HERNANDEZ, T. & GARCIA, C. Effect of long-term monoculture on microbiological and biochemical properties in semiarid soils. Comm. Soil Sci. Plant Anal., 32:537-552, 2001.
- RHEINHEIMER, D.S.; SANTOS, J.C.P.; FERNANDES, V.B.B.; MAFRA, A.L. & ALMEIDA, J.A. Modificações nos atributos químicos de solo sob campo nativo submetido à queima. Ci. Rural, 33:49-55, 2003.
- SAS. INSTITUTE. SAS User's guide: estatistics. 6.ed. Cary, 1990.
- SAUERBECK, D.R.; HELAL, H.M.; NONNEN, S. & ALLARD, J.L. Photosynthate consuption and carbon turnover in the rhizosphere depending on plant species and growth conditions. In: COLÓQUIO REGIONAL SOBRE MATÉRIA ORGÂNICA DO SOLO, 1., Piracicaba, 1982. Anais. Piracicaba, CENA, PROMOCET, 1982. p.171-174.
- SPARLING, G.P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive indicator of changes in soil organic matter. Aust. J. Soil Res., 30:95-207, 1992.
- TEDESCO, M.J.; GIANELLO, C.; BISSANI, C.A. & VOLKWEISS, S.J. Análises de solo, plantas e outros materiais. Porto Alegre, Universidade Federal do Rio Grande do Sul, 1985. 174p. (Boletim Técnico, 5)
- VANCE, E.D.; BROOKS, P.C. & JENKINSON, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem., 19:703-707, 1987.
- WARDLE, D.A. & HUNGRIA, M.A. A biomassa microbiana do solo e sua importância nos ecossistemas terrestres. In: ARAÚJO, R.S. & HUNGRIA, M., eds. Microrganismos de importância agrícola. Brasília, Embrapa-SPI, 1994. p.193-216.
- YAN, T.; YANG, L. & CAMPBELL, C.D. Microbial biomass and metabolic quocient of soils under diferent land use in three Gorges. Geoderma, 1979:1-10, 2003.
Datas de Publicação
-
Publicação nesta coleção
03 Mar 2006 -
Data do Fascículo
Out 2005