Abstracts
Soil can be either source or sink of methane (CH4), depending on the balance between methanogenesis and methanotrophy, which are determined by pedological, climatic and management factors. The objective of this study was to assess the impact of drainage of a highland Haplic Histosol on CH4 fluxes. Field research was carried out in Ponta Grossa (Paraná, Brazil) based on the measurement of CH4 fluxes by the static chamber method in natural and drained Histosol, over one year (17 sampling events). The natural Histosol showed net CH4 eflux, with rates varying from 238 µg m-2 h-1 CH4, in cool/cold periods, to 2,850 µg m-2 h-1 CH4, in warm/hot periods, resulting a cumulative emission of 116 kg ha-1 yr-1 CH4. In the opposite, the drained Histosol showed net influx of CH4 (-39 to -146 µg m-2 h-1), which resulted in a net consumption of 9 kg ha-1 yr-1 CH4. The main driving factors of CH4 consumption in the drained soil were the lowering of the water-table (on average -57 cm, vs -7 cm in natural soil) and the lower water content in the 0-10 cm layer (average of 5.5 kg kg-1, vs 9.9 kg kg-1 in natural soil). Although waterlogged Histosols of highland areas are regarded as CH4 sources, they fulfill fundamental functions in the ecosystem, such as the accumulation of organic carbon (581 Mg ha-1 C to a depth of 1 m) and water (8.6 million L ha-1 = 860 mm to a depth of 1 m). For this reason, these soils must not be drained as an alternative to mitigate CH4 emission, but effectively preserved.
greenhouse gas; water sources; water-table; gravimetric moisture; air temperature; rainfall
O solo pode atuar como fonte ou sumidouro de metano (CH4), dependendo do balanço entre metanogênese e metanotrofia, definido por fatores pedológicos, climáticos e de manejo. O objetivo deste estudo foi avaliar as implicações da drenagem do Organossolo Háplico hêmico, típico em campo hidrófilo de altitude sobre os fluxos de CH4. A pesquisa de campo foi conduzida no município de Ponta Grossa, PR, e envolveu avaliações de fluxos de CH4 pelo método da câmara estática em Organossolo natural e Organossolo drenado, por um período de um ano (17 coletas). No Organossolo natural, ocorreu efluxo líquido de CH4, com taxas variando entre 238 µg m-2 h-1 de CH4, em épocas mais frias, e 2.850 µg m-2 h-1 de CH4, em épocas mais quentes, totalizando emissão acumulada de 116 kg ha-1 ano-1 de CH4. Na área drenada, ocorreu influxo líquido (-39 a -146 µg m-2 h-1 de CH4), que totalizou em consumo de 9 kg ha-1 ano-1 de CH4. O rebaixamento do nível freático (em média -57 cm, contra -7 cm no solo natural) e a menor umidade gravimétrica na camada de 0-10 cm (média de 5,5 kg kg-1, contra 9,9 kg kg-1 do solo natural) foram os principais fatores determinantes do consumo de CH4, na área drenada. Apesar de os Organossolos em campo hidrófilo atuarem como fonte de CH4, esses possuem importantes funções no ecossistema, como acumular carbono orgânico (581 Mg ha-1 de C até 1 m) e armazenar água (8,6 milhões de L = 860 mm até 1 m). Por essa razão, não devem ser drenados, como alternativa para redução da emissão de metano, mas efetivamente preservados.
gás de efeito estufa; fontes de água; nível freático; umidade gravimétrica; temperatura do ar e precipitação pluviométrica
DIVISÃO 2 - PROCESSOS E PROPRIEDADES DO SOLO
COMISSÃO 2.4 - QUÍMICA DO SOLO
Methane fluxes from waterlogged and drained Histosols of highland areas1 1 Part of the thesis submitted by the first author to obtain a doctorate in Forestry Science, sub-area Nature Conservation. Post-graduation course in Forestry, sector of Agricultural Sciences, Federal University of Paraná - UFPR. Received for publication on November 1 st, 2012 and approved on November 5, 2013.
Fluxos de metano em Organossolo natural e após drenagem
Marcos Fernando Glück RachwalI; Josiléia Acordi ZanattaI; Jeferson DieckowII; Genuir Luis DenegaIII; Gustavo Ribas CurcioI; Cimélio BayerIV
IResearcher, Embrapa Florestas. Estrada da Ribeira, km 111. CEP 83411-000 Colombo (PR), Brazil. E-mail: marcos.rachwal@embrapa.br, josileia.zanatta@embrapa.br, gustavo.curcio@embrapa.br
IIAdjunct Professor, Department of Soil Science and Agricultural Engeneering, UFPR. Rua dos Funcionários, 1540. CEP 80035-050 Curitiba (PR), Brazil. Researcher supported by CNPq. E-mail: jefersondieckow@ufpr.br
IIIDoctoral student in Soil Science, Federal University of Rio Grande do Sul - UFRGS. Av. Bento Gonçalves, 7712. CEP 91540-000 Porto Alegre (RS), Brazil. E-mail: genuirluis@gmail.com
IVAssociate Professor, Department of Soil Science, UFRGS. Researcher supported by CNPq. E-mail: cimelio.bayer@ufrgs.br
SUMMARY
Soil can be either source or sink of methane (CH4), depending on the balance between methanogenesis and methanotrophy, which are determined by pedological, climatic and management factors. The objective of this study was to assess the impact of drainage of a highland Haplic Histosol on CH4 fluxes. Field research was carried out in Ponta Grossa (Paraná, Brazil) based on the measurement of CH4 fluxes by the static chamber method in natural and drained Histosol, over one year (17 sampling events). The natural Histosol showed net CH4 eflux, with rates varying from 238 µg m-2 h-1 CH4, in cool/cold periods, to 2,850 µg m-2 h-1 CH4, in warm/hot periods, resulting a cumulative emission of 116 kg ha-1 yr-1 CH4. In the opposite, the drained Histosol showed net influx of CH4 (-39 to -146 µg m-2 h-1), which resulted in a net consumption of 9 kg ha-1 yr-1 CH4. The main driving factors of CH4 consumption in the drained soil were the lowering of the water-table (on average -57 cm, vs -7 cm in natural soil) and the lower water content in the 0-10 cm layer (average of 5.5 kg kg-1, vs 9.9 kg kg-1 in natural soil). Although waterlogged Histosols of highland areas are regarded as CH4 sources, they fulfill fundamental functions in the ecosystem, such as the accumulation of organic carbon (581 Mg ha-1 C to a depth of 1 m) and water (8.6 million L ha-1 = 860 mm to a depth of 1 m). For this reason, these soils must not be drained as an alternative to mitigate CH4 emission, but effectively preserved.
Index terms: greenhouse gas, water sources, water-table, gravimetric moisture, air temperature, rainfall.
RESUMO
O solo pode atuar como fonte ou sumidouro de metano (CH4), dependendo do balanço entre metanogênese e metanotrofia, definido por fatores pedológicos, climáticos e de manejo. O objetivo deste estudo foi avaliar as implicações da drenagem do Organossolo Háplico hêmico, típico em campo hidrófilo de altitude sobre os fluxos de CH4. A pesquisa de campo foi conduzida no município de Ponta Grossa, PR, e envolveu avaliações de fluxos de CH4 pelo método da câmara estática em Organossolo natural e Organossolo drenado, por um período de um ano (17 coletas). No Organossolo natural, ocorreu efluxo líquido de CH4, com taxas variando entre 238 µg m-2 h-1 de CH4, em épocas mais frias, e 2.850 µg m-2 h-1 de CH4, em épocas mais quentes, totalizando emissão acumulada de 116 kg ha-1 ano-1 de CH4. Na área drenada, ocorreu influxo líquido (-39 a -146 µg m-2 h-1 de CH4), que totalizou em consumo de 9 kg ha-1 ano-1 de CH4. O rebaixamento do nível freático (em média -57 cm, contra -7 cm no solo natural) e a menor umidade gravimétrica na camada de 0-10 cm (média de 5,5 kg kg-1, contra 9,9 kg kg-1 do solo natural) foram os principais fatores determinantes do consumo de CH4, na área drenada. Apesar de os Organossolos em campo hidrófilo atuarem como fonte de CH4, esses possuem importantes funções no ecossistema, como acumular carbono orgânico (581 Mg ha-1 de C até 1 m) e armazenar água (8,6 milhões de L = 860 mm até 1 m). Por essa razão, não devem ser drenados, como alternativa para redução da emissão de metano, mas efetivamente preservados.
Termos de indexação: gás de efeito estufa, fontes de água, nível freático, umidade gravimétrica, temperatura do ar e precipitação pluviométrica.
INTRODUCTION
Methane (CH4) emissions, which are affected by land use changes, account for 15 to 20 % of the global greenhouse gas emissions (Wuebbles et al., 2000; IPCC, 2007), and approximately one third of the global CH4 emissions come from natural ecosystems such as wetlands and lakes (Forster et al., 2007). Hydromorphic soils cover 3 % of the terrestrial surface and 1 % of the Brazilian territory (Valladares, 2003). These soils have a greater capacity to store carbon (C) than freely drained soils (Raich & Potter, 1995; Alongi et al., 2001; Whiting & Chanton, 2001), but on the other hand can be significant CH4 sources to the atmosphere (Hayes & Clapp, 2001; Whalen, 2005).
Soil CH4 fluxes result from the balance between methanogenesis and methanotrophy i.e., between gas production and consumption, respectively. Soils become a net source of CH4 when methaneproduction by methanogenic surpasses consumption by methanotrophic bacteria (Le Mer & Roger, 2001). These processes are regulated by oxygen (O2) supply and availability of labile carbon (C), being methanogenesis predominant under anaerobic conditions. The CH4 flux in waterlogged areas is controlled by soil properties and processes (Roulet & Moore, 1995; Whalen, 2005; Jauhiainen et al., 2008), by microbiological factors (Le Mer & Roger, 2001; Kögel-Knabner et al., 2010; Page & Dalal, 2011), climatic factors (von Arnold et al., 2005; Dalal & Allen, 2008; Jiang et al., 2009), vegetation (Furukawa et al., 2005; Koh et al., 2009) and land management (Elder & Lal, 2008; Huang et al., 2010). Precipitation and air temperature are important climatic factors that can affect soil CH4 emissions, although with great variability (Le Mer & Roger, 2001; Whalen, 2005; Dalal & Allen, 2008; Koh et al., 2009; Huang et al., 2010).
The water-table level and its fluctuation alter the intensity and duration of CH4 production and oxidation processes (Furukawa et al., 2005; von Arnold et al., 2005; Jauhiainen et al., 2008). Methane production reaches a maximum near 20 cm below the water-table, while oxidation occurs to a depth of 10 cm (Kettunen et al., 1999). However, CH4 emissions decrease logarithmically with the lowering of the water-table (Moore & Dalva, 1993; Furukawa et al., 2005), which can occur with drainage. On the other hand, the drainage of these soils may affect its capability of fixing C and storing water. Huang et al. (2010) estimated that the conversion of marshes into croplands reduced the soil organic C stock by approximately 50 %, with 70 % of this loss recorded to a depth of 20 cm. For gravimetric water content, Andrade (2010) reported a reduction from 6 to 2 kg kg-1 in the surface 10 cm, after the drainage of a Histosol.
In Paraná, headwater streams are common in landscapes above 800 m altitude. In these areas, Histosols, which occur in waterlogged grasslands, are important C and water reservoirs and, according to Kozera (2008) and Andrade (2010), play a significant role in biodiversity protection. Depending on the decomposition degree of the organic matter, these soils can store a water amount equivalent to 4.5 to 30 times its dry weight (Soil Survey Staff, 1992). Scheer et al. (2011) reported that waterlogged highland Histosols can store 314.7 Mg ha-1 C and 3.4 million L ha-1 water to a depth of 58 cm. With regard to the richness of plant species, Andrade (2010) and Kozera (2008) reported, respectively, the occurrence of 146 and 87 species on Histosols with highland grassland vegetation.
However, the ecological functions of most of these highland headwater streams have been badly damaged by agricultural activities. In Brazil, data on CH4 fluxes from Histosols in humid environments with small natural oscillation of the water table and low annual temperatures, and on the effects of drainage, are practically non-existent.
Based on the hypothesis that drainage reduces CH4 emissions from Histosols, the purpose of this study was to quantify and compare CH4 fluxes in a highland Haplic Histosol, either under natural waterlogged condition or after drainage, considering the influence of the water-table level, soil moisture, air temperature, and rainfall.
MATERIAL AND METHODS
Field experiment
The experiment was conducted on a waterlogged montane grassland on an experimental farm of Embrapa SPM-EPGA (25o 09' 59" S; 50o 04' 45" W; 857 m altitude), in the municipality of Ponta Grossa, on the second plateau of the State of Paraná, in the region of Campos Gerais, Brazil. The study location is one of the headwaters of the Cará-Cará river, a tributary of the Tibagi river. The climate is Cfb (Köppen), mesothermal humid, with no dry season, with a mean air temperature below 22 oC in the warmer months. The landscape is a distal reverse of cuesta, modeled on clay shales of the Ponta Grossa Formation, Paraná Group, Paleozoic. According to the Brazilian System of Soil Classification (Santos et al., 2006), the soil is classified as Haplic Histosol on a subtropical waterlogged montane grassland, with a flat relief and a sequential arrangement of five histic horizons (H1 to H5), thickness 20 cm, to a depth of 1 m. The main physical and chemical characteristics of natural soil were assessed on October 26, 2010, shortly after the beginning of the experiment (Table 1).
Two treatments were established. The first was Histosol under its natural waterlogged condition, without intervention, in a 15 × 15 m plot. The second was drained Histosol, in a 15 × 10 m plot, 45 m away from the first, in the same topographic elevation and with the same phytophysiognomy. Trapezoidal channels (upper width 1.5 m, lower width 1.2 m, depth 0.65 m) were dug in the perimeter of the area, with a backhoe-bulldozer combination, on September 8 and 9, 2010. The drainage area was relatively small to minimize the disturbance of the waterlogged grassland.
Air sample collection
For the evaluation of CH4 emissions, air samples were collected by the static-chamber method (Gomes et al., 2009; Elder & Lal, 2008). Immediately after draining the soil, six circular metal bases (internal diameter 38 cm) were inserted 15 cm deep into the soil, in each treatment plot. Each base corresponded to one replication (n = 6). At the soil surface, the bases had four round holes (diameter 2 cm) to allow water flow between the inside and outside. These holes were plugged with a rubber stopper during air collection.
Seventeen samplings were carried out over one year (September 10, 2010 to September 26, 2011), in the first month on the 1st , 4th , 8th and 14th days, followed by 13 samplings at intervals of approximately 30 days. At each sampling, PVC chambers (height 40 cm) were set on the metal bases, which contained a channel on the top (height 4 cm × width 6 cm). This channel was filled with water to seal the chamber-base system. In the upper part, the chambers were equipped with a thermometer, a fan to mix the air and a three-way stopcock attached to a rubber septum for sampling, which was done using polypropylene syringes of 20 mL. Air samples were collected 0, 15, 30 and 45 min after closing the chamber, between 9:00 and 11:00 am.
The samples were packed in polystyrene ice boxes and sent immediately to the Laboratory of Environmental Biogeochemistry, UFRGS, Porto Alegre, Rio Grande do Sul. The CH4 concentration was determined by a gas chromatograph (Shimadzu GC -2014) equipped with Porapak-Q columns at a temperature of 70 °C. The carrier gas was N2 at a flow rate of 30 mL min-1. The temperature of the methanator was 380 °C and FID detector 250 °C.
The CH4 fluxes at each sampling were calculated from the increase or reduction rate of the gas concentration in the chamber. The resulting fluxes were considered representative of the average daily flux, as proposed by Jantalia et al. (2008), and the cumulative net efflux or influx per season and year were calculated from the integration of the daily fluxes in the respective periods.
Soil and climate parameters
At each air sampling, the soil moisture and the lowering of the water-table were assessed. Three soil samples were collected from the 0-10 cm layer of each treatment plot with a soil auger, for gravimetric moisture determination (at 105 oC). To monitor the lowering of the water-table, four inspection wells consisting of PVC pipes (diameter 75 mm, length 1.20 m) were installed in each plot. The tube had four round holes (diameter ~ 1 cm), at every 10 cm along the tube length to allow a rapid water flow. Each tube was installed between two chambers and reached the mineral horizon (Cg). The height of the water-table was measured with a rigid tape.
Air temperature and pluvial precipitation were monitored daily by a portable meteorological station (Vantage Vue, Davis instruments), installed approximately 1 km away from the experimental area.
Statistical analysis
The data of CH4 fluxes, cumulative CH4 emission and consumption, gravimetric moisture, and water-table level were subjected to analysis of variance followed by Tukey's test (p<0.05). A Pearson's correlation matrix was constructed to evaluate the correlation between fluxes and data of air temperature, pluvial precipitation, gravimetric soil moisture, and water-table level. Statistical analyses were performed using the statistical software "R" (R Development Core Team, 2011).
RESULTS AND DISCUSSION
CH4 fluxes in natural Histosol
The CH4 fluxes in natural waterlogged Histosol varied considerably over the seasons, with lower emission rates in the late winter and early spring of 2010 and higher rates in late summer and early fall, reaching 2850 mg m-2 h-1 CH4 (Figure 1). This seasonal variation is associated with the temperature effect on the emissions since methanogenic organisms are sensitive to its variations (Dunfield et al., 1993; van Hulzena et al., 1999; Megonigal & Schlesinger, 2002). Under colder conditions, the methanogenic activity decreased, as occurred on July 27, 2011 (after 291 days), when frost occurred and temperature at sampling was 4.1 °C. On this date, the flux was reduced to 559 mg m-2 h-1 CH4 (Figure 1).
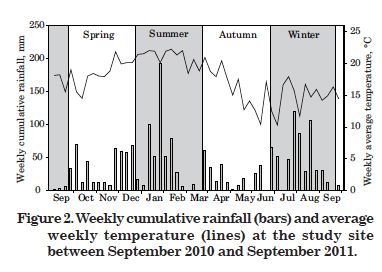
Although the correlation between CH4 emission rates and air temperature was significant and the above results are consistent with what was expected throughout the seasons, the correlation coefficient between these variables was low (r = 0.26, p<0.05) (Table 2). Two not mutually exclusive explanations are discussed. The first is that, apart from the variability of the ratio between CH4 flux and temperature, other environmental factors also regulate CH4 emissions (Le Mer & Roger, 2001; Joabsson & Christensen, 2001; Whalen, 2005; Dalal & Allen, 2008; Koh et al., 2009). The second is that there seemed to be a delay in the peak CH4 emissions, recorded in late summer and early fall (Figure 1), in relation to higher temperatures, observed mainly in late spring and in the two first thirds of summer (Figure 2). Both situations may have contributed to reduce the correlation coefficient, partially masking the temperature effect on the emissions.
The annual cumulative rainfall of 1,832 mm did not correlate with CH4 emission from the natural Histosol. The emission peak observed in late summer (Figure 1), for example, coincided with a period of low rainfall (Figure 2) and the highest emission peak recorded in early fall (Figure 1), in the first week of April, did not coincide with the period of maximum rainfall in the last week of March (Figure 2). It is likely that the flooded soil condition suppressed any influence of pluvial precipitation, so that methanogenesis was not affected by the rainfall regime.
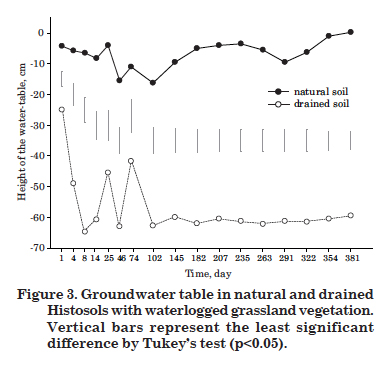
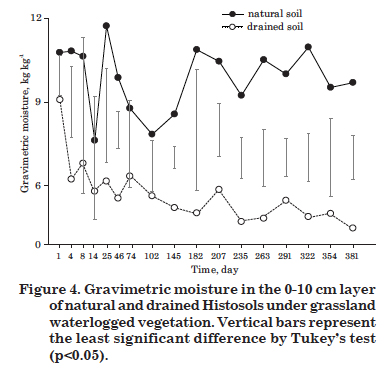
The natural Histosol was practically water-saturated for most of the study period, with an average water -table level of 7 cm below the surface (Figure 3). The correlation between water-table lowering and CH4 emissions was significant, but poor (r = -0.20, p<0.01) (Table 2), possibly due to the slight variation in the water-table level over time (-0.3 to -16.3 cm). The gravimetric soil moisture at 0-10 cm (Figure 4) had no significant effect on CH4 emission from the natural Histosol (Table 2), which agrees with the reduced effect of the water-table level.
Temperature and precipitation effects on CH4 emission from flooded Histosols reported in literature, either under natural conditions or irrigated rice cultivation, are not consistent. Some studies report positive effects of both temperature and pluvial precipitation (Jiang et al., 2009), others only of precipitation but not of temperature (Inubushi et al., 2003; Furukawa et al., 2005), while others of temperature only (von Arnold et al., 2005).
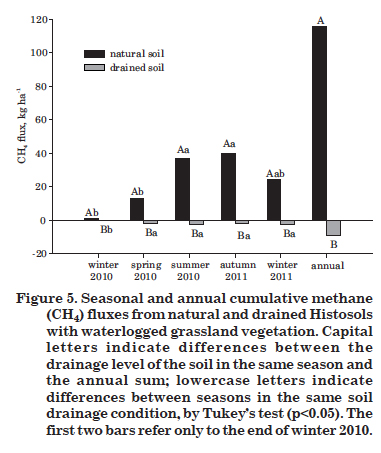
The cumulative annual emission from the natural Histosol was 116 kg ha-1 CH4 (Figure 5). The values of cumulative annual emissions from natural Histosols reported in the literature vary. In China, Jiang et al. (2009) found annual emissions of 199 kg ha-1 CH4. Dalal & Allen (2008) estimated cumulative annual emissions of 0.27 kg ha-1 in Malaysia, 64.5 kg ha-1 in Japan and 126.1 kg ha-1 in Sweden, based on research by Melling et al. (2005), Inubushi et al. (2005) and Ström et al. (2005), respectively.
In paddy rice fields, emissions reached 489 kg ha-1 yr-1 CH4 in a Histosol in Indonesia (Furukawa et al., 2005) and 586 kg ha-1 yr-1 CH4 in a Planosol (Sousa, 2013) and 623 kg ha-1 yr-1 CH4 in a Gleysol, the latter two in Rio Grande do Sul, Brazil (Zschornack, 2011). These values indicate that methane emission is much higher from paddy rice soils than from the natural Histosol in this study.
Implications of drainage on CH4 fluxes
There was a significant and consistent lowering of the water-table level in the drained compared to the natural Histosol (Figure 3). While in the natural soil the average level of the water-table was -7 cm, it was -57 cm in the drained. This reduced the gravimetric moisture in the 0-10 cm layer of the drained soil, to almost half of that observed under natural conditions (Figure 4). Consequently, the drained soil had a CH4 inflow (negative flow) over practically the entire evaluation period (Figure 1), i.e., there was methane consumption due to the oxidative activity of methanotrophic bacteria favored by the aerobic conditions induced by water removal.
One day after drainage, the water-table sank to 25 cm below the surface (Figure 3), but this was still not enough to reduce the CH4 fluxes (Figure 1). However, when the level sank to 49 cm below the surface four days after drainage (Figure 3), CH4 consumption (-39 µg m-2 h-1 CH4, the negative sign means consumption) started (Figure 1). On the eighth day, the water-table sank to 65 cm below the surface and consumption increased to -99 µg m-2 h-1 CH4. The high total porosity (0.78 to 0.95 m3 m-3) and high saturated hydraulic conductivity (130 to 720 mm h-1) (Table 1) contributed to the rapid drainage, so that within approximately one week, net consumption of CH4 was being recorded. Parallel to this, the microbial community certainly adapted very quickly to the aerobic conditions created by drainage, so that the methanotrophic supplanted the methanogenic bacteria. Possibly this was not a very difficult process, because according to Whalen (2005), methanogens and methanotrophs respond promptly to fluctuations in the water-table.
From the eighth day onwards, the water-table remained almost constant and the CH4 flux variations were not significant (-96 and -146 µg m-2 h-1 CH4) until the end of the measurements.
In general, the results of CH4 emission reduction followed by methane consumption fully agree with those of other studies that evaluated the effect of lowering the water-table level by draining in flooded soils in several parts of the world (Martikainen et al., 1995; Nykanen et al., 1995; Maljanen et al., 2004; von Arnold et al., 2005; Furukawa et al., 2005; Bridgham et al., 2006; Elder & Lal, 2008; Jauhiainen et al., 2008; Jiang et al., 2009; Huang et al., 2010; Page & Dalal, 2011). All these authors showed that small water-table decreases cause drastic reductions in methane emissions, demonstrating the strong effect of drainage on CH4 fluxes.
The occurrence of net CH4 consumption does not mean, however, that the methanogenic activity was completely suppressed, but rather that the activity of methanotrophs surpassed that of methanogens. Anaerobic bacteria such as methanogens seem to remain viable even when soil is not water-saturated (Kettunen et al., 1999), while methanotrophs can survive under anoxic conditions (Roslev & King, 1994). In swamps of Florida, increases in CH4 emissions due to reduced aerobic conditions were more related to the lower activity of methanotrophs than to the increased performance of methanogens (King et al., 1990; Sass et al., 1990).
In drained Histosols, a CH4 influx occurred and was positively correlated with water-table lowering and negatively with gravimetric moisture at 0-10 cm (Table 2). The moisture reduction was a substantial factor in methane fluxes in this soil, since the average gravimetric moisture in drained soil was 5.5 kg kg-1, which is below the mean of the natural soil (9.9 kg kg-1).
Contrary to the natural Histosol, the fluxes varied little in the drained soil throughout the seasons (Figure 1). The influence of air temperature was very small, since the correlation coefficient between it and methane flux was even lower than in the natural Histosol (Table 2). The pluvial precipitation did not affect CH4 consumption significantly in drained Histosols. This suggests that these climatic factors were not significant for the activity of methanotrophs in these soils.
Because there are other factors affecting methanogenesis and methanotrophy, e.g., variations in microbial community structure and in organic matter composition, there may be a nonlinear relationship between CH4 flux and temperature (Wilson et al., 1989; Joabsson & Christensen, 2001).
In drained Histosols, the cumulative CH4 consumption was almost the same in all seasons of the year (oscillating between -2.05 and -2.37 kg ha-1 yr-1 CH4), so that each season contributed with approximately 25 % of the total, which was 9 kg ha-1 yr-1 CH4 (Figure 5).
Effects of drainage on the ecological functions of Histosols
The annual emission of 116 kg ha-1 CH4 from natural Histosols, considering the warming potential of 25 for CH4 (IPCC, 2007), is equivalent to 2.9 or 0.79 Mg ha-1 yr -1 CO2eq. Approximately one third of this emissions occurred in the summer (37 kg ha-1) and a second third in the fall (40 kg ha-1), indicating the importance of these seasons on the total emission (Figure 5).
However, under no circumstances does this emission represent a contribution to the increase of atmospheric CH4 observed since 1850 (IPCC, 2007), for being inherent to the original condition of the ecosystem and occurring naturally long before that year.
The drainage, in turn, contributes to an annual CH4 consumption of 9 kg ha-1 (~ 61 kg ha-1 C-CO2eq) (Figure 5). In China, Histosols that emitted 199 kg ha-1 yr-1 CH4 began to consume 1.4 kg ha-1 yr-1 CH4 after having been drained and converted into cropland (Jiang et al., 2009).
It is worth highlighting the essential function of natural Histosols in the ecosystem as C reservoir. In terms of the concentration of organic C and soil density along the profile (Table 1), Histosols have a C stock of 581 Mg ha-1 to a depth of 1 m, which is certainly fundamental for the sustainability of the ecosystem.
With drainage, a significant proportion of the C stock will be mineralized and emitted into the atmosphere as CO2. Huang et al. (2010) estimated that soils of wetlands converted to cropland between 1950 and 2000 lost about 50 % C. Other authors also reported an increase in CO2 emissions from Histosols (Lohila et al., 2003; Maljanen et al., 2002, 2004; von Arnold et al., 2005; Furukawa et al., 2005; Yamulki et al., 2012), and Joosten & Couwenberg (2008) stated the emission of 30 Mg CO2 ha-1 yr-1 from drained Histosols in a forest environment.
Aside from C pools, natural Histosols are an important water filter and reservoir. The high soil porosity (Table 1) provides the capacity to store 8.6 million L water per hectare to a depth of 1m [calculated as the total porosity (Table 1) filled with water]. This water storage is equivalent to a water-table level of 860 mm and, together with the reverse cuesta position, makes Histosols fundamental for downstream watersheds, whose hydrological structuring relies on the reverse cuesta of the second plateau of Paraná.
In view of the functions Histosols fulfill in the ecosystem, they should be preserved and not drained. Strategies for mitigating CH4 emissions should be concentrated on the modification of human activities and on no account on interventions in the areas of occurrence of Histosols.
CONCLUSIONS
1. Natural Histosols under waterlogged grassland are sources of atmospheric methane since methanogenesis is favored by waterlogging conditions. The draining of these soils, on the other hand, reduces moisture and promotes oxygen diffusion, resulting in methane consumption.
2. The methane emissions from natural Histosols under natural waterlogged grassland vary over the seasons, and are higher in warmer seasons.
3. Despite acting as a methane source, natural Histosols with waterlogged grassland vegetation should be preserved because of their important functions in the ecosystem, e.g., the retention of organic carbon and water. Instead of being drained, they should rather be effectively preserved.
ACKNOWLEDGEMENTS
The authors are indebted to Embrapa Serviço de Produtos e Mercado/Escritório de Ponta Grossa (Embrapa SPM-EPGA) for granting access to the experimental area, to the Brazilian Council for Scientific and Technological Development (CNPq) for funding (Proc. 562627/2010-3), and to the research support foundation of Rio Grande do Sul (FAPERGS) whose financial support was essential for the laboratory infrastructure, particularly of the chromatographic analysis of greenhouse gases (Pronex Proc. 10/0054-7).
LITERATURE CITED
- ALONGI, D.M.; WATTAYAKORN, G.; PFITZNER, J.: TIRENDI, F.; ZAGORSKIS, I.; BRUNSKILL, G.J.; DAVIDSON, A. & CLOUGH, B.F. Organic carbon accumulation and metabolic pathways in sediments of mangrove forests in Southern Thailand. Mar. Geol., 179:85-103, 2001.
- ANDRADE, B.O. A vegetação das nascentes do rio Tibagi e sua relação com o regime hídrico dos solos. Curitiba, Universidade Federal do Paraná, 2010. 130p. (Dissertação de Mestrado)
- BRIDGHAM, S.D.; MEGONIGAL, J.P.; KELLER, J.K.; BLISS, N.B. & TRETTIN, C. The carbon balance of North American wetlands. Wetlands, 26:889-916, 2006.
- DALAL, R.C. & ALLEN, D.E. Greenhouse gas fluxes from natural ecosystems. Aust. J. Bot., 56:369-407, 2008.
- DUNFIELD, P.; KNOWLES, R.; DUMONT, R. & MOORE, T.R. Methane production and consumption in temperate and subarctic peat soils - response to temperature and pH. Soil Biol. Biochem., 25:321-326, 1993.
- ELDER, J.W. & LAL, R. Tillage effects on gaseous emissions from an intensively farmed organic soil in North Central Ohio. Soil Till. Res., 98:45-55, 2008.
- CLAESSEN, M.E.C., org. Manual de métodos de análises de solo. 2.ed. Rio de Janeiro, Embrapa-CNPS, 1997. 212p. (Documentos, 1)
- FORSTER, P.; RAMASWAMY, V.; ARTAXO, P.; BERNSTEN, T.; BETTS, R.; FAHEY, D.W.; HAYWOOD, J.; LEAN, J.; LOWE, D.C.; MYHRE, G.; NGANGA, J.; PRINN, R.; RAGA, G.; SCHUTZ, M. & van DORLAND, R. Changes in atmospheric constituents and in radiative forcing. In: SOLOMON, S.; QIN, D.; MANNING, M.; CHEN, Z.; MARQUIS, M.; AVERYT, K.B.; TIGNOR, M. & MILLER, H.L., eds. Climate change 2007: The physical science basis: Contribution of Working Group 1 to the Fourth Assessment Report of the Intergovernmental Panel on Climate Changes. Cambridge, University Press, 2007. p.129-234.
- FURUKAWA, Y.; INUBUSHI, K.; ALI, M.; ITANG, A.M. & TSURUTA, H. Effect of changing groundwater levels caused by land-use changes on greenhouse gas fluxes from tropical peat lands. Nutr. Cycl. Agroecosyst., 71:81-91, 2005.
- GOMES, J.; BAYER, C.; COSTA, F.S.; PICCOLO, M.C.; VIEIRA, F.C.B. & SIX, J. Soil nitrous oxide emission as affected by long term tillage, crop rotations and fertilization in a subtropical environment. Soil Till. Res., 106:36-44, 2009.
- HAYES, M.H.B. & CLAPP, C.E. Humic substances: considerations of compositions, aspects of structure and environmental influences. Soil Sci., 166:723-727, 2001.
- HUANG, Y.; SUN, W.J.; ZHANG, W.; YU, Y.Q.; SU, Y.H. & SONG, C.C. Marshland conversion to cropland in northeast China from 1950 to 2000 reduced the greenhouse effect. Global Change Biol., 16:680-695, 2010.
- INUBUSHI, K.; FURUKAWA, Y.; HADI, A.; PURNOMO, E. & TSURUTA, H. Seasonal changes of CO2, CH4 and N2O fluxes in relation to land-use changes in tropical peatlands located in coastal area of South Kalimantan. Chemosphere, 52:603-608, 2003.
- inubushi, k.; otake, s.; furukawa, y.; shibasaki, n.; ali, m.; itang, a.m. & tsuruta, h. Factors influencing methane emission from peat soil: Comparison of tropical and temperate wetlands. Nutr. cycl. Agroecosyst., 71:93-99, 2005.
- INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE - IPCC. Climate Change 2007: The physical science basis: Summary for policymakers. Intergovernmental Panel on Climate Change. Geneva, 2007.
- JANTALIA, C.P.; SANTOS, H.P.; URQUIAGA, S.; BODDEY, R.M. & ALVES, B.J.R. Fluxes of nitrous oxide from soil under different crop rotations and tillage systems in the South of Brazil. Nutr. Cycl. Agroecosyst., 82:161-173, 2008.
- JAUHIAINEN, J.; LIMIN, S.; SILVENNOINEN, H. & VASANDER, H. Carbon dioxide and methane fluxes in drained tropical peat before and after hydrological restoration. Ecology, 89:3503-3514, 2008.
- JIANG, C.; WANG, Y.; HAO, Q. & SONG, C. Effect of land-use change on CH4 and N2O emissions from freshwater marsh in Northeast China. Atmos. Environ., 43:3305-3309, 2009.
- JOABSSON, A. & CHRISTENSEN, T.R. Methane emissions from wetlands and their relationship with vascular plants: An Arctic example. Global Change Biol., 7:919-932, 2001.
- JOOSTEN, H. & COUWENBERG, J. Peatlands and carbon. In: PARISH, F.; SIRIN, A.; CHARMAN, D.; JOOSTEN, H.; MINAYEVA, T.; SILVIUS, M. & STRINGER, L., eds. Assessment on peatlands, biodiversity and climate change. Kuala Lumpur, Global Environment Centre, 2008. p.99-117.
- KETTUNEN, A.; KAITALA, V.; LEHTINEN, A.; LOHILA, A.; ALM, J.; SILVOLA, J. & MARTIKAINEN, P.J. Methane production and oxidation potentials in relation to water table fluctuations in two boreal mires. Soil Biol. Biochem., 31:1741-1749, 1999.
- KING, G.M.; ROSLEV, P. & SKOVGAARD, H. Distribution and rate of methane oxidation in sediments of the Florida Everglades. Appl. Environ. Microbiol., 56:2902-2911, 1990.
- KOH, H.S.; OCHS, C.A. & YU, K.W. Hydrologic gradient and vegetation controls on CH4 and CO2 fluxes in a spring-fed forested wetland. Hydrobiologia, 630:271-286, 2009.
- KÖGEL-KNABNER, I.; AMELUNG, W.; CAO, Z.H.; FIEDLER, S.; FRENZEL, P.; JAHN, R.; KALBITZ, K.; KOLBL, A. & SCHLOTER, M. Biogeochemistry of paddy soils. Geoderma, 157:1-14, 2010.
- KOZERA, C. Florística e fitossociologia de uma formação pioneira com influência fluvial e de uma estepe gramíneo-lenhosa em diferentes unidades geopedológicas, município de Balsa Nova, Paraná - Brasil. Curitiba, Universidade Federal do Paraná, 2008. 267p. (Tese de Doutorado)
- LE MER, J. & ROGER, P. Production, oxidation, emission and consumption of methane by soils: A review. Eur. J. Soil Biol., 37:25-50, 2001.
- LOHILA, A.; AURELA, M.; REGINA, K. & LAURILA, T. Soil and total ecosystem respiration in agricultural fields: Effects of soil and crop type. Plant Soil, 251:303-317, 2003.
- MALJANEN, M.; MARTIKAINEN, P.J.; AALTONEN, H. & SILVOLA, J. Short-term variation in fluxes of carbon dioxide, nitrous oxide and methane in cultivated and forested organic boreal soils. Soil Biol. Biochem., 34:577-584, 2002.
- MALJANEN, M.; KOMULAINEN, V.M.; HYTONEN, J.; MARTIKAINEN, P.J. & LAINE, J. Carbon dioxide, nitrous oxide and methane dynamics in boreal organic agricultural soils with different soil characteristics. Soil Biol. Biochem., 36:1801-1808, 2004.
- MARTIKAINEN, P.J.; NYKANEN, H.; ALM, J. & SILVOLA, J. Changes in fluxes of carbon dioxide, methane and nitrous oxide due to forest drainage of mire sites of different trophy. Plant Soil, 168-169:571-577, 1995.
- MEGONIGAL, J.P. & SCHLESINGER, W.H. Methane-limited methanotrophy in tidal freshwater swamps. Global Biogeochem. Cycles, 16:1088, 2002.
- MELLING, L.; HATANO, R. & GOH, K.J. Methane fluxes from three ecosystems in tropical peatland of Sarawak, Malaysia. Soil Biol. Biochem., 37:1445-1453, 2005.
- MOORE, T.R. & DALVA, M. The influence of temperature and water-table position on carbon-dioxide and methane emission from laboratory columns of peat land soils. J. Soil Sci., 44:641-664, 1993.
- NYKANEN, H.; ALM, J.; LANG, K.; SILVOLA, J. & MARTIKAINEN, P.J. Emissions of CH4, N2O and CO2 from a virgin fen and a fen drained for grassland in Finland. J. Biogeogr., 22:351-357, 1995.
- PAGE, K.L. & DALAL, R.C. Contribution of natural and drained wetland systems to carbon stocks, CO2, N2O and CH4 fluxes: An Australian perspective. Soil Res., 49:377-388, 2011.
- R DEVELOPMENT CORE TEAM. R: A language and environment for statistical computing. Viena: R Foundation for Statistical Computing, 2011. Available at: <http://www.R-project.org/>. Accessed: May 2012.
- RAICH, J.W. & POTTER, C.S. Global patterns of carbon dioxide emissions from soils. Global Biogeochem. Cycles, 9:23-36, 1995.
- ROULET, N.T. & MOORE, T.R. The effect of forestry drainage practices on the emission of methane from northern peatlands. Can. J. For. Res., 25:491-499, 1995.
- ROSLEV, P. & KING, G.M. Survival and recovery of methanotrophic bacteria starved under oxic and anoxic conditions. Appl. Environ. Microbiol., 60:2602-2608, 1994.
- SANTOS, H.G.; JACOMINE, P.K.T.; ANJOS, L.H.C.; OLIVEIRA, V.A.; OLIVEIRA, J.B.; COELHO, R.M.; LUMBRERAS, J.F. & CUNHA, T.J.F., eds. Sistema brasileiro de classificação de solos. 2.ed. Rio de Janeiro, Embrapa Solos, 2006. 306p.
- SASS, R.L.; FISHER, F.M.; HARCOMBE, P.A. & TURNER, F.T. Methane production and emission in a Texas rice-field. Global Biogeochem. Cycles, 4:47-68, 1990.
- SCHEER, M.B.; CURCIO, G.R. & RODERJAN, C.V. Funcionalidades ambientais de solos altomontanos na Serra da Igreja, Paraná. R. Bras. Ci. Solo, 35:1013-1026, 2011.
- SOIL SURVEY STAFF. Keys to soil taxonomy. 5.ed. Virginia, Pocahontas Press, 1992. 541p.
- SOUSA, E.L. Emissão de óxido nitroso e metano em área de cultivo com arroz irrigado com diferentes manejos da palha pós-colheita. Santa Maria, Universidade Federal de Santa Maria, 2013. 139p. (Tese de Doutorado)
- STRÖM, L.; MASTEPANOV, M. & CHRISTENSEN, T.R. Species-specific effects of vascular plants on carbon turnover and methane emissions from wetlands. Biogeochemistry, 75:65-82, 2005.
- VALLADARES, G.S. Caraterização de Organossolos, auxílio à sua classificação. Seropédica, Universidade Federal Rural do Rio de Janeiro, 2003. 129p. (Tese de Doutorado)
- van HULZENA, J.B.; SEGERSA, R.; van BODEGOMA, P.M. & LEFFELAAR, P.A. Temperature effects on soil methane production: An explanation for observed variability. Soil Biol. Biochem., 31:1919-1929, 1999.
- von ARNOLD, K.; NILSSON, M.; HANELL, B.; WESLIEN, P. & KLEMEDTSSON, L. Fluxes of CO2, CH4 and N2O from drained organic soils in deciduous forests. Soil Biol. Biochem., 37:1059-1071, 2005.
- WHALEN, S.C. Biogeochemistry of methane exchange between natural wetlands and the atmosphere. Environ. Eng. Sci., 22:73-94, 2005.
- WHITING, G.J. & CHANTON, J.P. Greenhouse carbon balance of wetlands: Methane emission versus carbon sequestration. Tellus. Series B, Chem. Phys. Meteorol., 53:521-528, 2001.
- WILSON, J.O.; CRILL, P.M.; BARTLETT, K.B.; SEBACHER, D.I.; HARRISS, R.C. & SASS, R.L. Seasonal variation of methane emissions from a temperate swamp. Biogeochemistry, 8:55-71, 1989.
- WUEBBLES, D.J.; HAYHOE, K.A.S. & KOTAMARTHI, R. Methane in the global environment. In: KHALIL, M.A.K., ed. Atmospheric Methane: Its role in the global environment. Berlin, Springer-Verlag, 2000. p.304-341.
- YAMULKI, S.; ANDERSON, R.; PEACE, A.; MORISON, J.I.L. Soil CO2, CH4, and N2O fluxes from an afforested lowland raised peatbog in Scotland: Implications for drainage and restoration. Biogeosci. Disc., 9:7313-7351, 2012.
- ZSCHORNACK, T. Emissões de metano e de óxido nitroso em sistemas de produção de arroz irrigado no sul do Brasil e potencial de mitigação por práticas de manejo. Porto Alegre, Universidade Federal do Rio Grande do Sul, 2011. 101p. (Tese de Doutorado)
Publication Dates
-
Publication in this collection
18 June 2014 -
Date of issue
Apr 2014
History
-
Received
01 Nov 2012 -
Accepted
05 Nov 2013