Resumos
Bactérias diazotróficas não simbióticas (BDNS) atuam no desenvolvimento das plantas por meio da fixação biológica de nitrogênio e também pela produção e liberação de substâncias reguladoras do crescimento vegetal. Este estudo objetivou avaliar a densidade e diversidade fenotípica desse grupo de bactérias em fragmentos de solo da Reserva Biológica Serra dos Toledos e entorno, em Itajubá/MG. Essa reserva localiza-se na Área de Proteção Ambiental da Mantiqueira, constituindo-se em uma importante área de recarga e de abrigo à flora e fauna endêmicas. Amostras de solo superficial foram coletadas em áreas com diferentes declividades na reserva, em épocas representativas das estações de inverno (setembro/2006) e verão (abril/2007). A densidade, avaliada pelo número mais provável, utilizando os meios de cultura NFb, JNFb e Fam, para Azospirillum spp., Herbaspirillum spp. e A. amazonense, respectivamente, variou de 0,12 a 75,60 (NMP x 10(5)) bactérias g-1 solo seco. Foram obtidos 172 e 174 isolados, respectivamente para as amostras de inverno e verão, dos quais 30 e 55 % apresentaram similaridade igual ou superior a 70 % com as estirpes-tipo Azospirillum brasilense, A. amazonense, A. lipoferum, Herbaspirillum seropedicae e Burkholderia brasilensis. O resultado do comportamento dos isolados com base na tolerância à salinidade nem sempre foi semelhante ao obtido pelas características fenotípicas culturais a 70 % de similaridade, sendo indicado para estudos complementares de diversidade desses organismos. As BDNS apresentam potencial de utilização em estudos de avaliação da qualidade e sustentabilidade de ecossistemas. No entanto, apesar da alta densidade e diversidade fenotípica em solos da reserva, maiores valores foram obtidos no entorno, evidenciando o efeito positivo da cobertura vegetal do tipo gramíneas sobre elas, independentemente da variação climática.
fixação biológica de nitrogênio; reserva biológica; Azospirillum; Herbaspirillum; Burkholderia
Non-symbiotic diazotrophic bacteria (NSDB) influence plant growth through biological nitrogen fixation and also production and release of plant growth-regulating substances. The aim of this study was to evaluate the density and phenotypic diversity of this bacterial group in fragments of the "Reserva Biológica Serra dos Toledos" and surroundings, in Itajubá/MG. This reserve is located in the environmental protection area of Mantiqueira and represents an important recharge area and shelter for endemic flora and fauna. Topsoil samples were collected in areas with different slopes in the reserve and surroundings, at the end of the winter season (September/2006) and of the summer (April/2007). Population densities were evaluated by the Most-Probable-Number method (MPN) using the growth media NFb, JNFb and Fam for Azospirillum spp., Herbaspirillum spp. and A. amazonense, respectively, and a range of 0.12 to 75.60 (MPN x 10(5)) bacteria g-1 dry soil was observed. In September/2006, 172 isolates were obtained, of which 30 % had a similarity of at least 70 % with the reference strains. In April/2007, 174 isolates were obtained, in 55 % of which a similarity of at least 70 % was observed with the reference strains evaluated (Azospirillum brasilense, A. amazonense, A. lipoferum, Herbaspirillum seropedicae and Burkholderia brasilensis). The results based on salt tolerance of the isolates were not always similar to those related to phenotypic similarity of 70 % of the cultures; so salt tolerance is recommended for additional diversity studies on these NSDB. The NSDB are indicated for studies on quality and sustainability of ecosystems. However, despite the high bacteria population density and phenotypic diversity in soils of the reserve, higher values were observed in the neighboring areas, showing the positive effect of a grass cover, regardless of climatic variations.
biological nitrogen fixation; biological reserve; Azospirillum; Herbaspirillum; Burkholderia
DIVISÃO 2 - PROCESSOS E PROPRIEDADES DO SOLO
2.1 - BIOLOGIA DO SOLO
Densidade e diversidade fenotípica de bactérias diazotróficas não simbióticas em solos da Reserva Biológica Serra dos Toledos, Itajubá (MG)1 1 Trabalho apresentado na FERTBIO 2008, Londrina (PR).
Density and phenotypic diversity of non-symbiotic diazotrophic bacteria in soils of the Biological Reserve Serra dos Toledos, Itajubá (MG)
Talita Filomena SilvaI; Rogério MelloniII
IEngenheira ambiental e mestranda em Meio Ambiente e Recursos Hídricos, Universidade Federal de Itajubá UNIFEI. Av. BPS 1303, Pinheirinho, CEP 37500903 Itajubá (MG). E-mail: taeam@unifei.edu.br
IIDocente do Instituto de Recursos Naturais, UNIFEI. E-mail: rmelloni@unifei.edu.br
RESUMO
Bactérias diazotróficas não simbióticas (BDNS) atuam no desenvolvimento das plantas por meio da fixação biológica de nitrogênio e também pela produção e liberação de substâncias reguladoras do crescimento vegetal. Este estudo objetivou avaliar a densidade e diversidade fenotípica desse grupo de bactérias em fragmentos de solo da Reserva Biológica Serra dos Toledos e entorno, em Itajubá/MG. Essa reserva localiza-se na Área de Proteção Ambiental da Mantiqueira, constituindo-se em uma importante área de recarga e de abrigo à flora e fauna endêmicas. Amostras de solo superficial foram coletadas em áreas com diferentes declividades na reserva, em épocas representativas das estações de inverno (setembro/2006) e verão (abril/2007). A densidade, avaliada pelo número mais provável, utilizando os meios de cultura NFb, JNFb e Fam, para Azospirillum spp., Herbaspirillum spp. e A. amazonense, respectivamente, variou de 0,12 a 75,60 (NMP x 105) bactérias g-1 solo seco. Foram obtidos 172 e 174 isolados, respectivamente para as amostras de inverno e verão, dos quais 30 e 55 % apresentaram similaridade igual ou superior a 70 % com as estirpes-tipo Azospirillum brasilense, A. amazonense, A. lipoferum, Herbaspirillum seropedicae e Burkholderia brasilensis. O resultado do comportamento dos isolados com base na tolerância à salinidade nem sempre foi semelhante ao obtido pelas características fenotípicas culturais a 70 % de similaridade, sendo indicado para estudos complementares de diversidade desses organismos. As BDNS apresentam potencial de utilização em estudos de avaliação da qualidade e sustentabilidade de ecossistemas. No entanto, apesar da alta densidade e diversidade fenotípica em solos da reserva, maiores valores foram obtidos no entorno, evidenciando o efeito positivo da cobertura vegetal do tipo gramíneas sobre elas, independentemente da variação climática.
Termos de indexação: fixação biológica de nitrogênio, reserva biológica, Azospirillum, Herbaspirillum, Burkholderia.
SUMMARY
Non-symbiotic diazotrophic bacteria (NSDB) influence plant growth through biological nitrogen fixation and also production and release of plant growth-regulating substances. The aim of this study was to evaluate the density and phenotypic diversity of this bacterial group in fragments of the "Reserva Biológica Serra dos Toledos" and surroundings, in Itajubá/MG. This reserve is located in the environmental protection area of Mantiqueira and represents an important recharge area and shelter for endemic flora and fauna. Topsoil samples were collected in areas with different slopes in the reserve and surroundings, at the end of the winter season (September/2006) and of the summer (April/2007). Population densities were evaluated by the Most-Probable-Number method (MPN) using the growth media NFb, JNFb and Fam for Azospirillum spp., Herbaspirillum spp. and A. amazonense, respectively, and a range of 0.12 to 75.60 (MPN x 105) bacteria g-1 dry soil was observed. In September/2006, 172 isolates were obtained, of which 30 % had a similarity of at least 70 % with the reference strains. In April/2007, 174 isolates were obtained, in 55 % of which a similarity of at least 70 % was observed with the reference strains evaluated (Azospirillum brasilense, A. amazonense, A. lipoferum, Herbaspirillum seropedicae and Burkholderia brasilensis). The results based on salt tolerance of the isolates were not always similar to those related to phenotypic similarity of 70 % of the cultures; so salt tolerance is recommended for additional diversity studies on these NSDB. The NSDB are indicated for studies on quality and sustainability of ecosystems. However, despite the high bacteria population density and phenotypic diversity in soils of the reserve, higher values were observed in the neighboring areas, showing the positive effect of a grass cover, regardless of climatic variations.
Index terms: biological nitrogen fixation, biological reserve, Azospirillum, Herbaspirillum, Burkholderia.
INTRODUÇÃO
O N é geralmente considerado o nutriente mais limitante para o crescimento de plantas no seu ambiente natural (Buchanan et al., 2000). Na natureza, esse elemento é encontrado em abundância em uma forma quimicamente estável (N2), a qual não é acessível à maioria dos seres vivos, sendo necessária a sua transformação para uma forma combinada que facilite sua assimilação (Marin et al., 1999). Segundo Moreira & Siqueira (2006), apenas uma pequena parcela das espécies de procariotos possui a enzima nitrogenase, que é capaz de reduzir o N2 para a forma inorgânica combinada NH3, disponível às plantas e outros organismos, sendo estes organismos denominados diazotróficos e o mecanismo responsável pela incorporação de N à biomassa é chamado de fixação biológica de N2 (FBN).
Entre os sistemas biológicos capazes de aproveitar o N diretamente da atmosfera, a simbiose rizóbio-leguminosa tornou-se o mais especializado, com destaque para a cultura da soja, onde até 94 % do N requerido pelas cultivares mais produtivas pode ser fornecido pela FBN (Hungria et al., 2006). Um dos grandes interesses dos pesquisadores é a extensão desse processo biológico para outras culturas, como gramíneas e cereais, principalmente as de grande importância socioeconômica (Sala et al., 2005). As associações entre bactérias diazotróficas e não leguminosas são denominadas comumente de simbiose associativa ou fixação de N2 associativa (Baldani et al., 1997a). Essas bactérias diazotróficas não simbióticas (BDNS), assim chamadas por Kennedy et al. (2004), apesar de serem aeróbias, apenas conseguem fixar o N2 quando este se encontra como única fonte de N em condições microaerófilas (Döbereiner, 1992).
Entre as BDNS mais estudadas destacam-se aquelas pertencentes aos gêneros Azospirillum, Herbaspirillum e Burkholderia, que já foram isoladas de plantas como arroz (Rodrigues et al., 2006), milho (Perin et al., 2006), cana-de-açúcar (Reis Junior et al., 2000) e Brachiaria spp. (Reis Junior et al., 2004). Estudos consideram bactérias do gênero Azospirillum como diazotróficas facultativas, sendo capazes de colonizar raízes de plantas não leguminosas interna e externamente (Baldani et al., 1997a). O gênero Herbaspirillum, apesar de também sobreviver no solo, é comumente encontrado no interior de raízes e também na parte aérea das plantas (Olivares et al., 1997), assim como o gênero Burkholderia, que teve sua capacidade diazotrófica descoberta na década de 90 (Baldani et al., 1997b).
As BDNS podem desempenhar importante papel na sustentabilidade dos ecossistemas, uma vez que incorporam N2 por meio da fixação biológica, podendo produzir e liberar substâncias reguladoras do crescimento vegetal (Dobbelaere et al., 2003), como auxinas, giberelinas e citocininas, as quais contribuem para melhorar a nutrição mineral e utilização de água pelas plantas (Bazzicalupo & Okon, 2000). No entanto, Bashan & Holguin (1997) relataram que a ocorrência e a atividade dessas bactérias no solo e na planta, ressalvando as condições de ambiente favoráveis, são fortemente influenciadas por estresses físicos (baixa umidade e alta temperatura), químicos (acidez e baixos teores de nutrientes e C) e biológicos (espécie vegetal não hospedeira).
Diversos trabalhos têm sido realizados nos últimos anos nessa área de BDNS no Brasil, principalmente em culturas como trigo (Sala et al., 2007), milho (Reis Junior et al., 2008), cana-de-açúcar (Oliveira et al., 2009), arroz (Ferreira et al., 2010), abacaxi (Baldotto et al., 2010), entre outras, evidenciando o potencial dessas bactérias no crescimento das culturas.
A Reserva Biológica Serra dos Toledos localiza-se na Área de Proteção Ambiental da Mantiqueira, sendo o maior trecho de Mata Atlântica preservada da região sul mineira, abrigando grande diversidade de espécies da fauna e flora endêmicas (Itajubá, 2002a). É um ecossistema rico em mananciais e participa do fornecimento de água potável ao município de Itajubá/MG. A avaliação das BDNS dentro da reserva foi definida em razão da importância na incorporação de N ao solo e devido à possibilidade de esses microrganismos indicarem variações de atributos edáficos, comuns em áreas de interferência antrópica. Esses estudos, aliados a outros, servirão como ferramentas para a elaboração de um plano estratégico de gerenciamento da reserva.
O objetivo do presente estudo foi avaliar a densidade e diversidade fenotípica de isolados de BDNS pertencentes aos gêneros Azospirillum, Herbaspirillum e Burkholderia, em solos sob gradientes de declividade na Reserva Biológica Serra dos Toledos e entorno, em duas épocas contrastantes (setembro/2006 e abril/2007).
MATERIAL E MÉTODOS
Após levantamento de dados sobre mapas de declividades da Reserva Biológica Serra dos Toledos, foram selecionadas três áreas no seu interior, representativas dessas variações de gradientes. Uma área foi selecionada fora da reserva e sob ação antropogênica, para confronto dos resultados. As áreas estudadas foram: Área Plana (P): área em relevo suave a suave ondulado, 0,13 m m-1 de declive (22 º 25 ' 48,8 " sul e 45 º 22 ' 18,3 " oeste) e altitude de 1.311 m; Área Mediana (M): área em relevo ondulado, declive de 0,35 m m-1 (22 º 25 ' 33,3 " sul e 45 º 22 ' 20,9 " oeste) e altitude de 1.306 m; Área Inclinada (I): área em relevo montanhoso, 0,56 m m-1 de declive (22 º 25 ' 45,5 " sul e 45 º 22 ' 17,0 " oeste) e altitude de 1.307 m; e Área Fora (F): área de média a baixa declividade, localizada fora da reserva (22 º 25 ' 28,2 " sul e 45 º 22 ' 15,0 " oeste), com altitude de 1.307 m, onde se pode observar a presença de gado e vegetação de campo (pasto abandonado em sucessão, com domínio de braquiária (Brachiaria spp.)). Estudo realizado pelo Instituto Estadual de Florestas (IEF) nesse ecossistema revela a presença de espécies importantes, como peroba (Aspidosperma parvifolium), angico (Parapiptadenia rigida), jacarandá (Jacaranda mimosifolia), cedro (Cedrela fissilis), jequitibá (Cariniana estrellensis), cajarana (Spondias dulcis), candeia (Arisarum vulgare), canelas (Nectandra lanceolata), araucária (Araucaria angustifolia) e grande variedade de bromélias (Vriesea sp.) e orquídeas (Dendrobium sp.), e baixa densidade de gramíneas no sub-bosque em virtude do elevado sombreamento. Sua cobertura vegetal original é representada pelo contato Floresta Ombrófila Densa/Floresta Ombrófila Mista, sendo atualmente a maior parte composta por vegetação secundária (Itajubá, 2002a).
Sob influência da elevada altitude da região, o clima da reserva é do tipo tropical temperado, com oscilações bruscas de temperatura e predominância de ventos nordeste (NE), com temperatura média de 25 ºC e precipitação pluvial de 1.410 mm ano-1; as estações do ano são bem definidas, com maiores temperaturas e precipitação pluvial durante os meses de verão, e clima frio e seco nos meses de inverno, com temperaturas abaixo de 10 ºC (Itajubá, 2002b).
Cada área foi subdividida em cinco subáreas, obtendo-se cinco amostras compostas, formadas por cinco subamostras cada. As subamostras foram retiradas em ziguezague, na profundidade de 0 a 10 cm, em setembro/2006 (representativa do inverno) e abril/2007 (representativa do verão), na distância de 10 m entre elas. As amostras de solo foram acondicionadas em sacos plásticos e encaminhadas ao Laboratório de Microbiologia, da UNIFEI, peneiradas em malha de 4 mm e mantidas a 4 ºC até a determinação dos atributos microbiológicos. Para cada amostra, determinou-se a umidade atual do solo. O solo de todas as áreas possui mesma formação litológica (gnaisses finos, escuros a claros, muito deformados), sendo classificado como Argissolo Vermelho-Amarelo alumínico (Nassar, 2007).
Para a avaliação da densidade de BDNS, foram preparados os meios de cultura com consistência semissólida NFb para Azospirillum spp. e JNFb para Herbaspirillum spp., conforme descrito em Döbereiner et al. (1995), e o meio Fam para Azospirillum amazonense, descrito em Magalhães (1983). Foram realizadas diluições sucessivas das amostras de solo em solução salina, de 10-1 a 10-5; dessas suspensões, 0,1 mL foi inoculado em frascos contendo os meios de cultura semissólidos, com três repetições por diluição. Essas culturas foram mantidas sob incubação por 7 dias a 28 ºC, avaliando-se a presença ou ausência de crescimento bacteriano pela formação de película próxima à superfície no meio. As densidades de bactérias foram avaliadas por meio do método do número mais provável (NMP), para cada subárea, obtendo-se o NMP por grama de solo úmido por meio do programa MPNES (Woomer et al., 1990). Assim, com os dados de umidade, pôde-se obter o NMP por grama de solo seco. A análise de variância, assim como as pressuposições desse modelo (normalidade dos dados e resíduos e homogeneidade de variâncias), e testes de comparações múltiplas (Duncan a 5 %) foram feitos utilizando-se o programa R (R Development Core Team, 2009) e seu complementar Tinn R, seguindo-se a transformação dos dados para log(NMP). Cada área amostrada foi considerada tratamento (com cinco repetições por área/tratamento), sendo todas analisadas em delineamento inteiramente ao acaso.
Para obtenção dos isolados de BDNS, procedeu-se ao método utilizado por Melloni et al. (2004). Os isolados crescidos no meio batata foram caracterizados quanto a cor, diâmetro e consistência das colônias. Os isolados que apresentaram as mesmas características fenotípicas foram unidos em um grupo fenotípico cultural. Em função do número de isolados obtidos por grupo fenotípico, pôde-se calcular o índice de diversidade de Shannon & Weaver, que relaciona a riqueza e uniformidade de espécies (Odum, 1983), para cada área de estudo. As estirpes-tipo de Azospirillum lipoferum (BR11080), Azospirillum brasilense (BR11001), Azospirillum amazonense (BR11040), Herbaspirillum seropedicae (BR11175) e Burkholderia brasilensis (BR11340) também foram caracterizadas fenotipicamente em meio batata, para comparação com os isolados previamente obtidos das amostras de solo. Todos os isolados e estirpes-tipo foram agrupados em dendrograma de similaridade pelo método Complete Linkage e Distância Euclidiana (Everitt, 1993), por meio do programa STATISTICA 5.0 (Statistica, 1995), de acordo com os valores atribuídos para as características fenotípicas (Quadro 1).
Os resultados dos atributos microbiológicos foram submetidos à análise de componentes principais (PCA), utilizando-se o programa PC-ORD 3.12 (McCune & Mefford, 1997). Essa técnica é frequentemente utilizada para reduzir o número de variáveis totais para discussão, distribuídas de acordo com componentes principais (CP) de maior correlação dos dados (Baretta et al., 2008), tornando a análise mais eficiente e mantendo a maioria ou todas as informações originais (Reyment & Jöreskog, 1993).
O teste de tolerância à salinidade é recomendado por Nóbrega et al. (2004) para estudos de diversidade fenotípica desse grupo bacteriano e foi empregado no presente trabalho a fim de avaliar o comportamento dos isolados e estirpes-tipo em relação aos dendrogramas elaborados por meio de atributos ligados aos aspectos morfológicos das colônias. Para essa análise, selecionaram-se, no máximo, três isolados de cada grupo fenotípico cultural que apresentaram similaridade igual ou superior a 70 % com as estirpes-tipo, tanto no dendrograma de similaridade para a coleta de setembro/2006 quanto no dendrograma referente à coleta de abril/2007. Nesse teste, porções das colônias dos isolados e estirpes-tipo em estudo foram transferidas para microtubos de ensaio contendo 1,5 mL de meio batata líquido, sendo estes submetidos à agitação em temperatura ambiente por 3 dias, tempo necessário para que atingissem a fase logarítmica (Nóbrega et al., 2004). Alíquotas de 25 mL das suspensões de bactérias foram inoculadas e espalhadas em miniplacas de Petri contendo meio de cultura batata sólido, modificado por adição de NaCl com concentrações finais de 0, 10, 20, 40 e 80 g L-1, incubados a 28 ºC por 4 dias. Os tratamentos foram distribuídos ao acaso, com três repetições. A tolerância das bactérias ao NaCl foi observada por meio da porcentagem de crescimento da bactéria em relação à área superficial do meio de cultura. Assim como proposto por Trannin et al. (2001), foram atribuídos valores ou notas às médias dos percentuais de crescimento para cada concentração de NaCl: 1 (0-20 %), 2 (20-40 %), 3 (40-60 %), 4 (60-80 %) e 5 (80-100 %).
RESULTADOS E DISCUSSÃO
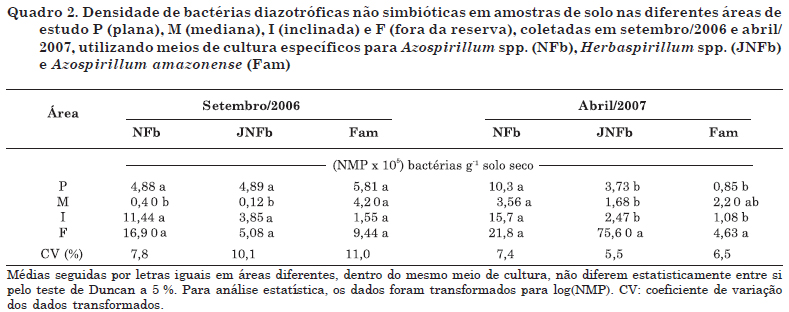
Para a coleta de setembro/2006 (Quadro 2), exceto a área com declividade mediana (M) que apresentou menor densidade de BDNS nos meios NFb e JNFb, não houve diferença significativa entre as demais nos três meios utilizados.
Na coleta de abril/2007 (Quadro 2), o maior valor de densidade foi obtida nos solos localizados fora da reserva no meio JNFb, o que possivelmente ocorreu devido à maior ocupação por hospedeiros (gramíneas) nesse local, em razão das condições ligadas à maior temperatura e umidade nesse período. Esse resultado corrobora aquele apresentado por Melloni et al. (2004), os quais encontraram forte correlação entre presença de gramíneas e densidade dessas bactérias, em solos de mineração em reabilitação, mostrando que a presença do hospedeiro é mais importante do que atributos físicos e químicos do solo. Segundo dados climatológicos da reserva, obtidos pelo CPTEC (2010), as condições climáticas do verão (290 mm e 22 ºC, respectivamente para precipitação total mensal e temperatura média) são comparativamente maiores que as do inverno (27 mm e 16 ºC) e podem estar ligadas à maior densidade bacteriana nesse período (Moreira & Siqueira, 2006). De acordo com Melloni (2007), temperaturas e umidades mais elevadas e revegetação com gramíneas do tipo capim-gordura e braquiária contribuíram para a maior multiplicação dessas BDNS e maior facilidade de isolamento em laboratório.
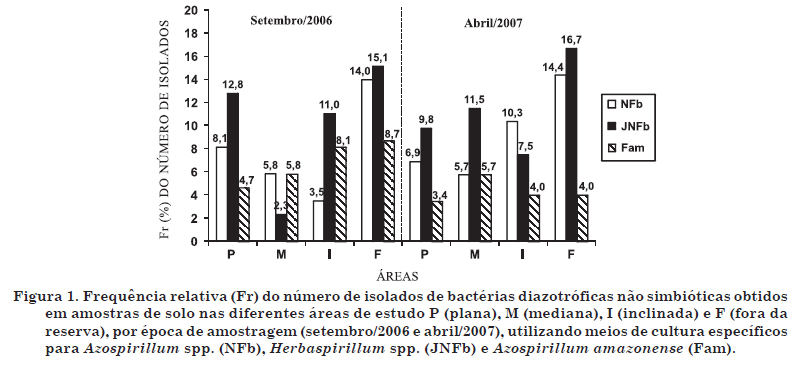
Notou-se efeito marcante no número obtido de isolados em amostras do solo fora da reserva (Figura 1), principalmente nos meios NFb e JNFb. De acordo com as características fenotípicas culturais de cada isolado, puderam-se estabelecer os grupos fenotípicos culturais por época de amostragem, conforme demonstrado nos quadros 3 e 4, juntamente com as descrições das características fenotípicas das estirpes-tipo. Dessa forma, foi observado grande número de grupos fenotípicos, onde 172 isolados foram distribuídos em 61 grupos (31, 16, 23 e 39 grupos nas áreas plana, mediana, inclinada e fora, respectivamente) para as amostras de setembro/2006, e 174 isolados em 37 grupos (16, 14, 19 e 23 nas áreas plana, mediana, inclinada e fora, respectivamente) para as amostras de abril/2007. O maior número de grupos fenotípicos em setembro/2006 esteve ligado aos maiores valores do índice de diversidade de Shannon & Weaver (H') (Quadros 3 e 4), em todas as áreas, quando comparados aos da segunda coleta. Assim como observado por Melloni et al. (2004), houve queda de cerca de 40 % no número de grupos na segunda época, evidenciando a dinâmica temporal dessa comunidade, em função das condições ligadas à temperatura e umidade, características de cada época do ano.
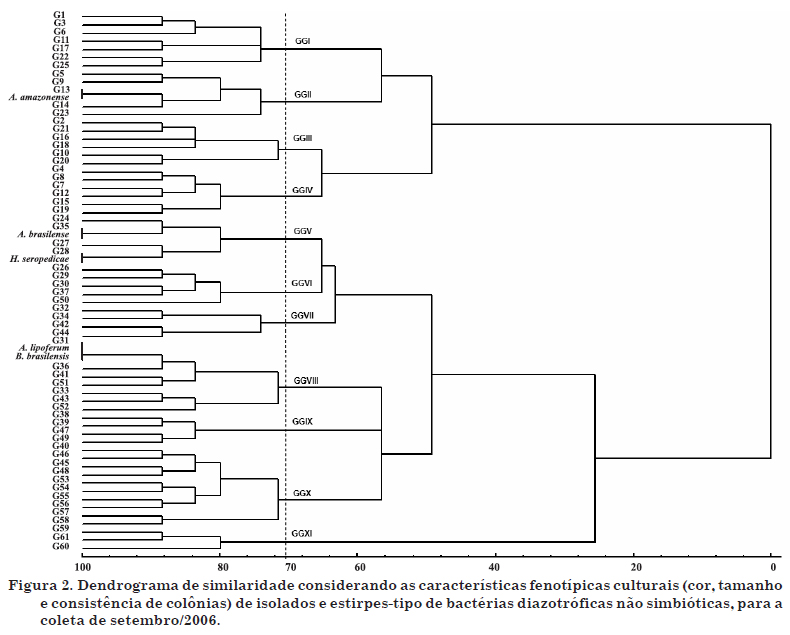
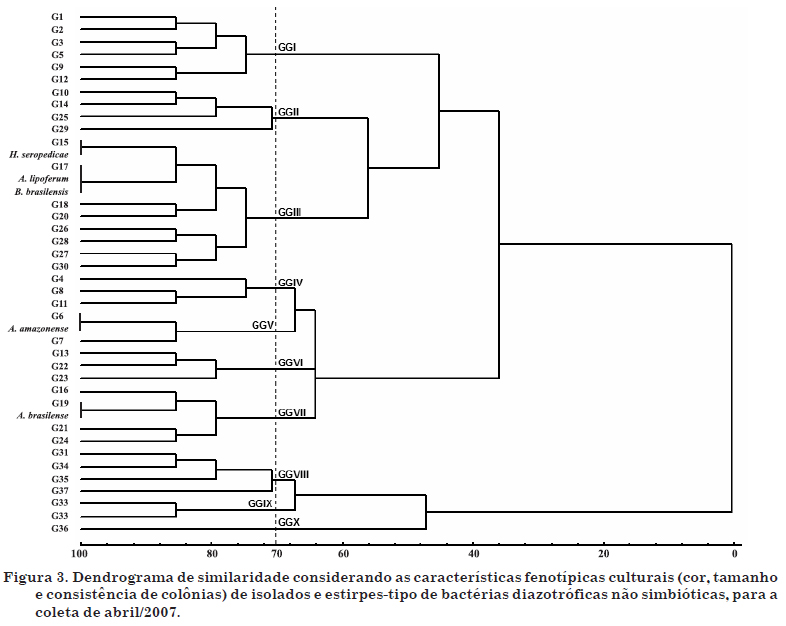
Os dendrogramas de similaridade estão representados nas figuras 2 e 3. Para análise da coerência taxonômica entre os grupos culturais, foi feito um estudo de similaridade igual ou superior a 70 %, onde se observaram 11 grandes grupos (GG) culturais no inverno e 10 no verão. Os GG são aqueles formados pela interseção com a linha de 70 % de similaridade. Dos 11 GG formados na época de inverno, três apresentaram similaridade igual ou superior a 70 % com as cinco estirpes-tipo, compreendendo 52 isolados. Na época de verão, três GG apresentaram similaridade igual ou superior a 70 % com as estirpes-tipo analisadas, com um total de 95 isolados. Esse resultado comprova a variação sazonal dos isolados em relação às suas características fenotípicas. Assim, pôde-se obter a distribuição dos grupos fenotípicos culturais que apresentaram similaridade igual ou superior a 70 % com as estirpes-tipo, por área analisada, para as duas épocas de amostragem (Quadro 5). O maior número de grupos fenotípicos culturais foi obtido na área fora da reserva (13 grupos), a qual apresentava, conforme já discutido, maior ocorrência de hospedeiros (braquiária) e, consequentemente, maior densidade radicular de gramíneas com contribuição ao aumento desses microrganismos (Melloni, 2007).
Do total de isolados obtidos nas épocas de inverno e verão, 30 % e 55 %, respectivamente, apresentaram similaridade igual ou superior a 70 % em relação às estirpes-tipo em estudo. Embora o maior número de isolados tenha sido obtido com a utilização do meio JNFb (Figura 1), a análise de similaridade (Figuras 2 e 3) indica que esses não necessariamente apresentaram maior similaridade a Herbaspirillum, concordando com os resultados obtidos por Melloni et al. (2004). Segundo esses autores, a falta de especificidade do meio JNFb, já considerada por Baldani et al. (1999), indica que o meio JNFb permite o crescimento de outras bactérias, principalmente aquelas capazes de tolerar a maior acidez inicial desse meio, como as espécies do gênero Azospirillum, o que pode explicar a similaridade igual ou superior a 70 % da estirpe-tipo H. seropedicae BR11175 com a estirpe A. brasilense BR11001 na época de inverno e com a estirpe A. lipoferum BR11080 na época de verão. Do mesmo modo, Magalhães & Döbereiner (1984) mostraram que, mesmo sendo as espécies de Azospirillum favorecidas nos meios NFb e Fam, outros microrganismos diazotróficos podem apresentar crescimento, como ocorreu no presente estudo, onde as estirpes de A. lipoferum BR11080 e B. brasilensis BR11340 apresentaram alta similaridade. A grande maioria dos isolados não apresentou similaridade com as estirpes-tipo em questão, podendo pertencer a outras espécies de diazotróficos não simbióticos.
Com relação ao teste de tolerância à salinidade (Figuras 4 e 5), na coleta de setembro/2006 (Figura 4), a maioria dos isolados selecionados do GGII apresentou queda no crescimento na concentração de 40 g L-1 de NaCl, comportamento bastante diferenciado daquele apresentado pela estirpe-tipo A. amazonense, cuja sensibilidade ao NaCl foi bem maior, apresentando redução no crescimento em concentrações de 10 g L-1, concordando com o resultado obtido por Nóbrega et al. (2004). Dessa forma, o comportamento dos isolados desse agrupamento é diferente daquele obtido pela característica fenotípica cultural a 70 % de similaridade, onde encontravam-se no mesmo GG da estirpe-tipo em questão.
No GGV, percebeu-se que a maior parte dos isolados apresentou redução de crescimento nas concentrações de 40 e 80 g L-1 de NaCl, o que corresponde ao comportamento das estirpes-tipo A. brasilense e H. seropedicae, mostrando tendência de mesmo comportamento fenotípico a 70 % de similaridade. Os isolados do GGVIII, de modo geral, apresentaram queda de crescimento nas concentrações de 40 e 80 g L-1, aproximando-se da observada para a estirpe-tipo B. brasilensis nessa primeira concentração. Dentro de um mesmo grupo fenotípico cultural, os isolados apresentaram comportamentos diferenciados, mostrando assim a diferenciação em relação ao comportamento fenotípico cultural.
Na coleta de abril/2007 (Figura 5), os isolados do GGIII apresentaram comportamentos variados até mesmo dentro do grupo fenotípico cultural, com queda acentuada de crescimento na concentração de 40 g L-1, semelhante ao das estirpes-tipo. Os isolados do GGV apresentaram comportamento diferenciado da estirpe-tipo A. amazonense, mostrando resistência maior que esta em relação ao NaCl. A maioria dos isolados do GGVII apresentou comportamento semelhante ao da estirpe-tipo A. brasilense, com queda de crescimento na concentração de 40 g L-1.
Dessa forma, nem sempre um agrupamento com base na tolerância à salinidade foi semelhante àquele obtido pelas características fenotípicas culturais (a 70 % de similaridade). Essa afirmativa pode ser confirmada ao se confrontar o comportamento das estirpes-tipo A. lipoferum e B. brasilensis, que, apesar de terem apresentado características fenotípicas culturais semelhantes, mostraram diferentes tolerâncias à salinidade.
Portanto, em estudos de avaliação da diversidade fenotípica de BDNS, quanto maior o número de atributos microbiológicos utilizados, maior a representatividade da classificação, motivo pelo qual se utilizou o teste de tolerância ao NaCl na caracterização de espécies bacterianas, conforme recomendado por Nóbrega et al. (2004). A estirpe-tipo Burkholderia brasilensis BR11340, no presente estudo, apresentou mesma tolerância à salinidade (2 %) que no estudo realizado pelos últimos pesquisadores. A discrepância entre os resultados das demais estirpes-tipo provavelmente ocorreu devido a diferentes composições químicas nos meios de cultura utilizados, que podem tanto interferir na disponibilidade do sal quanto favorecer o crescimento das bactérias. Segundo Bashan & Holguin (1997), quanto maior a capacidade de Azospirillum em tolerar o estresse salino, maior será o acúmulo de solutos orgânicos e mais espessa será a camada de lipopolissacarídeos envolvendo a bactéria fato que pode ser estendido às demais bactérias aqui estudadas.
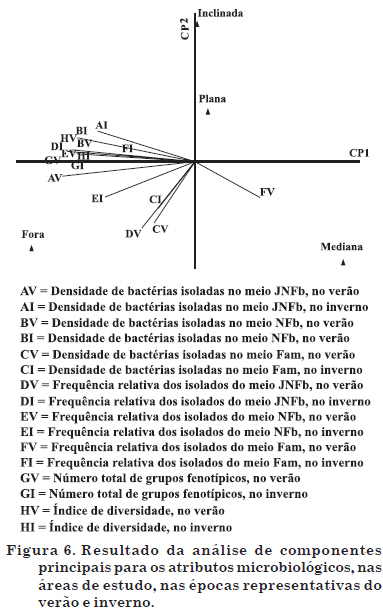
A análise multivariada (Figura 6) apresenta, de forma resumida, o comportamento dos atributos nas diferentes áreas de estudo. Os componentes principais CP1 e CP2 apresentaram 65,0 e 21,4 % da variância total dos dados, respectivamente. Em vista do elevado valor da variância total (86,4 %), somente esses dois CPs foram apresentados.
Com exceção do atributo "frequência relativa dos isolados do meio Fam, no verão" (FV), maior na área mediana dentro da reserva, todos os demais apresentaram maiores valores em amostras de solo da área de fora da reserva, independentemente da época de amostragem (verão ou inverno). Esse resultado confirma aqueles obtidos de densidade (Quadro 2), frequência relativa (Figura 1), índices de diversidade (Quadros 3 e 4) e total de isolados e de grupos funcionais (Quadro 5), bem como evidencia o efeito da cobertura vegetal do tipo gramíneas sobre todos os atributos avaliados, estimulando a densidade e diversidade de grupos fenotípicos, independentemente da época de amostragem do solo.
CONCLUSÕES
1. Maiores valores de densidade e diversidade fenotípica de bactérias diazotróficas não simbióticas foram obtidas na área de fora (entorno), quando comparadas a solos da Reserva, independentemente da época de amostragem de solo (verão e inverno).
2. A grande maioria dos isolados de bactérias diazotróficas não simbióticas não apresentou similaridade com as estirpes-tipo pesquisadas quanto às características fenotípicas culturais.
3. Novos estudos são sugeridos, visando à caracterização genética e avaliação do potencial do inóculo dos isolados de bactérias diazotróficas não simbióticas da Reserva e entorno em plantas de interesse ambiental e, ou, agrícola.
AGRADECIMENTOS
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e à Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG), pelo apoio.
LITERATURA CITADA
Recebido para publicação em março de 2010 e aprovado em janeiro de 2011.
- BALDANI, J.I.; CARUSO, L.; BALDANI, V.L.D.; GOI, S.R. & DÖBEREINER, J. Recent advances in BNF with non-legume plants. Soil Biol. Biochem., 29:922-928, 1997a.
- BALDANI, J.I.; AZEVEDO, M.S.; REIS, V.M.; TEIXEIRA, K.R.S.; OLIVARES, F.L.; GOI, S.R.; BALDANI, V.L.D. & DÖBEREINER, J. Fixação biológica de nitrogênio em gramíneas: Avanços e aplicações. In: SIQUEIRA, J.O.; MOREIRA, F.M.S.; LOPES, A.S.; GUILHERME, L.R.G.; FAQUIN, V.; FURTINI NETO, A.E. & CARVALHO, J.G., eds. Inter-relação fertilidade, biologia do solo e nutrição de plantas. Viçosa, MG, SBCS/UFLA/DCS, 1999. p.621-666.
- BALDANI, V.L.D.; OLIVEIRA, E.; BALOTA, E.; BALDANI, J.I.; KIRCHHOF, G. & DÖBEREINER, J. Burkholderia brasilensis sp. nov., uma nova espécie de bactéria diazotrófica endofítica. An. Acad. Bras. Ci., 69:116, 1997b.
- BALDOTTO, L.E.B.; BALDOTTO, M.A.; OLIVARES, F.L.; VIANA, A.P. & BRESSAN-SMITH, R. Seleção de bactérias promotoras de crescimento no abacaxizeiro cultivar 'Vitória' durante a aclimatização. R. Bras. Ci. Solo, 34:349-360, 2010.
- BARETTA, D.; BARETTA, C.R.D.M. & CARDOSO, E.J.B.N. Análise multivariada de atributos microbiológicos e químicos do solo em florestas com Araucaria angustifolia R. Bras. Ci. Solo, 32:2683-2691, 2008.
- BASHAN, Y. & HOLGUIN, G. Azospirillum-plant relationships: Enviromental and physiological advances (19901996). Can. J. Microbiol., 43:103-121, 1997.
- BAZZICALUPO, M. & OKON, Y. Associative and endophytic symbiosis. In: PEDROSA, F.; HUNGRIA, M.; YATES, M.G. & NEWTON, W.E., eds. Nitrogen fixation: From molecules to crop productivity. Dordrecht, Kluwer Academic Publishers, 2000. p.409-410.
- BUCHANAN, B.B.; GRUISSEM, W. & JONES, R.L. Biochemistry and molecular biology of plants. Rockville, American Society of Plant Physiologists, 2000. 1367p.
- CENTRO DE PREVISÃO DE TEMPO E ESTUDOS CLIMÁTICOS CPTEC. Plataformas de coleta de dados. Disponível em: <http://satelite.cptec.inpe.br/PCD/historico/consulta_pcdm.jsp>. Acesso em 6 de maio de 2010.
- DOBBELAERE, S.; VANDERLEYDEN, J. & OKON, Y. Plant growth-promoting effects of diazotrophs in the rhizosphere. Crit. Rev. Plant Sci., 22:107-149, 2003.
- DÖBEREINER, J. Recent changes in concepts of plant bacteria interactions: Endophytic N2 fixing bacteria. Ci. Cult., 44:310313, 1992.
- DÖBEREINER, J.; BALDANI, V.L.D. & BALDANI, J.I. Como isolar e identificar bactérias diazotróficas de plantas não-leguminosas. Itaguaí, Embrapa- CNPAB, 1995. 60p.
- EVERITT, B.S. Cluster analysis. New York, J. Wiley, 1993.
- FERREIRA, J.S.; BALDANI, J.I. & BALDANI, V.L.D. Seleção de inoculantes à base de turfa contendo bactérias diazotróficas em duas variedades de arroz. Acta Sci. Agron., 32:179-185, 2010.
- HUNGRIA, M.; FRANCHINI, J.C.; CAMPO, R.J.; CRISPINO, C.C.; MORAES, J.Z.; SIBALDELLI, R.N.R.; MENDES, I.C. & ARIHARA, J. Nitrogen nutrition of soybean in Brazil: Contributions of biological N2 fixation and of N fertilizer to grain yield. Can. J. Plant Sci., 86:927-939, 2006.
- ITAJUBÁ. Prefeitura Municipal. Elaboração do Plano de Manejo: Reserva Biológica Serra dos Toledos. Itajubá, Documento apresentado ao Fundo Nacional do Meio Ambiente_Edital 10/2001: Apoio à Gestão Integrada em Unidades de Conservação de Proteção Integral de Reservas Particulares do Patrimônio Natural, 2002a. (Administração 2001-2004)
- ITAJUBÁ. Secretaria Municipal da Educação. Atlas escolar, histórico e geográfico do Município de Itajubá. Itajubá, 2002b. (Administração 2001-2004)
- KENNEDY, I.R.; CHOUDHURY, A.T.M.A. & KECSKÉS, M.L. Non-symbiotic bacterial diazotrophs in crop-farming systems: Can their potential for plant growth promotion be better exploited? Soil Biol. Biochem., 36:1229-1244, 2004.
- MAGALHÃES, F.M.M. Caracterização e distribuição de uma nova espécie de bactéria fixadora de nitrogênio. Manaus, Universidade do Amazonas, 1983. 89p. (Tese de Mestrado)
- MAGALHÃES, F.M.M. & DÖBEREINER, J. Ocorrência de Azospirillum amazonense em alguns ecossistemas da Amazônia. R. Microbiol., 15:246-252, 1984.
- MARIN, V.A.; BALDANI, V.L.D.; TEIXEIRA, K.R.S. & BALDANI, J.I. Fixação biológica de nitrogênio de importância para a agricultura tropical. Seropédica, Embrapa Agrobiologia, 1999 (Série Documentos).
- McCUNE, B. & MEFFORD, M.J. Multivariate analysis of ecological data. Version 3.12. Gleneden Beach, MjM Software, 1997.
- MELLONI, R. Quantificação microbiana da qualidade do solo. In: SILVEIRA, A.P.D. & FREITAS, S.S., eds. Microbiota do solo e qualidade ambiental. Campinas, Instituto Agronômico, 2007. p.193-218.
- MELLONI, R.; NÓBREGA, R.S.A.; MOREIRA, F.M.S. & SIQUEIRA, J.O. Densidade e diversidade de bactérias diazotróficas endofíticas em solos de mineração de bauxita em reabilitação. R. Bras. Ci. Solo, 28:85-93, 2004.
- MOREIRA, F.M.S & SIQUEIRA, J.O. Microbiologia e bioquímica do solo. 2.ed. Lavras, Universidade Federal de Lavras, 2006. 729p.
- NASSAR, E.A. Influência do relevo na profundidade de solos da Reserva Biológica Serra dos Toledos. Itajubá, Universidade Federal de Itajubá, 2007. 142p. (Trabalho de Conclusão de Curso)
- NÓBREGA, R.S.A.; MOREIRA, F.M.S.; SIQUEIRA, J.O. & LIMA, A.S. Caracterização fenotípica e diversidade de bactérias diazotróficas associativas isoladas de solos em reabilitação após a mineração de bauxita. R. Bras. Ci. Solo, 28:269-279, 2004.
- ODUM, E.P. Ecologia. Rio de Janeiro, Guanabara, 1983.
- OLIVARES, F.L.; BALDANI, J.I.; JAMES, E.K. & DÖBEREINER, J. Infection of mottled stripe disease and resistant sugar varieties by endophitic diastrophic Herbaspirillum New Phytol., 135:723-737, 1997.
- OLIVEIRA, A.L.M.; STOFFELS, M.; SCHMID, M.; REIS, V.M.; BALDANI, J.I. & HARTMANN, A. Colonization of sugarcane plantlets by mixed inoculations with diazotrophic bacteria. Eur. J. Soil Biol., 45:106-113, 2009.
- PERIN, L.; MARTINEZ-AGUILAR, L.; PAREDESVALDEZ, G.; BALDANI, J.I.; ESTRADA DE LOS SANTOS, P.; REIS, V.M. & CABALLERO-MELLADO, J. Burkholderia silvatlantica sp. nov., a diazotrophic bacterium associated with sugar cane and maize. Int. J. Syst. Evol. Microbiol., 56:1931-1937, 2006.
- R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, 2009. Disponível em: <URL http://www.R-project.org>
- REIS JUNIOR, F.B.; SILVA, L.G.; REIS, V.M. & DÖBEREINER, J. Ocorrência de bactérias diazotróficas em diferentes genótipos de cana-de-açúcar. Pesq. Agropec. Bras., 35:985-994, 2000.
- REIS JUNIOR, F.B.; SILVA, M.F.; TEIXEIRA, K.R.S.; URQUIAGA, S. & REIS, V.M. Identificação de isolados de Azospirillum amazonense associados à Brachiaria spp. em diferentes épocas e condições de cultivo e produção de fitohormônio pela bactéria. R. Bras. Ci. Solo, 28:103-113, 2004.
- REIS JUNIOR, F.B.; MACHADO, C.T.T.; MACHADO, A.T. & SODEK, L. Inoculação de Azospirillum amazonense em dois genótipos de milho sob diferentes regimes de nitrogênio. R. Bras. Ci. Solo, 32:1139-1146, 2008.
- REYMENT, R. & JÖRESKOG, K.G. Applied factor analysis in the natural sciences. Cambridge, Cambridge University Press, 1993.
- RODRIGUES, L.S.; BALDANI, V.L.D.; REIS, V.M. & BALDANI, J.I. Diversidade de bactérias diazotróficas endofíticas dos gêneros Herbaspirillum e Burkholderia na cultura do arroz inundado. Pesq. Agropec. Bras., 41:275-284, 2006.
- SALA, V.M.R.; FREITAS, S.S.; DONZELI, V.P.; FREITAS, J.G.; GALLO, P.B. & SILVEIRA, A.P.D. Ocorrência e efeito de bactérias diazotróficas em genótipos de trigo. R. Bras. Ci. Solo, 29:345-352, 2005.
- SALA, V.M.R.; CARDOSO, E.J.B.N.; FREITAS, J.G. & SILVEIRA, A.P.D. Resposta de genótipos de trigo à inoculação de bactérias diazotróficas em condições de campo. Pesq. Agropec. Bras., 42:833-842, 2007.
- STATISTICA. Statistic Analysis. Versão 5.0. Cary, 1995.
- TRANNIN, I.C.B.; MOREIRA, F.M.S.; SIQUEIRA, J.O. & LIMA, A. Tolerância de estirpes e isolados de Bradyrhizobium e de Azorhizobium a zinco, cádmio e cobre "in vitro". R. Bras. Ci. Solo, 25:305-316, 2001.
- WOOMER, P.; BENNET, J. & YOST, R. Overcoming the inflexibility of most-probable-number procedures. Agron. J., 82:349-353, 1990.
Datas de Publicação
-
Publicação nesta coleção
30 Maio 2011 -
Data do Fascículo
Abr 2011
Histórico
-
Recebido
Mar 2010 -
Aceito
Jan 2011