Abstracts
The 13C natural abundance technique was applied to study C dynamics after land-use change from native savanna to Brachiaria, Pinus, and Eucalyptus in differently textured Cerrado Oxisols. But due to differences in the d13C signatures of subsoils under native savanna and under introduced species, C substitution could only be calculated based on results of cultivated soils nearby. It was estimated that after 20 years, Pinus C had replaced only 5 % of the native C in the 0-1.2 m layer, in which substitution was restricted to the top 0.4 m. Conversely, after 12 years, Brachiaria had replaced 21 % of Cerrado C to a depth of 1.2 m, where substitution decreased only slightly throughout the entire profile. The high d13C values in the subsoils of the cultivated sites led to the hypothesis that the natural vegetation there had been grassland rather than Cerrado sensu stricto, in spite of the comparable soil and site characteristics and the proximity of the studied sites. The hypothesis was tested using aerial photographs of 1964, which showed that the cultivated sites were located on a desiccated runoff head. The vegetation shift to a grass-dominated savanna formation might therefore have occurred in response to waterlogging and reduced soil aeration. A simple model was developed thereof, which ascribes the different Cerrado formations mainly to the plant-available water content and soil aeration. Soil fertility is considered of minor significance only, since at the studied native savanna sites tree density was independent of soil texture or nutrient status.
Cerrado formations; 13C natural abundance; waterlogging; Oxisol; soil fertility; land-use change
A abundância natural de 13C foi utilizada para estudar a dinâmica de carbono em latossolos de textura média e argilosa após plantação de pastagens (Brachiaria decumbens) e reflorestamentos (Pinus caribaea e Eucalyptus citriodora). Considerando as diferenças de d13C nos subsolos das savanas nativas e das espécies introduzidas, pôde-se calcular a substituição de carbono somente para os solos argilosos, baseando-se em resultados publicados de d13C em solos adjacentes. Estimou-se que somente 5 % do carbono nativo foram substituidos após 20 anos de plantação com pinus na camada de 0-1,2 m, ficando a substituição restrita aos primeiros 0,4 m do solo. Na pastagem de 12 anos, 21 % do carbono novo foram introduzidos na camada de de 0-1,2 m, sendo a substituição diminuída gradativamente pelo perfil inteiro. De acordo com os altos valores de d13C nos subsolos das áreas cultivadas, levantou-se a hipótese de que a vegetação natural teria sido de campo limpo em vez de cerrado stricto sensu apesar da semelhança dos solos e da proximidade dos sítios estudados. A hipótese foi testada com fotos aéreas de 1964, mostrando que as áreas cultivadas encontravam-se numa cabeceira de escoamento ressecada. Portanto, a vegetação dominada por gramíneas pode ter sido induzida pela hidromorfia e pela baixa aeração do solo. Com base nessas observações, desenvolveu-se um modelo simplificado, atribuindo às diferentes formações de cerrados o acesso de água para as plantas e a aeração do solo. A fertilidade do solo foi considerada de menor importância porque a densidade arbórea das áreas de cerrados encontrava-se independente da textura ou do nível de nutrientes dos solos.
formações de cerrados; abundância natural de 13C; hidromorfia; Latossólos; fertilidade do solo; manejo do solo
SEÇÃO V - GÊNESE, MORFOLOGIA E CLASSIFICAÇÃO DO SOLO
Geoecological drivers of cerrado heterogeneity and 13C natural abundance in oxisols after land-use change
Fatores geoecológicos das diferentes formações de cerrados - uma hipótese baseada em abundância natural de 13C e fotografia aérea
Henry Neufeldt
Senior Research Coordinator, Tyndall Institute for Climate Change Research, University of East Anglia, Norwich, NR4 7TJ, United Kingdom. E-mail: h.neufeldt@uea.ac.uk
SUMMARY
The 13C natural abundance technique was applied to study C dynamics after land-use change from native savanna to Brachiaria, Pinus, and Eucalyptus in differently textured Cerrado Oxisols. But due to differences in the d13C signatures of subsoils under native savanna and under introduced species, C substitution could only be calculated based on results of cultivated soils nearby. It was estimated that after 20 years, Pinus C had replaced only 5 % of the native C in the 01.2 m layer, in which substitution was restricted to the top 0.4 m. Conversely, after 12 years, Brachiaria had replaced 21 % of Cerrado C to a depth of 1.2 m, where substitution decreased only slightly throughout the entire profile. The high d13C values in the subsoils of the cultivated sites led to the hypothesis that the natural vegetation there had been grassland rather than Cerrado sensu stricto, in spite of the comparable soil and site characteristics and the proximity of the studied sites. The hypothesis was tested using aerial photographs of 1964, which showed that the cultivated sites were located on a desiccated runoff head. The vegetation shift to a grass-dominated savanna formation might therefore have occurred in response to waterlogging and reduced soil aeration. A simple model was developed thereof, which ascribes the different Cerrado formations mainly to the plant-available water content and soil aeration. Soil fertility is considered of minor significance only, since at the studied native savanna sites tree density was independent of soil texture or nutrient status.
Index terms: Cerrado formations; 13C natural abundance; waterlogging; Oxisol; soil fertility; land-use change.
RESUMO
A abundância natural de 13C foi utilizada para estudar a dinâmica de carbono em latossolos de textura média e argilosa após plantação de pastagens (Brachiaria decumbens) e reflorestamentos (Pinus caribaea e Eucalyptus citriodora). Considerando as diferenças de d13C nos subsolos das savanas nativas e das espécies introduzidas, pôde-se calcular a substituição de carbono somente para os solos argilosos, baseando-se em resultados publicados de d13C em solos adjacentes. Estimou-se que somente 5 % do carbono nativo foram substituidos após 20 anos de plantação com pinus na camada de 01,2 m, ficando a substituição restrita aos primeiros 0,4 m do solo. Na pastagem de 12 anos, 21 % do carbono novo foram introduzidos na camada de de 01,2 m, sendo a substituição diminuída gradativamente pelo perfil inteiro. De acordo com os altos valores de d13C nos subsolos das áreas cultivadas, levantou-se a hipótese de que a vegetação natural teria sido de campo limpo em vez de cerrado stricto sensu apesar da semelhança dos solos e da proximidade dos sítios estudados. A hipótese foi testada com fotos aéreas de 1964, mostrando que as áreas cultivadas encontravam-se numa cabeceira de escoamento ressecada. Portanto, a vegetação dominada por gramíneas pode ter sido induzida pela hidromorfia e pela baixa aeração do solo. Com base nessas observações, desenvolveu-se um modelo simplificado, atribuindo às diferentes formações de cerrados o acesso de água para as plantas e a aeração do solo. A fertilidade do solo foi considerada de menor importância porque a densidade arbórea das áreas de cerrados encontrava-se independente da textura ou do nível de nutrientes dos solos.
Termos de indexação: formações de cerrados; abundância natural de 13C; hidromorfia; Latossólos; fertilidade do solo; manejo do solo.
INTRODUCTION
The Cerrado region covers roughly two million square kilometers of Central Brazil, i.e. 20 % of the national territory, and is characterized by a seasonal climate, low-fertility soils, and a savanna-like vegetation (Adámoli et al., 1986). During the past four decades, the Cerrado region underwent severe changes as gradually more areas were cleared for cultivation. Today pastures cover around 400,000 km2, and approximately 150,000 km2 are cultivated with annual crops and tree plantations (Resck et al., 2000).
After land-use change, C derived from the introduced species (predominantly Brachiaria pastures, maize and soybean as annual crops, and Pinus or Eucalyptus in tree plantations) gradually replaces the original C. By the 13C natural abundance technique (Balesdent & Mariotti, 1996) this alteration can be calculated based on the physiologically different CO2 assimilation pathways between C-3 and C-4 plants. C-3 plants have d13C signatures ranging from -32 to -22 with an average of -27 , and C-4 plants have values ranging from -16 to -9 , with a mean of -13 (Balesdent & Mariotti, 1996).
Since Cerrado sensu stricto (s.s.) comprises both C-3 trees and shrubs and C-4 grasses, its d13C signature is between that of pure C-3 and C-4 vegetations. Roscoe et al. (2001) and Wilcke & Lilienfein (2004) used this feature to quantify the replacement of C derived from Cerrado vegetation after introduction of Brachiaria and Pinus on clayey Oxisols. Neufeldt (1998) applied the 13C technique to differently textured Cerrado Oxisols, but was not able to calculate C replacement due to surprisingly high subsoil d13C values on the sites with the introduced crops.
Neufeldt (1998) hypothesized that slightly higher groundwater levels at the catchment scale had induced a higher natural C-4 grass abundance in the past. Accordingly, catchment scale variations of the moisture balance could explain the occurrence of Cerrado savannas of varied densities on the nearly flat terrain (< 2 % inclination). Similarly, Ker & Resende (1996) observed that the Cerrado on hilltops was tree-dominated and gradually changed into grass-dominated Cerrado formations towards the effluents. In studies of forest-savanna boundaries within the Cerrado region, Emmerich (1989) found that the soil moisture balance was the main determinant factor of vegetation formation, so that mesophytic forests developed where the moisture balance was more stable, whereas soil fertility had no marked influence.
However, this hypothesis disagrees with the common belief that, within the Cerrado biome, there is a relation between, the shift from grass to tree dominated Cerrado formations(3) and soil fertility (Lopes & Cox, 1977; Haridasan, 2000), water availability (Alvim, 1996), or soil acidity and aluminum toxicity (Goodland & Pollard, 1973; Goodland & Ferri, 1979). Furthermore, frequent burning strongly affects height and density of the Cerrado vegetation (Eiten & Sambuichi, 1996; Roscoe et al., 2000), but fires are nowadays seen as a predominantly anthropogenic factor of Cerrado degradation (Alvim, 1996). These contrasting hypotheses on the determining factors of Cerrado formations call for more in-depth research.
For this study, d13C of soils under Cerrado, Brachiaria pastures, and Pinus and Eucalyptus plantations were analyzed and aerial photographs were assessed to test the hypothesis of waterlogging as a key factor of the varied catchment-scale Cerrado formations.
MATERIALS AND METHODS
Site location and history
The study area lies between 19.10 ° to 19.20 ° S and 48.12 ° to 48.18 °W, at a distance of approximately 25 km SSE from Uberlândia, Minas Gerais, Brazil. The mean annual temperature is 22 °C and average precipitation 1.650 mm, 90 % of which falls between October and April. Coarse-loamy, mixed, isohyperthermic Typic Haplustox and very fine, allitic, isohyperthermic Anionic Acrustox (Soil Survey Staff, 1997) were chosen for a regionally representative sampling (Neufeldt et al., 1999). According to the Brazilian taxonomy, the soils were classified as Latossolos vermelho-amarelos álicos A moderado textura média / muito argilosa fase cerrado (Camargo et al., 1986). On the clayey soil, a degraded Brachiaria decumbens pasture, a Pinus caribaea plantation and a Cerrado s.s. savanna were chosen, while on the coarse-loamy soil a degraded Brachiaria decumbens pasture, a Eucalyptus citriodora plantation and a Cerradão were selected. The sites were chosen for their longstanding management history and for being close to each other (< 2 km). The savanna sites differed in tree density, because no comparable sites could be identified within an acceptable distance. Detailed management histories as well as a discussion on the chemical and physical properties of the soils under study are provided in Neufeldt (1999). Table 1 gives an overview of site locations, management records, and soil properties.
Sampling and sample preparation
Soil samples were collected in March 1995, at the end of the rainy season, from each of the six sites. At five randomly selected points per site, samples were taken with an Edelman soil auger from the 00.1, 0.10.2, 0.20.4, 0.40.6, 0.60.8, 0.81, and 11.2 m layers and pooled. The samples were dried in an air-forced oven at 40 °C and passed though a 2 mm sieve to remove roots. For the chemical analyses, sub-samples from each depth were ground for greater homogeneity. Litter samples were collected using a 0.25 ´ 0.25 m2 frame. Thirty samples per treatment were randomly collected, pooled, and dried in an air-forced oven at 65 °C for at least 48 h. One litter sub-sample per treatment was finely ground for further analysis.
Analytical methods
The analytical methods for the determination of soil properties are described in Neufeldt (1999). The signature of stable carbon isotopes was obtained after burning samples of 20100 mg in the presence of CuO under a pure oxygen stream at 900 °C in a dry combustion furnace. In the resulting CO2,12C and 13C were determined with a mass spectrometer (Europa Scientific Roboprep Tracermass) and corrected for 17O influence (Craig, 1957). All analyses were performed in triplicate and presented as mean ± standard error. The d13C value was calculated according to the international V-PDB standard and expressed in tenths of a percent (), according to the equation:

Assessment of aerial photographs
Aerial photographs, taken long before any of the land-use systems were introduced, of flight VM AST-10 1370 PMW R-58, nr. 9298-9301, scale 1:50,000, from August 29, 1964, and of flight MG-103, nr. 100502-100507, nr. 100414-100417, and nr. 100525-100528, scale 1:15,000, from April 24, 1979, were analyzed visually. A stereoscope was used to assess natural differences between Cerrado formations as related to topography.
RESULTS
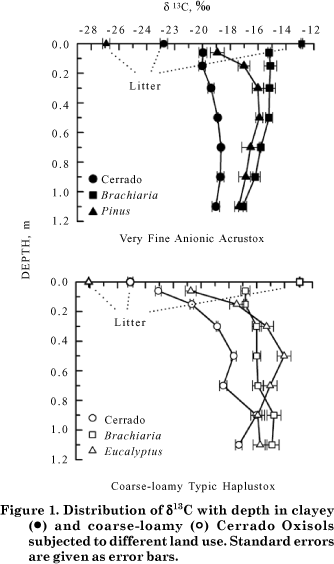
d 13 C signatures in the soil profile
Figure 1 shows the d13C signatures of the clayey and coarse-loamy Oxisols under different management systems in the 01.2 m soil layer. The d13C values of Cerrado litter on the clayey and coarse-loamy soil were -22.8 and -25.1 , respectively, and therefore typical for Cerrados (Roscoe et al., 2000; Wilcke & Lilienfein, 2004). The standards of Brachiaria, Pinus, and Eucalyptus litter were -12.9 , -26.9 , and -28.1 , respectively. The results are in line with the values obtained by Trouve et al. (1994), Roscoe et al. (2001), and Wilcke & Lilienfein (2004).
In the clayey soil under Cerrado s.s., d13C was -20.0 in the 00.2 m layer and increased slightly with depth, reaching -18.7 at 0.8 m. Below that, the signal remained almost constant. Conversely, in the coarse-loamy soil under Cerradão, d13C in the 00.1 m layer was -23.1 , which reflects the higher tree component, and asymptotically approached -17.3 in the 1.01.2 m layer. Between 0.6 and 1.0 m the signals were erratic, which might reflect past vegetation shifts or distinct rooting distributions. According to Wedin et al. (1995), the frequently observed d13C increase at greater depths is related to a gradual shift in the relative contributions of microbially-derived vs. plant-derived components, under the assumption that microbes show the same d13C signatures as the surrounding soil.
Despite lower d13C values of the litter, the soil signatures under Pinus and Eucalyptus clearly suggested enrichment of d13C throughout the profiles, in comparison to the respective Cerrado soils. There was a curvilinear signal increase in the upper 0.6 m ranging from -18.9 to -15.9 in the clayey soil and from -20.8 to -14.0 in the coarse-loamy soil, followed by a continuous decrease to -17.4 and -15.8 in the clayey and coarse-loamy soil, respectively. The signal range was therefore larger in the coarse-loamy soil.
The d13C signals in clayey soils under Brachiaria were constant at around -15.2 to a depth of 0.6 m and subsequently decreased to -17.1 in the deeper subsoil. Conversely, in the coarse-loamy soil under Brachiaria, signals increased stepwise from -16.8 in the 00.1 m layer to -14.9 in the 1.01.2 m layer.
Interpretation of aerial photographs
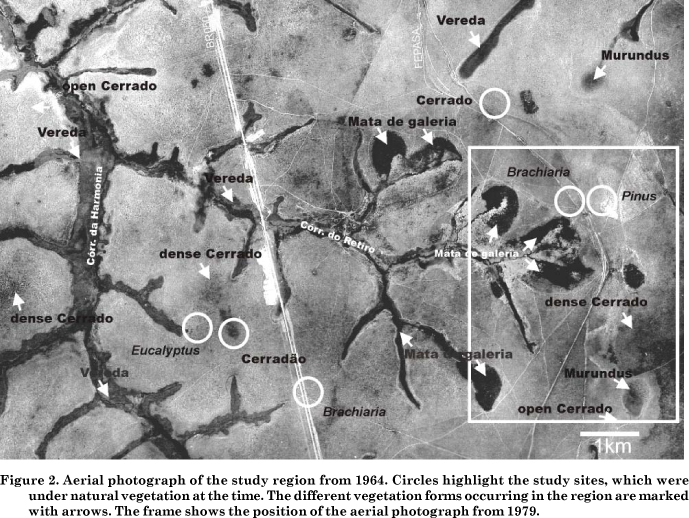
Figure 2 shows a section of an aerial photograph of 1964. Very clayey soils occur on the unconsolidated fine-textured Tertiary sediments of the tableland, to the right of the FEPASA railroad tracks at around 950 m altitude. Left hand of the tracks, the clay content of the soils continuously decreases as the sandstones of the Marília formation (Eocretaceous) are exposed (Nishyama, 1989), reaching an altitude of approximately 900 m at the BR 050 interstate highway (then under construction). Hence, the sites with clayey soils were located near the railroad tracks, whereas the sites with coarse-loamy soils were close to the road (site locations encircled).
Savanna dominates the natural vegetation between the effluents and is generally denser on hilltops (Figure 2). The other natural vegetation forms are closely associated to the drainage system. Mata de galeria (gallery forest) grows where inclination gives the runoff a downhill direction. Veredas, which are palm groves characterized by the Burití palm (Mauritia flexuosa), are found in shallow valleys with diffuse runoff and high groundwater levels throughout the whole year. Murundus are convex microstructures (probably termite mounds) of up to 1.5 m height and 20 m diameter within flat depressions or runoff heads with very low inclination (Ribeiro & Walter, 1998). The vegetation of the runoff heads is predominantly grassy with few or no woody components. In the study area, Murundus only occur on the fine-textured sediments of the tableland (Schneider, 1996). Figure 3 shows these structures more clearly, together with former runoff heads, later dried out, so that Pinus was planted on them in 1979.
DISCUSSION
Replacement of Cerrado C after land-use change
Since the cultivated soils and the adjacent Cerrado sites did not show similar subsoil d13C signatures, the Cerrado soils being depleted in d13C compared to the soil under introduced species, and hence lacking a common baseline, a direct calculation of C substitution after land-use change was not possible (Figure 1). On the clayey soils this was probably caused by a higher C-4 grass component in the past due to their proximity to an ancient runoff head (Figure 3). On the coarse-loamy soils, the Cerrado site seems to have been exceptionally dense, suggesting that the grass component had again been higher on the Eucalyptus and Brachiaria sites prior to cultivation (Figure 2).
An estimation of the C proportion derived from Pinus and Brachiaria in the clayey soils however seems possible, based on results of Wilcke & Lilienfein (2004) of comparable soils nearby. For the loamy soils, the calculation of C replacement by introduced species was not undertaken, since neither reference studies were available to compare the measured values with nor a similar subsoil d13C distribution between Cerradão, Eucalyptus, and Brachiaria that could serve as baseline.
Wilcke & Lilienfein (2004) showed that 20 yr of Pinus caribaea plantation on a clayey soil only led to C replacement in the topsoil (00.3 m), whereas C in the subsoil (0.32 m) remained unaffected by the land-use change. Similar results were obtained by Trouve et al. (1994) for 30 year-old Pinus caribaea stands in Congo. Wilcke & Lilienfein (2004) further reported that Brachiaria decumbens -derived C replaced 531 % of the original in the 0-2 m layer after 12 yr of cultivation. Roscoe et al. (2001) described a similar C substitution throughout the whole soil profile after 23 yr under Brachiaria spp. pasture in Sete Lagoas, MG, Brazil. Hence, C substitution rates for Pinus and for Brachiaria were comparable at different locations in Brazil and elsewhere, and it therefore seems valid to assume a similar response to the introduction of Pinus and Brachiaria on the study sites as well.
C substitution after vegetation change to Pinus was calculated using the measured d13C values, and assuming that no C replacement occurred below 0.4 m under the studied Pinus plantation. It was estimated that C derived from Pinus replaced 27 % of the original C in the 00.1 m layer, 10 % in the 0.1.0.2 m layer, and 1 % in the 0.2-0.4 m layer. In the total layer (01.2 m), only 5 % C were Pinus-derived after 20 yr. The estimated values strongly agree with the results of Wilcke & Lilienfein (2004) and Trouve et al. (1994), and probably reflect the fact that the Pinus litter accumulates on the soil surface, forming a thick organic layer (520 cm) that is only slowly incorporated into the soil (Neufeldt et al., 2002), and a comparatively low rooting intensity in the subsoil.
C substitution by Brachiaria on the clayey soil was calculated by relating the d13C pasture values to those of the corresponding layer of the Pinus site, because both appeared to have had a similar grassland vegetation in the past due to the adjacent runoff head (Figure 1). C substitution under Brachiaria was estimated to be nearly constant at about 24 % to 0.6 m depth, and then decreased gradually to 6 % in the 1.01.2 m layer. In the total layer (01.2 m), 21 % C were pasture-derived. Both the amounts of C replacement and their distribution in the profile are in close agreement with the results of Wilcke & Lilienfein (2004) and Roscoe et al. (2001). The high C substitution is a response to the profound and dense pasture rooting system and the relatively low recalcitrance of the soil organic matter despite high clay contents (Roscoe et al., 2001; Neufeldt et al., 2002).
Summing up, the underlying assumptions allowed the calculation of C replacements, which agree very well with published results. This suggests that the initial hypothesis, according to which the vegetation on the Brachiaria and Pinus sites had been dominated by C-4 grasses in the past due to waterlogging, appears to be correct. Aerial photographs (Figures 1 and 2), which were taken into consideration thereupon, reinforced the premise. However, analyses of stable isotopes and soil moisture contents along natural Cerrado vegetation gradients are required to verify the hypothesis.
Cerrado formations in response to geoecological drivers
Aerial photographs (Figure 2) and 13C natural abundance (Figure 1) indicated a strong heterogeneity of the studied savanna formations and suggest that high groundwater levels limit Cerrado tree growth in favor of grasses. This is confirmed by Eiten (1972), according to whom Cerrado trees are sensitive to waterlogging. Since the lateral extent of the groundwater influence is related to inclination, the affected zone may be very large on flat topographies. On the other hand, the coarse-loamy soils are clearly less fertile than clayey soils as indicated by their substantially lower cation exchange capacities (CEC) and C concentrations (Table 1). This does not seem to affect Cerrado physiognomy, as dense Cerrado s.s. and Cerradão frequently occur on the coarse-loamy soils derived from the Marília formation sandstones (Figure 1). Hence, on a catchment scale of several square kilometers, soil fertility does not seem to play such a prominent role for Cerrado density as frequently proposed (Goodland & Pollard, 1973; Lopes & Cox, 1977; Goodland & Ferri, 1979; Adámoli et al., 1986; Haridasan, 2000). This is consistent with Emmerich (1989) who showed that mainly the soil moisture balance determined the vegetation form along savanna-forest boundaries within the Cerrado region, whereas soil fertility played only a minor role. Recently, Ruggiero et al. (2002) studied the soil-vegetation relationships in Campo cerrado, Cerrado s.s., and Cerradão, but found no similar trends in the Cerrado formations in terms of nutrient status, base saturation, Al saturation, and CEC. However, in contrast to Emmerich (1989), they could clearly distinguish the soil fertility of the Cerrado formations from the adjacent semideciduous forest. The forest plots presented higher clay contents and CEC and the Cerrado higher Al saturation in surface soils.
Based on these observations, a simple model is proposed in an attempt to integrate the distribution of Cerrado formations. The model centers around the plant-available water content and soil aeration, in view of the plants' water stress during the dry season (Alvim, 1996) and the sensitivity of the Cerrado trees to waterlogging (Eiten, 1972), respectively. On the other hand, soil fertility is given a minor role only. The model is restricted to the catchment scale (up to several km2) and explicitly does not question the superordinate relevance of climate for the occurrence of the phytomorphological domains on the Brazilian territory. The Cerrado biome prevails where the amount and temporal distribution of precipitation is intermediate between the moist Amazonian and Atlantic forest biomes on the one hand and the dry Caatinga and Chaco biomes on the other (Ab'Sáber, 1971; Adámoli et al., 1986). The effects of fire on the Cerrado features are not discussed in this context either, since burning is seen as a predominantly anthropogenic cause of Cerrado degradation (Alvim, 1996; Eiten & Sambuichi, 1996).
Table 2 shows the different vegetation formations that occur naturally within the study region vs. different geoecological drivers. With exception of the Vereda, which is characterized by the Burití palm tree, the vegetation formations are in the order of increasing canopy height and tree density (Goodland & Ferri, 1979). While the Vereda is flooded for most of the year, the grasses on the Murundu and Campo limpo or Campo sujo may temporarily suffer from drought during the dry season due to the restricted rooting depth, which limits the plant-available water content. While in the study area rooting depth is restricted by a high groundwater level in the vicinity of the effluents, solid bedrock may limit root penetration in other regions. According to Eiten (1972), Campo sujo and Campo limpo are frequently found on very shallow soils (e.g. steep slopes or rock outcrops) with good or poor drainage. Goodland & Ferri (1979) suggested that the few dwarf trees on the Campo sujo might reflect faults within a laterite crust or the bedrock. As rooting depth and thus the plant-available water content increases, the grass-dominated Cerrado formations are gradually replaced by tree-dominated Cerrado formations. This corresponds to the fact that some Cerrado trees are very deep-rooting (Eiten, 1972). Under otherwise comparable conditions, soil fertility should also play a role for the distribution of the vegetation forms and certainly has an influence on the phytosociological composition of the Cerrado (Goodland & Pollard, 1973; Alvim, 1996). In conclusion, tree density rises with increasing plant-available water content provided that good internal drainage conditions a well aerated solum which does not impose physical resistance (e.g. waterlogging, laterite, bedrock) to deep rooting. The Mata de galeria is restricted to the effluents with directed runoff and occurs on deep, well-drained, and possibly more fertile soils. Plants of the Mata de galeria, which floristically belong to the mesophytic Atlantic forest biome (Eiten, 1972), have access to water from the capillary fringe of the stream and are therefore less exposed to drought during the dry season. Apparently, Cerrado vegetation cannot compete with the gallery forest under these circumstances. Similarly, Emmerich (1989) reported that this feature can be observed on the intra-montane planes west of Brasília (Cerrado biome), where additional water (and possibly nutrients) from the surrounding slopes allows mesophytic forest to thrive. The centers of these planes, where the groundwater level reaches its minimum are, however, dominated by dense Cerrado.
Recapitulating, the presence of different Cerrado formations can mainly be ascribed to good internal drainage and the absence of waterlogging or other physical barriers to rooting. The soil nutrient status or Al saturation, frequently cited to explain the occurrence of different Cerrado formations, are seen as less important.
Acknowledgements
The German Technical Cooperation (GTZ) enabled the conduction of this study "Soil Indicators of Sustainable Agropastoral Systems" with grant PN 94.7860.3-01.100, in cooperative work of the Institute of Soil Science and Soil Geography of the University of Bayreuth with the Brazilian agricultural research organization Embrapa Cerrados (CPAC) and the International Center for Tropical Agriculture (CIAT). I am further grateful to Marco Rondón of the CIAT for his valuable help with the 13C technique.
LITERATURE CITED
Received for publication in August of 2005 and aproved in August of 2006.
- AB'SÁBER, A.N. A organização natural das paisagens inter e subtropicais Brasileiras. In: SIMPÓSIO SOBRE O CERRADO, 3., São Paulo, 1971. Anais. São Paulo, Blücher, 1971. p.1-14.
- ADÁMOLI, J.; MACEDO, J.; AZEVEDO, L.G. & MADEIRA NETO, J.S. Caracterização da região do cerrado. In: GOEDERT, W.J., ed. Solos dos cerrados: tecnologias e estratégias de manejo. São Paulo: Nobel, 1986. p.33-74.
- ALVIM, P.T. Repensando a teoria da formação dos Campos Cerrados. In: Simpósio sobre o cerrado, 8., Planaltina, 1976. Anais. Embrapa-CPAC, Planaltina, 1996. p.56-58.
- BALESDENT, J. & MARIOTTI, A. Measurement of soil organic matter turnover using 13C natural abundance. In: BOUTTON, T.W. & YAMASAKI, S., eds. Mass spectrometry of soil. New York, Marcel Dekker,1996. p.83-111.
- CAMARGO, M.N.; KLAMT, E. & KAUFFMAN, J.H. Classificação de solos usada em levantamentos pedológicos no Brasil. Campinas, Sociedade Brasileira de Ciências do Solo, 1986. 24p.
- CRAIG, H. Isotopic standards for carbon and oxygen and correction factors for mass-spectrometric analysis of carbon dioxide. Geochim. Cosmochim. Acta, 12: 133-149, 1957.
- EITEN, G. The Cerrado vegetation of Brazil. Bot. Rev., 38: 201-341, 1972.
- EITEN, G. & SAMBUICHI, R.H.R. Effect of long-term periodic fire on plant diversity in a "Cerrado" region. In: SIMPÓSIO SOBRE O CERRADO, 8., Planaltina, 19896, Anais. Embrapa-CPAC, Planaltina, 1996. p.46-55.
- EMMERICH, K.H. Relief und quartäre reliefgenese als steuerfaktoren der wald-savannen-verteilung in brasilien. Bayreuther Geowiss. Arb., 14:9-24, 1989.
- GOODLAND, R.J. & POLLARD, R. The Brazilian cerrado vegetation: a fertility gradient. J. Ecol., 61:219-224, 1973.
- GOODLAND, R. & FERRI, M.G. Ecologia do cerrado. São Paulo, EDUSP, 1979. 193p.
- HARIDASAN, M. Nutrição mineral de plantas nativas do Cerrado. R. Bras. Fisiol. Veg., 12:54-64, 2000.
- KER, J.C. & RESENDE, M. Recursos edáficos dos Cerrados: ocorrência e potencial. In: Simpósio sobre o cerrado, 8., Planaltina, 1996. Anais. Embrapa-CPAC, Planaltina, 1996. p.15-19.
- LOPES, A.S. & COX, F.R. A survey of the fertility status of surface soils under 'cerrado' vegetation in Brazil. Soil Sci. Soc. Am. J., 41:741-747, 1977.
- NEUFELDET, H. Land-use effects on soil chemical and physical properties of cerrado Oxisols. Bayreuth, University of Bayreuth, 1998. 174p. (Tese de Doutorado)
- NEUFELDET, H. Physical and chemical properties of selected Oxisols in the Brazilian Cerrados. In: THOMAS, R. & AYARZA, A., ed. Sustainable land management for the Oxisols of the Latin American Savannas. Cali, CIAT, 1999. p.37-50. (CIAT Publication, 312)
- NEUFELDET, H.; SCHNEIDER, M.O. & ZECH, W. Oxisol development along a compound catena of the Araguari river, Central Brazil. In: THOMAS, R. & AYARZA, A., ed. Sustainable Land Management for the Oxisols of the Latin American Savannas. CIAT publication no. 312, Cali, CIAT, 1999. p.10-21.
- NEUFELDET, H.; RESCK, D.V.S. & AYARZA, M.A. Texture and land-use effects on soil organic matter in Cerrado Oxisols, Central Brazil. Geoderma, 107:151-164, 2002.
- NISHYAMA, L. Geologia do município de Uberlândia e áreas adjacentes. Sociedade Natureza, 1:9-16, 1989.
- RESCK, D.V.S.; VASCONCELLOS, C.A.; VILELA, L. & MACEDO M.C.M. Impact of conversion of Brazilian Cerrados to cropland and pastureland on soil carbon pools and dynamics. In: LAL, R.; KIMBLE, J.M. & STEWART, B.A., ed. Global climate change and tropical ecosystems. Boca Raton, CRC Press, 2000. p.169-196.
- RIBEIRO, J.F. & WALTER, B.M.T. Fitofisionomias do bioma cerrado. In: SANO, S.M. & ALMEIDA, S.P., eds. Cerrado: ambiente e flora. Planaltina, Embrapa-CPAC, 1998. p.89-168.
- ROESCOE, R.; BUURMAN, P.; VELTHORST, E.J. & PEREIRA, J.A.A. Effects of fire on soil organic matter in a "cerrado sensu-stricto" from Southeast Brazil as revealed by changes in d 13C. Geoderma, 95:141-160, 2000.
- ROSCOE, R.; BUURMAN, P.; VELTHORST, E.J. & VASCONCELLOS, C.A. Soil organic matter dynamics in density and particle size fractions as revealed by the 13C/12C isotopic ratio in a Cerrado Oxisol. Geoderma, 104:185-202, 2001.
- RUGGIERO, P.G.C.; BATALHA, M.A.; PIVELLO, V.R. & MEIRELLES, S.T. Soil-vegetation relationships in cerrado (Brazilian savanna) and semideciduous forest, Southeastern Brazil. Plant Ecol., 160:1-16, 2002.
- SCHNEIDER, M.O. Bacia do rio Uberabinha: Uso agrícola do solo e meio ambiente. Uberlândia, Unversidade Federal de Uberlândia, 1996. 90p. (Tese de Doutorado)
- SOIL SURVEY STAFF Keys to Soil Taxonomy. 7. ed., SMSS, Blacksburg, Pocahontas Press, 1997. 326p. (Tech. Monography No, 19)
- TROUVE, C.; MARIOTTI, A.; SCHWARTZ, D. & GUILLET, B. Soil organic carbon dynamics under Eucalyptus and Pinus planted on savannas in the Congo. Soil Biol. Biochem., 26:287-295, 1994.
- WEDIN, D.A.; TIESZEN, L.L.; DEWEY, B. & PASTOR, J. Carbon isotope dynamics during grass decomposition and soil organic matter formation. Ecology, 76:1383-1392, 1995.
- WILCKE, W. & LILIENFEIN, J. Soil carbon-13 natural abundance under native and managed vegetation in Brazil. Soil Sci. Soc. Am. J., 68:827-832, 2004.
Publication Dates
-
Publication in this collection
14 Dec 2006 -
Date of issue
Oct 2006
History
-
Accepted
Aug 2006 -
Received
Aug 2005