Resumos
Apesar de ainda existirem ecossistemas altomontanos no Paraná em excelente estado de conservação, iminentes ameaças antrópicas e a fragilidade desses ambientes têm sido motivos de preocupação. Este trabalho teve os seguintes objetivos: caracterizar solos de área representativa dos campos e florestas altomontanas ocorrentes na Serra da Igreja; apontar quais os possíveis fatores pedológicos que resultam nessas diferentes fitotipias; e caracterizar algumas das suas funcionalidades ambientais (estoque de C e de água). Os principais solos encontrados nos campos foram Organossolos Fólicos fíbricos/sápricos (líticos e típicos) e Organossolos Háplicos fíbricos/sápricos (típicos e térricos) e, nas florestas altomontanas, Gleissolos Háplicos alíticos típicos. Ambas as classes são de solos distróficos, extremamente ácidos, com alta saturação por Al trocável e altos teores de C orgânico total. A distribuição das florestas altomontanas está fortemente controlada por vales e colos de cumeeiras, os quais estão sujeitos a processos morfogenéticos que resultam em solos com horizontes minerais. Já os campos estão estabelecidos em topos, onde processos pedogenéticos promoveram espessamento de horizontes hísticos, os quais, em função de suas características intrínsecas, aliadas aos fortes ventos, parecem conter com sucesso o avanço da floresta sobre o campo. Os estoques de C por unidade de área nos solos dos campos são superiores aos dos solos das florestas altomontanas, sendo ambos considerados altos quando comparados aos dados de outros ecossistemas, sendo duas a três vezes maiores do que os encontrados em solos de ecossistemas de altitudes mais baixas na mesma latitude. Também foi constatada alta capacidade de retenção hídrica devido à porosidade total verificada nos horizontes hísticos, os quais têm o potencial de reter em média 12 vezes seu volume em água.
campos altomontanos; Floresta Ombrófila Densa Altomontana; estoques de carbono no solo; Organossolos; retenção hídrica do solo
Although in Paraná some upper montane ecosystems are still in excellent conditions, imminent anthropogenic threats and the vulnerability of these environments have raised concern. The objectives of this study were: (1) to characterize representative soils of high altitude grasslands and upper montane rain forests in the Serra da Igreja; (2) identify the possible pedological factors underlying the different vegetation types and; (3) describe some of the environmental roles of the soils of these areas (carbon stock and water retention). The main soil types of the grasslands were fibric/sapric Folic and Haplic Histosols and the upper montane forests typic alitic Haplic Gleysols. Both soils are dystrophic, extremely acid, with high exchangeable Al saturation and very high levels of total organic C. The upper montane forests are mostly found in valleys influenced by morphogenetic processes that result in soils with mineral horizons. The grasslands are located on flat tops, where pedogenetic processes led to thicker histic horizons. This fact, combined with a great wind exposure, seem to prevent the normal expansion of tree species. Carbon stocks per unit area in the grassland soils are higher than those of the upper montane forests. Furthermore, C values in both areas are two- to threefold higher than those reported for soil ecosystems at low altitudes at the same latitude. Another important environmental feature of these soils is the high water retention capacity, due to the total porosity values in the histic horizons, which can retain water up to approximately 12 times their volume.
High altitude grasslands; montane tropical rain forest; soil carbon stocks; histosols; soil water retention
COMISSÃO 1.2 - LEVANTAMENTO E CLASSIFICAÇÃO DO SOLO
Funcionalidades ambientais de solos altomontanos na Serra da Igreja, Paraná1 1 Parte da tese de Doutorado do primeiro autor apresentada ao Programa de Pós-Graduação em Engenharia Florestal, Univer-sidade Federal do Paraná - UFPR. Recebido para publicação em 12 de maio de 2010 e aprovado em 25 de março de 2011.
Environmental functionalities of upper montane soils in Serra da Igreja, southern Brazil
Maurício Bergamini ScheerI, II; Gustavo Ribas CurcioIII; Carlos Vellozo RoderjanIII
IPesquisador da Assessoria de Pesquisa e Desenvolvimento - APD/DMA da Companhia de Saneamento do Paraná - SANEPAR. Rua Engenheiros Rebouças 1376, Rebouças, CEP 80215-900 Curitiba (PR), Brasil. E-mail: mauriciobs@sanepar.com.br
IIEmpresa Brasileira de Pesquisa Agropecuária - Embrapa Florestas. Estrada da Ribeira, km 111, Caixa Postal 319, CEP 83411-0 Colombo (PR), Brasil. E-mail: curcio@embrapa.cnpf.br
IIIProfessor do Programa de Pós-Graduação em Engenharia Florestal, Universidade Federal do Paraná - UFPR. Av. Pref. Lothário Meissner 900, Jardim Botânico, Campus III. CEP 80210-170 Curitiba (PR), Brasil. E-mail: roderjan@ufpr.br
RESUMO
Apesar de ainda existirem ecossistemas altomontanos no Paraná em excelente estado de conservação, iminentes ameaças antrópicas e a fragilidade desses ambientes têm sido motivos de preocupação. Este trabalho teve os seguintes objetivos: caracterizar solos de área representativa dos campos e florestas altomontanas ocorrentes na Serra da Igreja; apontar quais os possíveis fatores pedológicos que resultam nessas diferentes fitotipias; e caracterizar algumas das suas funcionalidades ambientais (estoque de C e de água). Os principais solos encontrados nos campos foram Organossolos Fólicos fíbricos/sápricos (líticos e típicos) e Organossolos Háplicos fíbricos/sápricos (típicos e térricos) e, nas florestas altomontanas, Gleissolos Háplicos alíticos típicos. Ambas as classes são de solos distróficos, extremamente ácidos, com alta saturação por Al trocável e altos teores de C orgânico total. A distribuição das florestas altomontanas está fortemente controlada por vales e colos de cumeeiras, os quais estão sujeitos a processos morfogenéticos que resultam em solos com horizontes minerais. Já os campos estão estabelecidos em topos, onde processos pedogenéticos promoveram espessamento de horizontes hísticos, os quais, em função de suas características intrínsecas, aliadas aos fortes ventos, parecem conter com sucesso o avanço da floresta sobre o campo. Os estoques de C por unidade de área nos solos dos campos são superiores aos dos solos das florestas altomontanas, sendo ambos considerados altos quando comparados aos dados de outros ecossistemas, sendo duas a três vezes maiores do que os encontrados em solos de ecossistemas de altitudes mais baixas na mesma latitude. Também foi constatada alta capacidade de retenção hídrica devido à porosidade total verificada nos horizontes hísticos, os quais têm o potencial de reter em média 12 vezes seu volume em água.
Termos de indexação: campos altomontanos, Floresta Ombrófila Densa Altomontana, estoques de carbono no solo, Organossolos, retenção hídrica do solo.
SUMMARY
Although in Paraná some upper montane ecosystems are still in excellent conditions, imminent anthropogenic threats and the vulnerability of these environments have raised concern. The objectives of this study were: (1) to characterize representative soils of high altitude grasslands and upper montane rain forests in the Serra da Igreja; (2) identify the possible pedological factors underlying the different vegetation types and; (3) describe some of the environmental roles of the soils of these areas (carbon stock and water retention). The main soil types of the grasslands were fibric/sapric Folic and Haplic Histosols and the upper montane forests typic alitic Haplic Gleysols. Both soils are dystrophic, extremely acid, with high exchangeable Al saturation and very high levels of total organic C. The upper montane forests are mostly found in valleys influenced by morphogenetic processes that result in soils with mineral horizons. The grasslands are located on flat tops, where pedogenetic processes led to thicker histic horizons. This fact, combined with a great wind exposure, seem to prevent the normal expansion of tree species. Carbon stocks per unit area in the grassland soils are higher than those of the upper montane forests. Furthermore, C values in both areas are two- to threefold higher than those reported for soil ecosystems at low altitudes at the same latitude. Another important environmental feature of these soils is the high water retention capacity, due to the total porosity values in the histic horizons, which can retain water up to approximately 12 times their volume.
Index terms: High altitude grasslands, montane tropical rain forest, soil carbon stocks, histosols, soil water retention.
INTRODUÇÃO
A matéria orgânica do solo representa a maior parte do C orgânico continental, e seus estoques até 1 m de profundidade alcançam aproximadamente 1.500 x 10 15g (Pg = petagramas) (Batjes, 1996; Jobbagy & Jackson, 2000). Considerando o solo sob vegetação nativa dentro dos 30 cm superficiais, o Brasil estoca 5 % dessa reserva de C mundial (Batjes, 1996), indicando que mesmo um único país pode ter grande influência nos fluxos de C global (Bernoux et al., 2002).
Uma vez que comportam horizontes hísticos e húmicos, riquíssimos em matéria orgânica, os solos de ecossistemas altomontanos desempenham importante papel na imobilização de C, bem como na regulação de fluxos hídricos (Bruijnzeel, 2000; Simas et al., 2005; Benites et al., 2007). O grande acúmulo de matéria orgânica nesses ambientes deve-se às condições desfavoráveis para a decomposição microbiana, como baixas temperaturas, alta umidade, baixa respiração, falta de nutrientes disponíveis e altos níveis de Al (Zech et al., 1997; Simas et al., 2005; Benites et al., 2007). O acúmulo de compostos orgânicos fortemente humificados no solo, na forma relativamente estável de complexos organometálicos, incluindo principalmente Al e Fe, dificulta que os microrganismos utilizem essa matéria orgânica como substrato (Volkoff et al., 1984; Zech et al., 1997; Simas et al., 2005; Don et al., 2007).
As substâncias húmicas contribuem para aumentar a quantidade de nutrientes e a capacidade de troca catiônica e de retenção de água no solo, que, por sua vez, propiciam o desenvolvimento da vegetação, a qual devolve a matéria orgânica com a deposição de sua fitomassa aérea e morte das raízes, garantindo a sustentabilidade desses ambientes (Simas et al., 2005; Benites et al., 2007). Durante períodos chuvosos os solos altomontanos são encharcados, sendo gradualmente drenados ao longo do tempo, regulando os fluxos hídricos das cabeceiras das bacias hidrográficas e, juntamente com a vegetação, minimizando riscos de erosão (Bruijnzeel & Proctor, 1995; Benites et al., 2007; Martínez et al., 2009). Em períodos menos chuvosos, a interceptação direta da água presente nas nuvens, comum em ambientes altomontanos (Hamilton et al., 1995; Bruijnzeel & Proctor, 1995), pode ser retida pela vegetação e solos sotopostos e encaminhada lentamente às nascentes dos rios logo a jusante, evitando-se maiores perdas.
Estimativas dos estoques de C nos solos em escalas regionais, nacionais e globais têm enorme importância para se ter acesso às mudanças nos fluxos de C (Batjes & Dijkshoorn, 1999; Yimer et al., 2006). Para melhorar a capacidade de prever, de propor e de executar ações para amenizar as consequências das mudanças climáticas globais relacionadas aos gases de efeito estufa, a sociedade depende, em parte, do conhecimento desses estoques em diferentes tipos de solos e de vegetação e dos fatores que influenciam sua distribuição (Batjes & Dijkshoorn, 1999; Bernoux et al., 2002; Novaes Filho et al., 2007). Considerando que grande parte da matéria orgânica desses solos está nos 30 cm superficiais, há grande potencial de liberação de CO2 para a atmosfera caso a vegetação seja queimada ou cortada e a área convertida para a agricultura ou pecuária (Yiemer et al., 2006). A perda de ecossistemas naturais pode implicar significante prejuízo econômico para a sociedade em termos de serviços ecossistêmicos prestados pela matéria orgânica do solo (Martínez et al., 2009). Essa necessidade elevou o número de estudos com esse propósito para diversas áreas no mundo (Garcia-Pausas et al., 2007).
Solos de ambientes altomontanos ainda são muito pouco estudados (Bernoux et al., 2002; Simas et al., 2005; Becker et al., 2007; Arteaga et al., 2008) e, por isso, sua importância ainda não é devidamente reconhecida (Benites et al., 2007).
Na região Sul do Brasil, alguns trabalhos caracterizaram solos sob florestas altomontanas (Roderjan, 1994; Ghani, 1996; Rocha, 1999; Portes, 2000, Falkenberg, 2003; Wisniewski et al., 2005; Vashchenko et al., 2007), e informações sobre solos de campos altomontanos ainda são mais escassas (Wisniewski et al., 2005; Vashchenko et al., 2007).
Segundo os trabalhos realizados no Paraná, para ambas as formações altomontanas, os solos são pouco desenvolvidos, extremamente ácidos, com altos teores de C, baixa saturação por bases e alta saturação por Al trocável. Essas informações são semelhantes às de solos de diferentes regiões altomontanas do Brasil, em diferentes litotipias e tipos vegetacionais, como complexos rupestres de altitude na região Sudeste (Simas et al., 2005; Benites et al., 2007; Silva et al., 2009) e florestas nebulares nos Aparados da Serra Geral, em Santa Catarina (Falkenberg, 2003).
No entanto, no Paraná ainda não existem trabalhos específicos sobre solos de florestas altomontanas ocorrentes sobre substratos graníticos, muito menos sobre solos de campos altomontanos que abranjam as diversas feições geomórficas apresentadas ao longo de suas áreas de ocorrência originais.
Dando continuidade aos estudos sobre ambientes altomontanos no Paraná, realizados por Mocochinski & Scheer (2008), por Scheer & Mocochinski (2009) e por Scheer (2010), os objetivos deste trabalho foram: caracterizar solos de área representativa dos campos altomontanos e da Floresta Ombrófila Densa Altomontana ocorrentes na Serra da Igreja; apontar quais os possíveis fatores pedológicos que resultam nas diferentes fitotipias; e estimar seus potenciais de estocar C e água.
MATERIAL E MÉTODOS
Área de estudo
A área de estudo está localizada em um dos picos da Serra da Igreja, subserra da Serra do Mar, a 1.335 m s.n.m., coordenadas 25 ° 36 ' S e 48 ° 51 ' W (Figura 1). A Serra da Igreja está inserida na Área de Proteção Ambiental - APA de Guaratuba, nos municípios de Guaratuba, Morretes e São José dos Pinhais. Assim como a maioria das montanhas da Serra do Mar no Paraná, suas porções superiores são formadas por granitos alcalinos encaixados em terrenos metamórficos de alto grau com contatos claramente falhados, em consonância com sua origem tectônica (MINEROPAR, 2001). O Granito Serra da Igreja, como outros granitos da Serra do Mar, é incidido por falhamentos do Ciclo Brasiliano/Pan-Africano (direção NE-SW) e do Arco de Ponta Grossa (na direção NW-SE) (Melo et al., 2000).
O topo da citada serra é constituído por duas fitotipias: florestal (Floresta Ombrófila Densa) e estépica (campos). Os campos estão localizados em topos de cumeeira, com ausência de pedregosidade e rochosidade, em relevos convexados, suave ondulados a forte ondulados, dominantemente em feições geomórficas de divergência hídrica. A floresta ocupa as posições de colo de cumeeira relacionadas a lineamentos geológicos, em relevos forte ondulados e montanhosos, explicitamente em feições de convergência hídrica (Figuras 2 e 3).
Nesta serra, a Floresta Ombrófila Densa Atlântica cobre sua maior extensão, atingindo, em sua formação altomontana, as porções mais elevadas das encostas, acima de 1.200 m s.n.m., estando bem preservada devido ao seu difícil acesso. Em alguns trechos, nas encostas a oeste das montanhas do lado ocidental da serra, ocorre o ecótono entre essa e a Floresta Ombrófila Mista Montana, a aproximadamente 1.100 m s.n.m. Já nas porções mais elevadas das montanhas, a partir dos 1.300 m s.n.m. passam a ocorrer campos altomontanos em excelente estado de conservação, sem indícios de interferência antrópica.
O clima das florestas altomontanas da Serra do Mar paranaense é classificado como Cfb, segundo sistema de Köppen, sendo subtropical, sempre úmido e com a temperatura média do mês mais frio abaixo de 18 °C e superior a - 3 ºC, e a média do mês mais quente, inferior a 22 ºC (Roderjan, 1994). Roderjan & Grodski (1999) observaram temperatura mínima absoluta de - 5 ºC, média anual de 13,4 ºC e máxima absoluta de 30 ºC para patamares altomontanos a 1.385 m s.n.m., em ambiente florestal, no Morro do Anhangava, no município de Quatro Barras, a aproximadamente 25 km ao norte da Serra da Igreja, no Paraná. As precipitações na Serra do Mar são bem distribuídas ao longo do ano e apresentam grande variação em função da topografia local. Medições na região litorânea ultrapassam 2.000 mm anuais, e nas encostas da serra os valores chegam a 3.500 mm (Maack, 2002).
Os pontos amostrais fizeram parte de transecto sobreposto a cumeeiras aguçadas (1.335 m s.n.m.), em campos altomontanos sobre Organossolos dispostos em rampas convexas divergentes de pequeno dimensionamento geográfico (1,5 ha), os quais, após abrupta ruptura de declive, dão lugar a rampas côncavas- convergentes, nas classes de declive forte ondulado e montanhoso, constituídas por Gleissolos com horizontes hísticos sob floresta altomontana (Figuras 1 e 2).
Os campos altomontanos na área de estudo são constituídos por plantas herbáceas com aproximadamente 70 cm de altura, onde é dominante Lagenocarpus triquetrus (Boeck.) Kuntze (Cyperaceae), além de arbustos de pequeno porte, como Croton mullerianus L.R. Lima (Euphorbiaceae) (em média, 50 cm de altura) e Tibouchina dubia (Cham.) Cogn. (Melastomataceae), entre outros. Espécies arbóreas com participação na estrutura das florestas altomontanas adjacentes com altura aproximada de 5 m, como Myrsine altomontana M.F. Freitas & Kin.-Gouv. (Myrsinaceae) e Tabebuia catarinensis A. Gentry (Bignoniaceae), também ocorrem nos campos, porém esparsamente, com pequena cobertura e na forma de arbustos com altura média de 60 cm. Em levantamento florístico realizado no local por Mocochinski & Scheer (2008), foram detectadas 99 espécies de plantas vasculares em área aproximada de 1,5 ha.
A mudança de vegetação de campo para floresta é abrupta, observando-se área ecotonal com fisionomia arbustiva com espécies das duas formações num gradiente de poucos metros (2 a 5 m). Segundo Scheer (2010), Siphoneugena reitzii D. Legrand (Myrtaceae), Ilex microdonta Reissek (Aquifoliaceae) e Drimys angustifolia Miers. (Winteraceae) perfazem 50 % da percentagem de importância fitossociológica arbórea (PAP > 10 cm). A área basal é de 30,5 m2 ha-1, e a densidade, de 4.696 ind ha-1.
Coleta de amostras e análises dos dados
Foram inicialmente realizadas tradagens no solo em forma sequencial, em transecto abrangendo campos e florestas altomontanas (Figuras 1 e 2). Após mapeamento de solos detalhado, foram abertos, em sequência, três perfis de solos, sendo, respectivamente, dois Organossolos (no campo) e um Gleissolo (na floresta). Amostras complementares superficiais (de 0 a 20 cm) e subsuperficiais (de 20 a 40 cm), também foram coletados com trado tipo holandês. Nesses perfis foram realizadas descrições morfológicas, definidos os horizontes diagnósticos e coletadas amostras para fins de classificação dos solos, seguindo o procedimento descrito por Santos et al. (2005). A classificação dos solos foi realizada de acordo com Embrapa (2006).
No campo, para analisar o estádio de decomposição da matéria orgânica, foi utilizada a escala de Von Post descrita por Embrapa (2006).
A partir da fração terra fina seca ao ar (TFSA) foram realizadas as análises físico-químicas. As análises granulométricas foram realizadas somente para a classificação de textura, segundo método do densímetro (Embrapa, 1997). Para amostras de horizontes hísticos, foram apenas determinados os percentuais de argila. As análises químicas, seguindo o método da Embrapa (1997), envolveram a determinação de pH em CaCl2, cátions trocáveis (Ca, Mg e K), acidez potencial (Al e H) e P disponível extraído em Mehlich-1. Foram obtidos os índices: soma de bases (S), capacidade de troca catiônica (T), saturação por bases (V %) e saturação por Al trocável (m %).
Amostras indeformadas também foram coletadas nos perfis por meio de anéis volumétricos e enviadas ao laboratório para caracterizações físico-hídricas, sendo determinadas as variáveis densidade global, o percentual de água disponível, a permeabilidade saturada, a porosidade total, a de aeração e a macroporosidade, segundo Embrapa (1997).
A determinação de C orgânico total por via seca foi realizada no Laboratório de Isótopos Estáveis do Centro de Energia Nuclear na Agricultura (CENA).
O estoque de C no solo (em kg C m-2) foi calculado segundo Batjes (1996) e Yimer et al. (2006). Primeiramente, foi multiplicado o teor de C orgânico total (em g kg-1) de cada sub-horizonte pela densidade do solo (em kg m-3) e pela sua espessura (em m). A densidade de partículas grosseiras ( > 2 mm), geralmente subtraídas da densidade do solo para utilizar a densidade somente da terra fina (fração < 2 mm) (Garcia-Pausas et al., 2007; Don et al., 2007), não foi necessária devido à sua inexistência ou a valores desprezíveis. Os dados foram extrapolados para Mg ha- 1. Nos pontos sequenciais analisados por tradagens, houve também a definição da espessura dos horizontes e foram calculados os estoques de C com base nos teores e densidades correspondentes aos perfis mais próximos e com características similares.
No campo altomontano, um levantamento prévio de forma sistemática (malha com 52 tradagens) em todo o topo da cumeeira estudada (dados não publicados) foi utilizado para validar a profundidade média obtida para a estimativa do potencial de estoque de C no solo.
Apenas como simples aproximações para extrapolar os possíveis estoques de C, por meio de fotos aéreas de 1980, na escala 1:25.000, do Instituto de Terras e Cartografia (ITC-PR), obteve-se uma estimativa da área dos campos altomontanos na Serra da Igreja. Para a floresta altomontana, além da fotointerpretação, foi utilizado como base o levantamento do Pró-Atlântica (2002). Com base nas áreas obtidas pelo Pró-Atlântica (2002), também foram estimados os estoques potenciais de C para essas duas fitotipias na Serra do Mar no Paraná. No entanto, ressalta-se que no presente trabalho foram apenas estimados os potenciais de estoque de C, sendo necessárias estimativas em escalas de maior detalhe, bem como considerar demais tipos de solos observados em outras porções da serra.
Para comparar as propriedades químicas entre os principais horizontes superficiais (0 a 20 cm) e subsuperficiais (20 a 40 cm) e os estoques de C no solo do campo e da floresta altomontana, foram realizados testes de U de Mann-Whitney (Wilcoxon). Para esses testes não paramétricos, as medianas são comparadas par a par. Foram calculados coeficientes de correlação de Spearman (Rs) para verificar o comportamento dos parâmetros químicos ao longo das profundidades nos perfis.
RESULTADOS E DISCUSSÃO
Solos dos campos altomontanos na Serra da Igreja
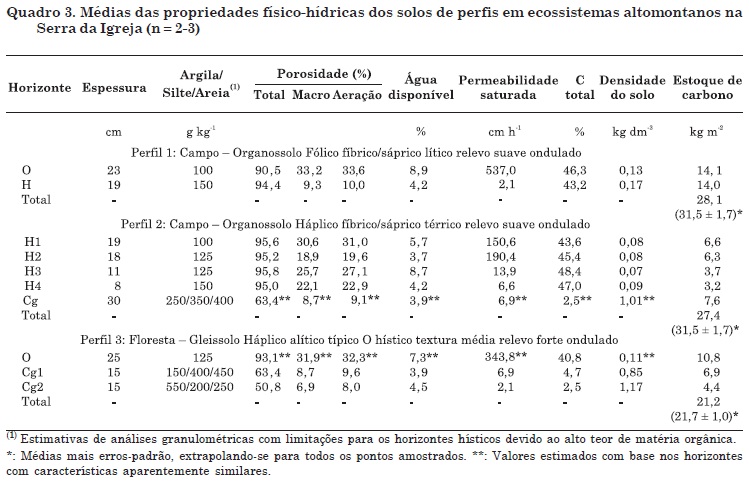
Nos solos dos campos altomontanos foram identificados apenas Organossolos (Figuras 1 e 2, Quadro 1). São constituídos essencialmente, ou na maior parte, por horizontes hísticos (O e H) com altos teores de C orgânico total (446,6 g kg-1). Apresentam cores úmidas pretas N2/ (úmidas) mais próximas à superfície e 10YR 2/1 (preto) a 3/2 (bruno-acinzentado muito escuro) nos demais sub-horizontes hísticos. O solo do perfil 1 foi classificado como Organossolo Fólico fíbrico/sáprico lítico, e o do perfil 2, como Organossolo Háplico fíbrico/sáprico térrico. O perfil 1, apesar de considerado solo hidromórfico sazonal, encontra-se na bordadura de topo aplainado, em local mais bem drenado do que o perfil 2. É composto por dois sub-horizontes hísticos (O e H) em contato lítico (Quadro 1).
No Organossolo Háplico fíbrico/sáprico térrico foram identificados quatro sub-horizontes hísticos assentes em horizonte Cg de cor cinza-escura (10YR 4/1), com textura média e espessura maior do que 30 cm dentro de 1 m de profundidade, o que o classifica como térrico (Quadro 1). Esse ambiente redutor é causado pela permanência de saturação hídrica devido ao leve abaciamento local, combinado à presença de horizontes minerais com baixa permeabilidade saturada na parte inferior do perfil, retardando a saída de água do sistema (Quadro 3). Em ambos os solos há predominância de material orgânico fíbrico herbáceo em superfície. Todos os horizontes de ambos os solos são extremamente ácidos (Quadros 1 e 2), coincidindo com as características de solos de demais trabalhos realizados em campos altomontanos (Wisniewski et al., 2005;Vashchenko et al., 2007) e de solos associados aos complexos rupestres de altitude (Benites et al., 2003, 2005, 2007; Simas et al., 2005).
Os horizontes hísticos dos Organossolos apresentaram Al, saturações por Al e T (potenciais) maiores nas partes mais profundas dos perfis, principalmente no perfil 2 (Rs = 1; p < 0,01; Quadros 1 e 2), devido à maior presença da fração argila no sistema (Quadro 3). As bases dos sub-horizontes hísticos inferiores provavelmente foram lixiviadas, conforme sua diminuição de teores, na ordem de duas a cinco vezes com a profundidade (Quadro 2). Os níveis de P, considerados de baixos a médios de acordo com Serrat et al. (2006), e as saturações por bases já bastante baixas em superfície diminuem significativamente com a profundidade (Rs = - 1; p < 0,01; Quadros 1 e 2).
Solos da Floresta Ombrófila Densa Altomontana na Serra da Igreja
Os solos representados pelo perfil 3 foram classificados como Gleissolos Háplicos alíticos típicos (Quadro 1), por apresentarem horizontes Cg com textura média, subjacentes aos horizontes O hísticos (N2/) com menos de 40 cm de espessura e com altos teores de C orgânico total (408,2 g kg-1). Segundo Embrapa (2006), são classificados como rasos a pouco profundos, fato perfeitamente coerente com a imposição geomórfica. Foram identificados dois sub-horizontes minerais gleizados: Cg1 com cor cinza-escura 10YR 4/1 e Cg2 bruno-amarelada 10YR 5/3, indicando a ação de fluxos hídricos subsuperficiais. O relevo forte ondulado, em rampa côncava-convergente, enseja a formação desses horizontes minerais por meio da ação combinada de processos morfogenéticos (coluviamento) e pedogenéticos. Assim como os Organossolos situados em campos, ambos os horizontes, orgânicos e minerais, são extremamente ácidos e com baixa saturação por bases, como solos descritos em outras florestas altomontanas (Roderjan, 1994; Ghani, 1996; Rocha, 1999; Portes, 2000; Wisniewski et al., 2005; Vashchenko et al., 2007).
O horizonte orgânico desses solos apresenta maior acidez potencial devido aos íons H+, provenientes dos grupos carboxílicos e fenólicos da matéria orgânica (Zech et al., 1997), e aos maiores teores de bases trocáveis, de P e de T. Os horizontes gleizados, altamente dessaturados por bases, apresentam os maiores valores de Al e de saturação por Al (caráter alítico), sugerindo que a floresta cicle, sobretudo, no volume compreendido pelo horizonte hístico e serapilheira. Os teores de P são ligeiramente superiores no horizonte O hístico na floresta (Gleissolo) do que nos hísticos superficiais dos campos (Organossolos), embora não tenham sido encontradas diferenças estatisticamente significativas. Gleissolos em florestas altomontanas no Paraná também foram encontrados por Rocha (1999) e por Wisniewski et al. (2005), porém não foram os solos predominantes nesses levantamentos.
Características gerais e diferenças encontradas entre solos das florestas e dos campos altomontanos
Conforme verificado nos itens anteriores, os solos que constituem os ecossistemas altomontanos na Serra da Igreja são distróficos e apresentam alta saturação por Al trocável. A baixa fertilidade encontrada em solos de ambientes altomontanos é relacionada à perda de nutrientes por lixiviação, pela elevada drenagem e pelo baixo conteúdo de nutrientes no material parental (Benites et al., 2007). Segundo esses autores, a acidez favorece a dissolução de caulinita e de aluminossilicatos, e o Al satura o complexo de troca.
A alta capacidade de troca catiônica e acidez potenciais nos horizontes superficiais evidencia que a maior parte das cargas negativas é proveniente de compostos orgânicos (Simas et al., 2005). A ciclagem de nutrientes é essencial, pois, conforme pode ser visualizado no quadro 1 (segundo considerações de Serrat et al., 2006; teores de Ca - baixos a muito baixos, e de Mg e K, muito altos), as maiores quantidades de nutrientes estão nos horizontes orgânicos superficiais (Benites et al., 2007), e um mecanismo para reduzir perdas é a formação de um sistema com alta densidade de raízes (Zech et al., 1997; Simas et al., 2005), o qual aumenta a eficiência na absorção de hidrossolúveis. Apesar das altas saturações por Al, grande parte desse elemento está complexada com a matéria orgânica, controlando os níveis na solução desses solos extremamente ácidos e evitando sua toxicidade para as plantas (Stevenson, 1982; Pavan, 1983; Zech et al., 1997; Simas et al., 2005; Don et al., 2007).
A gênese processada em hidromorfismos sazonais desses solos está condicionada ao fator altitude, em que se verificam temperaturas baixas e neblina frequente, causando acúmulo de matéria orgânica pela baixa taxa de mineralização (Rocha, 1999), além de elevado índice pluvial. O acúmulo de materiais orgânicos sobrepuja a taxa de decomposição, pois, apesar de haver transformação biológica, ao mesmo tempo, há preservação de níveis consideráveis de C orgânico na forma humificada (Ghani, 1996).
De acordo com os levantamentos já realizados nos campos altomontanos no Paraná, parecem predominar Neossolos Litólicos hísticos e Organossolos, ambos solos com horizontes hísticos, sem a presença de horizontes minerais ou com sua ocorrência somente após os 40 cm de profundidade. Conforme já comentado, a espessura do horizonte hístico maior do que 20 cm sem horizonte mineral dentro dos 40 cm (Organossolos) parece dificultar o estabelecimento físico de indivíduos arbóreos com porte maior do que 1 m. A gênese ascensional dos horizontes hísticos (em alguns casos, alcançando mais do que 50 cm de profundidade) e sua baixa densidade global, aliada à alta densidade de raízes de espécies típicas de campo e à exposição aos ventos fortes nas cumeeiras aguçadas do campo estudado, parecem não permitir o crescimento e o estabelecimento de indivíduos arbóreos de maior porte para propiciar o avanço da floresta altomontana. Se mudanças climáticas futuras resultarem em aumento da temperatura média anual e diminuição na precipitação pluvial na região, esses campos altomontanos correm o risco de desaparecer ou de perder grande parte de sua área para as florestas devido à oxidação, contração e consequente erosão dos horizontes hísticos.
Mais áreas altomontanas devem ser investigadas para se saber se a floresta já alcançou seu limite máximo de expansão sobre os campos altomontanos no Holoceno tardio. Em solos associados aos complexos rupestres de altitude na região Sudeste do Brasil, Benites et al. (2007) constataram que, com o desenvolvimento da paisagem, algumas turfeiras (Organossolos) secam suficientemente pelo estabelecimento de um sistema de drenagem, propiciando o crescimento de vegetação florestal, mas mantendo os grandes estoques de C. A fragilidade dos ecossistemas altomontanos é agravada pelo impacto de trilhas e acampamentos. Foi observado em serras vizinhas com maior visitação que, em períodos de estiagem, embora muito pouco comuns, os ambientes altomontanos apresentam alta suscetibilidade a incêndios antrópicos. Isso se deve à grande quantidade de material com alto grau de combustibilidade, implicando enorme risco de perda desses ambientes de baixa resiliência e de suas funcionalidades ambientais.
Os horizontes hísticos dos Organossolos abaixo dos 20 cm já apresentam mudanças químicas consideráveis em relação aos horizontes minerais encontrados em solos da floresta altomontana (Quadros 1 e 2). No entanto, os Organossolos Fólicos Sápricos líticos foram caracterizados por Rocha (1999) em florestas altomontanas, porém relativamente pouco desenvolvidas (com altas densidades e baixas áreas basais), apresentando horizontes hísticos entre 20 e 35 cm de espessura. Como nos trabalhos de Roderjan (1994) e Rocha (1999), também foram encontradas estruturas grumosas de matriz sáprica entre raízes finas. Segundo Rocha (1999), essas estruturas podem ser observadas com maior facilidade em relevos mais declivosos, onde as árvores, à medida que envelhecem, vão se curvando e, com isso, suspendendo seu sistema radicial, formando espaços vazios entre o solo e a rocha abaixo. Esses espaços e as maiores declividades proporcionam melhor drenagem e consequente aeração, podendo propiciar o avanço da floresta sobre os campos. Roderjan (1994) e Rocha (1999) também observaram Cambissolos, e Rocha (1999) também detectou Argissolos e Gleissolos em vales com maior coluvionamento, apresentando florestas altomontanas relativamente com baixas densidades e altas áreas basais. O caráter epigleico descrito por esses trabalhos limita o desenvolvimento da floresta, resultando em baixas densidades e áreas basais. Roderjan (1994), Portes (2000) e Vashchenko et al. (2007) encontraram Neossolos Litólicos (distróficos típicos e húmicos típicos) em situações de rampas convexas divergentes em encostas com florestas altomontanas nos picos Camacuã, Camapuã e Tucum, a 40 km ao norte da área estudada. Os horizontes minerais (A húmico ou C) desses solos parecem promover maior agregação mecânica, resultando numa maior capacidade de sustentação das árvores do que os Neossolos Litólicos hísticos (Portes, 2000), mais comuns em campos altomontanos (Vashchenko et al., 2007).
No entanto, conforme já comentado, florestas sobre horizontes hísticos pouco espessos diretamente sobre rocha - Neossolos Litólicos hísticos ( < 20 cm) com inclusão de Organossolos Fólicos com poucos centímetros a mais do que o solo anterior também podem dar suporte a florestas altomontanas, porém de pequeno porte, com altas densidades e baixas áreas basais (Roderjan, 1994; Rocha, 1999).
Estoques de carbono nos solos
Os estoques de C nos solos do campo altomontano, incluindo os dados dos perfis (Quadro 3) e das tradagens, variaram de 274,3 a 410,6 Mg ha-1 C (média mais erro-padrão: 314,7 ± 17,2 Mg ha-1 C). Na floresta altomontana os estoques variaram de 190,1 a 233,4 Mg ha-1 C (217,2 ± 10,3 Mg ha-1 C), sendo estatisticamente maiores no campo do que na floresta (teste U; p < 0,05) (Quadro 3). No levantamento prévio em toda a cumeeira estudada, obteve-se uma média de profundidade dos horizontes hísticos muito próxima (54 ± 3,7 cm) à utilizada nas estimativas supracitadas.
Segundo levantamento realizado no Paraná pelo Pró-Atlântica (2002), os campos altomontanos e a Floresta Ombrófila Densa Altomontana ocupam atualmente 1.627 e 5.760 ha, respectivamente. Apesar de os solos dos campos altomontanos estocarem mais C por unidade de área do que os das florestas altomontanas, a maior área destas no Paraná resulta numa maior quantidade estocada (1,25 x 10 6 t de C; Mt = megatoneladas) do que a dos campos (0,54 Mt de C). No entanto, para uma estimativa mais precisa desses estoques, são necessários mais levantamentos, uma vez que são muito comuns diferenças significativas nos estoques de C encontradas nos solos em diferentes exposições, posições topográficas e materiais parentais numa mesma montanha (Yimer et al., 2006; Garcia-Pausas et al., 2007).
Pela estimativa realizada por fotos horizontais e aéreas de 1980 do Instituto de Terras e Cartografia (ITC-PR), obteve-se uma área de aproximadamente 8 ha de campos altomontanos na Serra da Igreja, distribuídos em pelo menos 18 pequenos fragmentos, encravados em ambientes sob dominância da floresta altomontana. Com base no levantamento do Pró-Atlântica (2002), foi estimada uma área de pelo menos 400 ha para a floresta altomontana. Isso corresponde a aproximadamente 2.518 e 86.880 Mg de C estocadas nos solos dos campos e florestas altomontanas da Serra da Igreja, respectivamente. Contudo, ressalta-se que no presente trabalho foram estimados apenas os potenciais de estoques de C, sendo necessárias estimativas em escalas de maior detalhe, considerando a dificuldade de mapear inúmeros outros pequenos fragmentos de campos e de separar as florestas montanas das altomontanas, da sua transição, dos afloramentos de rocha e da vegetação rupestre, bem como considerar demais tipos de solos.
Apesar de apresentarem teores de C pelo menos 10 vezes menores do que os dos horizontes hísticos, os horizontes minerais gleizados dos Gleissolos da floresta altomontana possuem quase a metade do estoque de C do solo (48 %) . Essa compensação deve-se às densidades do solo 10 vezes superiores às dos horizontes hísticos (Quadro 3). Nos campos, somente os Organossolos Háplicos sápricos térricos apresentam contribuição significativa dos horizontes gleizados para o estoque de C total do solo (em média, 28 %). Os horizontes gleizados dos demais Organossolos mostraram pequena contribuição, entre 0 e 5 % (Quadro 3).
Ambos os valores, principalmente os dos campos, podem ser considerados altos, pois são da mesma magnitude dos estoques de C encontrados por Yimer et al. (2006), em florestas na Etiópia (326 a 460 Mg ha-1 C), em altitudes muito mais elevadas (de 2390 a 3580 m s.n.m.) e em temperaturas médias anuais um pouco menores do que as do presente estudo (entre 9,2 e 12,1 °C), porém em baixas latitudes e baixa precipitação anual (985 mm). Os valores calculados para os campos altomontanos na Serra da Igreja são mais do que o dobro dos encontrados por Garcia-Pausas et al. (2007) em campos alpinos e subalpinos da Espanha (153 Mg ha-1 C), em altitudes e latitudes superiores às do presente estudo, precipitação similar e temperaturas médias anuais entre -0,7 e 5 °C. Nesse caso, as baixas temperaturas devem resultar em menor produtividade primária e, consequentemente, em menor acúmulo de matéria orgânica no solo do que as áreas do presente estudo. Reservas de C muito menores (entre 48 e 86 Mg ha-1 C) foram encontradas em campos manejados na Alemanha, com baixas temperaturas (média anual de 8 °C), porém com baixa precipitação média anual (em torno de 600 mm) (Don et al., 2007).
Os estoques de C no solo para os principais sítios da Floresta Amazônica (Batjes & Dijksboorn, 1999; Novaes Filho et al., 2007) também são várias vezes menores (entre 40 e 98 Mg ha-1 C) que os do presente estudo, provavelmente devido às elevadas médias anuais de temperaturas e precipitações, as quais favorecem a rápida decomposição com menores taxas de humificação. Entretanto, sobre Organossolos, Batjes & Dijksboorn (1999) estimaram valores muito maiores (724 Mg ha-1 C) que os estimados na Serra da Igreja, certamente devido à baixa decomposição, por se tratar de solos hidromórficos e pela consideração de valores médios de densidade superiores.
Os solos altomontanos ora estudados estocam mais C do que solos em diversas formações nativas (de 37 a 118 Mg ha-1 C), em altitudes mais baixas na mesma latitude, muitas delas com maior produtividade primária, conforme dados de Britez et al. (2006) e Balbinot (2009).
Comparações com os estoques de carbono na biomassa acima do solo em florestas
Os valores médios de C estocado no solo dos campos altomontanos e das florestas altomontanas na Serra da Igreja (314,7 e 217,2 Mg ha-1 C, respectivamente) são superiores às maiores estimativas dos estoques para a fitomassa da própria Floresta Ombrófila Densa Submontana (em torno de 200 Mg ha- 1 C, média próxima a 120 Mg ha-1 C), formação que apresenta uma das maiores biomassas acima do solo (Tiepolo et al., 2002; Vieira et al., 2008). Britez et al. (2006), com base em dados de diversos inventários fitossociológicos, para diversas regiões fitoecológicas do Bioma Mata Atlântica no Paraná, estimaram estoques de C acima do solo de 26 Mg ha-1 C (formação secundária em estádio inicial) a 153 Mg ha-1 C (floresta bem desenvolvida), ambos em região da Floresta Ombrófila Densa. Novamente, essas informações relevam a grande importância dos estoques de C nos solos dos ecossistemas altomontanos, considerando que o solo do campo tem a capacidade média de estocar mais do que o dobro do estoque presente na biomassa viva acima do solo da Floresta Ombrófila Densa Submontana, a qual, possivelmente, apresenta a maior produtividade primária entre as formações mencionadas.
Estoque potencial de água
Os horizontes superficiais hísticos apresentam alta permeabilidade saturada (até 537 cm h-1; Quadro 3). No entanto, à medida que a água permeia os horizontes subjacentes, principalmente o hístico mais profundo (sáprico) no caso dos Organosssolos e dos horizontes gleizados nos Gleissolos, há brusca diminuição da permeabilidade (2 a 6 cm/h), favorecendo maior "retenção hídrica" no volume. Nos perfis abertos em Organossolos foi possível, por diferenças das cores de N2/ para 10YR 2/1 a 10YR 3/2, perceber fluxos hídricos laterais de subsuperfície nos horizontes hísticos subsuperficiais (hêmicos e sápricos).
A permeabilidade muito alta dos horizontes superficiais deve-se às suas características fíbricas, as quais possuem as menores densidades (0,132 a 0,08 kg dm-3), maior macroporosidade e aeração, bem como, provavelmente, ótima conectividade dos poros. A despeito da grande capacidade de retenção hídrica dos horizontes hísticos, os teores de água disponível são baixos (Quadro 3).
Os horizontes gleis dos Gleissolos apresentaram menores porosidades totais, além de menor percentual de macroporos, o que incorre em menor capacidade de aeração e diminuição da permeabilidade saturada - fato relacionado à mudança de textura e estrutura dos horizontes (Quadro 3).
Segundo Machado & Favaretto (2006), o aumento da disponibilidade de água em solos com maiores teores de matéria orgânica deve-se à influência desta na estrutura do solo e não da água retida no material orgânico. A matéria orgânica apresenta elevada umidade na capacidade de campo, porém a umidade no ponto de murcha também aumenta proporcionalmente.
A retenção hídrica máxima (umidade de saturação em volume) pode ser considerada a porosidade total do solo. No entanto, a saturação por água de todos os espaços porosos pode não ocorrer plenamente, pois alguns microporos podem permanecer com ar mesmo após o processo de saturação (Machado & Favaretto, 2006).
Uma simples estimativa da capacidade de armazenamento de água pode ser calculada por meio da umidade volumétrica, obtida pela diferença entre a porosidade total e a porosidade de aeração (Prevedello, 1996). Considerando que pelo menos 95 % dos horizontes hísticos dos Organossolos são poros (Quadro 3) e que a profundidade média é de 58 cm e sua umidade volumétrica é de 71 %, significa que cada metro quadrado de solo pode reter 411,8 L de água (4.118 m3 ha-1). Isso significa que a fase sólida dos horizontes hísticos dos Organossolos dos campos altomontanos retém aproximadamente 12 vezes (6 a 16) seu volume em água (ou de 4 a 10 vezes seu peso em água), e isso deve-se, sobretudo, à sua estrutura; mais especificamente, as estimativas para os Organossolos analisados (perfis 1 e 2) foram de 2.912 e 5.567 m3 ha-1, respectivamente.
Para o Gleissolo (perfil 3), a capacidade de armazenamento de água foi estimada em 2.969 m3 ha-1. Por se tratar de horizontes minerais, a fase sólida dos horizontes gleizados (entre 26 e 49 %) é muito superior à dos horizontes hísticos (entre 4 e 10 %). Portanto, esses horizontes retêm apenas entre 0,9 e 1,5 vezes seu volume em água (ou de 0,36 a 0,63 seu peso em água). Apesar de os valores de umidade volumétrica (média de 48 %) serem menores do que os encontrados para os horizontes hísticos, devido à sua menor porosidade total e de aeração, os horizontes gleizados contribuem com uma importante parcela no armazenamento de água nos solos altomontanos, sendo 29 % no perfil 2 (Organossolo do campo) e 49 % no perfil 3 (Gleissolo na floresta).
Esses resultados indicam alta capacidade dos solos altomontanos de retenção de água e regulação dos fluxos hídricos nas cabeceiras de bacias hidrográficas e estão próximos dos valores apresentados por Soil Survey Staff (1992), em que o material orgânico fíbrico é o que possui a maior capacidade de armazenar água: de 8,5 a 30 vezes seu peso seco, ao passo que o hêmico pode reter de 4,5 a 8,5 vezes, e o sáprico, até 4,5 vezes.
CONCLUSÕES
1. Os solos altomontanos da Serra da Igreja são distróficos, extremamente ácidos, e apresentam alta saturação por Al trocável e altos teores de C orgânico total.
2. As florestas estão fortemente estabelecidas por controle estrutural de vales e colos relacionados a maior ação de processos morfogenéticos, os quais resultam em solos com horizontes minerais e maior acúmulo de argila, como os Gleissolos Háplicos alíticos típicos encontrados no presente estudo. A maior densidade desses horizontes proporciona maior consistência para a condição de suporte e desenvolvimento de espécies arbóreas. Portanto, nos ambientes altomontanos, o espessamento dos horizontes minerais e o adelgaçamento dos orgânicos promovem o desenvolvimento da floresta.
3. Os campos altomontanos ocupam cumeeiras aguçadas sobre Organossolos Fólicos fíbricos/sápricos (líticos e típicos) e Organossolos Háplicos fíbricos/ sápricos (típicos e térricos) dispostos em rampas convexas divergentes, nas quais os processos pedogenéticos promoveram maior espessura de horizontes hísticos. As baixas densidades e menores consistências desses horizontes, associadas às condições de hidromorfia plena sazonal e à presença de fortes ventos, proporcionam forte restrição à ocupação desses ambientes pela floresta. Nos locais estudados, somente após abrupta ruptura de declive, rampas côncavas-convergentes propiciam o desenvolvimento da floresta sobre Gleissolos com horizontes hísticos.
4. Os estoques de C por unidade de área nos solos dos campos são superiores (314,7 Mg ha-1 C) aos das florestas altomontanas (217,2 Mg ha-1 C), sendo ambos considerados altos, comparados aos dados de outros ecossistemas disponíveis na literatura, bem como superiores aos estoques estimados para a biomassa aérea de florestas submontanas, as quais apresentam alta produtividade primária.
5. Os altos potenciais de armazenamento hídrico dos solos altomontanos estudados (entre 291,2 e 556,7 L m-2 ) estão fortemente relacionados às altas porosidades totais dos horizontes hísticos em conjunto com horizontes gleizados. Essas características, bem como a capacidade de filtro desses solos, pelo alto conteúdo de materiais orgânicos, também devem ser reconhecidas como importantes funcionalidades desses ambientes, as quais implicam na necessidade de sua preservação.
AGRADECIMENTOS
À Fundação O Boticário de Proteção à Natureza, pelo patrocínio, e à Sociedade Chauá, pelo apoio ao projeto, em especial a Christopher Blum. Somos gratos ao auxílio de campo de Marcos Rachwal, Marcos de Souza, Letícia Sousa, Yury Vashchenko, Anette Bonnet e Marcia de Lima. Agradecemos também as contribuições de Renato Dedecek, Celso Prevedello, Renato Goldenberg, Alexandre Uhlmann, Franklin Galvão, Luiz Carlos Pessenda, Kelly Gütseit, Alan Mocochinski, Celina Wisniewski e dos revisores anônimos da revista.
LITERATURA CITADA
FUNCIONALIDADES AMBIENTAIS DE SOLOS ALTOMONTANOS NA SERRA DA IGREJA, PARANÁ 1125
- ARTEAGA, G.A.; CALDERÓN, N.E.G.; KRASILNIKOV, P.V.; SEDOV, S.N.; TARGULIAN, V.O. & ROSAS, N.V. Soil altitudinal sequence on base-poor parent material in a montane cloud forest in Sierra Juárez, Southern Mexico. Geoderma, 144:593-612, 2008.
- BALBINOT, R. Carbono, nitrogênio e razões isotópicas δ13C e δ15N no solo e vegetação de estágios sucessionais de Floresta Ombrófila Densa Submontana. Curitiba, Universidade Federal do Paraná, 2009. 103p. (Tese de Doutorado)
- BATJES, N.H. Total carbon and nitrogen in the soils of the world. Europ. J. Soil Sci., 47:151-163, 1996.
- BATJES, N.H. & DIJKSHOORN, J.A. Carbon and nitrogen stocks in the soils of the Amazon region. Geoderma, 89:273-286, 1999.
- BECKER, A.; KÖRNER, C.; BRUN, J.; GUISAN, A. & TAPPEINER, U. Ecological and land use studies along elevational gradients. Mt. Res. Dev., 27:58-65, 2007.
- BENITES, V.M.; CAIAFA, A.N.; MENDONÇA, E.S.; SCHAEFFER, C.E.G.R. & KER, J.C. Solos e vegetação nos Complexos Rupestres de Altitude da Mantiqueira e do Espinhaço. R. Flor. Amb., 10:76-85, 2003.
- BENITES, V.M.; MENDONÇA, E.S.; SCHAEFER, C.E.G.R.; NOVONTNY, E.H.; REIS, E.L. & KER, J.C. Properties of black soil humic acids from high altitude rocky complexes in Brazil. Geoderma, 127:104-113, 2005.
- BENITES, V.M.; SCHAEFER, C.E.G.R.; SIMAS, F.N.B. & SANTOS, H.G. Soils associated with rock outcrops in the Brazilian mountain ranges Mantiqueira and Espinhaço. R. Bras. Bot., 30:569-577, 2007.
- BERNOUX, M.; CARVALHO, M.C.S.; VOLKOFF, B. & CERRI, C.C. Brazil's soil carbon stocks. Soil Sci. Soc. Am. J., 66:888-896, 2002.
- BRITEZ, R.M.; BORGO, M.; TIEPOLO, G.; FERRETI, A.; CALMON, M. & HIGA, R. Estoque e incremento de carbono em florestas e povoamentos de espécies arbóreas com ênfase na Floresta Atlântica do sul do Brasil. Colombo, Embrapa Florestas, 2006.
- BRUIJNZEEL, L.A. Hydrology of tropical montane cloud forests: a reassessment. In: INTERNATIONAL COLLOQUIUM, 2., Paris, 2000. Proceedings... Paris, UNESCO, 2000. p.353-383.
- BRUIJNZEEL, L.A. & PROCTOR, J. Hydrology and biogeochemistry of tropical montane cloud forests: What do we really know? In: HAMILTON, L.S.; JUVIK J.O. & SCATENA, F.N., ed. Tropical montane cloud forests. New York, Springer Verlag, 1995. p.38-78.
- DON, A.; SHUMACHER, J.; SHERER-LORENZEN, M.; SCHOLTEN, T. & SCHULZE, E. Spatial and vertical variation of soil carbon at two grassland sites - Implications for measuring soil carbon stocks. Geoderma, 141:272-282, 2007.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Centro Nacional de Pesquisa de Solos. Sistema brasileiro de classificação de solos. 2.ed. Rio de Janeiro, 2006. 306p.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Centro Nacional de Pesquisa de Solos. Manual de métodos de análises de solos. 2.ed. Rio de Janeiro, 1997. 212p.
- FALKENBERG, D.B. Matinhas nebulares e vegetação rupícola dos Aparados da Serra Geral (SC/RS), sul do Brasil. Campinas, Universidade de Campinas, 2003. 558p. (Tese de Doutorado)
- GARCIA-PAUSAS, J.; CASALS, P.; CAMARERO, L.L.; HUGUET, C.; SEBASTIÀ, M.A.T.; THOMPSON, R. & ROMANYÀ, J. Soil organic carbon storage in mountain grasslands of the Pyrenees: effects of climate and topography. Biogeochemistry, 82:279-289, 2007.
- GHANI, N.L.B. Caracterização morfológica, física, química, mineralógica, gênese e classificação de solos altimontanos derivados de riólito e migmatito da Serra do Mar - PR. Curitiba, Universidade Federal do Paraná, 1996. 174p. (Tese de Mestrado)
- HAMILTON, L.S.; JUVIK, J.O. & SCATENA, F.N. The Puerto Rico tropical cloud forest symposium: introduction and workshop synthesis In: HAMILTON, L.S., JUVIK, J.O. & SCATENA, F.N. Tropical montane cloud forests. New York, Springer Verlag, 1995. p.1-23.
- JOBBÁGY, E.G. & JACKSON, R.B. The vertical distribuition of soil organic carbon and its relation to climate and vegetation. Ecol. Applic., 10:423-436, 2000.
- MAACK, R. Geografia física do Estado do Paraná. Curitiba, Imprensa Oficial, 2002. 440p.
- MARTÍNEZ, M.L.; PÉREZ-MAQUEO, O.; VÁZQUEZ, G.; CASTILLO-CAMPOS, G.; GARCÍA-FRANCO, J.; MEHLTRETER, K.; EQUIHUA, M. & LANDGRAVE, R. Effects of land use change on biodiversity and ecosystem services in tropical montane cloud forests of Mexico. For. Ecol. Manag., 258:1856-1863, 2009.
- MELO, M.S.; GIANNINI, P.C.F. & PESSENDA, L.C.R. Gênese e evolução da Lagoa Dourada, Ponta Grossa, PR. R. Inst. Geol., 21:17-31, 2000.
- MINEROPAR MINERAIS DO PARANÁ S.A. Atlasgeo.pdf. Atlas geológico do estado do Paraná. Curitiba, 2001.
- MOCOCHINSKI, A.Y. & SCHEER, M.B. Campos de altitude na Serra do Mar paranaense: Aspectos florísticos. R. Flor., 38:625-640, 2008.
- MACHADO, M.A.M. & FAVARETTO, R. Atributos físicos do solo relacionados ao manejo e conservação dos solos. In: LIMA, M.R. et al., eds. Recomendações de manejo do solo: Aspectos tecnológicos e metodológicos. Curitiba, Universidade Federal do Paraná, 2006. p.233-254.
- NOVAES FILHO, J.P.N.; SELVA, E.C.; COUTO, E.G.; LEHMANN, J.; JOHNSON, M.S. & RIHA, S.J. Distribuição espacial de carbono em solo sob floresta primária na Amazônia Meridional. Soc. Invest. Flor., 31:83-92, 2007.
- PORTES, M.C.G.O. Deposição de serapilheira e decomposição foliar em floresta ombrófila densa altomontana, Morro Anhangava, Serra da Baitaca, Quatro Barras - PR. Curitiba, Universidade Federal do Paraná, 2000. 90p. (Tese de Mestrado)
- PAVAN, M.A. Alumínio em solos ácidos do Paraná: relação entre o alumínio não-trocável, trocável e solúvel, com o pH, CTC, porcentagem de saturação de Al e matéria orgânica. R. Bras. Ci. Solo, 7:39-46, 1983.
- PREVEDELLO, C.L. Física do solo: Com prolemas resolvidos. Curitiba, C.L. Prevedello, 1996. 446p.
- PROGRAMA DE PROTEÇÃO DA FLORESTA ATLÂNTICA - PRÓ-ATLÂNTICA. Projeto Carta Geológica. Cartas: MI-2844-2 Ariri, MI-2843-1 Represa do Capivari, MI-2843-3 Morretes, MI-2858-1 Mundo Novo e MI-2858-3 Pedra Branca do Araraquara. Escala 1:50000. Curitiba, SEMA, 2002.
- ROCHA, M.R.L. Caracterização fitossociológica e pedológica de uma Floresta Ombrófila Densa no Parque Estadual do Pico do Marumbi - Morretes, PR. Curitiba, Universidade Federal do Paraná, 1999. 80p. (Tese de Mestrado)
- RODERJAN, C.V. A Floresta Ombrófila Densa Altomontana no Morro Anhangava, Quatro Barras, PR: Aspectos climáticos, pedológicos e fitossociológicos. Curitiba, Universidade Federal do Paraná, 1994. 119p. (Tese de Doutorado)
- RODERJAN, C.V. & GRODSKI, L. Acompanhamento meteorológico em um ambiente de Floresta Ombrófila Densa Altomontana no morro Anhangava, Mun. de Quatro Barras - PR, no ano de 1993. Cader. Biodivers., 2:27-34, 1999.
- SANTOS, R.D.; LEMOS, R.C.; SANTOS, H.G.; KER, J.C. & ANJOS, L.H.C. Manual de descrição e coleta de solo no campo. 5.ed. Viçosa, MG, Sociedade Brasileira de Ciência do Solo, 2005. 92p.
- SCHEER, M.B. & MOCOCHINSKI, A.Y. Florística vascular da Floresta Ombrófila Densa Altomontana de quatro serras no Paraná. 2009. Biota Neotrop., 9. Disponível em: <http://www.biotaneotropica.org.br/v9n2/en/abstract?article+bn00609022009>
- SCHEER, M.B. Ambientes altomontanos no Paraná: Florística vascular, estrutura arbórea, relações pedológicas e datações por 14C. Curitiba, Universidade Federal do Paraná, 2010. 153p. (Tese de Doutorado)
- SERRAT, B.M.; KRIEGER, K.I. & MOTTA, A.C.V. Considerações sobre a interpretação de análise de solos (com exemplos). In: LIMA, M.R. et al., eds. Recomendações de manejo do solo: Aspectos tecnológicos e metodológicos. Curitiba, Universidade Federal do Paraná, 2006. p.125-143.
- SILVA, A.C.; HORÁK, I.; CORTIZAS, A.M.; VIDAL-TORRADO, P.; RACEDO, J.R.; GRAZZIOTTI, P.H.; SILVA, E.B. & FERREIRA, C.A. Turfeiras da Serra do Espinhaço Meridional-MG: I - caracterização e classificação. R. Bras. Ci. Solo, 33:1385-1398, 2009.
- SIMAS, F.N.B.; SCHAEFER, C.E.G.R.; FERNANDES, E.I.; CHAGAS, A.C. & BRANDÃO, P.C. Chemistry, mineralogy and micropedology of highland soils on crystalline rocks of Serra da Mantiqueira, southeastern Brazil. Geoderma, 125:187-201, 2005.
- SOIL SURVEY STAFF. Keys to soil taxonomy. 5.ed. Virginia, Pocahontas Press, 1992. 541p.
- STEVENSON, F.J. Humus chemistry: Genesis, composition, reactions. New York, Wiley-Interscience, 1982. 443p.
- TIEPOLO, G.; CALMON, M. & FERETTI, A.R. Measuring and monitoring carbon stocks at the Guaraqueçaba Climate Action Project, Paraná, Brazil. In: INTERNATIONAL SYMPOSIUM ON FOREST CARBON SEQUESTRATION AND MONITORING. Extension Serie Taiwan Forestry Research Institute, 153:98-115, 2002.
- VASHCHENKO, Y.; PIOVESAN, R.P.; LIMA, M.R & FAVARETTO, N. Solos e vegetação dos Picos Camacuã, Camapuã e Tucum - Campina Grande Do Sul-PR. Sci. Agr., 8:411-419, 2007.
- VIEIRA, S. M. et al. Estimation of biomass and carbon stocks: the case of the Atlantic Forest. 2008. Biota Neotrop., 8(2). Available from: <http://www.biotaneotropica.org.br/v8n2/en/abstract?point-of-view+bn00108022008>
- VOLKOFF, B.; CERRI, C.C. & MELFI, A.J. Húmus e mineralogia dos horizontes superficiais de três solos de campos de altitude dos estados de Minas Gerais, Paraná e Santa Catarina. R. Bras. Ci. Solo, 8:277-283, 1984.
- WISNIEWSKI, C.; TEMPESTA, P.B. & RODRIGUES, V.M.R. Solos e vegetação em uma topossequência do morro Mãe Catira, alto da Serra da Graciosa, Morretes PR. In: CONGRESSO BRASILEIRO DE CIÊNCIA DO SOLO, 30., Recife, 2005. Anais... Viçosa, MG, Sociedade Brasileira de Ciência do Solo, 2005.
- YIMER, F.; LEDIN, S. & ABDLKADIR, A. Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetation in the Bale Mountains, Ethiopia. Geoderma, 135:335-344, 2006.
- ZECH, W.; SENESI, N.; GUGGENBERGER, G.; KAISER, K.; LEHMANN, K.; MILTNER, A.; MIANO, T.M. & SCHROTH, G. Factors controlling humification and mineralizing of soil organic matter in the tropics. Geoderma, 79:117-161, 1997.
Datas de Publicação
-
Publicação nesta coleção
21 Out 2011 -
Data do Fascículo
Ago 2011
Histórico
-
Recebido
12 Maio 2010 -
Aceito
25 Maio 2011